
Plant-Derived Smoke Ameliorates Salt Stress in Wheat by Enhancing Expressions of Stress-Responsive Genes and Antioxidant Enzymatic Activity

 , , , , ,
, , , , ,  and
and 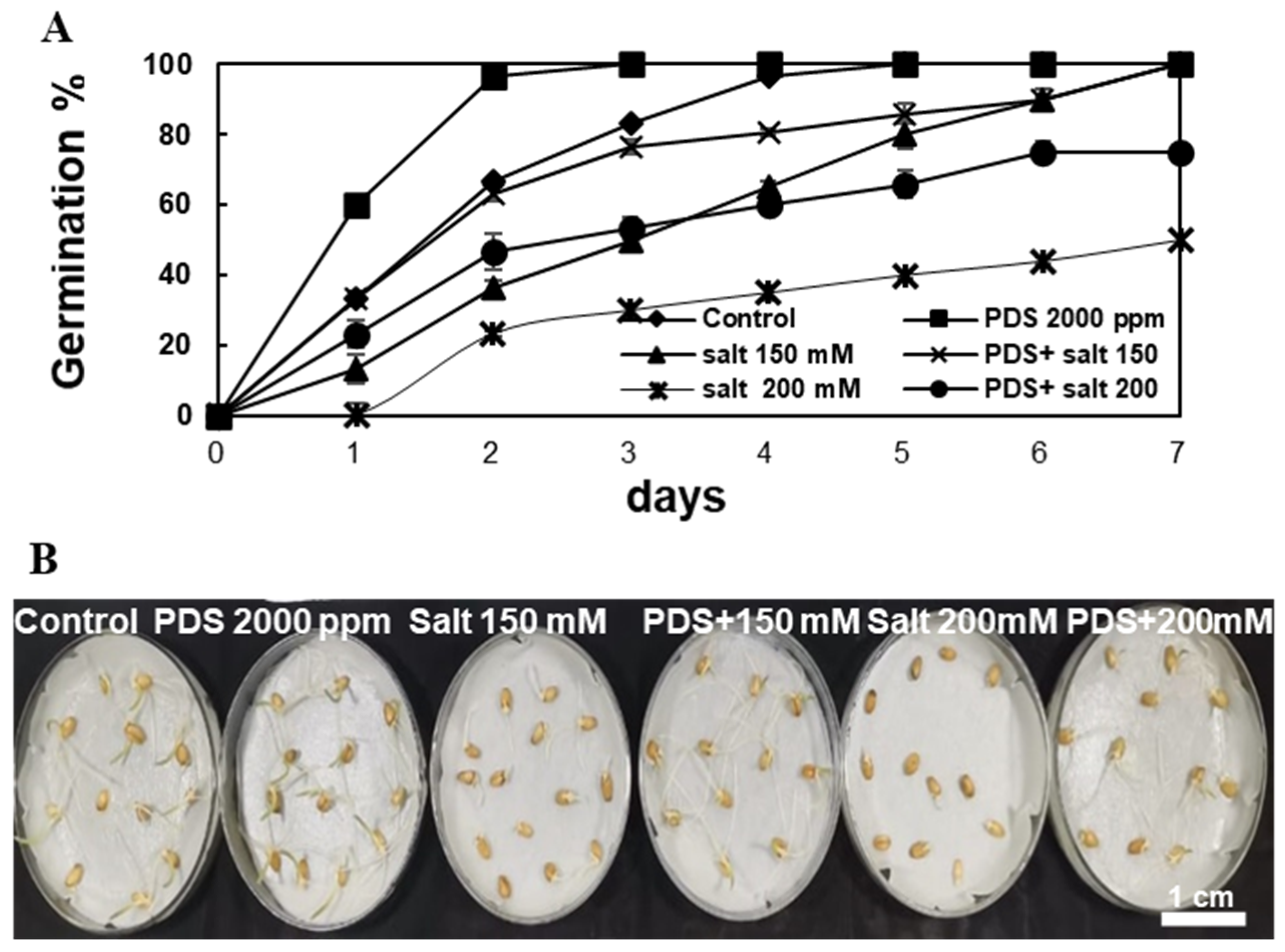{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Plant-Derived Smoke Solution Enhances Seed Germination under Normal and Salt Stress Condition
2.2. Plant-Derived Smoke Solution Modulates Expression of Germination Marker Genes under Salt Stress
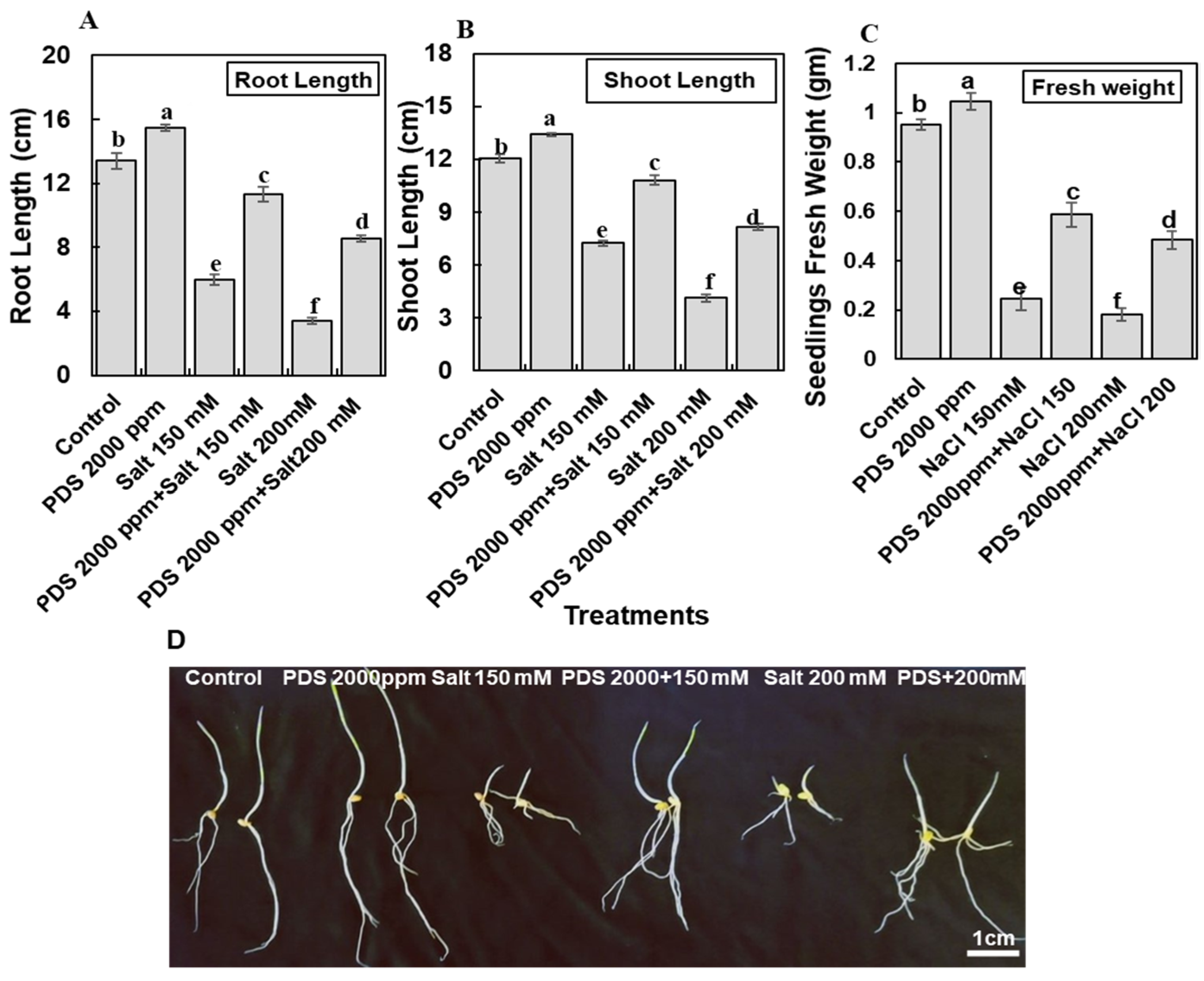
2.3. Application of PDS Improves Wheat Plantlets Growth and Fresh Weight under Salt Stress
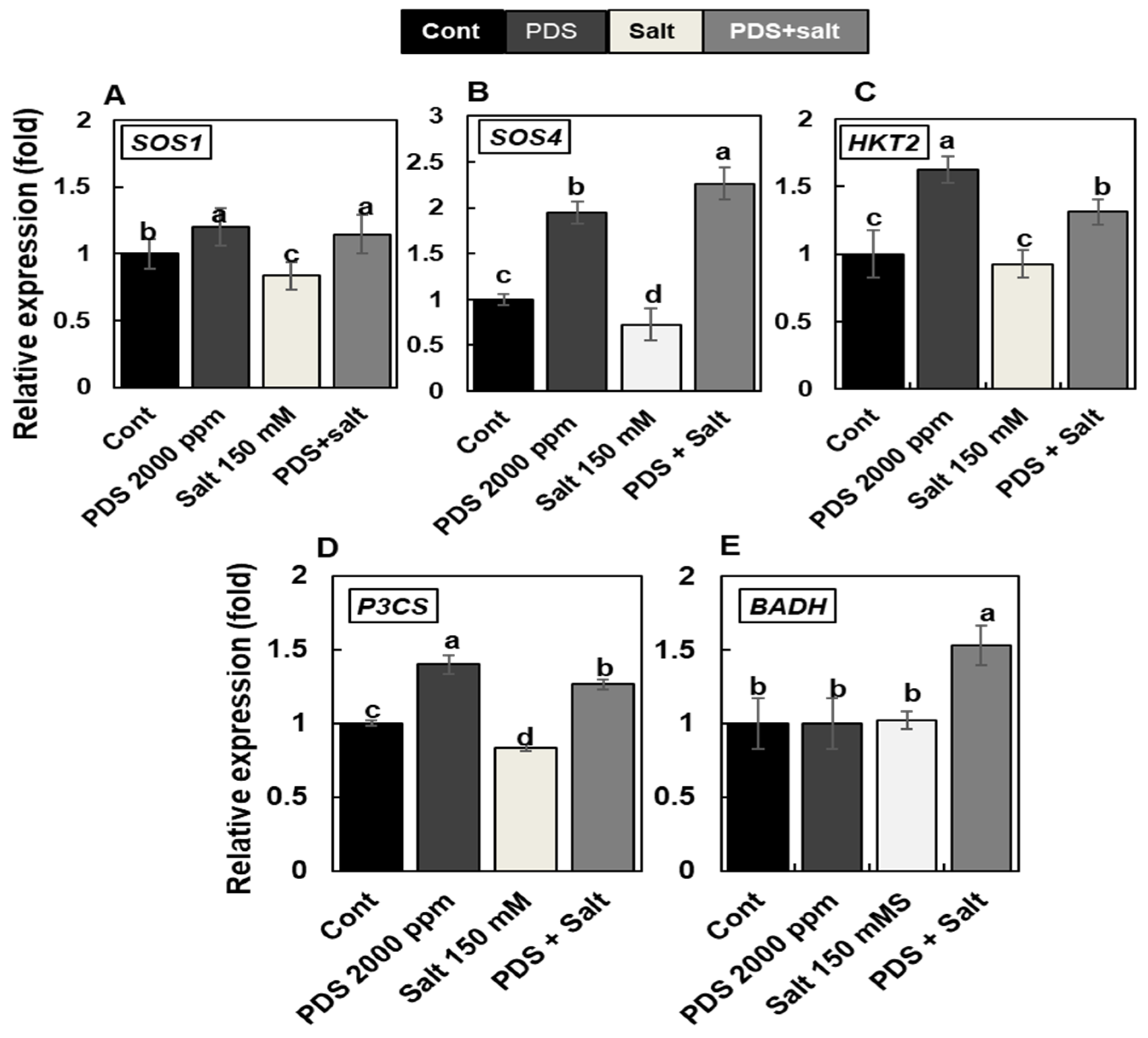
2.4. Plant-Derived Smoke Solution Modulates Expression of Salt Stress Marker Genes
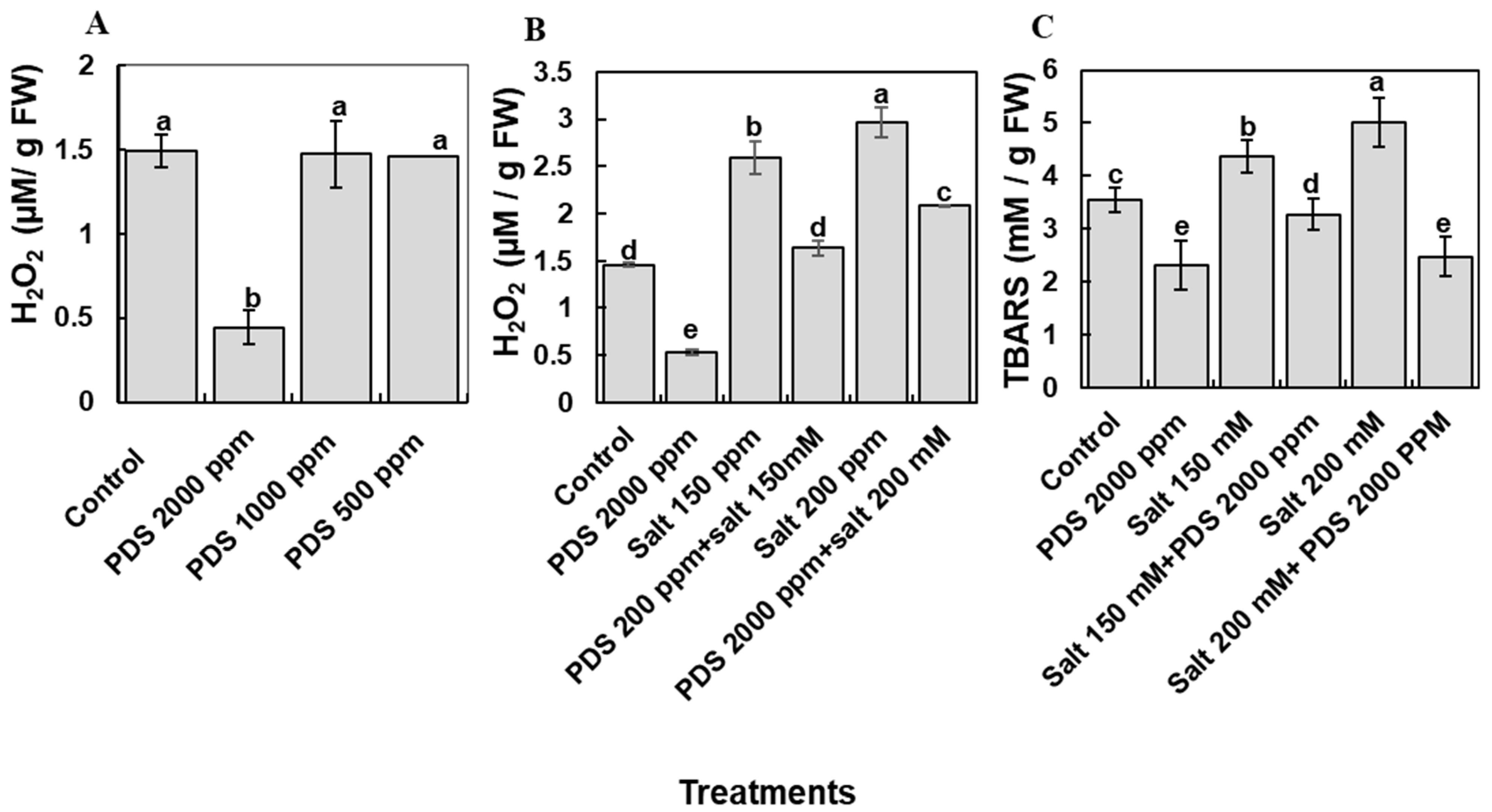
2.5. Plant-Derived Smoke Solution Reduces Oxidative Stress (H2O2) and Lipid Peroxidation
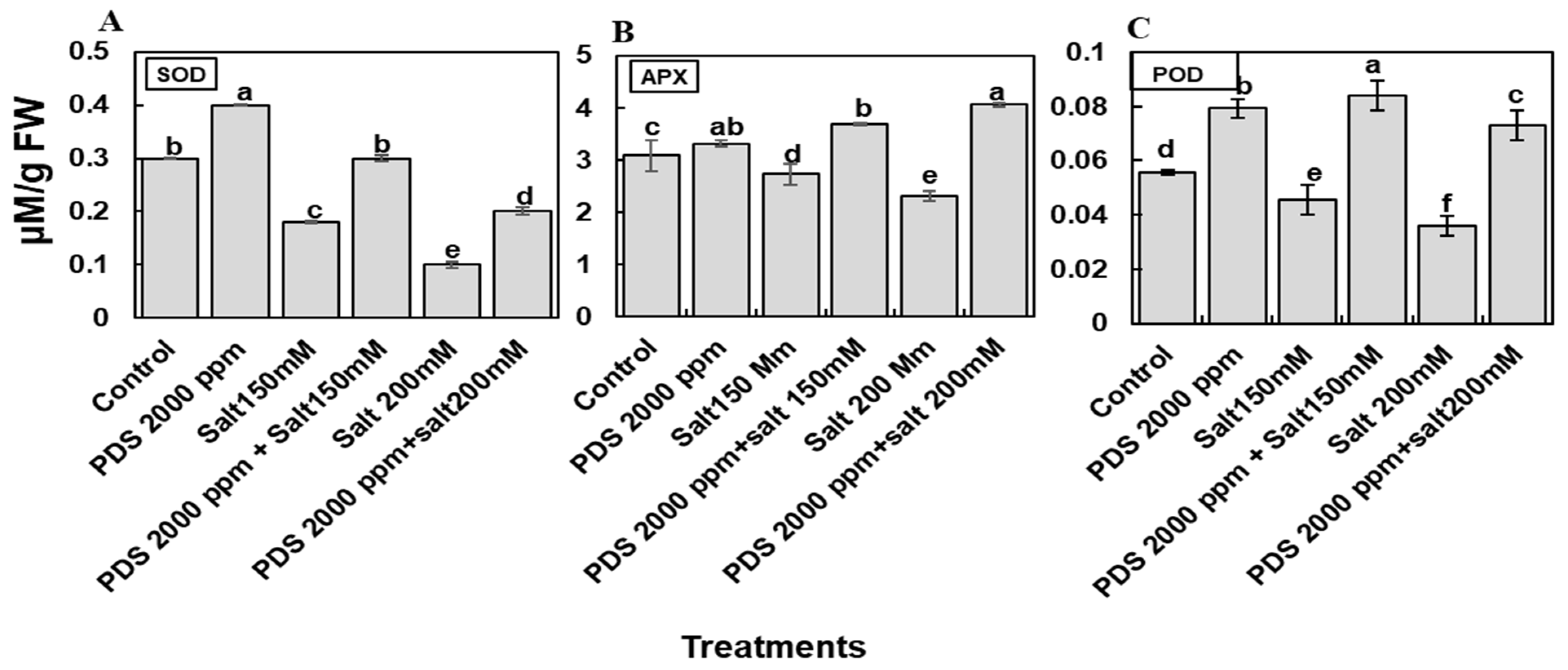
2.6. Plant-Derived Smoke Solution Reduces Oxidative Stress by Enhancing Levels of Antioxidative Enzymes
3. Discussion
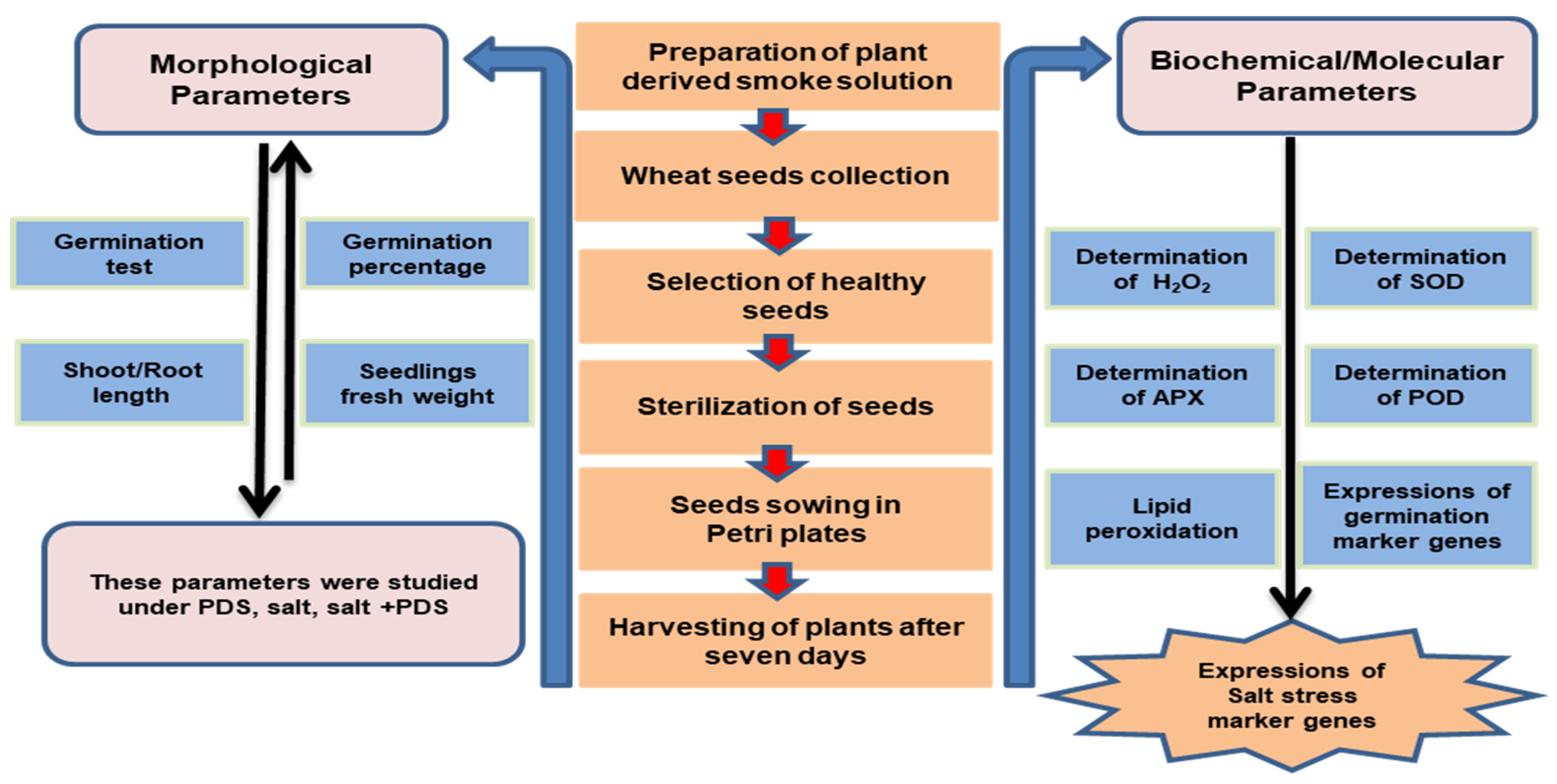
4. Materials and Methods
4.1. Preparation of PDS and Salt Solutions
4.2. Plant Material and Treatment with Plant-Derived Smoke Solution and Salt Stress
4.3. Total RNA Extraction and Analysis of qRT-PCR
4.4. Measurement of H2O2 and Lipid Peroxidation
4.5. Quantification of Antioxidant Enzymes
4.6. Measurement of Superoxide Dismutase (SOD) Activity
4.7. Quantification of Guaiacol Peroxidase (POD) and Ascorbate Peroxidase (APX) Activity
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ganie, S.A.; Reddy, A.S. Stress-Induced Changes in Alternative Splicing Landscape in Rice: Functional Significance of Splice Isoforms in Stress Tolerance. Biology 2021, 10, 309. [Google Scholar] [CrossRef]
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, P.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R.; Mark, T. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xu, W.; Velten, J.; Xin, Z.; Stout, J. Characterization of maize inbred lines for drought and heat tolerance. J. Soil Water Conser. 2012, 67, 354–364. [Google Scholar] [CrossRef] [Green Version]
