
The Genetic Diversity and Population Structure of Different Geographical Populations of Bottle Gourd (Lagenaria siceraria) Accessions Based on Genotyping-by-Sequencing




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. GBS, Read Clustering, and SNP Calling
2.3. Analysis of Genetic Diversity Parameters and Molecular Variance
2.4. Population Structure and Genetic Relationship
2.5. Mining of Simple Sequence Repeats Markers
3. Results
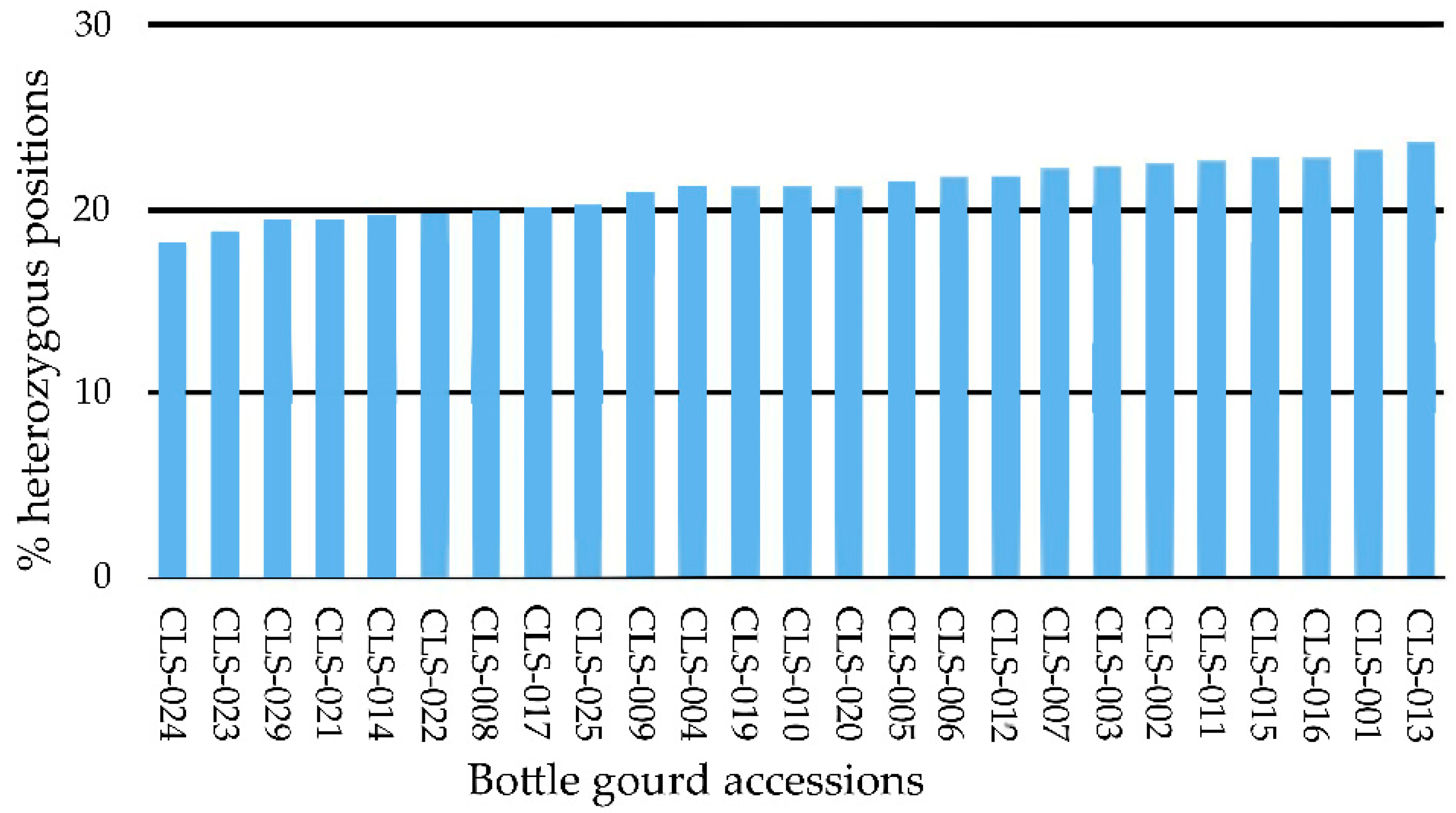
3.1. GBS Analysis
3.2. AMOVA
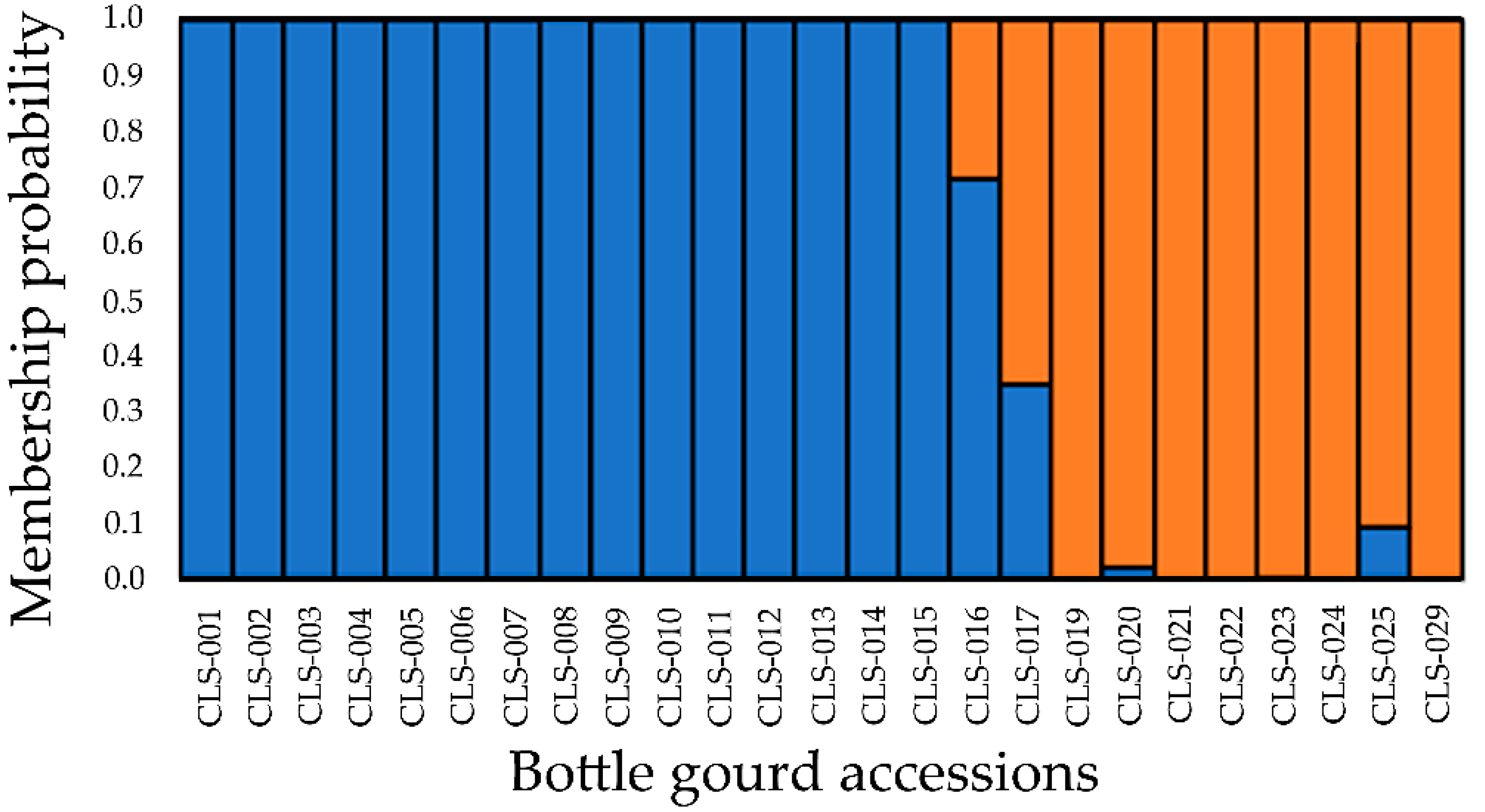
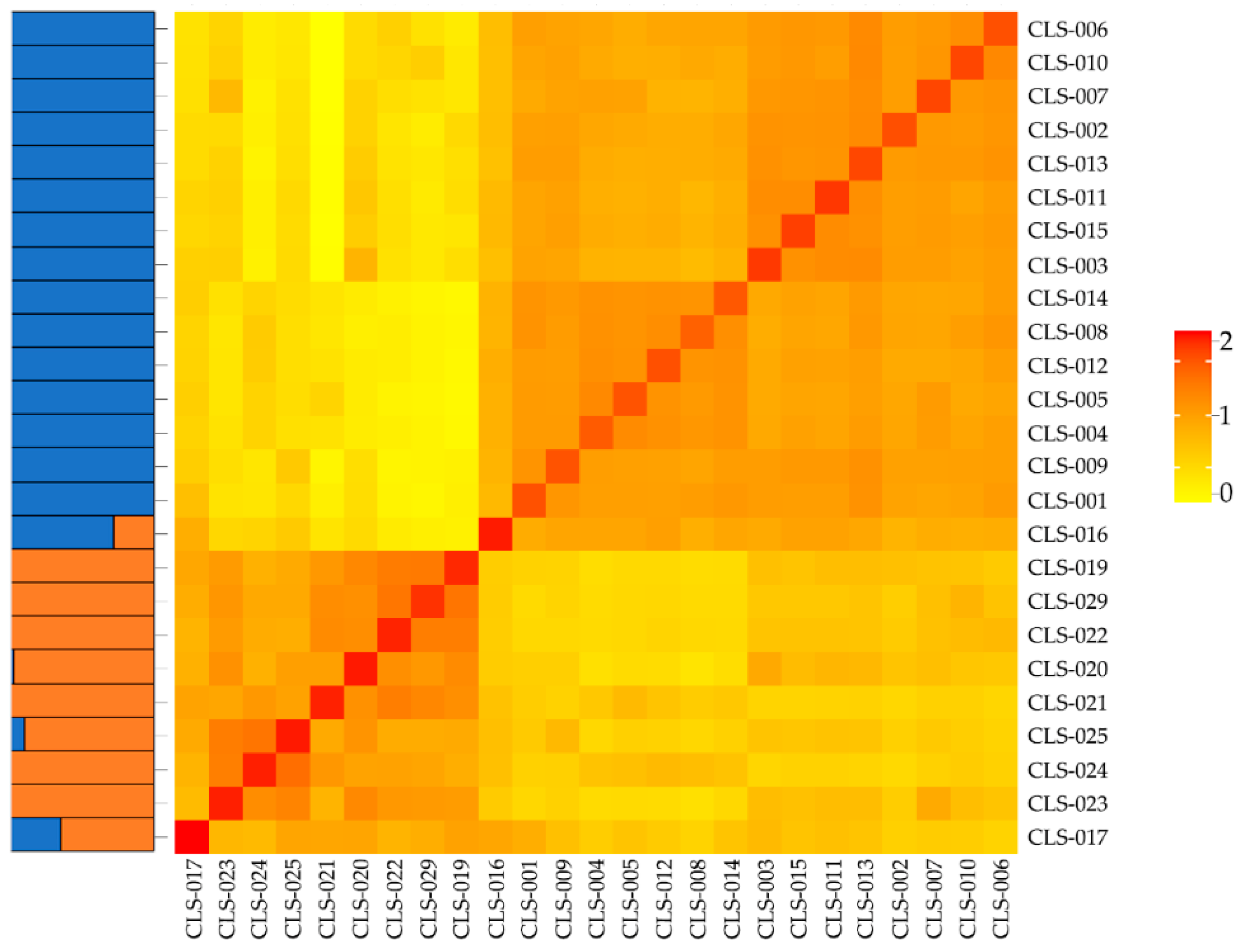
3.3. Population Structure
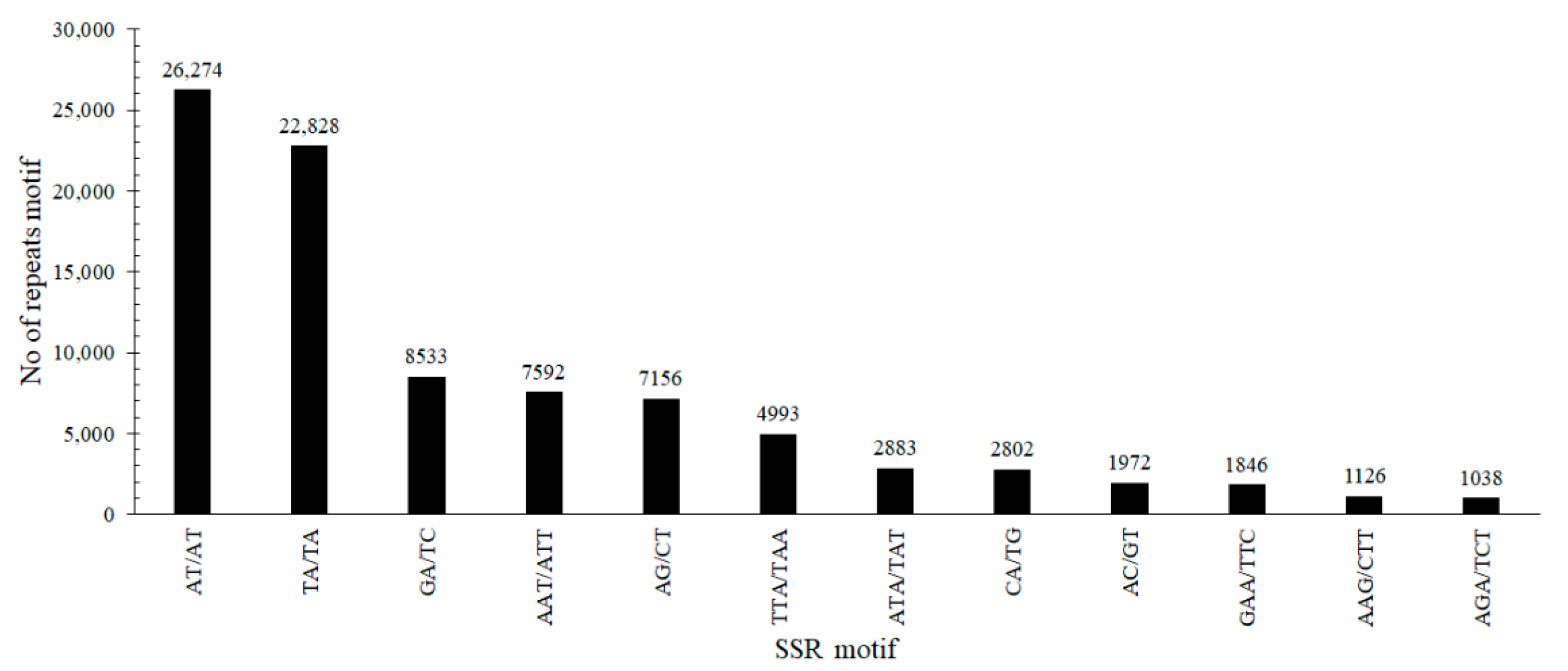
3.4. Bottle Gourd SSR Locus Identification and the Frequency of SSRs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Achigan-Dako, E.G.; Fuchs, J.; Ahanchede, A.; Blattner, F.R. Flow cytometric analysis in Lagenaria siceraria (Cucurbitaceae) indicates correlation of genome size with usage types and growing elevation. Plant Syst. Evol. 2018, 276, 9–19. [Google Scholar] [CrossRef]
- Kalpana, V.N.; Alarjani, K.M.; Rajeswari, V.D. Enhancing malaria control using Lagenaria siceraria and its mediated zinc oxide nanoparticles against the vector Anopheles stephensi and its parasite Plasmodium falciparum. Sci. Rep. 2020, 10, 21568. [Google Scholar] [CrossRef] [PubMed]
- Yetisir, H.; Sari, N. Effect of different rootstock on plant growth, yield and quality of watermelon. Aust. J. Exp. Agric. 2003, 43, 1269–1274. [Google Scholar] [CrossRef]
- King, S.R.; Davis, A.R.; Liu, W.; Levi, A. Grafting for disease resistance. HortScience 2008, 43, 1673–1676. [Google Scholar] [CrossRef] [Green Version]
- Ulas, A.; Doganci, E.; Ulas, F.; Yetisir, H. Root-growth characteristics contributing to genotypic variation in nitrogen efficiency of bottle gourd and rootstock potential for watermelon. Plants 2019, 8, 77. [Google Scholar] [CrossRef] [Green Version]
