
Pre-Harvest Fruit Splitting of Citrus


Abstract
:1. What Is Pre-Harvest Fruit Splitting?
2. Affected Cultivars
3. Underlying Mechanisms
4. Initiation
5. Symptom Expression
5.1. Fruit Cell Division (Stage I)
5.2. Cell Expansion Stage (Stage II)
5.3. Cell Maturation Stage (Stage III)
6. Factors Affecting Severity
6.1. Crop Load
6.2. Fruit Shape
6.3. Fluctuating Water Status
6.4. Humidity and Light
6.5. Peel Quality
7. What Steps Can Be Taken to Mitigate the Effects of PFS?
7.1. Mineral Nutrients
7.1.1. Boron
7.1.2. Calcium
7.1.3. Nitrogen
7.1.4. Phosphorus
7.1.5. Potassium
7.1.6. Zinc
7.1.7. Ending Remarks
7.2. Water (Rainfall, Irrigation, Flood, or Drought)
7.3. Regulation of Crop Load
7.4. Plant Growth Regulators
7.4.1. Gibberellic Acid (i.e., Gibberellins, GA, GA3, or A3)
7.4.2. 2,4-Dichlorophenoxyacetic Acid (2,4-D)
7.4.3. Other Plant Growth Regulators
7.4.4. Combinations of Plant Growth Regulators
7.5. Pesticides
7.6. Canopy Position
8. Summary and Possible Future Directions for Research
9. Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Erickson, L. The general physiology of citrus. In The Citrus Industry Vol. II; Reuther, W., Batchelor, L., Webber, H., Eds.; University of California: Berkeley, CA, USA, 1968; pp. 116–117. [Google Scholar]
- Coit, J. Citrus Fruits; MacMillan: New York, NY, USA, 1925. [Google Scholar]
- Cronjé, P.J.R.; Stander, O.P.J.; Theron, K.I. Fruit splitting in citrus. Hortic. Rev. 2013, 41, 177–200. [Google Scholar]
- Iglesias, D.J.; Cercós, M.; Colmenero-Flores, J.M.; Naranjo, M.A.; Ríos, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; Tadeo, F.R.; et al. Physiology of citrus fruiting. Braz. J. Plant Physiol. 2007, 19, 333–362. [Google Scholar] [CrossRef]
- Li, J.; Chen, J. Citrus fruit-cracking: Causes and occurrence. Hortic. Plant J. 2017, 3, 255–260. [Google Scholar] [CrossRef]
- Coetzee, J.; Land, G.; Gilfillan, I.; Bower, J.P.; Holmden, E.; Chamberlain, J.; Krajewski, A. Production Guidelines for Export Citrus. Vol II. Citriculture—Cultural Practices; Outspan International Ltd.: Paarl, South Africa, 1996. [Google Scholar]
- Agusti, M.; Martínez-Fuentes, A.; Mesejo, C. Citrus fruit quality. Physiological basis and techniques of improvement. Agrociencia 2002, VI, 1–16. [Google Scholar]
- Zur, N.; Shlizerman, L.; Ben-Ari, G.; Sadka, A. Use of Magnetic Resonance Imaging (MRI) to Study and Predict Fruit Splitting in Citrus. Hortic. J. 2017, 86, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Opara, L.U.; Studman, C.J.; Banks, N.H. Fruit skin splitting and cracking. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1996; Volume 19, pp. 217–262. [Google Scholar]
- Kaur, R.; Kaur, N.; Singh, H. Pericarp and pedicel anatomy in relation to fruit cracking in lemon (Citrus limon L. Burm.). Sci. Hortic. 2019, 246, 462–468. [Google Scholar] [CrossRef]
- Sdoodee, S.; Chiarawipa, R. Fruit splitting occurrence of Shogun mandarin (Citrus reticulata Blanco cv. Shogun) in southern Thailand and alleviation by calcium and boron sprays. Songklanakarin J. Sci. Technol. 2005, 27, 719–730. [Google Scholar]
- Yara. Preventing Peel Plugging and Creasing. Available online: https://www.yara.us/crop-nutrition/citrus/preventing-peel-plugging-and-creasing/ (accessed on 19 June 2022).
