
Effects of Microbial Fertilizer on Soil Fertility and Alfalfa Rhizosphere Microbiota in Alpine Grassland

Abstract
:1. Introduction
2. Methods and Materials
2.1. Experimental Material
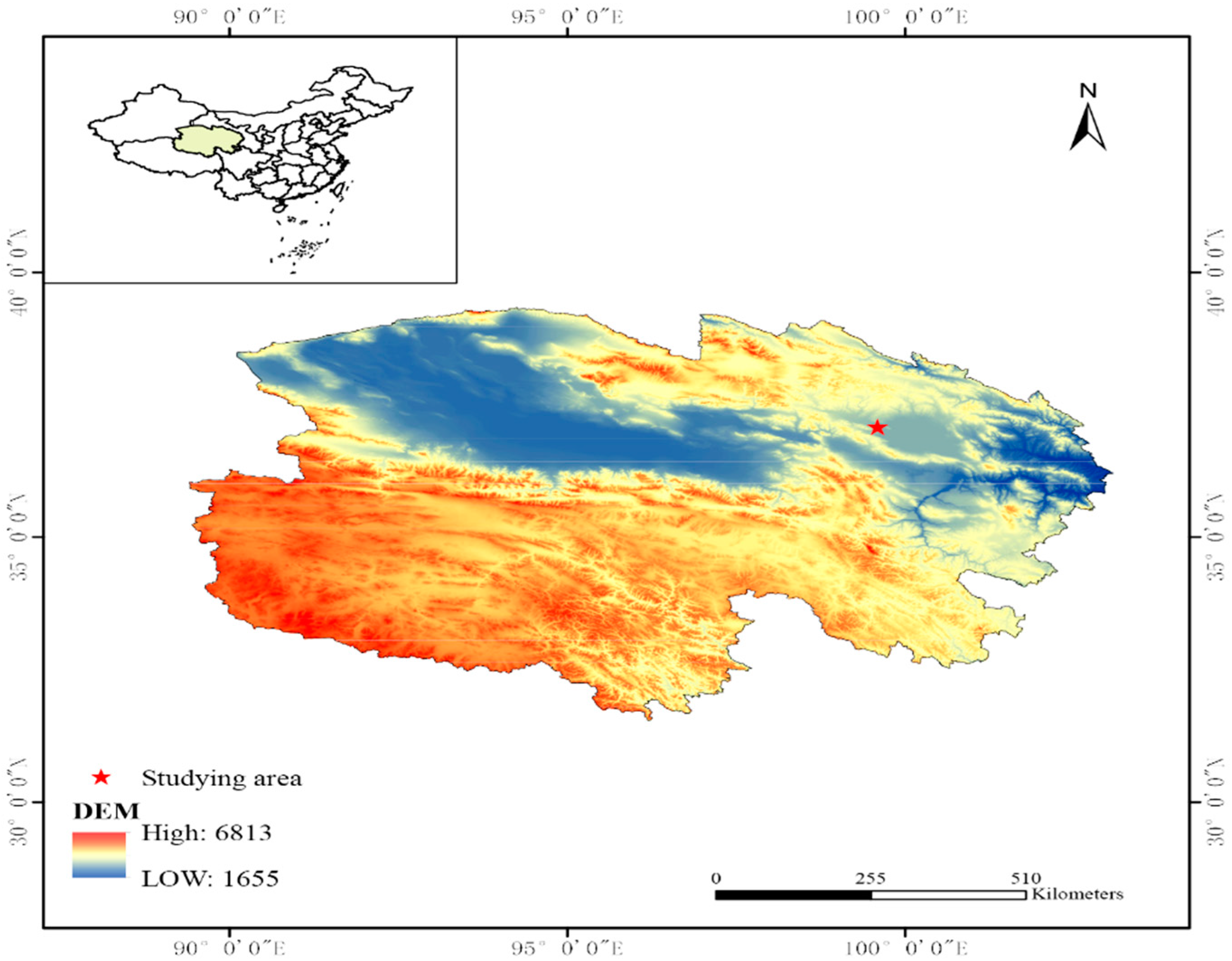
2.2. Overview of the Study Area
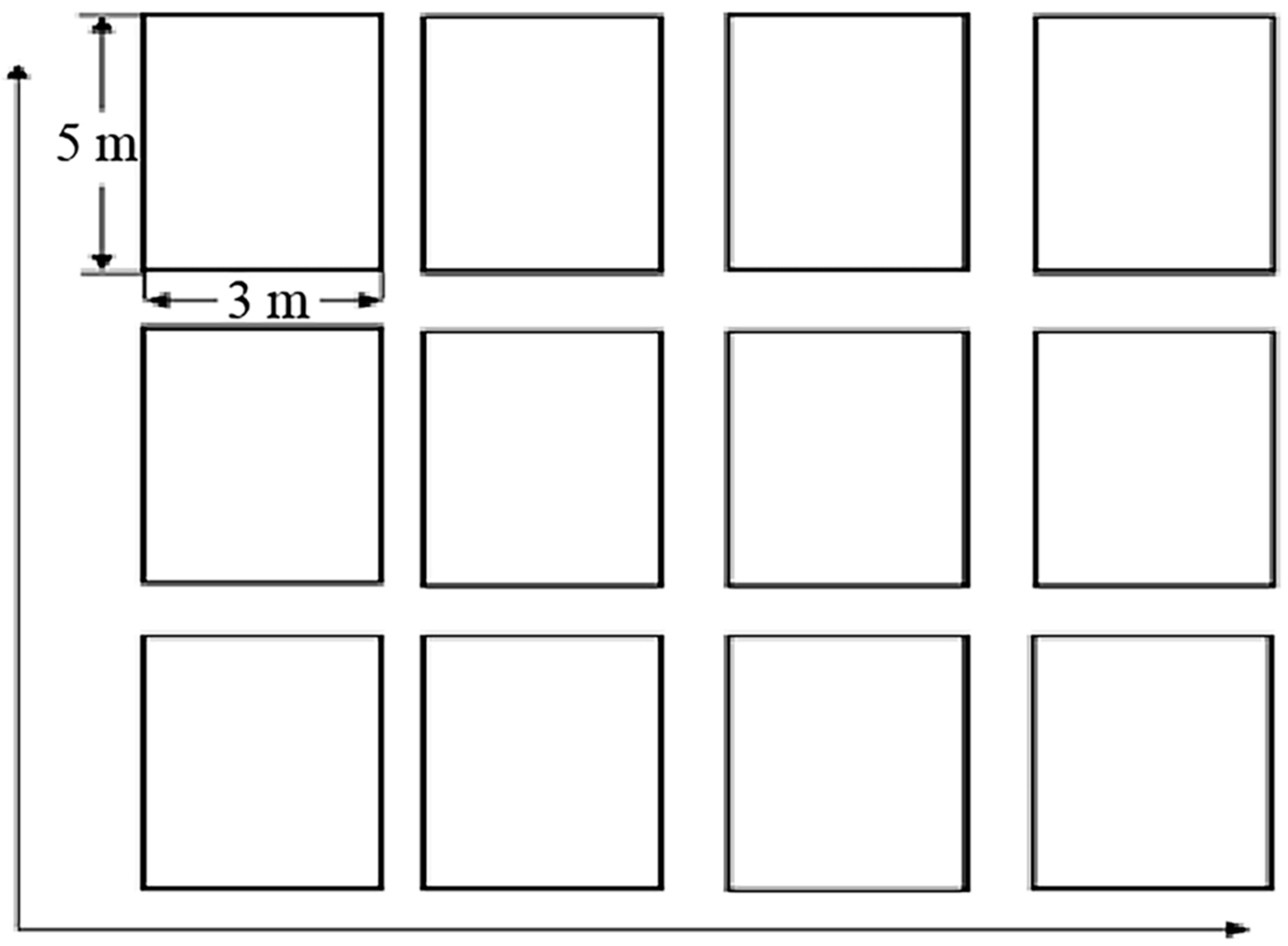
2.3. Experimental Design
2.4. Measurement Index and Method
2.4.1. Root Traits and Soil Physicochemical Index
2.4.2. DNA Extraction and Amplicon Sequencing of 16S rRNA Gene and ITS rRNA Gene
2.5. Processing of the Sequencing Data
3. Results
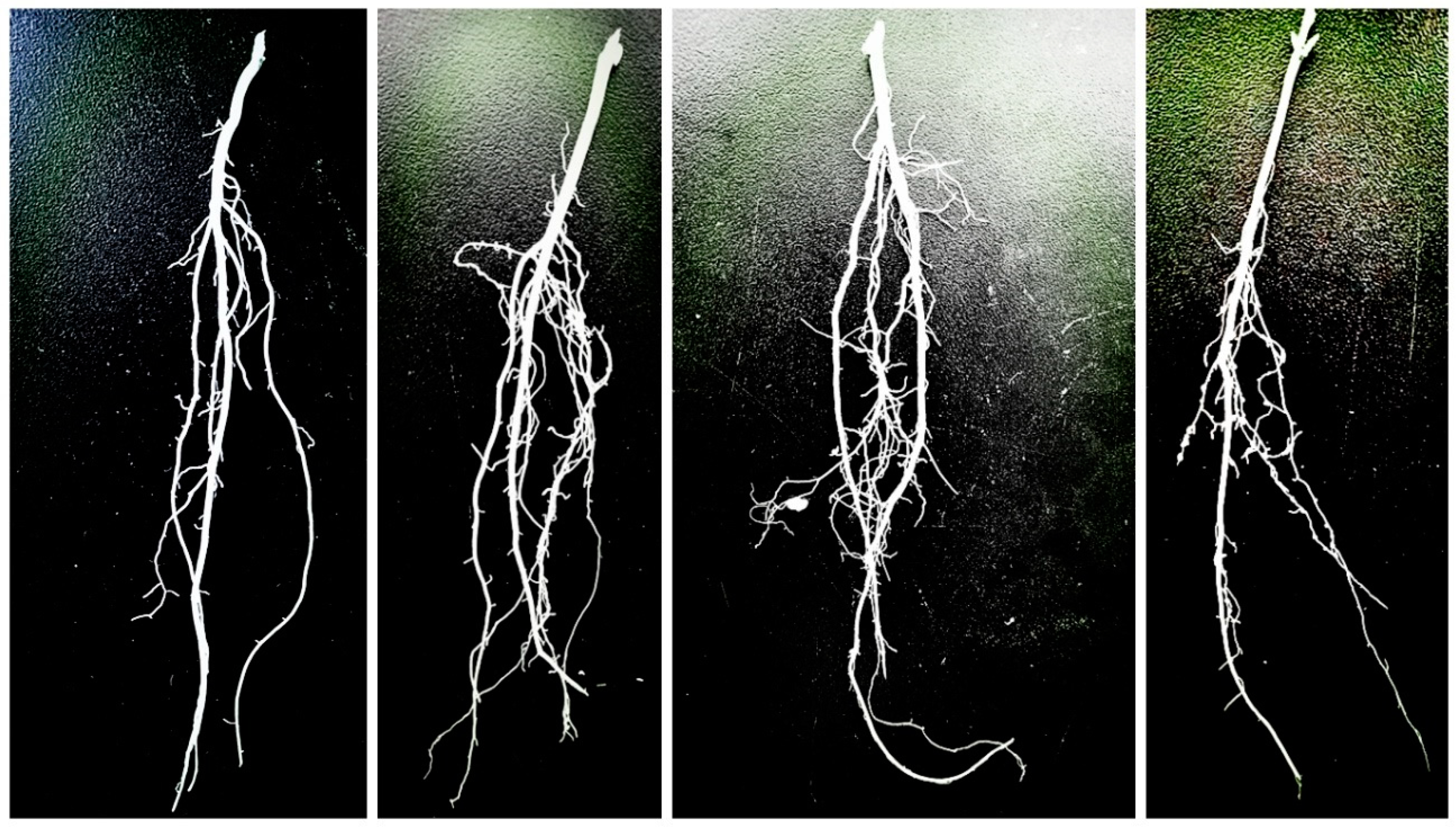
3.1. Analysis of Root System Characteristics after Different Gradient Treatments
3.2. Analysis of Soil Physical and Chemical Properties
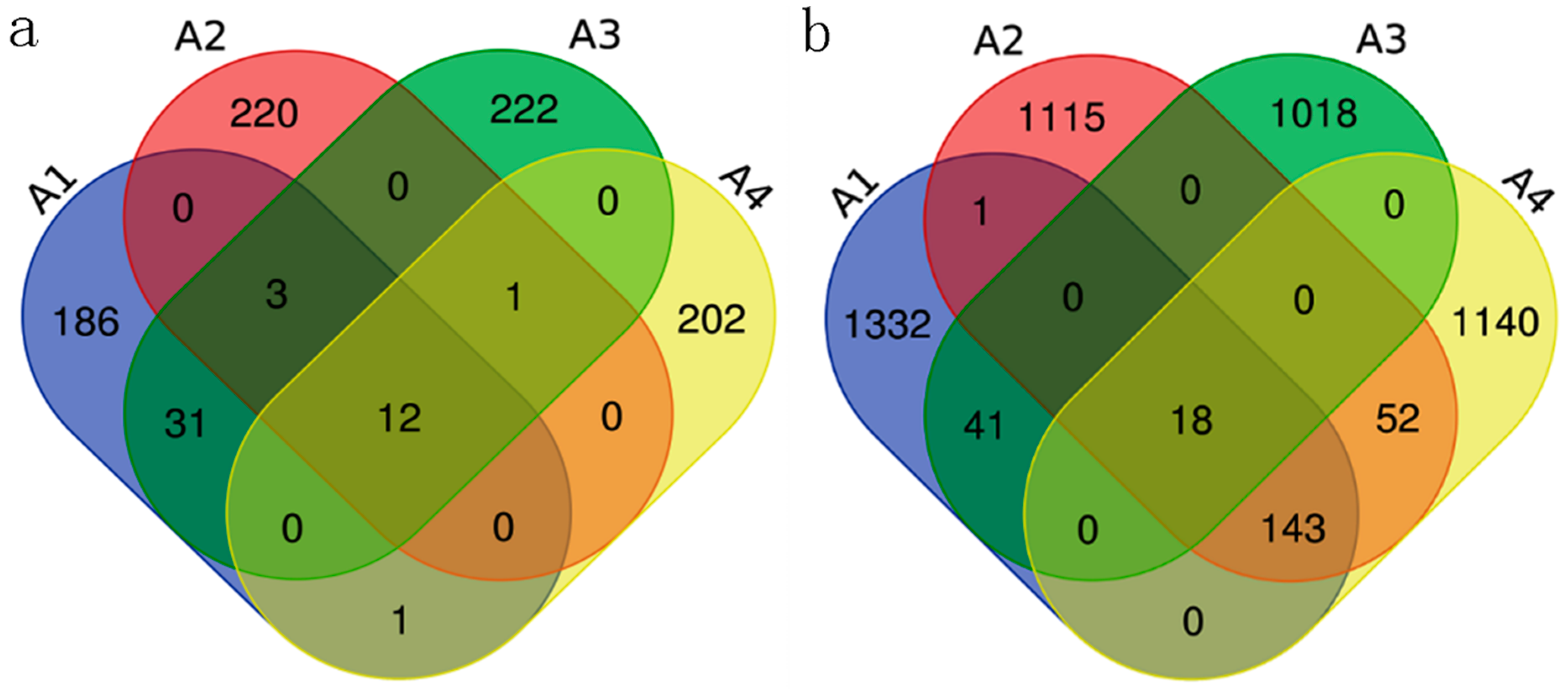
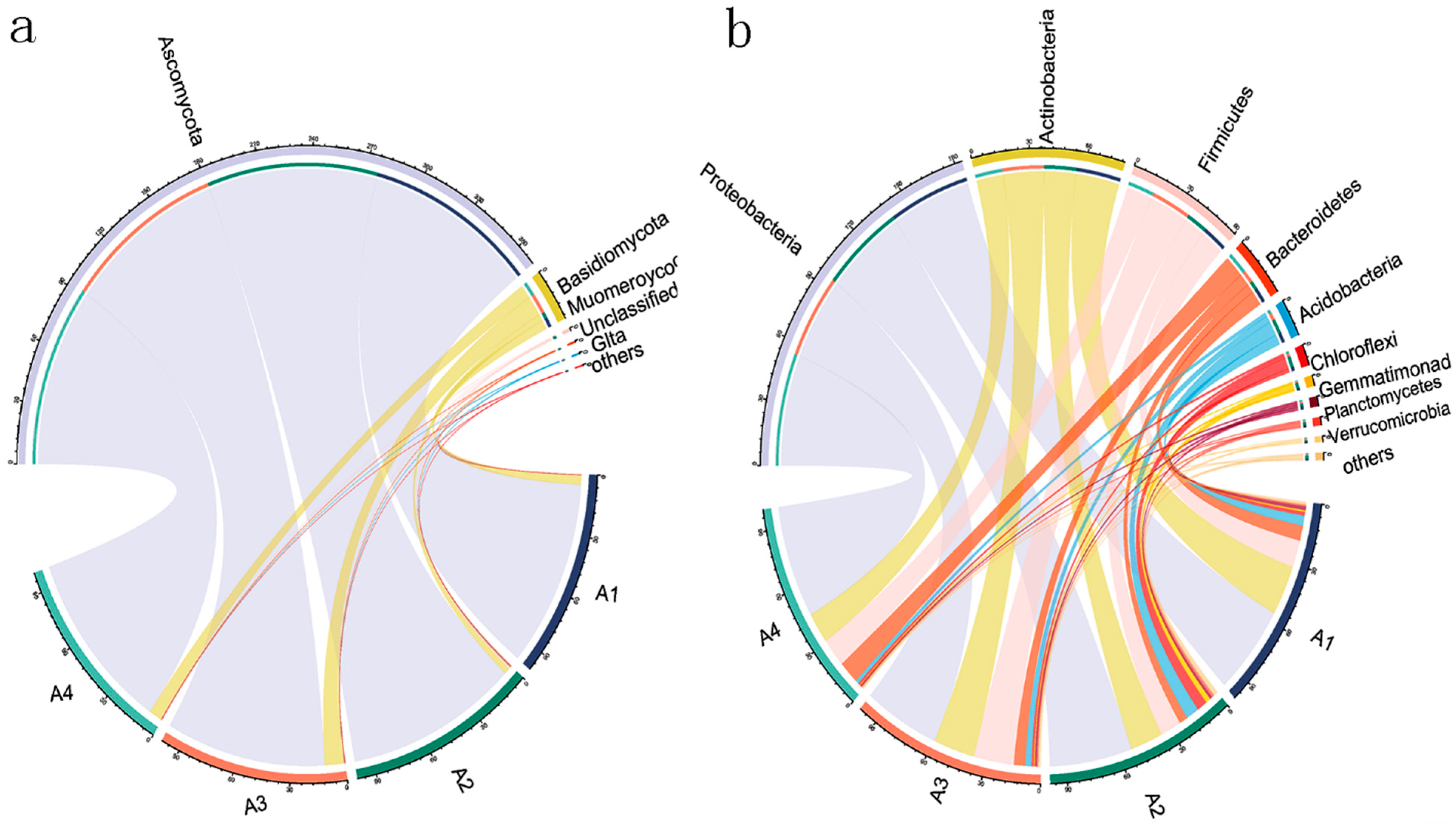
3.3. Soil Microbial Community Composition and Structure Analysis
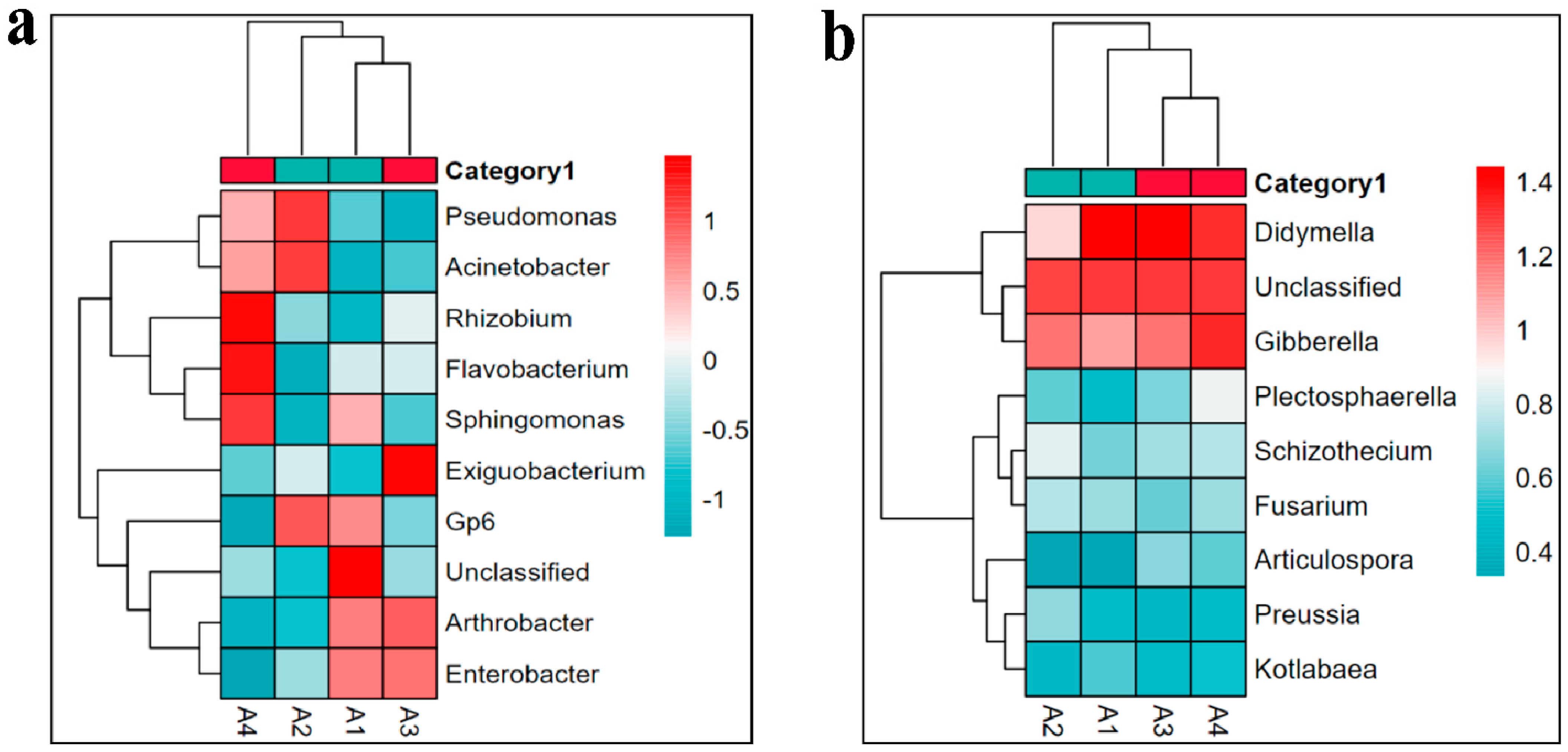
3.4. Biome Diversity Analysis
3.5. The Relationship between Soil Microorganisms and Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kumawat, K.C.; Nagpal, S.; Sharma, P. Potential of plant growth-promoting rhizobacteria-plant interactions in mitigating salt stress for sustainable agriculture: A review. Pedosphere 2022, 32, 223–245. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Knight, M.I.; Moate, P.J.; Jacobs, J.L. An alternative approach for sustainable sheep meat production: Implications for food security. J. Anim. Sci. Biotechnol. 2021, 12, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Rong-Zhen, Z.; Yi, F.; Hai-Xia, S.; Min, W.; Dao-Wei, Z. Rumen methane output and fermentation characteristics of gramineous forage and leguminous forage at differing harvest dates determined using an in vitro gas production technique. J. Integr. Agric. 2016, 15, 414–423. [Google Scholar]
- Xu, J.; Yue, X. Study on the Ecotoxicity of Three Veterinary Antibiotics Against Alfalfa (Medicago sativa L.). Agric. Biotechnol. 2019, 8, 166–168. [Google Scholar]
