Optimized Bioconversion of Xylose Derived from Pre-Treated Crop Residues into Xylitol by Using Candida boidinii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Lignocellulosic Materials
2.3. Compositional Analysis
2.4. Preparation of Xylose-Rich Hydrolysates from Wheat Bran and Rice Straw
2.5. Treatments and Supplementations of the Xylose-Rich Hydrolysates before Xylitol Fermentation
2.6. Inoculum Preparation
2.7. Xylitol Fermentation
2.8. Analytical Methods
2.8.1. Concentration of Sugars, Alcohols, Organic Acids, Phenols, and Total Protein
2.8.2. Cell Concentration
2.8.3. Thin-Layer Chromatography
2.8.4. Determination of Oxygen Transfer Rate
2.9. Calculation of Xylose Yield, Xylitol Yield, Xylitol Volumetric Productivity, Specific Xylitol Yield and Combined Severity Factor
2.10. Statistical Evaluations and Optimisation
3. Results
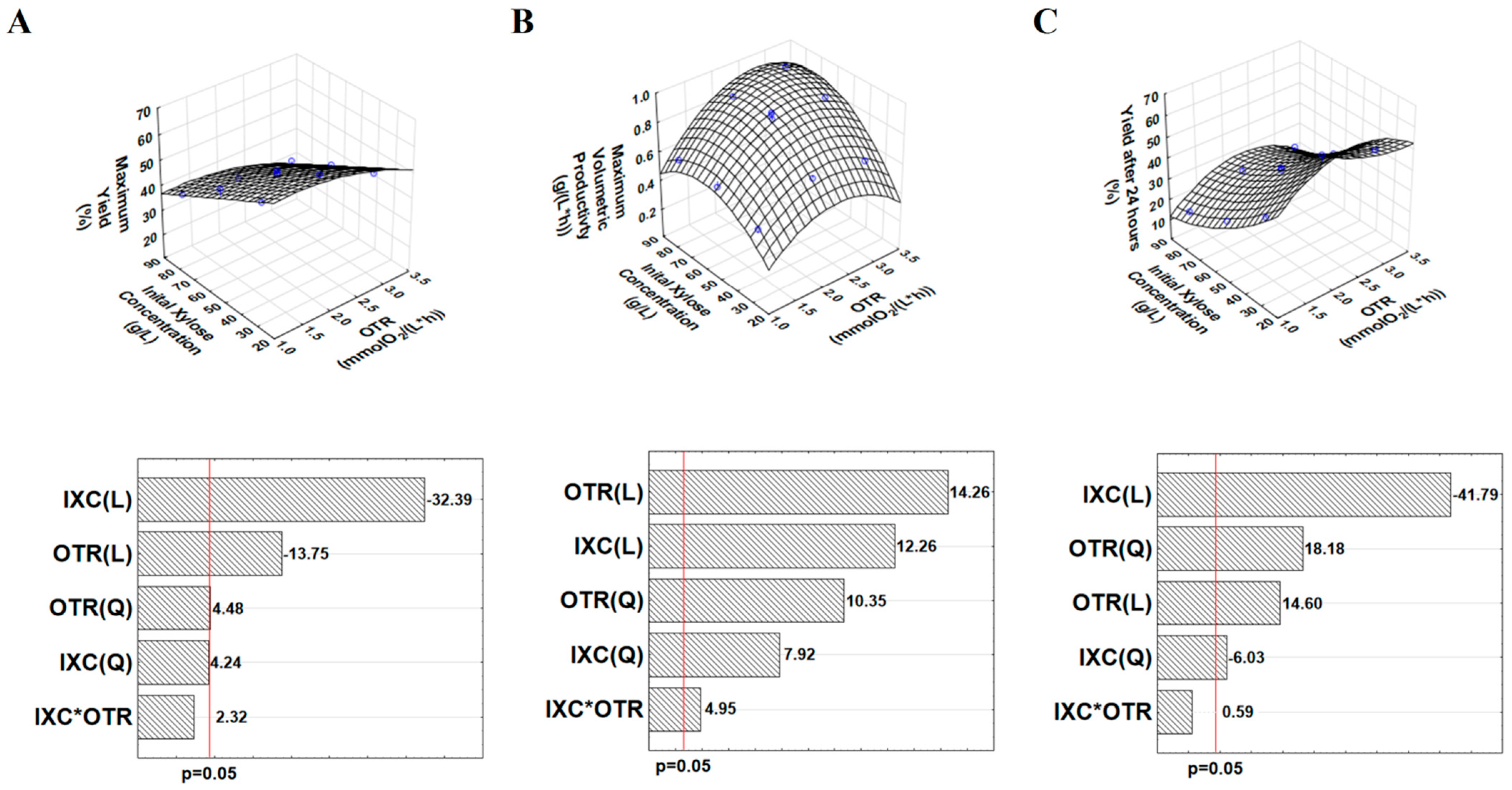
3.1. Investigating the Effects of OTR and Initial Xylose Concentration on Xylitol Production by Candida boidinii