- Garcia-Luis, A.; Duarte, A.M.M.; Kanduser, M.; Guardiola, J.L. The anatomy of the fruit in relation to the propensity of citrus species to split. Sci. Hortic. 2001, 87, 33–52. [Google Scholar] [CrossRef] [Green Version]
- Wen, M.-X.; Shi, X.-J. Influence of calcium on fruit cracking of Jincheng orange and its physiological mechanism. Sci. Agric. Sin. 2012, 45, 1127–1134. [Google Scholar] [CrossRef]
- Kumar, A.; Chahal, T.S. Studies on diseases and disorders of different citrus germplasm under natural conditions. J. Entomol. Zool. Stud. 2018, 6, 406–409. [Google Scholar]
- Mesejo, C.; Reig, C.; Martinez-Fuentes, A.; Gambetta, G.; Gravina, A.; Agusti, M. Tree water status influences fruit splitting in Citrus. Sci. Hortic. 2016, 209, 96–104. [Google Scholar] [CrossRef]
- De Cicco, V.; Intrigliolo, F.; Ippolito, A.; Vanadia, S.; Guiffrida, A. Factors in Navelina orange splitting. In Proceedings of the 6th International Citrus Congress, Tel Aviv, Israel, 6–11 March 1988; pp. 535–540. [Google Scholar]
- Zhou, J.; Xiang, B.; Lan, W.; Wang, X.; Hu, X. Effects of fertilizers with different proportions of N, P and K on citrus fruit cracking. Agric. Sci. Technol. 2014, 15, 665–667. [Google Scholar]
- Habibi, S.; Ebadi, A.; Ladanmoghadam, A.R.; Rayatpanah, S. Effect of Plant Growth Regulators on Fruit Splinting in Thompson Navel Orange. Acta Sci. Pol. Hortorum Cultus 2021, 20, 83–92. [Google Scholar] [CrossRef]
- Omar, A.; El-Enin, M.S.A. Foliar spray with different agrochemicals on fruit quality and exportability of ‘Washington’ navel orange fruit (Citrus sinensis L.). Int. J. Fruit Sci. 2017, 17, 280–295. [Google Scholar] [CrossRef]
- El-Tanany, M.M.; Messih, M.N.A.; Shama, M.A. Effect of foliar application with potassium, calcium, and magnesium on yield, fruit quality and mineral composition of Washington navel orange trees. Alex. Sci. Exch. J. 2017, 32, 65–75. [Google Scholar]
- El Rahman, A.; Hoda, G.F.; Mohamed, M.; Tayh, E.A.H. Effect of GA3, and potassium nitrate in different dates on fruit set, yield and splitting of Washington navel orange. Nat. Sci. 2012, 10, 148–157. [Google Scholar]
- El-Sayed, S.A. Some factors affecting orange fruit splitting of Washington navel orange under Kaferelshikh conditions. B- The effect of climate conditions and fruit position on the tree canopy. J. Plant Prod. Mansoura Univ. 2016, 7, 339–342. [Google Scholar]
- Wagner, V.A. The navel-end-rot, splitting, and large-end problems of Washington navel oranges in the Kat River Valley. Plant Ind. Ser. Sci. Bull. 1938, 192, 5–20. [Google Scholar]
- Rabe, E.; van Rensburg, P.; van der Walt, H.; Bower, J. Factors influencing preharvest fruit splitting in Ellendale (C. reticulata). HortScience 1990, 25, 1135. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Sha, C.; Shu, Q. Integrated Physiological and Metabolomic Analyses of the Effect of Potassium Fertilizer on Citrus Fruit Splitting. Plants 2022, 11, 499. [Google Scholar] [CrossRef]
- Barry, G.H.; Bower, J.P. Manipulation of fruit set and stylar-end fruit split in ‘Nova’ mandarin hybrid. Sci. Hortic. 1997, 70, 243–250. [Google Scholar] [CrossRef]