- Aslam, W.; Noor, R.S.; Hussain, F.; Ameen, M.; Ullah, S.; Chen, H. Evaluating morphological growth, yield, and postharvest fruit quality of cucumber (Cucumis sativus L.) grafted on cucurbitaceous rootstocks. Agriculture 2020, 10, 101. [Google Scholar] [CrossRef] [Green Version]
- Guler, Z.; Candir, E.; Yetisir, H.; Karaca, F.; Solmaz, I. Volatile organic compounds in watermelon (Citrullus lanatus) grafted onto 21 local and two commercial bottle gourd (Lagenaria siceraria) rootstocks. J. Hortic. Sci. Biotechnol. 2014, 89, 448–452. [Google Scholar] [CrossRef]
- Guler, Z.; Karaca, F.; Yetisir, H. Volatile compounds in the peel and flesh of cucumber (cucumis sativus L.) grafted onto bottle gourd (Lagenaria siceraria) rootstocks. J. Hortic. Sci. Biotechnol. 2013, 88, 123–128. [Google Scholar] [CrossRef]
- Decker-Walters, D.; Wilkins-Ellert, M. Discovery and genetic assessment of wild bottle gourd from Zimbabwe. Econ. Bot. 2004, 58, 501–508. [Google Scholar] [CrossRef]
- Erickson, D.L.; Smith, B.D.; Clarke, A.C.; Sandweiss, D.H.; Tuross, N. An Asian origin for a 10,000-year-old domesticated plant in the Americas. Proc. Natl. Acad. Sci. USA 2005, 102, 18315–18320. [Google Scholar] [CrossRef] [Green Version]
- Kobiakova, J.A. The bottle gourd. Bull. Appl. Bot. Genet. Plant. Breed. 1930, 23, 475–520. [Google Scholar]
- Schlumbaum, A.; Vandorpe, P. A short history of Lagenaria siceraria (bottle gourd) in the Roman provinces: Morphotypes and archaeogenetics. Veg. Hist. Archaeobot. 2012, 21, 499–509. [Google Scholar] [CrossRef] [Green Version]
- Mashilo, J.; Shimelis, H.; Odindo, A. Phenotypic and genotypic characterization of bottle gourd [Lagenaria siceraria (Molina) Standl.] and implications for breeding: A Review. Sci. Hortic. 2017, 222, 136–144. [Google Scholar] [CrossRef]
- Sivaraj, N.; Pandravada, S.R. Morphological diversity for fruit characters in bottle gourd germplasm from tribal pockets of Telangana region of Andhra Pradesh, India. Asian Agrihist. 2005, 9, 305–310. [Google Scholar]