3.2. Xylose-Rich Hydrolysates from Rice Straw and Wheat Bran
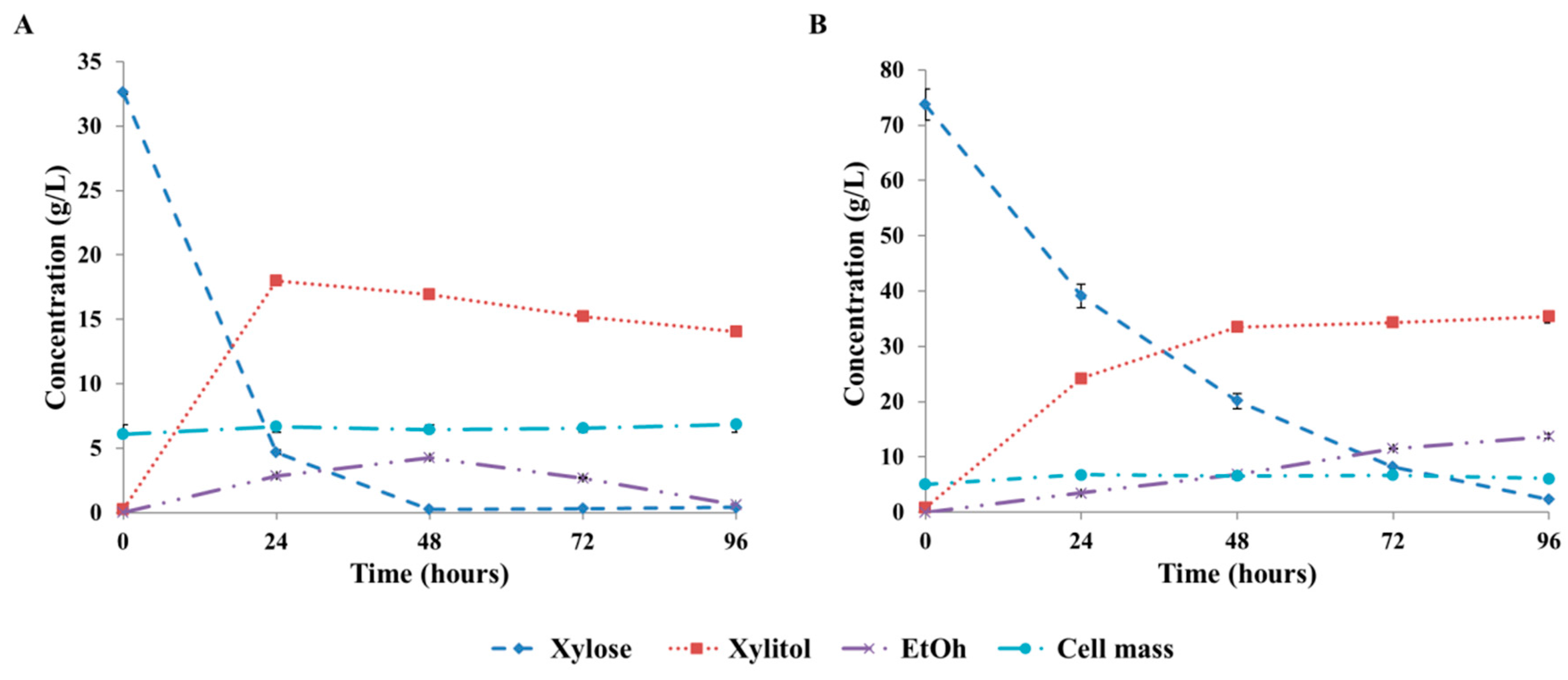
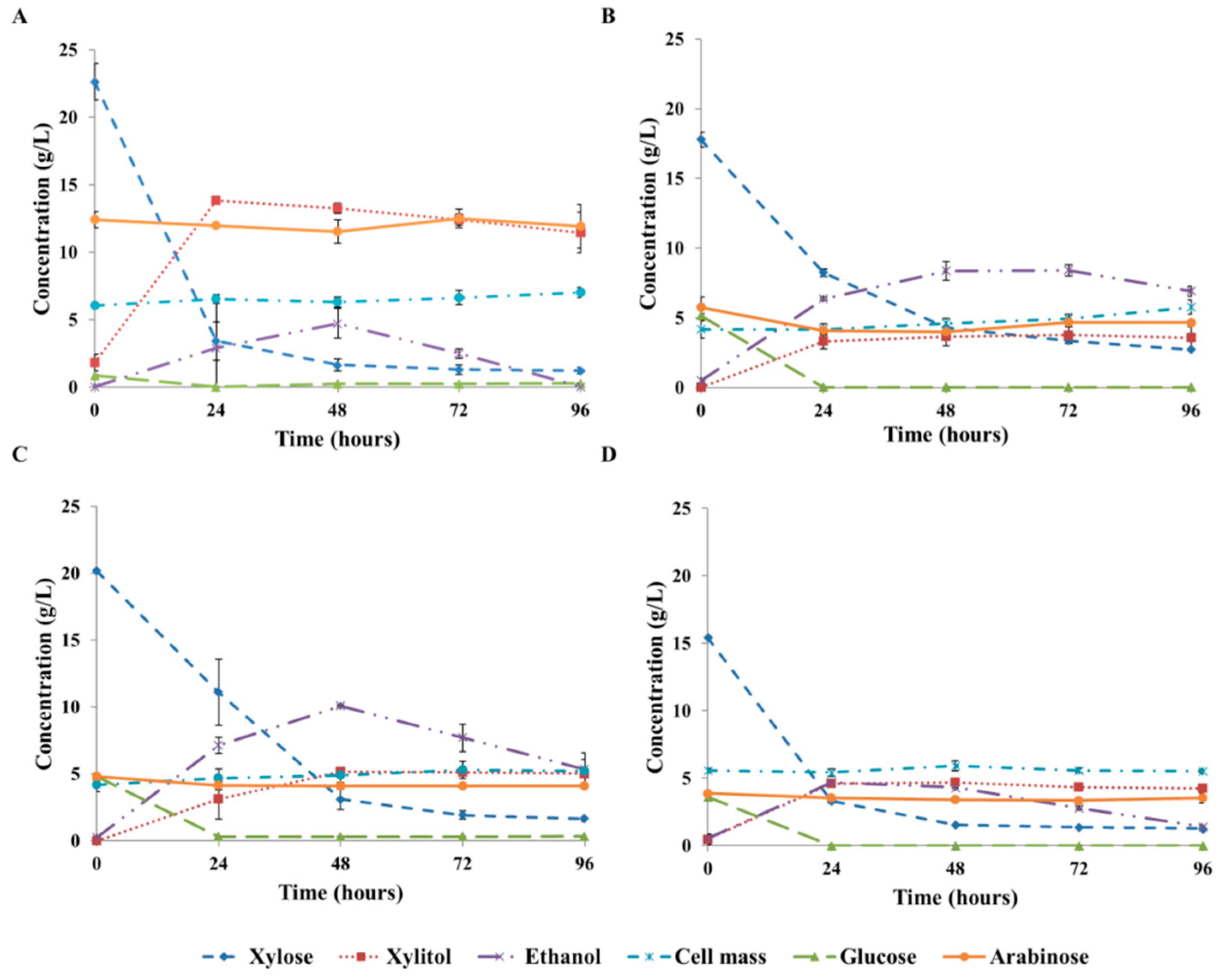
3.3. Xylitol Fermentation on Xylose-Rich Hydrolysates of Rice Straw and Wheat Bran
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ubando, A.T.; Felix, C.B.; Chen, W.H. Biorefineries in circular bioeconomy: A comprehensive review. Bioresour. Technol. 2020, 299, 122585. [Google Scholar] [CrossRef] [PubMed]
- Fehér, C.; Barta, Z.; Réczey, K. Process considerations of a biorefinery producing value-added products from corn fibre. Period. Polytech. Chem. Eng. 2012, 56, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Dahmen, N.; Lewandowski, I.; Zibek, S.; Weidtmann, A. Integrated lignocellulosic value chains in a growing bioeconomy: Status quo and perspectives. GCB Bioenergy 2019, 11, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Piotrowski, S.; Essel, R.; Carus, M.; Dammer, L.; Engel, L. Nachhaltig nutzbare Potenziale für Biokraftstoffe in Nutzungskonkurrenz zur Lebens-und Futtermittelproduktion, Bioenergie sowie zur stofflichen Nutzung in Deutschland, Europa und der Welt. Eur. Welt Nova-Institut 2015, 22501112, 1–252. [Google Scholar]
- Isikgor, F.H.; Becer, C.R. Lignocellulosic biomass: A sustainable platform for the production of bio-based chemicals and polymers. Polym. Chem. 2015, 6, 4497–4559. [Google Scholar] [CrossRef] [Green Version]
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Alexandri, M.; López-Gómez, J.P.; Olszewska-Widdrat, A.; Venus, J. Valorising agro-industrial wastes within the circular bioeconomy concept: The case of defatted rice bran with emphasis on bioconversion strategies. Fermentation 2020, 6, 42. [Google Scholar] [CrossRef]
- FAO. Cereal Supply and Demand Brief. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 27 November 2020).