- Tasker, A. Noxious weed control in the USA. Pestic. Outlook 2000, 11, 104–106. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Z. China’s alfalfa market and imports: Development, trends, and potential impacts of the U.S.–China trade dispute and retaliations. J. Integr. Agric. 2020, 19, 1149–1158. [Google Scholar] [CrossRef]
- Wei, J.; Bai, R.; Gao, X.; Li, Z.; Zhao, C. Research Progress on the Factors Affecting the Quality of Scutellaria baicalensis Georgi. Med. Plant 2021, 12, 63–66, 69. [Google Scholar]
- Sehrawat, A.; Sindhu, S.S.; Glick, B.R. Hydrogen cyanide production by soil bacteria: Biological control of pests and promotion of plant growth in sustainable agriculture. Pedosphere 2022, 32, 15–38. [Google Scholar] [CrossRef]
- Shahid, M.; Khan, M.S. Fungicide tolerant Bradyrhizobium japonicum mitigate toxicity and enhance greengram production under hexaconazole stress. J. Environ. Sci. 2019, 78, 92–108. [Google Scholar] [CrossRef]
- Wang, Y.; Kamagata, Y.; Li, M.; Han, F.; Wang, F.; Xiao, X. New approaches for archaeal genome-guided cultivation. Sci. China (Earth Sci.) 2021, 64, 1658–1673. [Google Scholar] [CrossRef]
- Liu, X.; Wang, W.; Li, J.; Li, Y.; Zhang, J.; Tan, H. A widespread response of Gram-negative bacterial acyl-homoserine lactone receptors to Gram-positive Streptomyces γ-butyrolactone signaling molecules. Sci. China (Life Sci.) 2021, 64, 1575–1589. [Google Scholar] [CrossRef] [PubMed]
- Sidjabat, H.E.; Kamolvit, W.; Wailan, A.; Paterson, D.L. Multi-drug-resistant Gram-negative bacteria. Microbiol. Aust. 2013, 34, 43–46. [Google Scholar] [CrossRef] [Green Version]
- Jérémie, B.; Pilar, B.-S.; Caroline, H.; Christine, L.-B.; Martin, C. Complexity of miRNA-dependent regulation in root symbiosis. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1570–1579. [Google Scholar]
- Rose, S.L.; Youngberg, C.T. Tripartite associations in snowbrush (Ceanothus velutinus): Effect of vesicular–arbuscular mycorrhizae on growth, nodulation, and nitrogen fixation. Botany 1981, 59, 34–39. [Google Scholar] [CrossRef]
- Yang, J.; Lan, L.; Jin, Y.; Yu, N.; Wang, D.; Wang, E. Mechanisms underlying legume–rhizobium symbioses. J. Integr. Plant Biol. 2022, 64, 244–267. [Google Scholar] [CrossRef] [PubMed]