- Greenberg, J.; Kaplan, I.; Fainzack, M.; Egozi, Y.; Giladi, B. Effects of auxin sprays on yield, fruit size, fruit splitting and the incidence of creasing of ‘Nova’ mandarin. Acta Hortic. 2006, 727, 249–254. [Google Scholar] [CrossRef]
- García-Luis, A.; Duarte, A.M.M.; Porras, I.; Garcialidon, A.; Guardiola, J.L. Fruit splitting in ‘Nova’ hybrid mandarine in relation to the anatomy of the fruit and fruit-set treatments. Sci. Hortic. 1994, 57, 215–231. [Google Scholar] [CrossRef]
- Almela, V.; Zaragoza, S.; Primo-Millo, E.; Agusti, M. Hormonal control of splitting in ‘Nova’ mandarin fruit. J. Hortic. Sci. 1994, 69, 969–973. [Google Scholar]
- Alikhani, M.; Babakhani, B.; Golein, B.; Asadi, M.; Rahdari, P. Foliar application of potassium nitrate and 2,4-dichlorophenoxyacetic acid affect some fruit splitting related characteristics and biochemical traits of mandarin cv. ‘page’. EurAsian J. BioSciences 2020, 14, 4251–4260. [Google Scholar]
- Hardiyanto; Devy, F.N. Application of K, Ca, and Mg on peel thickness and fruit cracking incidence of citrus. Russ. J. Agric. Socio-Econ. Sci. 2019, 87, 45–56. [Google Scholar] [CrossRef]
- Savreet, S.; Bal, J.S. Quality improvement in lemon (Citrus limon (L.) Burm.) through integrated management of fruit cracking. Afr. J. Agric. Res. 2013, 8, 3552–3557. [Google Scholar] [CrossRef] [Green Version]
- Khehra, S.; Bal, J.S. Influence of foliar sprays on fruit cracking in lemon. Int. J. Plant Anim. Environ. Sci. 2014, 4, 124–128. [Google Scholar]
- Sandhu, S.; Bal, J.S. Response of lemon cv. Baramasi to foliar feeding of nutrients. Indian J. Hortic. 2012, 69, 281–283. [Google Scholar]
- Josan, J.S.; Sandhu, A.S.; Singh, R.; Dhillon, D.S. Effect of plant growth regulators and nutrients on fruit cracking in lemon. Indian J. Hortic. 1995, 52, 121–124. [Google Scholar]
- Josan, J.S.; Sandhu, A.S. Studies on the causes and periodical incidence of fruit cracking in lemon (Citrus limon (L.) Burm). Haryana J. Hortic. Sci. 1999, 28, 158–160. [Google Scholar]
- Sandhu, S. Improving lemon [Citrus limon (L.) Burm.] quality using growth regulators. J. Hortic. Sci. 2013, 8, 88–90. [Google Scholar]
- Randhawa, G.S.; Singh, J.P.; Malik, R.S. Fruit cracking in some tree fruits with special reference to lemon (Citrus limon). Indian J. Hortic. 1958, 15, 6–9. [Google Scholar]
- Devi, K.; Kumar, R.; Wali, V.K.; Bakshi, P.; Sharma, N.; Arya, V.M. Effect of foliar nutrition and growth regulators on nutrient status and fruit quality of Eureka lemon (Citrus limon). Indian J. Agric. Sci. 2018, 88, 704–708. [Google Scholar]
- Sharma, R.R.; Saxena, S.K.; Goswami, A.M.; Shukla, A.K. Effect of foliar application of calcium chloride on fruit cracking, yield and quality of Kagzi Kalan lemon. Indian J. Hortic. 2002, 59, 145–149. [Google Scholar]
- Bhatt, B.B.; Rawat, S.S.; Naithani, D.C.; Kumar, D.; Singh, K.K. Effect of foliar application of bio-regulators and nutrients on growth and yield characters of lemon (Citrus limon Burma.) c.v. plant lemon-1 under subtropical condition of Garhwal region. Plant Arch. 2016, 16, 821–825. [Google Scholar]
- Hardy, S.; Barkley, P.; Treeby, M.T.; Smith, M.; Sanderson, G. Australian Mandarin Production Manual; New South Wales Department of Primary Industries: Orange, NSW, Australia, 2017. [Google Scholar]