- Bandumula, N. Rice Production in Asia: Key to Global Food Security. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1323–1328. [Google Scholar] [CrossRef]
- Sharma, I.; Tyagi, B.S.; Singh, G.; Venkatesh, K.; Gupta, O.P. Enhancing wheat production—A global perspective. Indian J. Agric. Sci. 2015, 85, 3–13. [Google Scholar]
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef]
- Prückler, M.; Siebenhandl-Ehn, S.; Apprich, S.; Höltinger, S.; Haas, C.; Schmid, E.; Kneifel, W. Wheat bran-based biorefinery 1: Composition of wheat bran and strategies of functionalization. LWT Food Sci. Technol. 2014, 56, 211–221. [Google Scholar] [CrossRef]
- Kim, S.; Dale, B.E. Global potential bioethanol production from wasted crops and crop residues. Biomass Bioenergy 2004, 26, 361–375. [Google Scholar] [CrossRef]
- Balan, V.; Da Costa Sousa, L.; Chundawat, S.P.S.; Vismeh, R.; Jones, A.D.; Dale, B.E. Mushroom spent straw: A potential substrate for an ethanol-based biorefinery. J. Ind. Microbiol. Biotechnol. 2008, 35, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Koegelenberg, D.; Chimphango, A.F.A. Effects of wheat-bran arabinoxylan as partial flour replacer on bread properties. Food Chem. 2017, 221, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Gautam, A.; Dutt, D. Biotechnological Transformation of Lignocellulosic Biomass in to Industrial Products: An Overview. Adv. Biosci. Biotechnol. 2016, 7, 149–168. [Google Scholar] [CrossRef] [Green Version]
- Bedő, S.; Antal, B.; Rozbach, M.; Fehér, A.; Fehér, C. Optimised fractionation of wheat bran for arabinose biopurification and xylitol fermentation by Ogataea zsoltii within a biorefinery process. Ind. Crops Prod. 2019, 139, 111504. [Google Scholar] [CrossRef]
- Liaw, W.-C.; Chen, C.-S.; Chang, W.-S.; Chen, K.-P. Xylitol Production from Rice Straw Hemicellulose Hydrolyzate by Polyacrylic Hydrogel Thin Films with Immobilized Candida subtropicalis WF79. J. Biosci. Bioeng. 2008, 105, 97–105. [Google Scholar] [CrossRef]
- Ylikahri, R. Metabolic and Nutritional Aspects of Xylitol. In Advances in Food Research; Chichester, C.O., Ed.; Academic Press: Cambridge, MA, USA, 1979; Volume 25, pp. 159–180. [Google Scholar]
- Mussatto, S.I. Application of Xylitol in Food Formulations and Benefits for Health. In D-Xylitol; Springer: Berlin/Heidelberg, Germany, 2012; pp. 309–323. [Google Scholar]
- Ravichandran, K.; Jain, J.; Sadhu, B.J.; Gunasekaran, S.; Poojitha, M.C.; Majumder, P.; Isaac, N. The efficacy of xylitol based oral hygiene products on salivary parameters—An invivo study. Int. J. Res. Pharm. Sci. 2020, 11, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Delgado Arcaño, Y.; Valmaña García, O.D.; Mandelli, D.; Carvalho, W.A.; Magalhães Pontes, L.A. Xylitol: A review on the progress and challenges of its production by chemical route. Catal. Today 2018, 344, 2–14. [Google Scholar] [CrossRef]
- Mikkola, J.P.; Sjöholm, R.; Salmi, T.; Mäki-Arvela, P. Xylose hydrogenation: Kinetic and NMR studies of the reaction mechanisms. Catal. Today 1999, 48, 73–81. [Google Scholar] [CrossRef]