- Shupu, W.; Xin, G.; Jiaqiang, L.; Na, Z.; Zengkun, G.; Baijun, S. Ecological environment quality evaluation of the Sahel region in Africa based on remote sensing ecological index. J. Arid. Land 2022, 14, 14–33. [Google Scholar]
- Chen, L.; He, F.; Long, R.; Zhang, F.; Li, M.; Wang, Z.; Kang, J.; Yang, Q. A global alfalfa diversity panel reveals genomic selection signatures in Chinese varieties and genomic associations with root development. J. Integr. Plant Biol. 2021, 63, 1937–1951. [Google Scholar] [CrossRef]
- Wang, K.; Liu, Y.; Teng, F.; Cen, H.; Yan, J.; Lin, S.; Li, D.; Zhang, W. Heterogeneous expression of Osa-MIR156bc increases abiotic stress resistance and forage quality of alfalfa. Crop J. 2021, 9, 1135–1144. [Google Scholar] [CrossRef]
- Kibalou, P.K.; Ruixian, L.; Qun, G.; Juwu, G.; Junwen, L.; Quanwei, L.; Pengtao, L.; Youlu, Y.; Wankui, G. Current advances in pathogen-plant interaction between Verticillium dahliae and cotton provide new insight in the disease management. J. Cotton Res. 2021, 4, 282–294. [Google Scholar]
- Wang, X.; Chang, S.; Lu, J.; Fray, R.; Grierson, D.; Han, Y. Plant genetic engineering and genetically modified crop breeding: History and current status. Front. Agric. Sci. Eng. 2017, 4, 5–27. [Google Scholar] [CrossRef] [Green Version]
- Chandarana, K.A.; Amaresan, N. Soil protists: An untapped microbial resource of agriculture and environmental importance. Pedosphere 2022, 32, 184–197. [Google Scholar] [CrossRef]
- Han, C.; Liu, Y.; Zhang, C.; Li, Y.; Zhou, T.; Khan, S.; Chen, N.; Zhao, C. Effects of three coniferous plantation species on plant-soil feedbacks and soil physical and chemical properties in semi-arid mountain ecosystems. For. Ecosyst. 2021, 8, 22–34. [Google Scholar] [CrossRef]
- Chen, T.; Xiao, H. Coordinated Development of Urbanization and Animal Husbandry Industrialization in Western Regions of China. Asian Agric. Res. 2016, 8, 27–30, 35. [Google Scholar]
