
Nitrogen Source Influences the Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Macadamia integrifolia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growing Conditions
2.2. Processing of Plant and Media Samples
2.3. Analysis of Plant Growth and Physiology
2.4. Physicochemical Analysis of Leachates, Plant Material and Potting Media
2.5. Isotopic Analysis
2.6. Genetic Identification of PGPR SOS3 Copies
2.7. Assessment of Microbial-Biomass N
2.8. Statistical Analysis
3. Results
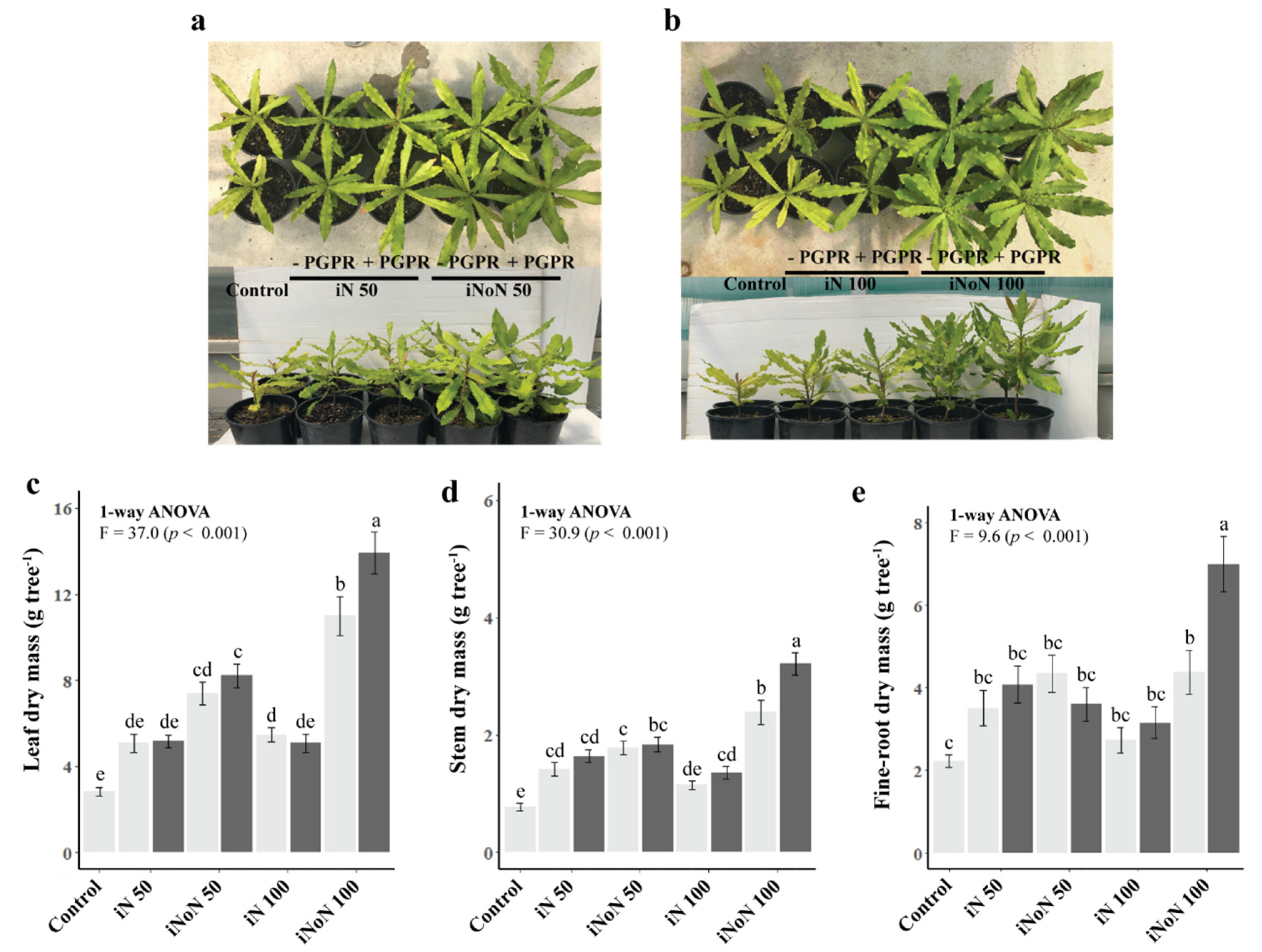
3.1. Tree Growth, Root Responses and Nutrient Status
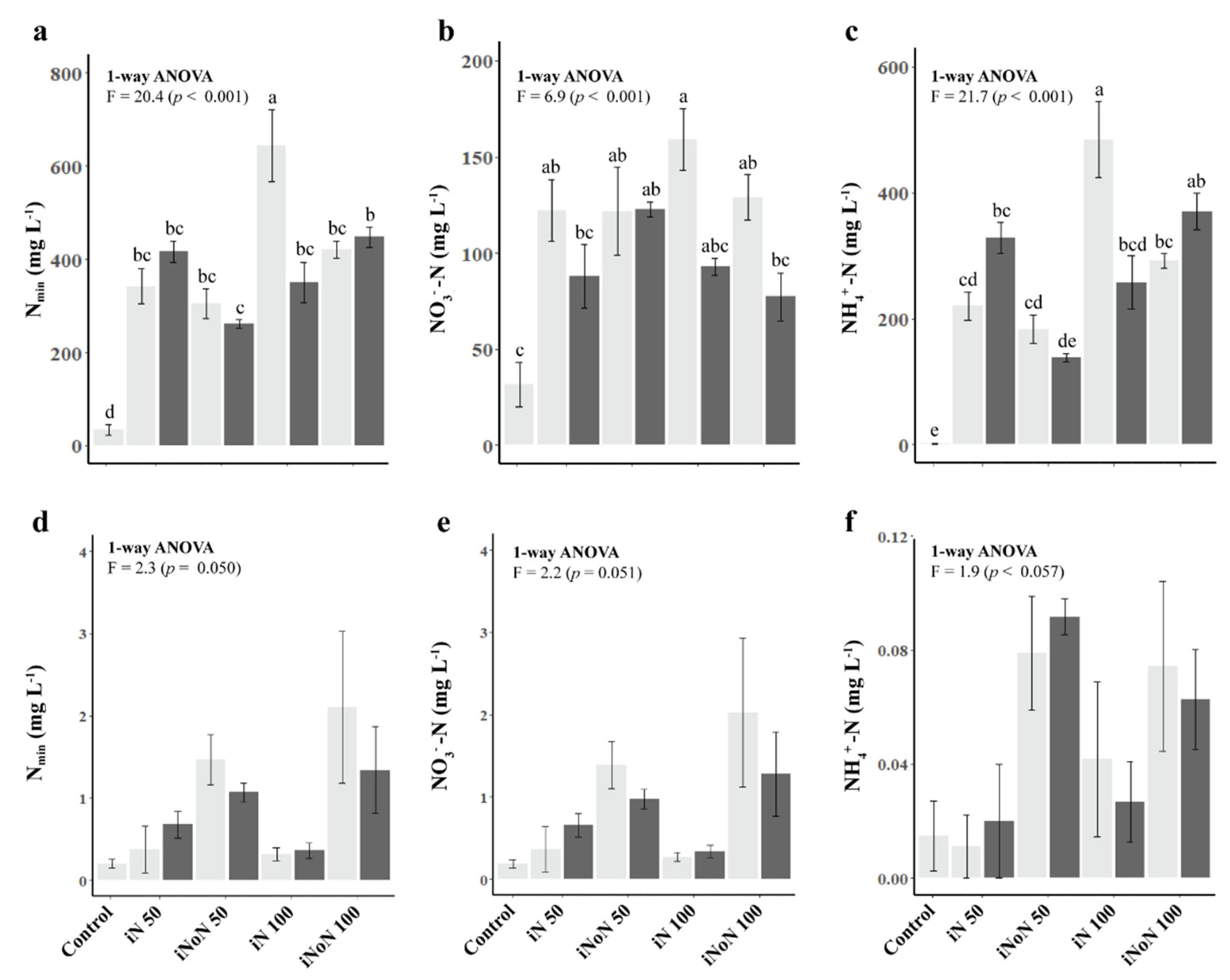
3.2. Mineral N in Soil-Leachate
3.3. Potting Media Physicochemical Properties
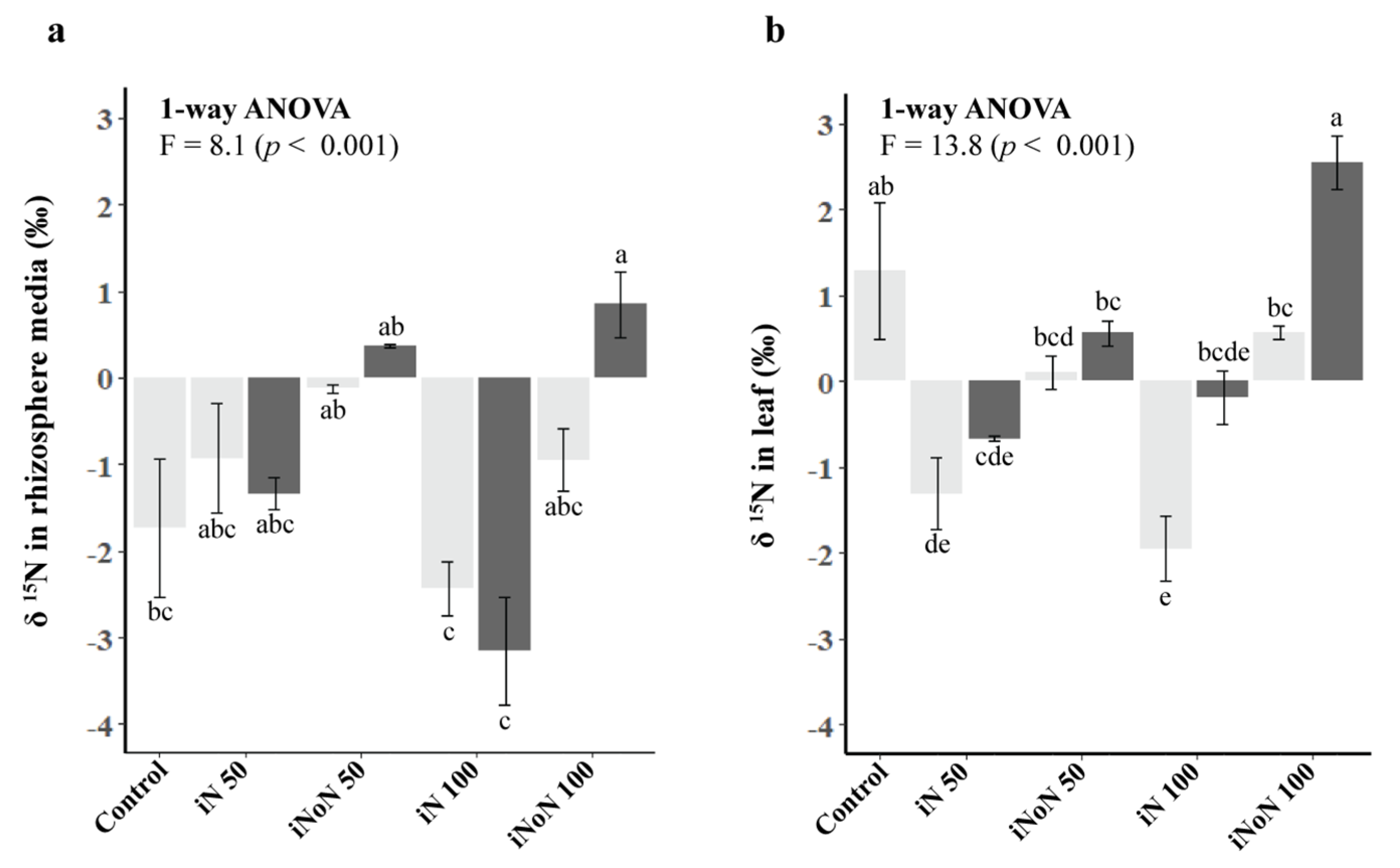
3.4. δ15N Isotope Signatures
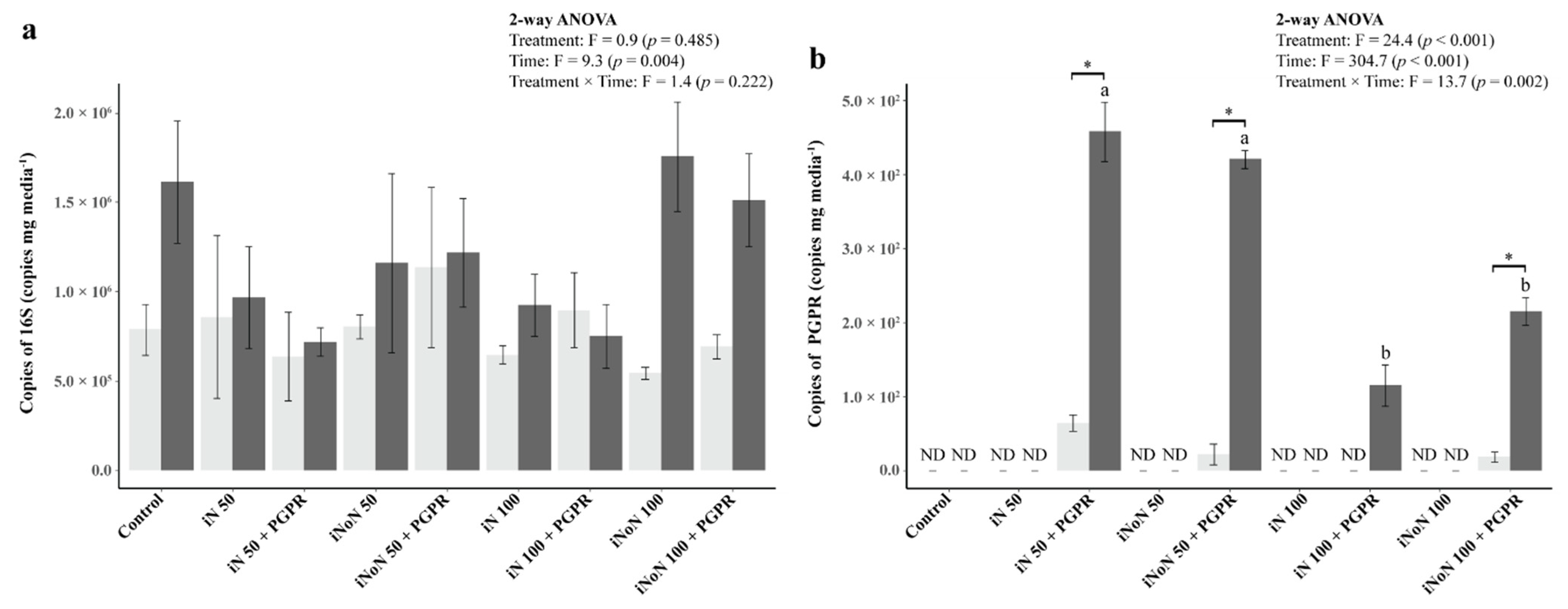
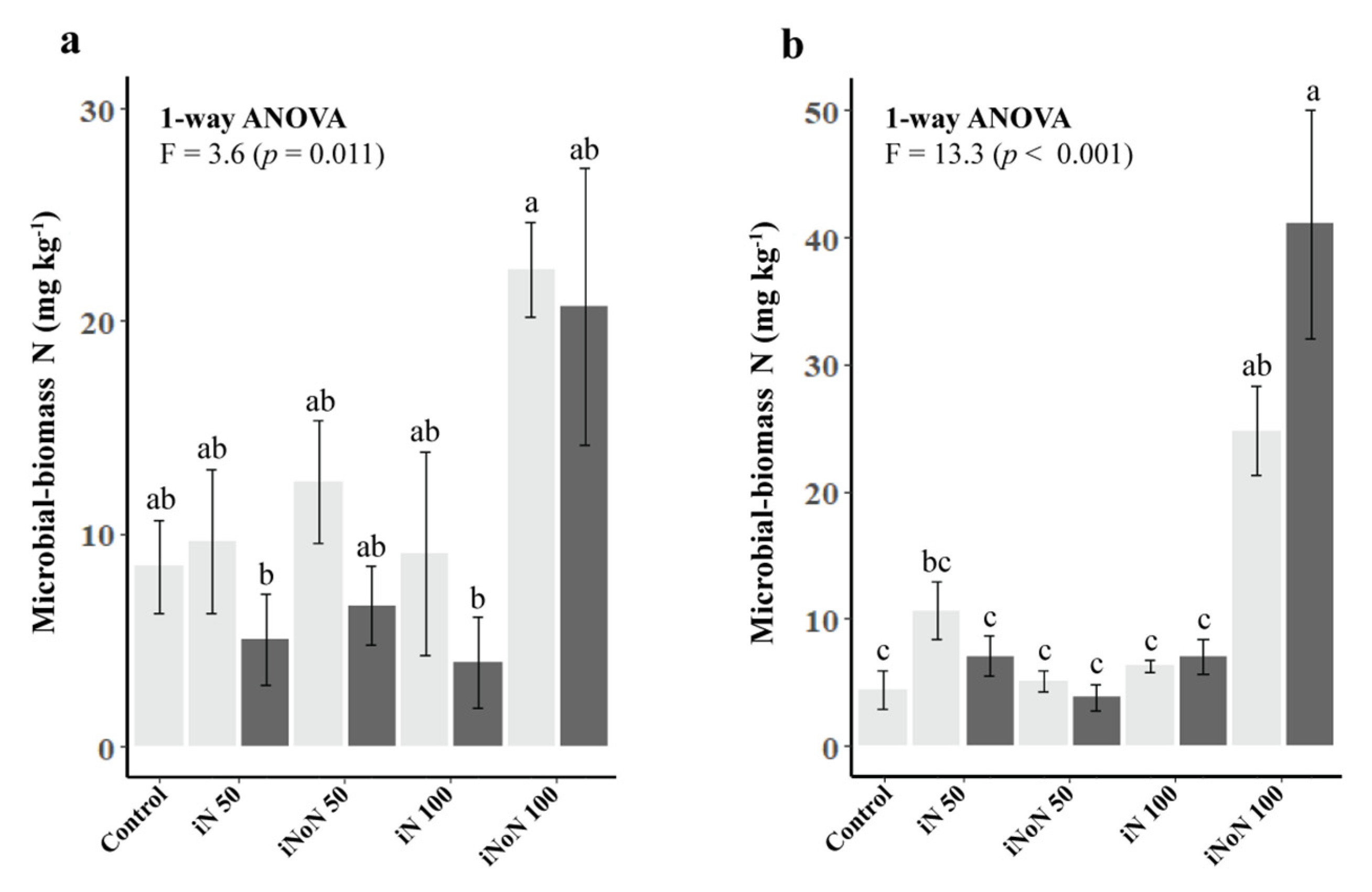
3.5. Microbial Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Technical report: Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737. [Google Scholar] [CrossRef]
- Rockström, J.; Steffen, W.; Noone, K.; Persson, A.; Chapin, F.S.; Lambin, E.F.; Lenton, T.M.; Scheffer, M.; Folke, C.; Schellnhuber, H.J.; et al. A safe operating space for humanity. Nat. Cell Biol. 2009, 461, 472–475. [Google Scholar] [CrossRef]
- Gruber, N.; Galloway, J.N. An Earth-system perspective of the global nitrogen cycle. Nat. Cell Biol. 2008, 451, 293–296. [Google Scholar] [CrossRef]
- Majsztrik, J.C.; Fernandez, R.T.; Fisher, P.R.; Hitchcock, D.R.; Lea-Cox, J.; Owen, J.S.; Oki, L.R.; White, S.A. Water use and treatment in container-grown specialty crop production: A review. Water Air Soil Pollut. 2017, 228, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.L.; Polasky, S. Agricultural sustainability and intensive production practices. Nat. Cell Biol. 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Bolan, N.; Szogi, A.; Chuasavathi, T.; Seshadri, B.; Rothrock, M.J.; Panneerselvam, P. Uses and management of poultry litter. Worlds Poult. Sci. J. 2010, 66, 673–698. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Maiti, S.K. Effect of organic manures on the growth of Cymbopogon citratus and Chrysopogon zizanioides for the Phytoremediation of chromite-asbestos mine waste: A pot scale experiment. Int. J. Phytoremediat. 2014, 17, 437–447. [Google Scholar] [CrossRef]