- Stander, O.P.J. Fruit Split and Fruit Size Studies on Citrus; Stellenbosch University: Stellenbosch, South African, 2013. [Google Scholar]
- Rodríguez-Gamir, J.; Intrigliolo, D.S.; Primo-Millo, E.; Forner-Giner, M.A. Relationships between xylem anatomy, root hydraulic conductivity, leaf/root ratio and transpiration in citrus trees on different rootstocks. Physiol. Plant. 2010, 139, 159–169. [Google Scholar] [CrossRef]
- Vasconcellos, L.A.B.C.; Castle, W.S. Trunk xylem anatomy of mature healthy and blighted grapefruit trees on several rootstocks. J. Am. Soc. Hortic. Sci. 1994, 119, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Bain, J.M. Morphological, anatomical, and physiological changes in the developing fruit of the Valencia orange, Citrus sinensis (L.) Osbeck. Aust. J. Bot. 1958, 6, 1–24. [Google Scholar] [CrossRef]
- Stander, O.P.J. An investigation of excessive fruit drop in the easter and western cape during the 2016/2017 season. S. Afr. Fruit J. 2017, 16, 102–107. [Google Scholar]
- Stander, O.P.J.; Theron, K.; Cronjé, P.J.R. Foliar 2,4-D application after physiological fruit drop reduces fruit splitting of mandarin. HortTechnology 2014, 24, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Lima, J.E.O.; Davies, F.S. Secondary-fruit ontogeny in navel orange. Am. J. Bot. 1984, 71, 532–541. [Google Scholar] [CrossRef]
- Cook, A. California Citrus Culture; California State Printing Office: Sacramento, CA, USA, 1913. [Google Scholar]
- Considine, J.; Brown, K. Physical aspects of fruit growth: Theoretical analysis of distribution of surface growth forces in fruit in relation to cracking and splitting. Plant Physiol. 1981, 68, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Rabe, E.; Van Rensburg, P.J.J. Gibberellic acid sprays, girdling, flower thinning and potassium applications affect fruit splitting and yield in the ‘Ellendale’ tangor. J. Hortic. Sci. 1996, 71, 195–203. [Google Scholar] [CrossRef]
- Falivene, S. High-Density Planting and Pruning Case Study: Sunmar Orchards, Sunraysia. Available online: https://www.dpi.nsw.gov.au/agriculture/horticulture/citrus/content/crop-management/orchard-management-factsheets/high-density-planting-and-pruning-case-study-sunmar-orchards,-sunraysia (accessed on 13 June 2022).
- Reuther, W. Climate and citrus behaviour. In The Citrus Industry Vol III; Reuther, W., Batchelor, L.D., Webber, H.J., Eds.; University of California: Riverside, CA, USA, 1973; pp. 280–337. [Google Scholar]
- Bilal, H.M.; Zulfiqar, R.; Adan, M.; Umer, M.S.; Islam, H.; Zaheer, H.; Abbas, W.M.; Haider, F.; Ahmad, R.I. Impact of salinity on citrus production; A review. Int. J. Appl. Res. 2020, 6, 173–176. [Google Scholar]
- Monselise, S.P.; Goldschmidt, E.E. Alternate bearing in fruit trees. Hortic. Rev. 1982, 4, 128–173. [Google Scholar]
- Moss, G.I.; Bevington, K.B. Methods for controlling biennial bearing in “Late Valencia” sweet orange. In Proceedings of the Congreso Mundial de Citricultura, Murcia, Spain, 29 April–10 May 1973; pp. 367–372. [Google Scholar]
- Anonymous. Fruit Size Management Guide, Part 1. Available online: https://www.dpi.nsw.gov.au/__data/assets/pdf_file/0005/138830/Fruit-Size-Guide-PART-1.pdf (accessed on 13 June 2022).