- Mashilo, J.; Shimelis, H.; Odindo, A.; Amelework, B. Genetic diversity of South African bottle gourd [Lagenaria siceraria (Molina) standl.] landraces revealed by simple sequence repeat markers. HortScience 2016, 51, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Buthelezi, L.G.; Mavengahama, S.; Ntuli, N.R. Morphological variation and heritability studies of Lagenaria siceraria landraces from northern Kwazulu-Natal, South Africa. Biodiversitas 2019, 20, 922–930. [Google Scholar] [CrossRef] [Green Version]
- Sarao, N.K.; Pathak, M.; Kaur, N.; Kaur, K. Microsatellite-based DNA fingerprinting and genetic diversity of bottle gourd genotypes. Plant Genet. Resour. 2014, 12, 156–159. [Google Scholar] [CrossRef]
- Mashilo, J.; Shimelis, H.; Odindo, A. Genetic diversity of bottle gourd (Lagenaria siceraria (Molina) Standl.) landraces of South Africa assessed by morphological traits and simple sequence repeat markers. S. Afr. J. Plant Soil 2016, 33, 113–124. [Google Scholar] [CrossRef]
- Xu, P.; Xu, S.; Wu, X.; Tao, Y.; Wang, B.; Wang, S.; Li, G. Population genomic analyses from low-coverage RAD-Seq data: A case study on the non-model cucurbit bottle gourd. Plant J. 2014, 77, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Bhawna, M.; Abdin, Z.; Arya, L.; Saha, D.; Sureja, A.K.; Pandey, C.; Verma, M. Population structure and genetic diversity in bottle gourd [Lagenaria siceraria (Mol.) Standl.] germplasm from India assessed by ISSR markers. Plant Syst. Evol. 2014, 300, 767–773. [Google Scholar] [CrossRef]
- Konan, J.A.; Guyot, R.; Koffi, K.K.; Vroh-Bi, I.; Zoro, A.I.B. Molecular confirmation of varietal status in bottle gourd (Lagenaria siceraria) using genotyping-by-sequencing. Genome 2020, 63, 535–545. [Google Scholar] [CrossRef]