- Manlay, R.J.; Feller, C.; Swift, M. Historical evolution of soil organic matter concepts and their relationships with the fertility and sustainability of cropping systems. Agric. Ecosyst. Environ. 2007, 119, 217–233. [Google Scholar] [CrossRef]
- Gutser, R.; Ebertseder, T.; Weber, A.; Schraml, M.; Schmidhalter, U. Short-term and residual availability of nitrogen after long-term application of organic fertilizers on arable land. J. Plant Nutr. Soil Sci. 2005, 168, 439–446. [Google Scholar] [CrossRef]
- Sørensen, P.; Amato, M. Remineralisation and residual effects of N after application of pig slurry to soil. Eur. J. Agron. 2002, 16, 81–95. [Google Scholar] [CrossRef]
- Schimel, J.P.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Rees, R.; Castle, K. Nitrogen recovery in soils amended with organic manures combined with inorganic fertilisers. Agronomy 2002, 22, 739–746. [Google Scholar] [CrossRef]
- Wen, Z.; Shen, J.; Blackwell, M.; Li, H.; Zhao, B.; Yuan, H. Combined applications of nitrogen and phosphorus fertilizers with manure increase maize yield and nutrient uptake via stimulating root growth in a long-term experiment. Pedosphere 2016, 26, 62–73. [Google Scholar] [CrossRef]
- Zhong, W.; Gu, T.; Wang, W.; Zhang, B.; Lin, X.; Huang, Q.; Shen, W. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 2009, 326, 511–522. [Google Scholar] [CrossRef]
- Francioli, D.; Schulz, E.; Lentendu, G.; Wubet, T.; Buscot, F.; Reitz, T. Mineral vs. organic amendments: Microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 2016, 7, 1446. [Google Scholar] [CrossRef] [Green Version]
- Denk, T.R.; Mohn, J.; Decock, C.; Lewicka-Szczebak, D.; Harris, E.; Butterbach-Bahl, K.; Kiese, R.; Wolf, B. The nitrogen cycle: A review of isotope effects and isotope modeling approaches. Soil Biol. Biochem. 2017, 105, 121–137. [Google Scholar] [CrossRef] [Green Version]
- Gardner, J.B.; Drinkwater, L.E. The fate of nitrogen in grain cropping systems: A meta-analysis of15N field experiments. Ecol. Appl. 2009, 19, 2167–2184. [Google Scholar] [CrossRef] [PubMed]