- Anonymous. Fruit Size Management Guide, Part II. Available online: https://www.dpi.nsw.gov.au/__data/assets/pdf_file/0006/138831/Fruit-Size-Guide-PART-2.pdf (accessed on 13 June 2022).
- Embleton, T.W.; Jones, W.W.; Labanauskas, C.K.; Reuther, W. Leaf analysis as a diagnostic tool and guide to fertilization. In The Citrus Industry, Vol. III: Production Technology; Reuther, W., Ed.; University of California: Berkeley, CA, USA, 1973; pp. 183–210. [Google Scholar]
- Smith, P. Citrus nutrition. In Temperate to Tropical Fruit Nutrition; Childers, N., Ed.; Rutgers State University: Rutgers, NJ, USA, 1968; pp. 174–207. [Google Scholar]
- Jordan, L.; Day, B. Weed Control in Citrus; University of California: Berkeley, CA, USA, 1973; Volume 3. [Google Scholar]
- Brown, K.; Considine, J. Physical aspects of fruit growth: Stress distribution around lenticels. Plant Physiol. 1982, 69, 585–590. [Google Scholar] [CrossRef]
- Odemis, B.; Turhan, S.; Buyuktas, D. The effects of irrigation and fertilizer applications on yield, pomological characteristics and fruit cracking in Nova mandarin. Agric. Water Manag. 2014, 135, 54–60. [Google Scholar] [CrossRef]
- Scott, F.M.; Baker, K. Anatomy of Washington navel orange rind in relation to water spot. Bot. Gaz. 1947, 108, 459–475. [Google Scholar] [CrossRef]
- Matas, A.J.; Agusti, J.; Tadeo, F.R.; Talon, M.; Rose, J.K. Tissue-specific transcriptome profiling of the citrus fruit epidermis and subepidermis using laser capture microdissection. J. Exp. Bot. 2010, 61, 3321–3330. [Google Scholar] [CrossRef]
- Blevins, D.G.; Lukaszewski, K.M. Boron in plant structure and function. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 481–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wimmer, M.A.; Goldberg, S.; Gupta, U.C. Boron. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press: New York, NY, USA, 2015; pp. 305–346. [Google Scholar]
- Hanger, B.C. Movement of calcium in plants. Commun. Soil Sci. Plant Anal. 1979, 10, 171–193. [Google Scholar] [CrossRef]
- Rabe, E. Yield benefits associated with pre-bloom low-biuret urea sprays on Citrus spp. J. Hortic. Sci. 1994, 69, 495–500. [Google Scholar] [CrossRef]
- Quaggio, J.A.; Mattos, D.; Cantarella, H.; Almeida, E.L.E.; Cardoso, S.A.B. Lemon yield and fruit quality affected by NPK fertilization. Sci. Hortic. 2002, 96, 151–162. [Google Scholar] [CrossRef]
- Koo, R.C.J. Potassium nutrition and fruit splitting in ‘Hamlin’ orange. Annu. Res. Rep. Fla. Agric. Exp. Stn. 1961, 2, 223–224. [Google Scholar]
- Khadivi-Khub, A. Physiological and genetic factors influencing fruit cracking. Acta Physiol. Plant. 2015, 37, 1718. [Google Scholar] [CrossRef]
- Lewis, L.N.; McCarty, C.D. Pruning and girdling of citrus. In The Citrus Industry, Vol. III: Production Technology; Reuther, W., Ed.; University of California: Berkeley, CA, USA, 1973; pp. 211–229. [Google Scholar]
- Krezdorn, A.H.; Wiltbank, W.J. Annual girdling of ‘Orlando’ tangelos over an eight-year period. Annu. Meet. Fla. State Hortic. Soc. 1968, 3142, 29–35. [Google Scholar]
- Cohen, A. Recent developments in girdling of citrus trees. Proc. Int. Soc. Citric. 1981, 1, 196–199. [Google Scholar]
- Lange, J.H.; Skarup, O.; Vincent, A.P. The influence of cross-pollination and girdling on fruit set and seed content of citrus ‘Ortanique’. Sci. Hortic. 1974, 2, 285–292. [Google Scholar] [CrossRef]
- Morris, G. Improving Citrus Quality Using Gibberellic Acid. Available online: https://www.agric.wa.gov.au/citrus/improving-citrus-quality-using-gibberellic-acid?page=0%2C0 (accessed on 30 March 2021).