- Akram, M.S.; Shahid, M.; Tariq, M.; Azeem, M.; Javed, M.T.; Saleem, S.; Riaz, S. Deciphering Staphylococcus sciuri SAT-17 mediated anti-oxidative defense mechanisms and growth modulations in salt stressed maize (Zea mays L.). Front. Microbiol. 2016, 7, 867. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Shannon, M.C. Salinity effects on seedling growth and yield components of rice. Crop Sci. 2000, 40, 996–1003. [Google Scholar] [CrossRef] [Green Version]
- Bayuelo-Jimenez, J.S.; Craig, R.; Lynch, J.P. Salinity tolerance of Phaseolus species during germination and early seedling growth. Crop Sci. 2002, 42, 1584–1594. [Google Scholar] [CrossRef]
- Alam, M.Z.; Stuchbury, T.; Naylor, R.E.; Rashid, M.A. Effect of salinity on growth of some modern rice cultivars. J. Agron. 2004, 3, 223–236. [Google Scholar] [CrossRef]
- Omami, E.N. Effects of salinity stress on amaranth seed germination and seedling growth. N. Z. J. Crop Horticul. Sci. 2005, 34, 35–59. [Google Scholar]
- Jain, N.; Kulkarni, M.G.; Van Staden, J. A butenolide, isolated from smoke, can overcome the detrimental effects of extreme temperatures during tomato seed germination. Plant Growth Regul. 2006, 49, 263–267. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Ganie, S.A.; Molla, K.A.; Henry, R.J.; Bhat, K.V.; Mondal, T.K. Advances in understanding salt tolerance in rice. Theor. Appl. Genet. 2019, 132, 851–870. [Google Scholar] [CrossRef]
- Ganie, S.A.; Mazumder, A.; Kiran, K.; Hossain, F.; Sharma, R.; Mondal, T.K. Transcriptional dynamics of Zn-accumulation in developing kernels of maize reveals important Zn-uptake mechanisms. Genomics 2020, 112, 3435–3447. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Gu, W.; Meng, Y.; Li, J.; Li, L.; Wang, Y.; Qu, D.; Wei, S. Exogenous DCPTA ameliorates simulated drought conditions by improving the growth and photosynthetic capacity of maize seedlings. Nature 2017, 7, 12684. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Li, Y.; Zhu, J.K. Developing naturally stress-resistant crops for a sustainable agriculture. Nat. Plants 2018, 4, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Khan, A.L.; Ali, L.; Hussain, J.; Waqas, M.; Al-Harrasi, A.; Imran, Q.M.; Kim, Y.H.; Kang, S.M.; Yun, B.W.; et al. Hydroquinone; a novel bioactive compound from plant-derived smoke can cue seed germination of lettuce. Front. Chem. 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, T.H.; van Staden, J. Dormancy break of celery (Apium graveolens L.) seeds by plant-derived smoke extract. Plant Growth Regul. 1995, 17, 195–198. [Google Scholar] [CrossRef]