- Yadav, M.; Mishra, D.K.; Hwang, J.S. Catalytic hydrogenation of xylose to xylitol using ruthenium catalyst on NiO modified TiO2 support. Appl. Catal. A Gen. 2012, 425–426, 110–116. [Google Scholar] [CrossRef]
- Tran, L.H.; Yogo, M.; Ojima, H.; Idota, O.; Kawai, K.; Suzuki, T.; Takamizawa, K. The production of xylitol by enzymatic hydrolysis of agricultural wastes. Biotechnol. Bioprocess Eng. 2004, 9, 223–228. [Google Scholar] [CrossRef]
- Espinoza-Acosta, J.L. Biotechnological Production of Xylitol from Agricultural Waste. Biotecnia 2020, XXII, 126–134. [Google Scholar]
- Winkelhausen, E.; Pittman, P.; Kuzmanova, S.; Jeffries, T.W. Xylitol formation by Candida boidinii in oxygen limited chemostat culture. Biotechnol. Lett. 1996, 18, 753–758. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Roberto, I.C. Xylitol production from high xylose concentration: Evaluation of the fermentation in bioreactor under different stirring rates. J. Appl. Microbiol. 2003, 95, 331–337. [Google Scholar] [CrossRef]
- Tamburini, E.; Costa, S.; Marchetti, M.G.; Pedrini, P. Optimized production of xylitol from xylose using a hyper-acidophilic Candida tropicalis. Biomolecules 2015, 5, 1979–1989. [Google Scholar] [CrossRef] [Green Version]
- Nolleau, V.; Preziosi-Belloy, L.; Delgenes, J.P.; Navarro, J.M. Xylitol production from xylose by two yeast strains: Sugar tolerance. Curr. Microbiol. 1993, 27, 191–197. [Google Scholar] [CrossRef]
- Felipe Hernández-Pérez, A.; de Arruda, P.V.; Sene, L.; da Silva, S.S.; Kumar Chandel, A.; de Almeida Felipe, M. das G. Xylitol bioproduction: State-of-the-art, industrial paradigm shift, and opportunities for integrated biorefineries. Crit. Rev. Biotechnol. 2019, 39, 924–943. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Sugars, Byproducts, and Degradation Products in Liquid Fraction Process. Natl. Renew. Energy Lab. 2008, NREL/TP-510-42623, 1–11. [Google Scholar]
- Hames, B.; Scarlata, C.; Sluiter, A. Determination of protein content in biomass. Natl. Renew. Energy Lab. 2008, NREL/TP-510-42625, 1–5. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of ash in biomass. Natl. Renew. Energy Lab. 2008, NREL/TP-510-42622, 1–5. [Google Scholar]
- Jampatesh, S.; Sawisit, A.; Wong, N.; Jantama, S.S.; Jantama, K. Evaluation of inhibitory effect and feasible utilization of dilute acid-pretreated rice straws on succinate production by metabolically engineered Escherichia coli AS1600a. Bioresour. Technol. 2019, 273, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Palmarola-Adrados, B.; Chotěborská, P.; Galbe, M.; Zacchi, G. Ethanol production from non-starch carbohydrates of wheat bran. Bioresour. Technol. 2005, 96, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Cavka, A.; Jönsson, L.J.; Hong, F. Comparison of methods for detoxification of spruce hydrolysate for bacterial cellulose production. Microb. Cell Fact. 2013, 12, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Fehér, C.; Gazsó, Z.; Gál, B.; Kontra, A.; Barta, Z.; Réczey, K. Integrated Process of Arabinose Biopurification and Xylitol Fermentation Based on the Diverse Action of Candida boidinii. Chem. Biochem. Eng. Q. 2015, 29, 587–597. [Google Scholar] [CrossRef]