- Tan, D.; Wang, G.; Zhu, Y.; Chen, X.; Ma, J.; Li, R.; Xie, J. Thoughts on Present Situation of Development of Rural Animal Husbandry and Its Relationship with Construction of Ecological Agriculture and Revitalization of Rural Industry. Asian Agric. Res. 2021, 13, 20–23, 31. [Google Scholar]
- Yong-Fang, Z.; Chun-Yan, Z.; Bo, Z.; Man, Y.; Hui-Long, H.; Li-Li, Y.; Hua-Wei, G.; Yong-Zhe, G.; Zhang-Xiong, L.; Fu-Heng, L.; et al. Establishment and application of an accurate identification method for fragrant soybeans. J. Integr. Agric. 2021, 20, 1193–1203. [Google Scholar]
- Sun, X.; Wang, G.; Ye, Y.; Ma, Q.; Guan, Q.; Jones, D.L. Response of nitrogen fractions in the rhizosphere and bulk soil to organic mulching in an urban forest plantation. J. For. Res. 2021, 32, 2577–2588. [Google Scholar] [CrossRef]
- Wei-Hua, L.; Qi-Zhi, L. Changes in fungal community and diversity in strawberry rhizosphere soil after 12 years in the greenhouse. J. Integr. Agric. 2019, 18, 677–687. [Google Scholar]
- Sadeghian, F.; Jahandari, S.; Haddad, A.; Rasekh, H.; Li, J. Effects of variations of voltage and pH value on the shear strength of soil and durability of different electrodes and piles during electrokinetic phenomenon. J. Rock Mech. Geotech. Eng. 2022, 14, 625–636. [Google Scholar] [CrossRef]
- Li, J.; Shao, X.; Huang, D.; Liu, K.; Shang, J.; Zhang, Q.; Zhao, T.; Yang, X. Short-term biochar effect on soil physicochemical and microbiological properties of a degraded alpine grassland. Pedosphere 2022, 32, 426–437. [Google Scholar] [CrossRef]
- Manoj, S.; Thirumurugan, M.; Elango, L. Determination of distribution coefficient of uranium from physical and chemical properties of soil. Chemosphere 2020, 244, 125411. [Google Scholar] [CrossRef]
- Kai, F.; Zhaojing, Z.; Weiwei, C.; Wenzong, L.; Meiying, X.; Huaqun, Y.; Aijie, W.; Zhili, H.; Ye, D. Biodiversity and species competition regulate the resilience of microbial biofilm community. Mol. Ecol. 2017, 26, 6170–6182. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.T. Molecular Biodesign of Plant Leaves and Flowers. J. Plant Biotechnol. 2003, 5, 137–142. [Google Scholar]