- Doherty, L.C.; Cohn, M.A. Seed dormancy in red rice (Oryza sativa). XI. Commercial liquid smoke elicits germination. Seed Sci. Res. 2000, 10, 415–421. [Google Scholar] [CrossRef]
- Sparg, S.G.; Kulkarni, M.G.; van Staden, J. Aerosol smoke and smoke water stimulation of seedling vigor of a commercial maize cultivar. Crop Sci. 2006, 46, 1336–1340. [Google Scholar] [CrossRef]
- Aremu, A.O.; Bairu, J.F.X.; Finnie, V.; van Staden, J. Stimulatory role of smoke water and karrikinolide on the photosynthetic pigment and phenolic contents of micropropagated ‘Williams’ bananas. Plant Growth Regul. 2012, 67, 271–279. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Amoo, S.O.; Kandari, L.S.; van Staden, J. Seed germination and phytochemical evaluation in seedlings of Aloe arborescens Mill. Plant Biosyst. 2013, 148, 460–466. [Google Scholar] [CrossRef]
- Waheed, M.A.; Jamil, M.; Daud, M.; Shakir, S.; Rehman, S. Effect of plant-derived smoke solutions onphysiological and biochemical attributes of maize (Zea mays L.) under salt stress. Pak. J. Bot. 2016, 48, 1763–1774. [Google Scholar]
- Khatoon, A.; Rehman, S.U.; Aslam, M.M.; Jamil, M.; Komatsu, S. Plant-Derived Smoke Affects Biochemical Mechanism on Plant Growth and Seed Germination. Int. J. Mol. Sci. 2020, 21, 7760. [Google Scholar] [CrossRef]
- Jamil, M.; Kanwal, M.; Aslam, M.M.; Shakir, S.K.; Malook, I.; Tu, J.; Rehman, S. Effect of plant-derived smoke priming on physiological and biochemical characteristics of rice under salt stress condition. Aust. J. Crop. Sci. 2014, 8, 159–170. [Google Scholar]
- Shah, F.A.; Wei, X.; Wang, Q.; Liu, W.; Wang, D.; Yao, Y.; Hu, H.; Chen, X.; Huang, S.; Hou, J.; et al. Karrikin Improves Osmotic and Salt Stress Tolerance via the Regulation of the Redox Homeostasis in the Oil Plant Sapium sebiferum. Front. Plant Sci. 2020, 11, 216. [Google Scholar] [CrossRef]