- Ghio, S.; Ontañon, O.; Piccinni, F.E.; Marrero Díaz de Villegas, R.; Talia, P.; Grasso, D.H.; Campos, E. Paenibacillus sp. A59 GH10 and GH11 Extracellular Endoxylanases: Application in Biomass Bioconversion. Bioenergy Res. 2018, 11, 174–190. [Google Scholar] [CrossRef] [Green Version]
- WISE, W.S. The measurement of the aeration of biological culture media. J. Gen. Microbiol. 1950, 4, 167–177. [Google Scholar]
- Nishikawa, M.; Nakamura, M.; Yagi, H.; Hashimoto, K.; Nishikawa, M.; Yagi, H. Gas absorption in aerated mixing vessels. J. Chem. Eng. Jpn. 1981, 14, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Wyman, C.E.; Yang, B. Combined Severity Factor for Predicting Sugar Recovery in Acid-Catalyzed Pretreatment Followed by Enzymatic Hydrolysis. In Hydrothermal Processing in Biorefineries: Production of Bioethanol and High Added-Value Compounds of Second and Third Generation Biomass; Ruiz, H.A., Hedegaard Thomsen, M., Trajano, H.L., Eds.; Springer: Cham, Switzerland, 2017; pp. 161–180. ISBN 978-3-319-56457-9. [Google Scholar]
- Fehér, C.; Gazsó, Z.; Tatijarern, P.; Molnár, M.; Barta, Z.; Réczey, K. Investigation of selective arabinose release from corn fibre by acid hydrolysis under mild conditions. J. Chem. Technol. Biotechnol. 2015, 90, 896–906. [Google Scholar] [CrossRef]
- Vandeska, E.; Amartey, S.; Kuzmanova, S.; Jeffries, T. Effects of environmental conditions on production of xylitol by Candida boidinii. World J. Microbiol. Biotechnol. 1995, 11, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Vongsuvanlert, V.; Tani, Y. Xylitol production by a methanol yeast, Candida boidinii (Kloeckera sp.) No. 2201. J. Ferment. Bioeng. 1989, 67, 35–39. [Google Scholar] [CrossRef]
- Kim, J.H.; Han, K.C.; Koh, Y.H.; Ryu, Y.W.; Seo, J.H. Optimization of fed-batch fermentation for xylitol production by Candida tropicalis. J. Ind. Microbiol. Biotechnol. 2002, 29, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Aranda-Barradas, J.S.; Garibay-Orijel, C.; Badillo-Corona, J.A.; Salgado-Manjarrez, E. A stoichiometric analysis of biological xylitol production. Biochem. Eng. J. 2010, 50, 1–9. [Google Scholar] [CrossRef]
- Granström, T.B.; Izumori, K.; Leisola, M. A rare sugar xylitol. Part I: The biochemistry and biosynthesis of xylitol. Appl. Microbiol. Biotechnol. 2007, 74, 277–281. [Google Scholar] [CrossRef]
- Winkelhausen, E.; Amartey, S.A.; Kuzmanova, S. Xylitol production from D-xylose at different oxygen transfer coefficients in a batch bioreactor. Eng. Life Sci. 2004, 4, 150–154. [Google Scholar] [CrossRef]
- Santana, N.B.; Teixeira Dias, J.C.; Rezende, R.P.; Franco, M.; Silva Oliveira, L.K.; Souza, L.O. Production of xylitol and bio-detoxification of cocoa pod husk hemicellulose hydrolysate by Candida boidinii XM02G. PLoS ONE 2018, 13, e0195206. [Google Scholar] [CrossRef] [Green Version]
- López-Linares, J.C.; Ruiz, E.; Romero, I.; Castro, E.; Manzanares, P. Xylitol production from exhausted olive pomace by Candida boidinii. Appl. Sci. 2020, 10, 6966. [Google Scholar]