- Chalk, P.M.; Inácio, C.T.; Chen, D. An overview of contemporary advances in the usage of 15N natural abundance (δ15N) as a tracer of agro-ecosystem N cycle processes that impact the environment. Agric. Ecosyst. Environ. 2019, 283, 106570. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Genet. 2020, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lareen, A.; Burton, F.; Schäfer, P. Plant root-microbe communication in shaping root microbiomes. Plant Mol. Biol. 2016, 90, 575–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Vivanco, J.M.; Shen, Q. The unseen rhizosphere root–soil–microbe interactions for crop production. Curr. Opin. Microbiol. 2017, 37, 8–14. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Mano, H.; Morisaki, H. Endophytic bacteria in the rice plant. Microbes Environ. 2009, 24, 193. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Kloepper, J.W.; Ryu, C.-M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Bakker, P.A.H.M.; Pieterse, C.M.J. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [Green Version]
- Vessey, J.K. Plant growth promoting rhizobacteria as biofertilizers. Plant Soil 2003, 255, 571–586. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E.; Prabhu, S.R.; Hernandez, J. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2013, 378, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Yadav, R.; Suman, A.; Prasad, S.; Prakash, O. Effect of Gluconacetobacter diazotrophicus and Trichoderma viride on soil health, yield and N-economy of sugarcane cultivation under subtropical climatic conditions of India. Eur. J. Agron. 2009, 30, 296–303. [Google Scholar] [CrossRef]
- Riveracruz, M.; Trujillonarcia, A.; Cordovaballona, G.; Kohler, J.; Caravaca, F.; Roldan, A. Poultry manure and banana waste are effective biofertilizer carriers for promoting plant growth and soil sustainability in banana crops. Soil Biol. Biochem. 2008, 40, 3092–3095. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Redding, M.; Pratt, C.; Wang, W. Plant growth promoting rhizobacteria increase the efficiency of fertilisers while reducing nitrogen loss. J. Environ. Manag. 2019, 233, 337–341. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Watanarojanaporn, N.; Jaemsaeng, R. Plant growth promoting rhizobacteria enhance the efficiency of the combination of organic and chemical fertilisers in sugarcane. Open J. Ecol. 2020, 10, 440–444. [Google Scholar] [CrossRef]