- Ghebrehiwot, H.M.; Kulkarni, M.G.; Kirkman, K.P.; Staden, J.V. Smoke-water and a smoke-isolated butenolide improve germination and seedling vigour of Eragrostis tef (zucc.) trotter under high temperature and low osmotic potential. J. Agron. Crop Sci. 2008, 194, 270–277. [Google Scholar] [CrossRef]
- Nelson, D.C.; Scaffidi, A.; Dun, E.A.; Waters, M.T.; Flematti, G.R.; Dixon, K.W.; Beveridge, C.A.; Ghisalbert, E.L.; Smith, S.M. F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 8897–8902. [Google Scholar] [CrossRef] [Green Version]
- Sunmonu, T.O.; Kulkarni, M.G.; Staden, J.V. Smoke-water, karrikinolide and gibberellic acid stimulate growth in bean and maize seedlings by efficient starch mobilization and suppression of oxidative stress. S. Afr. J. Bot. 2016, 102, 4–11. [Google Scholar] [CrossRef]
- Shabir, S.; Ilyas, N.; Asif, S.; Iqbal, M.; Kanwal, S.; Ali, Z. Deciphering the role of plantoderived smoke solution in ameliorating saline stress and improving physiological, biochemical and growth responses of wheat. J. Plant Growth Regul. 2021, 10, 1007. [Google Scholar]
- Liu, T.; Hou, G.G.; Cardin, M.; Marquart, L.; Dubat, A. Quality attributes of whole-wheat flour tortillas with sprouted whole-wheat four substitution. LWT 2017, 77, 1–7. [Google Scholar] [CrossRef]
- Smol, J.P. Climate change: A planet in fux. Nature 2012, 483, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.W.; Ren, L.K.; Fan, J.W.; Li, Q.; Wang, K.J.; Guo, M.M.; Chen, F. Salt response of photosynthetic electron transport system in wheat cultivars with contrasting tolerance. Plant Soil Environ. 2016, 62, 515–521. [Google Scholar]
- Shi, H.; Ishitani, M.; Kim, C.; Zhu, J.K. The Arabidopsis thaliana salt tolerance gene SOS1 encodes a putative Na+/H+ antiporter. Proc. Natl. Acad. Sci. USA 2000, 97, 6896–6901. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.M.; Jamil, M.; Khatoon, A.; Hendawy, S.E.; Suhaibani, N.A.; Malook, I.; Rehman, S. Physiological and biochemical responses of maize (Zea mays L.) to plant derived smoke solution. Pak. J. Bot. 2017, 49, 435–443. [Google Scholar]
- Aslam, M.M.; Rehman, S.; Khatoon, A.; Jamil, M.; Yamaguchi, H.; Hitachi, K.; Komatsu, S. Molecular responses of maize shoot to a plant derived smoke solution. Int. J. Mol. Sci. 2019, 20, 1319. [Google Scholar] [CrossRef] [Green Version]