- Mayerhoff, Z.D.V.L.; Roberto, I.C.; Silva, S.S. Xylitol production from rice straw hemicellulose hydrolysate using different yeast strains. Biotechnol. Lett. 1997, 19, 407–409. [Google Scholar] [CrossRef]
- Dasgupta, D.; Bandhu, S.; Adhikari, D.K.; Ghosh, D. Challenges and prospects of xylitol production with whole cell bio-catalysis: A review. Microbiol. Res. 2017, 197, 9–21. [Google Scholar] [CrossRef]
- Silva, C.J.S.M.; Roberto, I.C. Optimization of xylitol production by Candida guilliermondii FTI 20037 using response surface methodology. Process Biochem. 2001, 36, 1119–1124. [Google Scholar] [CrossRef]
- Zeid, A.A.A.; El-Fouly, M.Z.; El-Zawahry, Y.A.; El-Mongy, T.M.; El-Aziz, A.B.A. Bioconversion of rice straw xylose to xylitol by a local strain of Candida Tropicalis. J. Appl. Sci. Res. 2008, 4, 975–986. [Google Scholar]
- Schneider, H. Selective removal of acetic acid from hardwood-spent sulfite liquor using a mutant yeast. Enzyme Microb. Technol. 1996, 19, 94–98. [Google Scholar] [CrossRef]
- Fehér, A.; Fehér, C.; Rozbach, M.; Rácz, G.; Fekete, M.; Hegedűs, L.; Barta, Z. Treatments of Lignocellulosic Hydrolysates and Continuous-Flow Hydrogenation of Xylose to Xylitol. Chem. Eng. Technol. 2018, 41, 496–503. [Google Scholar] [CrossRef]
- Hua, Y.; Wang, J.; Zhu, Y.; Zhang, B.; Kong, X.; Li, W.; Wang, D.; Hong, J. Release of glucose repression on xylose utilization in Kluyveromyces marxianus to enhance glucose-xylose co-utilization and xylitol production from corncob hydrolysate. Microb. Cell Fact. 2019, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Kogje, A.; Ghosalkar, A. Xylitol production by Saccharomyces cerevisiae overexpressing different xylose reductases using non-detoxified hemicellulosic hydrolysate of corncob. 3 Biotech 2016, 6, 127. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Type of Acid | Raw Material | Treatments | Xylose-Rich Hydrolysate | |
|---|---|---|---|---|
| First Step | Second Step | |||
| H3PO4 | ground rice straw | soaking (10% DM, 2N acid, 4 h, 25 °C) | in autoclave (soaked slurry, 121 °C, 30 min) | GRS/P |
| fine ground rice straw | soaking (10% DM, 2N acid, 4 h, 25 °C) | in autoclave (soaked slurry, 121 °C, 30 min) | FGRS/P | |
| H2SO4 | ground rice straw | in autoclave (10% DM, 1.5% acid, 121 °C, 30 min) | - | GRS/S |
| fine ground rice straw | in autoclave (10% DM, 1.5% acid, 121 °C, 30 min) | - | FGRS/S | |
| wheat bran | in water bath (10% DM, 90 °C, 1.16% acid, 50 min) | in autoclave (first solid residue, 7.5% DM, 1% acid sol., 121 °C, 30 min) | WB1/S | |
| wheat bran | in water bath (10% DM, 90 °C, 1.61% acid, 47 min) | in autoclave (first solid residue, 7.5% DM, 1% acid sol., 121 °C, 30 min) | WB2/S | |
| OTR | IXC (Nominal Values) | Max. Xylitol Yield | Time of Max. Xylitol Yield | Max. Xylitol Productivity | 24-h Xylitol Yield | Xylose at Max. Xylitol Yield | Spec. Xylitol Yield at Max. Xylitol Yield | Max. Ethanol |
|---|---|---|---|---|---|---|---|---|
| mmol O2/(L × h) | g/L | % | h | g/(L × h) | % | g/L | g/g | g/L |
| 3.1 | 30 | 49 | 24 | 0.61 | 49 * | 3 | 0.55 | 4 |