- Rayment, G.E.; Lyons, D.J. Soil Chemical Methods—Australasia; CSIRO Publishing: Collingwood, VIC, Australia, 2010; ISBN 9780643101364. [Google Scholar]
- Miller, R.O.; Kissel, D.E. Comparison of soil pH methods on soils of North America. Soil Sci. Soc. Am. J. 2010, 74, 310–316. [Google Scholar] [CrossRef]
- Munter, R.; Grande, R. Plant tissue and soil extract analysis by ICP—Atomic emission spectrometry. In Developments in Atomic Plasma Spectrochemical Analysis; Heyden: London, UK, 1981; pp. 653–672. [Google Scholar]
- Martinie, G.D.; Schilt, A.A. Wet oxidation efficiencies of perchloric acid mixtures for various organic substances and the identities of residual matter. Anal. Chem. 1976, 48, 70–74. [Google Scholar] [CrossRef]
- Amato, M. Assay for microbial biomass based on ninhydrin-reactive nitrogen in extracts of fumigated soils. Soil Biol. Biochem. 1988, 20, 107–114. [Google Scholar] [CrossRef]
- RStudio Team RStudio: Integrated Development for RStudio; PBC: Boston, MA, USA, 2020; Available online: http//www.rstudio.com (accessed on 25 May 2020).
- Rahman, M.; Sabir, A.A.; Mukta, J.A.; Alam Khan, M.; Mohi-Ud-Din, M.; Miah, G.; Rahman, M.; Islam, M.T. Plant probiotic bacteria Bacillus and Paraburkholderia improve growth, yield and content of antioxidants in strawberry fruit. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Poupin, M.J.; Timmermann, T.; Vega, A.; Zuñiga, A.; González, B. Effects of the plant growth-promoting bacterium Burkholderia phytofirmans PsJN throughout the life cycle of Arabidopsis thaliana. PLoS ONE 2013, 8, e69435. [Google Scholar] [CrossRef] [Green Version]
- Puri, A.; Padda, K.P.; Chanway, C.P. Sustaining the growth of Pinaceae trees under nutrient-limited edaphic conditions via plant-beneficial bacteria. PLoS ONE 2020, 15, e0238055. [Google Scholar] [CrossRef]