- Rehman, A.; Rehman, S.U.; Khatoon, A.; Qasim, M.; Itoh, T.; Iwasaki, Y.; Wang, X.; Sunohara, Y.; Matsumoto, H.; Komatsu, S. Proteomic analysis of the promotive effect of plant-derived smoke on plant growth of chickpea. J. Proteom. 2018, 176, 56–70. [Google Scholar] [CrossRef]
- Nazish, M.; Zafar, M.; Ahmad, M.; Sultana, S.; Ullah, R.; Alqahtani, A.S.; Ullah, F.; Ahmad, S.; Ashfaq, S.; Ullah, F. Palyno-morphological investigations of halophytic taxa of Amaranthaceae through SEM from Salt range of Northern Punjab, Pakistan. Microsc. Res. Tech. 2019, 82, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Downes, K.S.; Light, M.E.; Posta, M.; Kohout, L.; van Staden, J. Comparison of germination responses of Anigozanthos flavidus (Haemodoraceae), Gyrostemon racemiger and Gyrostemon ramulosus (Gyrostemonaceae) to smoke-water and the smoke-derived compounds karrikinolide (KAR1) and glyceronitrile. Ann. Bot. 2013, 111, 489–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J.L.S.; van Staden, J. Plant-derived smoke solutions stimulate the growth of Lycopersicon esculentum roots in vitro. Plant Growth Regul. 1998, 26, 77–83. [Google Scholar] [CrossRef]
- Van Staden, J.; Sparg, S.G.; Kulkarni, M.G.; Light, M.E. Post germination effects of the smoke-derived compound 3-methyl-2H-furo [2,3-c] pyran-2-one, and its potential as a preconditioning agent. Field Crop Res. 2006, 98, 98–105. [Google Scholar] [CrossRef]
- Gorni, P.H.; de Lima, G.R.; de Oliveira Pereira, L.M.; Spera, K.D.; de Marcos Lapaz, A.; Pacheco, A. Increasing plant performance, fruit production and nutritional value of tomato through foliar applied rutin. Sci. Hortic. 2021, 294, 110755. [Google Scholar] [CrossRef]
- Papenfus, H.B.; Kulkarni, G.; Stirk, W.A.; Rengasamy, K.R.R.; Salomon, M.V.; Piccoli, P.; Bottini, R.; van Staden, J. Interactions between a plant growth-promoting rhizobacterium and smoke-derived compounds and their effect on okra growth. J. Plant Nutr. Soil Sci. 2015, 178, 741–747. [Google Scholar] [CrossRef]