- Bhattacharjee, R.; Agre, P.; Bauchet, G.; de Koeyer, D.; Lopez-Montes, A.; Kumar, P.; Asiedu, R. Genotyping-by-sequencing to unlock genetic diversity and population structure in white yam (dioscorea rotundata poir). Agronomy 2020, 10, 1437. [Google Scholar] [CrossRef]
- Yang, X.; Tan, B.; Liu, H.; Zhu, W.; Xu, L.; Wang, Y.; Kang, H. Genetic Diversity and Population Structure of Asian and European Common Wheat Accessions Based on Genotyping-By-Sequencing. Front. Genet. 2020, 11, 1157. [Google Scholar] [CrossRef]
- Zhu, H.; Song, P.; Koo, D.H.; Guo, L.; Li, Y.; Sun, S.; Weng, Y.; Yang, L. Genome wide characterization of simple sequence repeats in watermelon genome and their application in comparative mapping and genetic diversity analysis. BMC Genom. 2016, 17, 557. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Dias, L.; Vilanova, S.; Fita, A.; Prohens, J.; Rodríguez-Burruezo, A. Genetic diversity, population structure, and relationships in a collection of pepper (Capsicum spp.) landraces from the Spanish centre of diversity revealed by genotyping-by-sequencing (GBS). Hortic. Res. 2019, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Shamimuzzaman, M.; Sun, H.; Salse, J.; Sui, X.; Wilder, A.; Fei, Z. The bottle gourd genome provides insights into Cucurbitaceae evolution and facilitates mapping of a Papaya ring-spot virus resistance locus. Plant J. 2017, 92, 963–975. [Google Scholar] [CrossRef] [Green Version]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; 1000 Genomes Project Analysis Group. The variant call format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef] [PubMed]
- Poland, J.A.; Rife, T.W. Genotyping-by-Sequencing for Plant Breeding and Genetics. Plant Genome 2012, 5, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Money, D.; Gardner, K.; Migicovsky, Z.; Schwaninger, H.; Zhong, G.Y.; Myles, S. LinkImpute: Fast and accurate genotype imputation for nonmodel organisms. G3 Genes Genom. Genet. 