- Paungfoo-Lonhienne, C.; Lonhienne, T.G.A.; Yeoh, Y.K.; Donose, B.C.; Webb, R.I.; Parsons, J.; Liao, W.; Sagulenko, E.; Lakshmanan, P.; Hugenholtz, P.; et al. Crosstalk between sugarcane and a plant-growth promoting Burkholderia species. Sci. Rep. 2016, 6, 37389. [Google Scholar] [CrossRef]
- Ritchie, G.A.; Tanaka, Y.; Meade, R.; Duke, S.D. Field survival and early height growth of Douglas-fir rooted cuttings: Relationship to stem diameter and root system quality. For. Ecol. Manag. 1993, 60, 237–256. [Google Scholar] [CrossRef]
- Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Küsel, K.; Rillig, M.C.; Rivett, D.; Salles, J.F.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukanova, K.; Сhеbоtаr, V.; Meyer, J.; Bibikova, T. Effect of plant growth-promoting Rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Zandi, P. Effects of exogenously applied plant growth regulators in combination with PGPR on the physiology and root growth of chickpea (Cicer arietinum) and their role in drought tolerance. J. Plant Interact. 2018, 13, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Chamam, A.; Sanguin, H.; Bellvert, F.; Meiffren, G.; Comte, G.; Wisniewski-Dyé, F.; Bertrand, C.; Prigent-Combaret, C. Plant secondary metabolite profiling evidences strain-dependent effect in the Azospirillum–Oryza sativa association. Phytochemistry 2013, 87, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Contesto, C.; Desbrosses, G.; Lefoulon, C.; Béna, G.; Borel, F.; Galland, M.; Gamet, L.; Varoquaux, F.; Touraine, B. Effects of rhizobacterial ACC deaminase activity on Arabidopsis indicate that ethylene mediates local root responses to plant growth-promoting rhizobacteria. Plant Sci. 2008, 175, 178–189. [Google Scholar] [CrossRef]
- Zúñiga, A.; Poupin, M.J.; Donoso, R.; Ledger, T.; Guiliani, N.; Gutiérrez, R.A.; González, B. Quorum sensing and indole-3-acetic acid degradation play a role in colonization and plant growth promotion of Arabidopsis thaliana by Burkholderia phytofirmans PsJN. Mol. Plant Microbe Interact. 2013, 26, 546–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef]
- Cambui, C.A.; Svennerstam, H.; Gruffman, L.; Nordin, A.; Ganeteg, U.; Näsholm, T. Patterns of plant biomass partitioning depend on nitrogen source. PLoS ONE 2011, 6, e19211. [Google Scholar] [CrossRef] [Green Version]