- Meng, Y.; Shuai, H.; Luo, X.; Chen, F.; Zhou, W.; Yang, W.; Shu, K. Karrikins: Regulators involved in phytohormone signaling networks during seed germination and seedling development. Front. Plant Sci. 2017, 7, 2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehnavi, A.R.; Zahedi, M.; Ludwiczak, A.; Perez, S.C.; Piernik, A. Effect of Salinity on Seed Germination and Seedling Development of Sorghum (Sorghum bicolor (L.) Moench) Genotypes. Agronomy 2020, 10, 859. [Google Scholar] [CrossRef]
- Iqbal, M.; Raza, A.; Zulfiqar, S.; Athar, H.R.; Zafar, Z.U. Physiological quality improvement of salt imbibed maize (Zea mays L.) seeds by priming with smoke water. Agrochimical 2013, 56, 247–255. [Google Scholar]
- Wang, M.; Xia, G. The landscape of molecular mechanisms for salt tolerance in wheat. Crop J. 2018, 6, 42–47. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Smith, M.W.; Brown, R.G.; Kamiya, Y.; Sun, T. Phytochrome regulation and differential expression of gibberellin 3β-hydroxylase genes in germinating Arabidopsis seeds. Plant Cell. 1998, 10, 2115–2126. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, K.; Job, C.; Groot, S.P.; Puype, M.; Demol, H.; Vandekerckhove, J.; Job, D. Proteomic analysis of Arabidopsis seed germination and priming. Plant Physiol. 2001, 126, 835–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nawaz, G.; Lee, K.; Park, S.J.; Kim, Y.; Kang, H. A chloroplast-targeted cabbage DEAD-box RNA helicase BrRH22 confers abiotic stress tolerance to transgenic Arabidopsis plants by affecting translation of chloroplast transcripts. Plant Physiol. Biochem. 2018, 127, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, G.; Sai, T.Z.T.; Lee, K.; Kim, Y.; Kang, H. Rice DEAD-box RNA helicase OsRH53 has negative impact on Arabidopsis response to abiotic stresses. J. Plant Growth Regul. 2018, 85, 153–163. [Google Scholar] [CrossRef]
- Ni, J.; Wang, Q.; Shah, F.A.; Liu, W.; Wang, D.; Huang, S.; Fu, S.; Wu, L. Exogenous melatonin confers cadmium tolerance by counterbalancing the hydrogen peroxide homeostasis in wheat seedlings. Molecules 2018, 23, 799. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, S.; Pandey, G.K.; Tuteja, N. Calcium-and salt-stress signaling in plants: Shedding light on SOS pathway. Arch. Biochem. Biophys. 2008, 471, 146–158. [Google Scholar] [CrossRef]