2015, 5, 2383–2390. [Google Scholar] [CrossRef] [Green Version]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, L. GMATA: An integrated software package for genome-scale SSR mining, marker development and viewing. Front. Plant Sci. 2016, 7, 1350. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, P.; Wu, X.; Wu, X.; Wang, B.; Huang, Y.; Li, G. GourdBase: A genome-centered multi-omics database for the bottle gourd (Lagenaria siceraria), an economically important cucurbit crop. Sci. Rep. 2018, 8, 3604. [Google Scholar] [CrossRef]
- Xu, P.; Wu, X.; Luo, J.; Wang, B.; Liu, Y.; Ehlers, J.D.; Li, G. Partial sequencing of the bottle gourd genome reveals markers useful for phylogenetic analysis and breeding. BMC Genom. 2011, 12, 467. [Google Scholar] [CrossRef] [Green Version]
- Scheben, A.; Batley, J.; Edwards, D. Genotyping-by-sequencing approaches to characterize crop genomes: Choosing the right tool for the right application. Plant. Biotechnol. J. 2017, 15, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Fountain, E.D.; Pauli, J.N.; Reid, B.N.; Palsbøll, P.J.; Peery, M.Z. Finding the right coverage: The impact of coverage and sequence quality on single nucleotide polymorphism genotyping error rates. Mol. Ecol. Resour. 2016, 16, 966–978. [Google Scholar] [CrossRef]
- Swarts, K.; Li, H.; Romero Navarro, J.A.; An, D.; Romay, M.C.; Hearne, S.; Bradbury, P.J. Novel Methods to Optimize Genotypic Imputation for Low-Coverage, Next-Generation Sequence Data in Crop Plants. Plant Genome 2014, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Zhang, Z.; Liu, L.; Staub, J.E.; Han, Y.; Cheng, Z.; Huang, S. An integrated genetic and cytogenetic map of the cucumber genome. PLoS ONE 2009, 4, e5795. [Google Scholar] [CrossRef]
- Zhu, H.; Guo, L.; Song, P.; Luan, F.; Hu, J.; Sun, X.; Yang, L. Development of genome-wide SSR markers in melon with their cross-species transferability analysis and utilization in genetic diversity study. Mol. Breed. 2016, 36, 1–14. [Google Scholar] [CrossRef]