- Ladaniya, M.S. Physiological disorders and their management. In Citrus Fruit Biology, Technology, and Evaluation; Elsevier: Boston, MA, USA, 2008; pp. 451–463. [Google Scholar]
- Jona, R.; Goren, R.; Marmora, M. Effect of gibberellin on cell-wall components of creasing peel in mature ‘Valencia’ orange. Sci. Hortic. 1989, 39, 105–115. [Google Scholar] [CrossRef]
- Bower, J.P.; Gilfillan, J.M.; Skinner, H. Fruit Splitting in “Valencia” and its relationship to the pectin status of the rind. In Proceedings of the International Society of Citriculture, Acireale, Italy, 8–13 March 1992; pp. 511–514. [Google Scholar]
- Mupambi, G.; Verreynne, J.S.; Stander, O.P.J.; Cronje, P.J.R. Optimal timing of application of 2,4-D on ‘Navel’ sweet orange Citrus sinensis (Osbeck) reduces the size of the navel-end. J. Hortic. Sci. Biotechnol. 2015, 90, 619–625. [Google Scholar] [CrossRef]
- Mesejo, C.; Martínez-Fuentes, A.; Juan, M.; Almela, V.; Agustí, M. Vascular tissues development of citrus fruit peduncle is promoted by synthetic auxins. Plant Growth Regul. 2003, 39, 131–135. [Google Scholar] [CrossRef]
- Anthony, M.F.; Coggins, C.W., Jr. The efficacy of five forms of 2,4-D in controlling preharvest fruit drop in citrus. Sci. Hortic. 1999, 81, 267–277. [Google Scholar] [CrossRef]
- Beck, N.G.; Arpaia, M.L.; Eckard, K.J.; Reints, J.S.; Lord, E.M. The effect of chlorpyrifos on flower and fruit development in grapefruit, Citrus paradisi MacFayden. Sci. Hortic. 1991, 47, 35–50. [Google Scholar] [CrossRef]
- Sinclair, W.B.; Lindgren, D.L. Ridges and sectors induced in the rind of citrus fruits by fumigation with hydrocyanic acid. Plant Physiol. 1943, 18, 99–106. [Google Scholar] [CrossRef]





{kind=link}
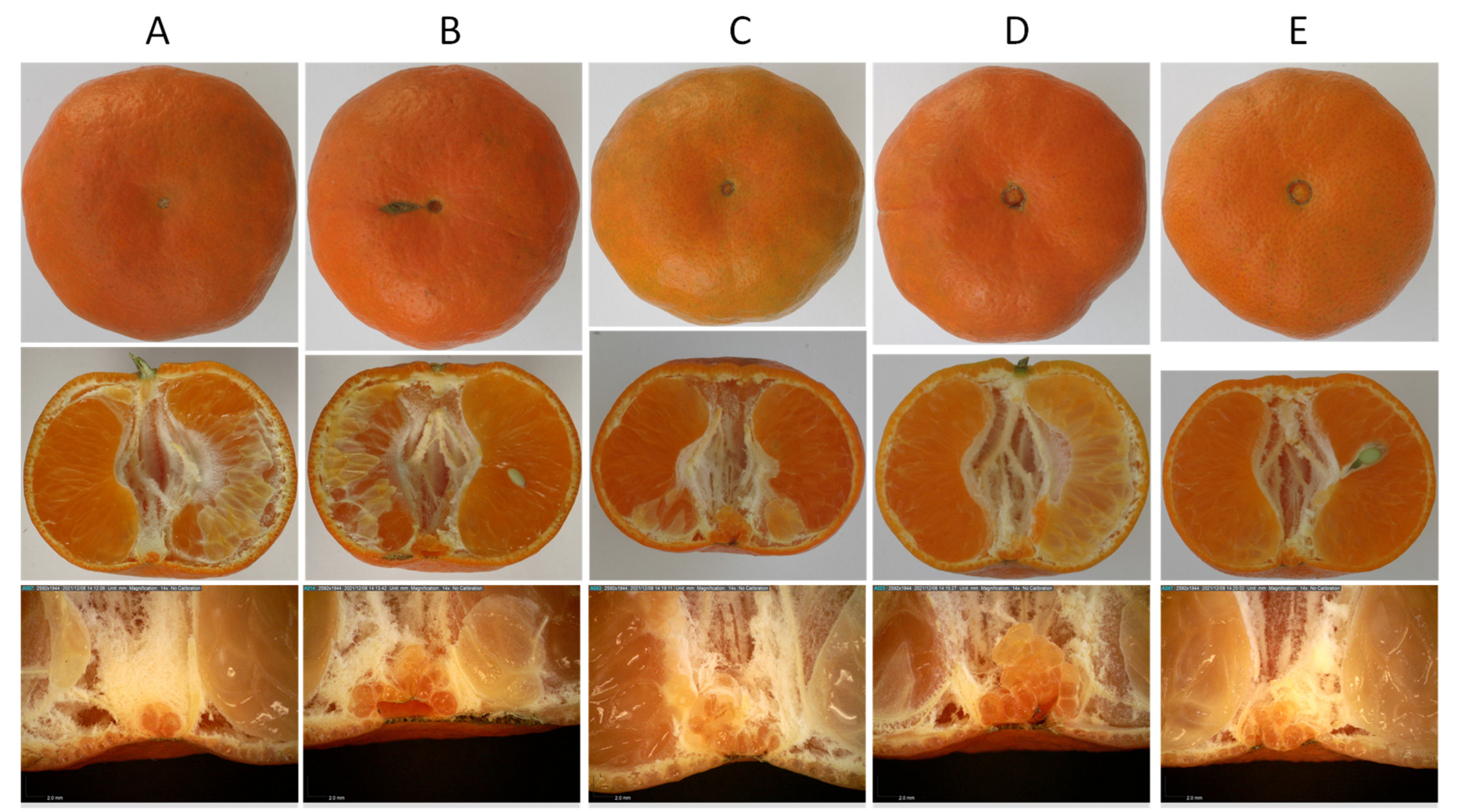{kind=link}
{kind=link}
