
A Novel Pinkish-White Flower Color Variant Is Caused by a New Allele of Flower Color Gene W1 in Wild Soybean (Glycine soja)

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. RT-PCR and Sequence Analysis
2.3. Multiple Alignment Analysis of W1 Proteins
2.4. Prediction by Protein Variation Effect Analyzer (PROVEAN)
2.5. SNP-Based Genetic Analysis
3. Results and Discussion
3.1. Genetic Analysis of New Pinkish-White Flower Variant of Wild Soybean
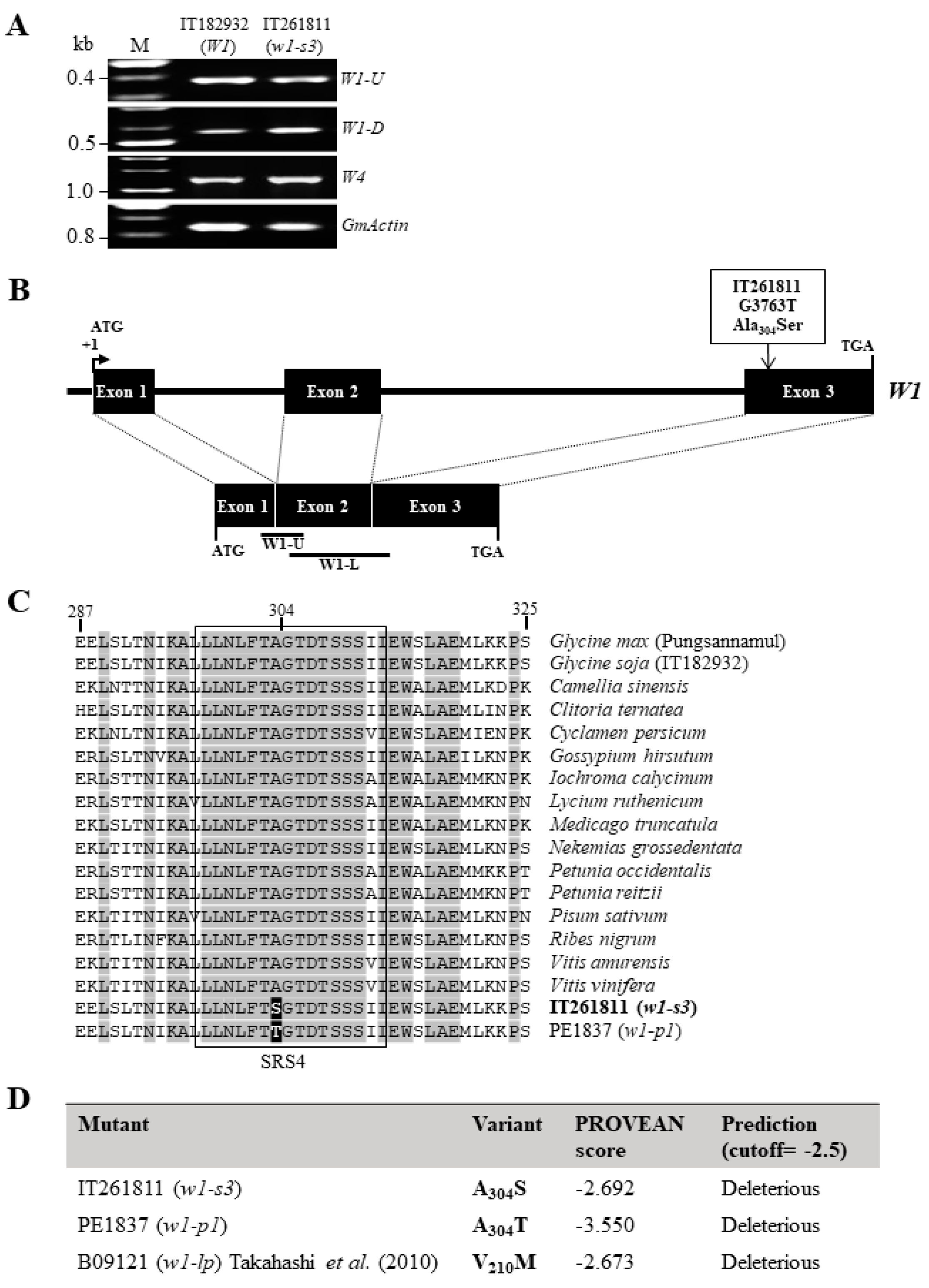
3.2. Molecular Analysis of the w1-s3 Variant
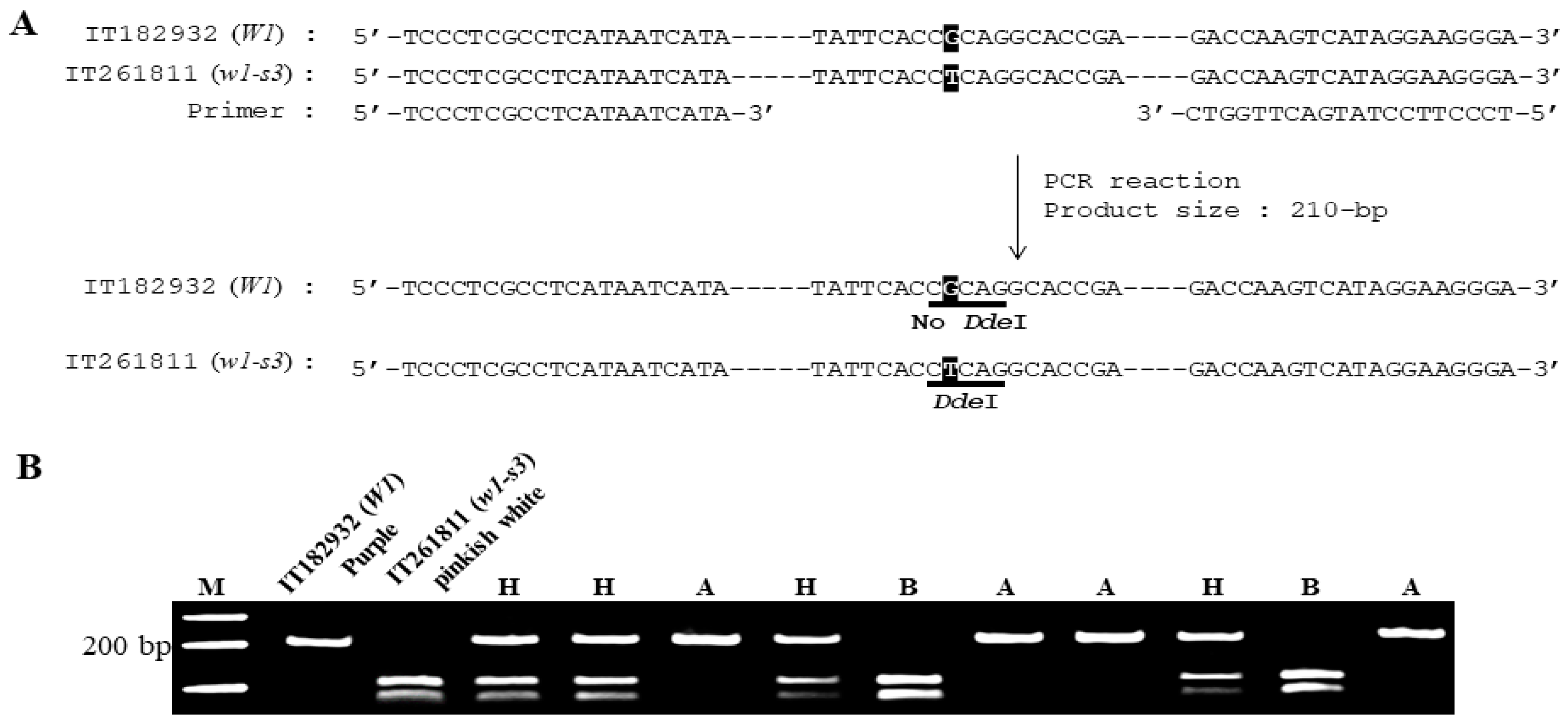
3.3. Co-Segregation of the W1 Polymorphism with Flower Color Phenotype
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Moreau, C.; Ambrose, M.J.; Turner, L.; Hill, L.; Ellis, T.H.; Hofer, J.M. The B gene of pea encodes a defective flavonoid 3’,5’-hydroxylase, and confers pink flower color. Plant Physiol. 2012, 159, 759–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buzzell, R.; Buttery, B.; MacTavish, D. Biochemical genetics of black pigmentation of soybean seed. J. Hered. 1987, 78, 53–54. [Google Scholar] [CrossRef]
- Tanner, G. Condensed tannins. In Plant Pigments and Their Manipulation; Davies, K.M., Ed.; Oxford: Blackwell, UK, 2004; Volume 14, pp. 154–184. [Google Scholar]
- Matsubara, K.; Kodama, H.; Kokubun, H.; Watanabe, H.; Ando, T. Two novel transposable elements in a cytochrome P450 gene govern anthocyanin biosynthesis of commercial petunias. Gene 2005, 358, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, T.; Nishihara, M.; Mishiba, K.; Hirano, H.; Yamamura, S. Two different transposable elements inserted in flavonoid 3’,5’-hydroxylase gene contribute to pink flower coloration in Gentiana scabra. Mol. Genet. Genom. 2006, 275, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Snowden, K.C.; Napoli, C.A. Psl: A novel Spm-like transposable element from Petunia hybrida. Plant J. 1998, 14, 43–54. [Google Scholar] [CrossRef]
- Park, G.T.; Sundaramoorthy, J.; Lee, J.-D.; Kim, J.H.; Seo, H.S.; Song, J.T. Elucidation of molecular identity of the W3 locus and its implication in determination of flower colors in soybean. PLoS ONE 2015, 10, e0142643. [Google Scholar] [CrossRef] [Green Version]