- Britto, D.T.; Kronzucker, H. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Lori, M.; Symanczik, S.; Mäder, P.; Efosa, N.; Jaenicke, S.; Buegger, F.; Tresch, S.; Goesmann, A.; Gattinger, A. Distinct nitrogen provisioning from organic amendments in soil as influenced by farming system and water regime. Front. Environ. Sci. 2018, 6. [Google Scholar] [CrossRef]
- Edmeades, D.C. The long-term effects of manures and fertilisers on soil productivity and quality: A review. Nutr. Cycl. Agroecosyst. 2003, 66, 165–180. [Google Scholar] [CrossRef]
- Kramer, S.B.; Reganold, J.P.; Glover, J.D.; Bohannan, B.J.M.; Mooney, H.A. Reduced nitrate leaching and enhanced denitrifier activity and efficiency in organically fertilized soils. Proc. Natl. Acad. Sci. USA 2006, 103, 4522–4527. [Google Scholar] [CrossRef] [Green Version]
- Florio, A.; Pommier, T.; Gervaix, J.; Bérard, A.; Le Roux, X. Soil C and N statuses determine the effect of maize inoculation by plant growth-promoting rhizobacteria on nitrifying and denitrifying communities. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szpak, P.; Longstaffe, F.J.; Millaire, J.-F.; White, C.D. Stable isotope biogeochemistry of seabird guano fertilization: Results from growth chamber studies with maize (Zea mays). PLoS ONE 2012, 7, e33741. [Google Scholar] [CrossRef]
- Karwat, H.; Egenolf, K.; Nuñez, J.; Rao, I.; Rasche, F.; Arango, J.; Moreta, D.; Arevalo, A.; Cadisch, G. Low 15N natural abundance in shoot tissue of Brachiaria humidicola is an indicator of reduced N losses due to Biological Nitrification Inhibition (BNI). Front. Microbiol. 2018, 9, 2383. [Google Scholar] [CrossRef]
- Pardo, L.H.; Hemond, H.F.; Montoya, J.P.; Fahey, T.J.; Siccama, T.G. Response of the natural abundance of 15N in forest soils and foliage to high nitrate loss following clear-cutting. Can. J. For. Res. 2002, 32, 1126–1136. [Google Scholar] [CrossRef]
- Högberg, P. Tansley Review No. 95 15 N natural abundance in soil-plant systems. New Phytol. 1997, 137, 179–203. [Google Scholar] [CrossRef]