- Gürcan, K.; Say, A.; Yetişir, H.; Denli, H. A study of genetic diversity in bottle gourd [Lagenaria siceraria (Molina) Standl.] population, and implication for the historical origins on bottle gourds in Turkey. Genet. Resour. Crop. Evol. 2015, 62, 321–333. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Genetic diversity in Egyptian bottle gourd genotypes based on ISSR markers. Ecol. Genet. Genom. 2021, 18, 6–11. [Google Scholar] [CrossRef]
- Mashilo, J.; Odindo, A.O.; Shimelis, H.A.; Musenge, P.; Tesfay, S.Z.; Magwaza, L.S. Drought tolerance of selected bottle gourd [Lagenaria siceraria (Molina) Standl.] landraces assessed by leaf gas exchange and photosynthetic efficiency. Plant Physiol. Biochem. 2017, 120, 75–87. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Singh, N.; Choudhury, D.R.; Singh, A.K.; Kumar, S.; Srinivasan, K.; Tyagi, R.K.; Singh, R. Comparison of SSR and SNP markers in estimation of genetic diversity and population structure of Indian rice varieties. PLoS ONE 2013, 8, e84136. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, J.; Fan, X.; Htwe, N.M.P.S.; Wang, S.; Huang, W.; Qiu, L. Assessing the numbers of SNPs needed to establish molecular IDs and characterize the genetic diversity of soybean cultivars derived from Tokachi nagaha. Crop. J. 2017, 5, 326–336. [Google Scholar] [CrossRef]
- Eltaher, S.; Sallam, A.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.; Baenziger, P.S. Genetic diversity and population structure of F3:6 Nebraska Winter wheat genotypes using genotyping-by-sequencing. Front. Genet. 2018, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Chesnokov, Y.; Artemyeva, A. Evaluation of the measure of polymorphism information of genetic diversity. Agric. Biol. 2015, 50, 571–578. [Google Scholar]