{kind=link}
| Type | Scion | Rootstock | Percentage | Citation |
|---|---|---|---|---|
| Orange | Beibei 447 Jincheng | 7–16 | [14] | |
| Early Gold | 0 | [15] | ||
| Hamlin | 0 | [15] | ||
| Itaborai | 10 | [15] | ||
| Midknight | 20 | [15] | ||
| Navel | Cleopatra mandarin | 3 | [16] | |
| Navel | FA-5 | 4 | [16] | |
| Navel | C-35 | 9 | [16] | |
| Navel | Carrizo citrange | 8 | [16] | |
| Navelina | <1–30 | [17] | ||
| NewHall | 27.5 | [18] | ||
| Olinda | 0 | [15] | ||
| Rhode Red | 0 | [15] | ||
| Ruby Nucellar | 0 | [15] | ||
| Thompson Navel | C. aurantium | 35–45 | [19] | |
| Trovita | 0 | [15] | ||
| Valencia | 3–30 | [6] | ||
| Vernia | 0 | [15] | ||
| Washington | 5.7–6.6 | [20] | ||
| Washington | Sour orange | 4.0–9.8 | [21] | |
| Washington | Sour orange | 7 | [22] | |
| Washington | Sour orange | 24–26 | [23] | |
| Washington | C. volkameriana | 18–20 | [23] | |
| Washington | 3.4–5.2 | [24] | ||
| Westin | 10 | [15] | ||
| Mandarin | Clementina de Nules | 0 | [13] | |
| Clementine | Poncirus trifoliata | 4 | [16] | |
| Clementine | Carrizo citrange | 11 | [16] | |
| Daisy | 10 | [15] | ||
| Ellendale | Carrizo citrange | 14–30 | [25] | |
| Ellendale | 0 | [13] | ||
| Ehime Kashi 34 | 31 | [26] | ||
| Fino | 0 | [13] | ||
| Kinnow | 0 | [15] | ||
| Nova | C. volkameriana | 30–55 | [8] | |
| Nova | FA-5 | 5 | [16] | |
| Nova | Carrizo citrange | 17 | [16] | |
| Nova | Carrizo citrange | 1.5–27 | [16] | |
| Nova | Poncirus trifoliata | 1.2–7.7 | [16] | |
| Nova | Troyer citrange | 0.10–26 | [27] | |
| Nova | Sour orange | 35 | [28] | |
| Nova | Cleopatra mandarin | 10–40 | [29] | |
| Nova | Troyer citrange | 5–30 | [29] | |
| Nova | 16–18 | [13] | ||
| Nova | 9 | [30] | ||
| Orogrande | 0 | [13] | ||
| Page | 33–50 | [31] | ||