- Wu, D.; Andales, A.A.; Yang, H.; Sun, Q.; Chen, S.; Guo, X.; Li, D.; Du, T. Linking Crop Water Productivity to Soil Physical, Chemical and Microbial Properties. Front. Agric. Sci. Eng. 2021, 8, 545–558. [Google Scholar] [CrossRef]
- Shao, Z.Q.; Zheng, C.C.; Postma, J.A.; Lu, W.L.; Gao, Q.; Gao, Y.Z.; Zhang, J.J. Nitrogen acquisition, fixation and transfer in maize/alfalfa intercrops are increased through root contact and morphological responses to interspecies competition. J. Integr. Agric. 2021, 20, 2240–2254. [Google Scholar] [CrossRef]
- Palansooriya, K.N.; Wong, J.T.F.; Hashimoto, Y.; Huang, L.; Rinklebe, J.; Chang, S.X.; Bolan, N.; Wang, H.; Ok, Y.S. Response of microbial communities to biochar-amended soils: A critical review. Biochar 2019, 1, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Ting-Ting, X.; An-Dong, C.; Chang-Ai, L.; Hong-Ling, Y.; Hong-Liang, W.; Sheng-Chang, H.; Jin-Yu, W.; Ming-Gang, X.; Qi-Mei, L. Increasing soil microbial biomass nitrogen in crop rotation systems by improving nitrogen resources under nitrogen application. J. Integr. Agric. 2022, 21, 1488–1500. [Google Scholar]
- Sazawa, K.; Sugano, T.; Kuramitz, H. High-heat Effects On the Physical and Chemical Properties of Soil Organic Matter and Its Water-soluble Components In Japan’s Forests: A Comprehensive Approach Using. Sci. Lett. 2020, 601–609. [Google Scholar]
- Sokołowska, J.; Józefowska, A.; Woźnica, K.; Zaleski, T. Succession from meadow to mature forest: Impacts on soil biological, chemical and physical properties—Evidence from the Pieniny Mountains, Poland. CATENA 2020, 189, 104503. [Google Scholar] [CrossRef]
- Xue, L.; Li, Q.; Chen, H. Effects of a Wildfire on Selected Physical, Chemical and Biochemical Soil Properties in a Pinus massoniana Forest in South China. Forests 2014, 5, 2947–2966. [Google Scholar] [CrossRef] [Green Version]
- Yun-Feng, C.; Xian-Ge, X.; Cheng, H.; Dong-Hai, L.; Yan, L.; Shuang-Lai, F. Effects of long-term straw incorporation on nematode community composition and metabolic footprint in a rice–wheat cropping system. J. Integr. Agric. 2021, 20, 2265–2276. [Google Scholar]