- Yetişir, H.; Şakar, M.; Serçe, S. Collection and morphological characterization of Lagenaria siceraria germplasm from the Mediterranean region of Turkey. Genet. Resour. Crop. Evol. 2008, 55, 1257–1266. [Google Scholar] [CrossRef]
- Yildiz, M.; Cuevas, H.E.; Sensoy, S.; Erdinc, C.; Baloch, F.S. Transferability of Cucurbita SSR markers for genetic diversity assessment of Turkish bottle gourd (Lagenaria siceraria) genetic resources. Biochem. Syst. Ecol. 2015, 59, 45–53. [Google Scholar] [CrossRef]
- Heiser, C.B. Tropical Forest Ecosystems in Africa and South America: A Comparative Review; Meggers, B.J., Ayensu, E.S., Eds.; Duckworth, W.D.: London, UK, 1973; pp. 121–128. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome Number | Number of SNPs Markers | PIC | MAF | Ho | He |
|---|---|---|---|---|---|
| 1 | 1285 | 0.298 | 0.236 | 0.184 | 0.159 |
| 2 | 1580 | 0.297 | 0.238 | 0.208 | 0.161 |
| 3 | 1305 | 0.305 | 0.239 | 0.179 | 0.162 |
| 4 | 1448 | 0.287 | 0.224 | 0.186 | 0.155 |
| 5 | 1272 | 0.292 | 0.228 | 0.177 | 0.157 |
| 6 | 1044 | 0.291 | 0.225 | 0.166 | 0.156 |
| 7 | 855 | 0.286 | 0.222 | 0.176 | 0.154 |
| 8 | 1103 | 0.275 | 0.211 | 0.150 | 0.147 |
| 9 | 1059 | 0.297 | 0.234 | 0.190 | 0.159 |
| 10 | 824 | 0.302 | 0.242 | 0.211 | 0.163 |
| 11 | 991 | 0.295 | 0.231 | 0.172 | 0.158 |
| Total/Average | 12766 | 0.293 | 0.230 | 0.183 | 0.157 |
| Component of Differentiation | DF | Mean Square | PVE (%) | Phi Statistics |
|---|---|---|---|---|
| Between populations | 2 | 28,618 | 35.9 | PT = 0.36 |
| Between samples within populations | 22 | 3958 | 16.3 | SP = 0.25 |
| Within samples | 25 | 2357 | 47.8 | ST = 0.52 |
| Total | - | - | 100 |
| Cluster | Individuals | Mean | Median | Mode * | SD | 95% Credible Interval | |
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| I | 9 | 0.40 | 0.40 | 0.40 | 0.0062 | 0.39 | 0.41 |
| II | 16 | 0.62 | 0.61 | 0.61 | 0.0052 | 0.60 | 0.62 |