- Yinglu, S.; Liu, X.; Fu, L.; Qin, P.; Li, T.; Ma, X.; Wang, X. Overexpression of TaBADH increases salt tolerance in Arabidopsis. Can. J. Plant Sci. 2019, 99, 546–555. [Google Scholar]
- Shi, H.; Zhu, J.K. SOS4, A Pyridoxal Kinase Gene, Is Required for Root Hair Development in Arabidopsis1. Plant Physiol. 2002, 129, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K.; Liu, J. An Arabidopsis mutant that requires increased calcium for potassium nutrition and salt tolerance. Proc. Natl. Acad. Sci. USA 1997, 94, 14960–14964. [Google Scholar]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidat. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Nita, M.; Grzybowski, A. The role of the reactive oxygen species and oxidative stress in the pathomechanism of the age-related ocular diseases and other pathologies of the anterior and posterior eye segments in adults. Oxidat. Med. Cell. Longev. 2016, 2016, 3164734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asada, K. The water-water cycle as alternative photon and electron sinks. Philos. Trans. R. Soc. Lond. 2000, 355, 1419–1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, F.R.; Lima, J.P.; Ferreira-Silva, S.L.; Viegas, R.A.; Silveira, J.A. Roots and leaves display contrasting oxidative response during salt stress and recovery in cowpea. Plant Physiol. 2007, 164, 591–600. [Google Scholar] [CrossRef]
- Mullineaux, P.M.; Karpinski, S.; Jiménez, A.; Cleary, S.P.; Robinson, C.; Creissen, G.P. Identification of cDNAS encoding plastid-targeted glutathione peroxidase. Plant J. 2010, 13, 375–379. [Google Scholar] [CrossRef]
- Chen, S.; Heuer, B. Effect of genotype and exogenous application of glycinebetaine on antioxidant enzyme activity in native gels of 7-day-old saltstressed tomato (Solanum lycopersicum) seedlings. Sci. Horticult. 2013, 162, 106–116. [Google Scholar] [CrossRef]
- Del Río, L.A.; Corpas, F.J.; López-Huertas, E.; Palma, J.M. Plant superoxide dismutases: Function under abiotic stress conditions. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer: Cham, Switzerland, 2018; pp. 1–26. [Google Scholar]