- Palmer, R.G.; Pfeiffer, T.W.; Buss, G.R.; Kilen, T.C. Qualitative Genetics. In Soybeans: Improvement, Production, and Uses, 3rd ed.; Shibles, M., Harper, J.E., Wilson, R.F., Shoemaker, R.C., Eds.; Madison Inc.: Madison, WI, USA, 2004. [Google Scholar]
- Takahashi, R.; Githiri, S.M.; Hatayama, K.; Dubouzet, E.G.; Shimada, N.; Aoki, T.; Ayabe, S.; Iwashina, T.; Toda, K.; Matsumura, H. A single-base deletion in soybean flavonol synthase gene is associated with magenta flower color. Plant Mol. Biol. 2007, 63, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Zabala, G.; Vodkin, L.O. A rearrangement resulting in small tandem repeats in the F3′ 5′ H gene of white flower genotypes is associated with the soybean W1 locus. Crop Sci. 2007, 47, S–113–S–124. [Google Scholar] [CrossRef]
- Park, G.T.; Sundaramoorthy, J.; Park, J.B.; Lee, J.D.; Choi, K.S.; Kim, J.H.; Seo, H.S.; Park, S.-K.; Song, J.T. Diversity of the W1 gene encoding flavonoid 3 ‘, 5 ‘-hydroxylase in white- and purple-flowered soybeans. Plant Genet. Resour. 2015, 13, 213–218. [Google Scholar] [CrossRef]
- Sundaramoorthy, J.; Park, G.T.; Lee, J.D.; Kim, J.H.; Seo, H.S.; Song, J.T. Genetic and molecular regulation of flower pigmentation in soybean. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 555–562. [Google Scholar] [CrossRef]
- Chen, Y.; Nelson, R.L. Identification and characterization of a white-flowered wild soybean plant. Crop Sci. 2004, 44, 339–342. [Google Scholar] [CrossRef]
- Sundaramoorthy, J.; Park, G.T.; Chang, J.H.; Lee, J.D.; Kim, J.H.; Seo, H.S.; Chung, G.; Song, J.T. Identification and molecular analysis of four new alleles at the W1 locus associated with flower color in soybean. PLoS ONE 2016, 11, e0159865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, R.; Morita, Y.; Nakayama, M.; Kanazawa, A.; Abe, J. An active CACTA-family transposable element is responsible for flower variegation in wild soybean Glycine soja. Plant Genome 2012, 5, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Dubouzet, J.G.; Matsumura, H.; Yasuda, K.; Iwashina, T. A new allele of flower color gene W1 encoding flavonoid 3 ‘5 ‘-hydroxylase