| SSR Motifs | Number of Repeat Units of Each SSR Motif | Frequency (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 5 | 6 | 7 | 8 | 9 | 10 | >10 | Total | ||
| Dinucleotide | 28,400 | 10,526 | 6872 | 4695 | 3539 | 2679 | 12,971 | 69,682 | 72.86 |
| Trinucleotide | 9670 | 4589 | 2399 | 1386 | 855 | 603 | 2139 | 21,641 | 22.63 |
| Tetranucleotide | 2372 | 537 | 160 | 68 | 28 | 12 | 26 | 3203 | 3.35 |
| Pentanucleotide | 424 | 125 | 33 | 10 | 1 | 3 | 3 | 599 | 0.63 |
| Hexanucleotide | 250 | 82 | 16 | 4 | 2 | 1 | 1 | 356 | 0.37 |
| Heptanucleotide | 105 | 20 | 15 | 2 | 1 | 2 | 9 | 154 | 0.16 |
| Total | 41,221 | 15,879 | 9495 | 6165 | 4426 | 3300 | 15,149 | 95,635 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras-Soto, R.; Salvatierra, A.; Maldonado, C.; Mashilo, J. The Genetic Diversity and Population Structure of Different Geographical Populations of Bottle Gourd (Lagenaria siceraria) Accessions Based on Genotyping-by-Sequencing. Agronomy 2021, 11, 1677. https://doi.org/10.3390/agronomy11081677
Contreras-Soto R, Salvatierra A, Maldonado C, Mashilo J. The Genetic Diversity and Population Structure of Different Geographical Populations of Bottle Gourd (Lagenaria siceraria) Accessions Based on Genotyping-by-Sequencing. Agronomy. 2021; 11(8):1677. https://doi.org/10.3390/agronomy11081677
Chicago/Turabian StyleContreras-Soto, Rodrigo, Ariel Salvatierra, Carlos Maldonado, and Jacob Mashilo. 2021. "The Genetic Diversity and Population Structure of Different Geographical Populations of Bottle Gourd (Lagenaria siceraria) Accessions Based on Genotyping-by-Sequencing" Agronomy 11, no. 8: 1677. https://doi.org/10.3390/agronomy11081677
APA StyleContreras-Soto, R., Salvatierra, A., Maldonado, C., & Mashilo, J. (2021). The Genetic Diversity and Population Structure of Different Geographical Populations of Bottle Gourd (Lagenaria siceraria) Accessions Based on Genotyping-by-Sequencing. Agronomy, 11(8), 1677. https://doi.org/10.3390/agronomy11081677