- Tieu, A.; Dixon, K.W.; Sivasithamparam, K.; Plummer, J.A.; Sieler, I.M. Germination of four species of native western Australian plant using plant-derived smoke. Aust. J. Bot. 1999, 47, 207–219. [Google Scholar] [CrossRef]
- Kim, J.S.; Park, S.J.; Kwak, K.J.; Kim, Y.O.; Kim, J.Y.; Song, J.; Jang, B.; Jung, C.H.; Kang, H. Cold shock domain proteins and glycine-rich RNA-binding protein from Arabidopsis thaliana can promote the cold adaptation process in Escherichia coli. Nucleic Acids Res. 2007, 35, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Le, T.N.N.; Kang, H. Artificial targeting of a nucleus-encoded RNA-binding protein AtRZ1a to chloroplasts affects flowering and ABA response of Arabidopsis thaliana. J. Plant Biol. 2017, 60, 278–284. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van Montagu, M.; Van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hayat, N.; Afroz, N.; Rehman, S.; Bukhari, S.H.; Iqbal, K.; Khatoon, A.; Taimur, N.; Sakhi, S.; Ahmad, N.; Ullah, R.; et al. Plant-Derived Smoke Ameliorates Salt Stress in Wheat by Enhancing Expressions of Stress-Responsive Genes and Antioxidant Enzymatic Activity. Agronomy 2022, 12, 28. https://doi.org/10.3390/agronomy12010028
Hayat N, Afroz N, Rehman S, Bukhari SH, Iqbal K, Khatoon A, Taimur N, Sakhi S, Ahmad N, Ullah R, et al. Plant-Derived Smoke Ameliorates Salt Stress in Wheat by Enhancing Expressions of Stress-Responsive Genes and Antioxidant Enzymatic Activity. Agronomy. 2022; 12(1):28. https://doi.org/10.3390/agronomy12010028
Chicago/Turabian StyleHayat, Nailla, Neelum Afroz, Shafiq Rehman, Syeda Huma Bukhari, Khalid Iqbal, Amana Khatoon, Nadia Taimur, Shazia Sakhi, Nisar Ahmad, Riaz Ullah, and et al. 2022. "Plant-Derived Smoke Ameliorates Salt Stress in Wheat by Enhancing Expressions of Stress-Responsive Genes and Antioxidant Enzymatic Activity" Agronomy 12, no. 1: 28. https://doi.org/10.3390/agronomy12010028
APA StyleHayat, N., Afroz, N., Rehman, S., Bukhari, S. H., Iqbal, K., Khatoon, A., Taimur, N., Sakhi, S., Ahmad, N., Ullah, R., Ali, E. A., Bari, A., Hussain, H., & Nawaz, G. (2022). Plant-Derived Smoke Ameliorates Salt Stress in Wheat by Enhancing Expressions of Stress-Responsive Genes and Antioxidant Enzymatic Activity. Agronomy, 12(1), 28. https://doi.org/10.3390/agronomy12010028