- Chen, B.; Pan, L.; Xing, Y.; Chen, J.; Deng, K.; Liu, Y. Study on the Activation Effect of Fermented Organic Fertilizers on Selenium in the Soil of Southern Dragon Fruit Orchards. Agric. Biotechnol. 2021, 10, 83–85. [Google Scholar]
- Li, F.; Lu, B.; Yang, L.; Yang, R.; Qian, F.; Xu, X.; Li, J. Effects of Partial Replacement of Chemical Fertilizer by Bio-organic Fertilizer Kunyijian on Yield and Quality of Tobacco Leaves. Asian Agric. Res. 2020, 12, 42–46. [Google Scholar]
- Geng, Y.; Yuan, Y.; Miao, Y.; Zhi, J.; Huang, M.; Zhang, Y.; Wang, H.; Shen, Q.; Zou, J.; Li, S. Decreased nitrous oxide emissions associated with functional microbial genes under bio-organic fertilizer application in vegetable fields. Pedosphere 2021, 31, 279–288. [Google Scholar] [CrossRef]
- Lan, J.; Xie, S. Effects of Organic Compound Fertilizers on Soil Water Infiltration Characteristics. Agric. Biotechnol. 2020, 9, 104–110, 167. [Google Scholar]
- Cheng, W.L.; Li, Z.E.N.G.; Xue, Y.A.N.G.; Huang, D.; Hao, Y.U.; Wen, C.H.E.N.; Zhang, J.B. Preparation and efficacy evaluation of Paenibacillus polymyxa KM2501-1 microbial organic fertilizer against root-knot nematodes. J. Integr. Agric. 2022, 21, 542–551. [Google Scholar] [CrossRef]
- Ma, Y.; Yin, S.L.; Zhang, Y.X. Research and Discussion on Physical and Chemical Properties of Cultivating Substrate with Facilities. Chin. For. Sci. Technol. 2008, 2, 49–55. [Google Scholar]
- Ndaba, B.; Roopnarain, A.; Rama, H.; Maaza, M. Biosynthesized metallic nanoparticles as fertilizers:An emerging precision agriculture strategy. J. Integr. Agric. 2022, 21, 1225–1242. [Google Scholar] [CrossRef]
- Meki, K.; Liu, Q.; Wu, S.; Yuan, Y. Plant-and microbe-assisted biochar amendment technology for petroleum hydrocarbon remediation in saline-sodic soils: A review. Pedosphere 2022, 32, 211–221. [Google Scholar] [CrossRef]
- Guo, M.; Gong, Z.; Miao, R.; Su, D.; Li, X.; Jia, C.; Zhuang, J. Impact of a bacterial consortium on the soil bacterial community structure and maize (Zea mays L.) cultivation. Sci. Rep. 2021, 11, 13092. [Google Scholar]