| Pontianak | 0 | [32] | ||
| Shogun | 52.2 | [11] | ||
| Terigas | 6.8–8.8 | [32] | ||
| W. Murcott | 2 | [15] | ||
| Lemon | Baramasi | 31.9–36 | [33] | |
| Baramasi | 31.9–34.1 | [34] | ||
| Baramasi | 33 | [35] | ||
| Baramasi | 33.5–34.0 | [36] | ||
| Baramasi | 26–34 | [37] | ||
| Baramasi | 37.6 | [38] | ||
| Baramasi | 0 | [15] | ||
| Columbia | 20 | [39] | ||
| Eureka | 0 | [15] | ||
| Eureka | 25.98 | [40] | ||
| Eureka | 22.5–35.0 | [39] | ||
| Genoa | 3.3–10.5 | [39] | ||
| Improved Meyer | 0 | [15] | ||
| Italian | 27.7–35.8 | [39] | ||
| Kagzi Kalan | 47.7–48.9 | [41] | ||
| Lisbon | 0 | [15] | ||
| Malta | 15.8–26.8 | [39] | ||
| Nepali oblong | 5.0–20.0 | [39] | ||
| Plant Lemon 1 | 13.2 | [42] | ||
| Punjab galgal | 0 | [15] | ||
| Seedless | 25.0 | [39] | ||
| Grapefruit | Flame | 0 | [15] | |
| Marsh Seedless | 0 | [15] | ||
| Rio Red | 0 | [15] | ||
| Red Blush | 0 | [15] | ||
| Star Ruby | 0 | [15] | ||
| Lime | Bearss | 0 | [15] | |
| Local | 0 | [15] | ||
| Mexican | 0 | [15] | ||
| Kumquat | Fortunella spp. | 0 | [15] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krajewski, A.; Ebert, T.; Schumann, A.; Waldo, L. Pre-Harvest Fruit Splitting of Citrus. Agronomy 2022, 12, 1505. https://doi.org/10.3390/agronomy12071505
Krajewski A, Ebert T, Schumann A, Waldo L. Pre-Harvest Fruit Splitting of Citrus. Agronomy. 2022; 12(7):1505. https://doi.org/10.3390/agronomy12071505
Chicago/Turabian StyleKrajewski, Andrew, Timothy Ebert, Arnold Schumann, and Laura Waldo. 2022. "Pre-Harvest Fruit Splitting of Citrus" Agronomy 12, no. 7: 1505. https://doi.org/10.3390/agronomy12071505
APA StyleKrajewski, A., Ebert, T., Schumann, A., & Waldo, L. (2022). Pre-Harvest Fruit Splitting of Citrus. Agronomy, 12(7), 1505. https://doi.org/10.3390/agronomy12071505