| 2.1 | 30 | 58 | 24 | 0.66 | 58 * | 4.5 | 0.71 | 2.7 |
| 1.1 | 30 | 58 | 48 | 0.49 | 41 | 1.4 | 0.61 | 2 |
| 3.1 | 55 | 42 | 72 | 0.88 | 35 | 0.2 | 0.43 | 2 |
| 2.1 | 55 | 50 | 72 | 0.88 | 39 | 0.5 | 0.51 | 6.1 |
| 2.1 | 55 | 48 | 72 | 0.91 | 40 | 0.4 | 0.49 | 5.9 |
| 2.1 | 55 | 49 | 72 | 0.92 | 40 | 0.6 | 0.5 | 6 |
| 1.1 | 55 | 52 | 72 | 0.59 | 26 | 5.3 | 0.58 | 5.7 |
| 3.1 | 80 | 34 | 96 | 0.93 | 27 | 9.7 | 0.39 | 3.1 |
| 2.1 | 80 | 36 | 96 | 0.87 | 26 | 19.8 | 0.49 | 5.9 |
| 1.1 | 80 | 39 | 96 | 0.6 | 18 | 30.3 | 0.64 | 2.8 |
| Conditions | 30 g/L IXC, 2.1 mmol O2/(L × h) OTR, Semi-Defined Medium | 71 g/L IXC, 2.1 mmol O2/(L × h) OTR, Semi-Defined Medium | ||||
|---|---|---|---|---|---|---|
| Parameters | Max.. Xylitol Yield | Max. Xylitol Productivity | 24-h Xylitol Yield | Max. Xylitol Yield | Max. Xylitol Productivity | 24-h Xylitol Yield |
| % | g/(L × h) | % | % | g/(L × h) | % | |
| Predicted | 57 | 0.68 | 55 | 42 | 0.92 | 32 |
| 95% prediction interval | 54–61 | 0.57–0.79 | 51–59 | 39–46 | 0.81–1.02 | 29–36 |
| Experimentally obtained | 55 (0.18) | 0.75 (0.00) | 55 (0.18) | 46 (1.67) | 1.01 (0.01) | 32 (1.43) |
| Composition (g/L) | ||||||
|---|---|---|---|---|---|---|
| WB1/S | WB2/S | GRS/S | FGRS/S | GRS/P | FGRS/P | |
| Glucose | 0.8 (0.1) | 1.0 (0.0) | 5.5 (0.1) | 5.3 (0.3) | n.m. | n.m. |
| Xylose | 22.6 (1.3) | 21.1 (0.4) | 20.6 (0.7) | 19.9 (0.5) | 11.1 (0.5) | 11.0 (0.2) |
| Arabinose | 12.4 (0.6) | 9.1 (1.3) | 3.9 (0.2) | 3.7 (0.2) | 3.3 (0.1) | 3.2 (0.1) |
| Protein | 7.3 (0.3) | 7.0 (0.4) | 0.3 (0.0) | n.m. | 0.7 (0.0) | n.m. |
| Acetic acid | 1.0 (0.1) | 0.8 (0.0) | 2.4 (0.2) | 2.4 (0.1) | 1.6 (0.1) | 1.3 (0.0) |
| Formic acid | n.d. | n.d. | 1.1 (0.2) | 1.1 (0.0) | 1.0 (0.1) | 1.0 (0.0) |
| HMF | n.d. | n.d. | 0.5 (0.1) | 0.5 (0.0) | 0.6 (0.0) | 0.5 (0.0) |
| Furfural | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Phenols | 1.4 (0.1) | 1.4 (0.1) | 1.4 (0.3) | 1.7 (0.1) | 1.3 (0.1) | 1.4 (0.0) |
| Total inhibitors | 2.5 | 2.4 | 5.4 | 5.7 | 4.5 | 4.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedő, S.; Fehér, A.; Khunnonkwao, P.; Jantama, K.; Fehér, C. Optimized Bioconversion of Xylose Derived from Pre-Treated Crop Residues into Xylitol by Using Candida boidinii. Agronomy 2021, 11, 79. https://doi.org/10.3390/agronomy11010079
Bedő S, Fehér A, Khunnonkwao P, Jantama K, Fehér C. Optimized Bioconversion of Xylose Derived from Pre-Treated Crop Residues into Xylitol by Using Candida boidinii. Agronomy. 2021; 11(1):79. https://doi.org/10.3390/agronomy11010079
Chicago/Turabian StyleBedő, Soma, Anikó Fehér, Panwana Khunnonkwao, Kaemwich Jantama, and Csaba Fehér. 2021. "Optimized Bioconversion of Xylose Derived from Pre-Treated Crop Residues into Xylitol by Using Candida boidinii" Agronomy 11, no. 1: 79. https://doi.org/10.3390/agronomy11010079
APA StyleBedő, S., Fehér, A., Khunnonkwao, P., Jantama, K., & Fehér, C. (2021). Optimized Bioconversion of Xylose Derived from Pre-Treated Crop Residues into Xylitol by Using Candida boidinii. Agronomy, 11(1), 79. https://doi.org/10.3390/agronomy11010079