- Hua, Z.; Jia-Rui, S.; Lu, Z.; Fang, B.; Xian-Zhu, Z. Effect of altitude and climatic parameters on shrub-meadow community composition and diversity in the dry valley region of the eastern Hengduan Mountains, China. J. Mt. Sci. 2022, 19, 1139–1155. [Google Scholar]
- Xiao, F.; Li, Y.; Li, G.; He, Y.; Lv, X.; Zhuang, L.; Pu, X. High throughput sequencing-based analysis of the soil bacterial community structure and functions of Tamarix shrubs in the lower reaches of the Tarim River. PeerJ. 2021, 9, e12105. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, A.; Fox, A.; Jongen, M.; Melo, J.; Musyoki, M.; Vieira, A.; Zimmermann, J.; Carlsson, G.; Cruz, C.; Lüscher, A.; et al. Soil bacteria respond to regional edapho-climatic conditions while soil fungi respond to management intensity in grasslands along a European transect. Appl. Soil Ecol. 2022, 170, 104264. [Google Scholar] [CrossRef]
- Li, K.; Han, X.; Ni, R.; Shi, G.; de-Miguel, S.; Li, C.; Shen, W.; Zhang, Y.; Zhang, X. Impact of Robinia pseudoacacia stand conversion on soil properties and bacterial community composition in Mount Tai, China. For. Ecosyst. 2021, 8, 253–264. [Google Scholar] [CrossRef]
- Wang, X.; Whalley, W.R.; Miller, A.J.; White, P.J.; Zhang, F.; Shen, J. Biology and chemistry of rhizosphere: Their role in nutrient mobilisation. Int. J. Microbiol. Res. 2020, 12, 1884–1889. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | Mixed Bacteria Gradient | Repetition | Numbering |
|---|---|---|---|
| Beilin 201 | 0 | 3 | A1 |
| 25% | 3 | A2 | |
| 50% | 3 | A3 | |
| 75% | 3 | A4 |
| Group | Main Root Length/cm | Main Root Diameter/mm | Lateral Root Number | Max. Root Width/cm | Nodule Number |
|---|---|---|---|---|---|
| A1 | 31.89 ± 1.108 a | 5.06 ± 0.248 a | 3.27 ± 0.656 a | 10.98 ± 1.341 ab | 3.33 ± 0.333 b |
| A2 | 33.43 ± 0.653 a | 5.41 ± 0.240 a | 3.00 ± 0.577 a | 10.20 ± 0.327 ab | 4.33 ± 0.881 ab |
| A3 | 34.19 ± 1.852 a | 5.49 ± 0.068 a | 3.60 ± 0.305 a | 12.43 ± 0.993 a | 6.00 ± 0.577 a |
| A4 | 30.93 ± 1.326 a | 5.01 ± 0.658 a | 2.60 ± 0.346 a | 8.55 ± 0.427 b | 4.00 ± 0 b |