- Perin, L.; Martinez-Aguilar, L.; Castro-Gonzalez, R.; Santos, P.E.-D.L.; Cabellos-Avelar, T.; Guedes, H.V.; Reis, V.M.; Caballero-Mellado, J. Diazotrophic Burkholderia species associated with field-grown maize and sugarcane. Appl. Environ. Microbiol. 2006, 72, 3103–3110. [Google Scholar] [CrossRef] [Green Version]
- Castro-González, R.; Martínez-Aguilar, L.; Ramírez-Trujillo, A.; Santos, P.E.-D.L.; Caballero-Mellado, J. High diversity of culturable Burkholderia species associated with sugarcane. Plant. Soil 2011, 345, 155–169. [Google Scholar] [CrossRef]
- Reed, S.C.; Cleveland, C.C.; Townsend, A.R. Functional ecology of free-living nitrogen fixation: A contemporary perspective. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 489–512. [Google Scholar] [CrossRef]
- Dobbelaere, S.; Vanderleyden, J.; Okon, Y. Plant growth-promoting effects of diazotrophs in the rhizosphere. Crit. Rev. Plant. Sci. 2003, 22, 107–149. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | PGPR Inoculation | N Level | N Form | N Content (g pot−1) | PGPR (g pot−1) |
|---|---|---|---|---|---|
| Control | - | Nil | Nil | 0 | - |
| iN 50 | −PGPR | Low | 100% Urea | 3 | - |
| +PGPR | Low | 100% Urea | 3 | 10 | |
| iNoN 50 | −PGPR | Low | 50% Urea, 50% EcoNPK™ | 3 | - |
| +PGPR | Low | 50% Urea, 50% EcoNPK™ | 3 | 10 | |
| iN 100 | −PGPR | High | 100% Urea | 6 | - |
| +PGPR | High | 100% Urea | 6 | 10 | |
| iNoN 100 | −PGPR | High | 50% Urea, 50% EcoNPK™ | 6 | - |
| +PGPR | High | 50% Urea, 50% EcoNPK™ | 6 | 10 |
| Control | iN 50 | iNoN 50 | iN 100 | iNoN 100 | F (p) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| - | −PGPR | +PGPR | −PGPR | +PGPR | −PGPR | +PGPR | −PGPR | +PGPR | ||
| Total biomass (g tree−1) | 7.7 ± 0.4d | 12.4 ± 1.0cd | 13.5 ± 0.9c | 16.4 ± 0.9bc | 16.7 ± 1.1bc | 11.6 ± 0.7cd | 11.8 ± 0.9cd | 21.2 ± 1.7b | 27.9 ± 1.8a | 28.3 (<0.001) |
| Chlorophyll | 24.1 ± 0.7d | 27.6 ± 0.6c | 27.8 ± 0.8c | 30.0 ± 0.9abc | 29.5 ± 0.8bc | 27.8 ± 0.8c | 28.0 ± 0.8c | 32.9 ± 0.9a | 32.4 ± 0.4ab | 13.2 (<0.001) |
| Shoot mass fraction (%) | 46.7 ± 1.2d | 53.0 ± 1.6cd | 51.6 ± 1.4cd | 55.9 ± 2.0bc | 60.7 ± 1.4ab | 57.1 ± 1.6abc | 55.0 ± 1.3bc | 63.8 ± 1.5a | 61.2 ± 1.4ab | 12.4 (<0.001) |