is responsible for light purple flowers in wild soybean Glycine soja. BMC Plant Biol. 2010, 10, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, D.; Atkinson, T.; Noll, A.; Johnson, C.; Espinosa, K.; Boelter, J.; Abel, S.; Dhatt, B.K.; Barta, T.; Singsaas, E.; et al. Soybean proteins GmTic110 and GmPsbP are crucial for chloroplast development and function. Plant Sci. 2016, 252, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Park, G.T.; Sundaramoorthy, J.; Lee, J.D.; Kim, J.H.; Park, S.K.; Seo, H.S.; Song, J.T. Association of new mutant allele, w4-nw, at W4 locus with near-white flower color in soybean. Crop Sci. 2016, 56, 1857–1864. [Google Scholar] [CrossRef]
- Choi, Y.; Sims, G.E.; Murphy, S.; Miller, J.R.; Chan, A.P. Predicting the functional effect of amino acid substitutions and indels. PLoS ONE 2012, 7, e46688. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwig, E.E.; Hinson, K. Inheritance of flower color of soybeans. Crop Sci. 1962, 2, 152–153. [Google Scholar] [CrossRef] [Green Version]
- Park, G.T.; Sundaramoorthy, J.; Lee, S.; Lee, J.D.; Kim, J.H.; Park, S.K.; Seo, H.S.; Chung, G.; Song, J.T. Color variation in a novel Glycine soja mutant w4-s1 with pinkish-white flowers is controlled by a single recessive allele at the W4 locus. Crop Sci. 2017, 57, 3112–3121. [Google Scholar] [CrossRef]
- Fang, C.; Ma, Y.; Wu, S.; Liu, Z.; Wang, Z.; Yang, R.; Hu, G.; Zhou, Z.; Yu, H.; Zhang, M.; et al. Genome-wide association studies dissect the genetic networks underlying agronomical traits in soybean. Genome Biol. 2017, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Joshi, T.; Wang, J.; Zhang, H.; Chen, S.; Zeng, S.; Xu, B.; Xu, D. The evolution of soybean knowledge base (SoyKB). In Plant Genomics Databases; Dijk, A.V., Ed.; Humana Press: New York, NY, USA, 2017; pp. 149–159. [Google Scholar]
- Valliyodan, B.; Qiu, D.; Patil, G.; Zeng, P.; Huang, J.; Dai, L.; Chen, C.; Li, Y.; Joshi, T.; Song, L.; et al. Landscape of genomic diversity and trait discovery in soybean. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, S.W.; Hayhurst, G.P.; Smith, G.; Lightfoot, T.; Wong, M.M.; Simula, A.P.; Acland, M.J.; Sternberg, M.J.E.; Lennard, M.S.; Tucker, G.T.; et al. Evidence that aspartic acid 301 is a critical substrate-contact residue in the active site of cytochrome P450 2D6. J. Biol. Chem. 1995, 270, 29055–29058. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Fang, Y.; Li, X.; Zhang, H.; Liu, M.; Yang, H.; Kang, Z.; Li, Y.; Wang, Y. Mechanism of the plant cytochrome P450 for herbicide resistance: A modelling study. J. Enzym. Inhib. Med. Chem. 2013, 28, 1182–1191. [Google Scholar] [CrossRef] [PubMed]
- Doss, C.G.P.; Nagasundaram, N.; Chakraborty, C.; Chen, L.; Zhu, H. Extrapolating the effect of deleterious nsSNPs in the binding adaptability of flavopiridol with CDK7 protein: A molecular dynamics approach. Hum. Genom. 2013, 7, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Flower Color | W1 | |
|---|---|---|
| Segregation Ratios | ||
| P/PW * | A/H/B ** | |
| Observed | 95:29 | 31:64:29 |
| Expected | 93:31 | 31:62:31 |
| Ratio | 3:1 | 1:2:1 |
| χ2 | 0.0056 | 0.0063 |
| Probability † | 0.94 | 0.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sundaramoorthy, J.; Park, G.-T.; Jo, H.; Lee, J.-D.; Seo, H.-S.; Song, J.-T. A Novel Pinkish-White Flower Color Variant Is Caused by a New Allele of Flower Color Gene W1 in Wild Soybean (Glycine soja). Agronomy 2021, 11, 1001. https://doi.org/10.3390/agronomy11051001
Sundaramoorthy J, Park G-T, Jo H, Lee J-D, Seo H-S, Song J-T. A Novel Pinkish-White Flower Color Variant Is Caused by a New Allele of Flower Color Gene W1 in Wild Soybean (Glycine soja). Agronomy. 2021; 11(5):1001. https://doi.org/10.3390/agronomy11051001
Chicago/Turabian StyleSundaramoorthy, Jagadeesh, Gyu-Tae Park, Hyun Jo, Jeong-Dong Lee, Hak-Soo Seo, and Jong-Tae Song. 2021. "A Novel Pinkish-White Flower Color Variant Is Caused by a New Allele of Flower Color Gene W1 in Wild Soybean (Glycine soja)" Agronomy 11, no. 5: 1001. https://doi.org/10.3390/agronomy11051001
APA StyleSundaramoorthy, J., Park, G. -T., Jo, H., Lee, J. -D., Seo, H. -S., & Song, J. -T. (2021). A Novel Pinkish-White Flower Color Variant Is Caused by a New Allele of Flower Color Gene W1 in Wild Soybean (Glycine soja). Agronomy, 11(5), 1001. https://doi.org/10.3390/agronomy11051001