| Root mass fraction (%) | 53.3 ± 1.2a | 47.0 ± 1.6ab | 48.4 ± 1.4ab | 44.1 ± 2.0bc | 39.3 ± 1.4cd | 42.9 ± 1.6bcd | 45.0 ± 1.3bc | 36.2 ± 1.5d | 38.8 ± 1.4cd | 12.4 (<0.001) |
| R:S | 1.17 ± 0.06a | 0.93 ± 0.07b | 0.97 ± 0.06ab | 0.83 ± 0.06bc | 0.67 ± 0.04cd | 0.78 ± 0.05bcd | 0.84 ± 0.04bc | 0.58 ± 0.04d | 0.65 ± 0.04cd | 12.2 (<0.001) |
| Fine-root mass fraction (g g root−1) | 0.54 ± 0.02ab | 0.57 ± 0.02ab | 0.59 ± 0.02ab | 0.58 ± 0.02ab | 0.53 ± 0.02b | 0.52 ± 0.03b | 0.56 ± 0.02ab | 0.53 ± 0.02b | 0.63 ± 0.02a | 2.8 (0.007) |
| N uptake 1 (mg N plant−1) | 36.4 ± 2.2d | 61.5 ± 7.7cd | 63.8 ± 5.2cd | 95.7 ± 7.4bc | 95.1 ± 8.3bc | 65.3 ± 5.8cd | 66.3 ± 3.8cd | 132.5 ± 13.5b | 190.2 ± 13.5a | 31.2 (<0.001) |
| P uptake 2 (mg P g dry mass leaf−1) | 24.8 ± 3.9c | 32.8 ± 6.0c | 31.2 ± 3.9c | 42.0 ± 5.1bc | 52.5 ± 5.0bc | 22.7 ± 3.4c | 31.8 ± 3.4c | 65.3 ± 8.8ab | 94.8 ± 14.2a | 11.7 (<0.001) |
| K uptake 3 (mg K g dry mass leaf−1) | 39.5 ± 5.4d | 57.3 ± 8cd | 55.0 ± 6.4cd | 83.5 ± 7.1bcd | 95.9 ± 9bc | 52.3 ± 5.7cd | 60.1 ± 5.8cd | 127.3 ± 13.8b | 181.6 ± 21.2a | 19.2 (<0.001) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallart, M.; Paungfoo-Lonhienne, C.; Gonzalez, A.; Trueman, S.J. Nitrogen Source Influences the Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Macadamia integrifolia. Agronomy 2021, 11, 1064. https://doi.org/10.3390/agronomy11061064
Gallart M, Paungfoo-Lonhienne C, Gonzalez A, Trueman SJ. Nitrogen Source Influences the Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Macadamia integrifolia. Agronomy. 2021; 11(6):1064. https://doi.org/10.3390/agronomy11061064
Chicago/Turabian StyleGallart, Marta, Chanyarat Paungfoo-Lonhienne, Axayacatl Gonzalez, and Stephen J. Trueman. 2021. "Nitrogen Source Influences the Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Macadamia integrifolia" Agronomy 11, no. 6: 1064. https://doi.org/10.3390/agronomy11061064
APA StyleGallart, M., Paungfoo-Lonhienne, C., Gonzalez, A., & Trueman, S. J. (2021). Nitrogen Source Influences the Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Macadamia integrifolia. Agronomy, 11(6), 1064. https://doi.org/10.3390/agronomy11061064