| Sample | Soil Moisture | Soil Conductivity (μS/cm) | TN (%) | Nitrate Nitrogen (mg/kg) | TOC (%) |
|---|---|---|---|---|---|
| A1 | 15.43 ± 2.46 a | 0.12 ± 0.04 a | 0.16 ± 119.41 b | 34.74 ± 4.35 a | 2.08 ± 0.07 b |
| A2 | 15.13 ± 2.04 a | 0.08 ± 0.03 a | 0.14 ± 6.51 ab | 32.61 ± 2.36 a | 1.65 ± 0.38 ab |
| A3 | 16.19 ± 0.85 a | 0.10 ± 0.02 a | 0.15 ± 54.24 b | 34.11 ± 0.70 a | 2.18 ± 0.26 b |
| A4 | 14.99 ± 0.91 a | 0.09 ± 0.01 a | 0.12 ± 196.27 a | 30.50 ± 3.10 a | 1.46 ± 0.35 a |
| Microbe | Samples | Shannon | Inv_Simpson | Observed_Richness | Simpson_Evenness |
|---|---|---|---|---|---|
| Bacter | A1 | 4.798 ± 0.503 a | 22.359 ± 4.912 a | 2061.67 ± 451.23 a | 0.011 ± 0.001 a |
| A2 | 4.681 ± 0.661 a | 27.653 ± 14.308 a | 1901.00 ± 347.15 a | 0.014 ± 0.005 a | |
| A3 | 4.346 ± 0.789 a | 28.078 ± 15.466 a | 1739.33 ± 435.12 a | 0.014 ± 0.006 a | |
| A4 | 4.665 ± 0.209 a | 28.871 ± 5.978 a | 1761.67 ± 128.23 a | 0.016 ± 0.002 a | |
| Fungi | A1 | 3.874 ± 0.551 a | 21.723 ± 11.377 a | 825.33 ± 79.55 a | 0.024 ± 0.011 a |
| A2 | 3.920 ± 0.633 a | 22.514 ± 8.982 a | 771.67 ± 159.91 a | 0.025 ± 0.008 a | |
| A3 | 4.389 ± 0.127 a | 26.132 ± 5.357 a | 909.00 ± 60.85 a | 0.028 ± 0.005 a | |
| A4 | 4.168 ± 0.157 a | 22.112 ± 1.632 a | 791.33 ± 82.49 a | 0.028 ± 0.004 a |
| Correlation | Shannon | Inv_Simpson | Observed_Richness | Pielou_Evenness | Soil Moisture | Soil Conductivity | TN | Nitrate Nitrogen | TOC |
|---|---|---|---|---|---|---|---|---|---|
| Shannon | 1 | 0.866 | 0.973 | 0.997 * | 0.908 | 0.807 | 0.084 | 0.295 | 0.587 |
| Inv_Simpson | 0.807 ** | 1 | 0.728 | 0.899 | 0.576 | 0.403 | 0.571 | 0.733 | 0.103 |
| Observed_richness | 0.941 ** | 0.621 * | 1 | 0.954 | 0.980 | 0.921 | −0.147 | 0.067 | 0.757 |
| Pielou_evenness | 0.985 ** | 0.827 ** | 0.902 ** | 1 | 0.875 | 0.762 | 0.155 | 0.363 | 0.527 |
| Soil Moisture | 0.502 | 0.427 | 0.595 * | 0.427 | 1 | 0.980 | −0.342 | −0.134 | 0.873 |
| Soil conductivity | 0.413 | 0.132 | 0.596 * | 0.434 | 0.45 | 1 | −0.521 | −0.327 | 0.952 |
| TN | −0.176 | −0.293 | −0.039 | −0.195 | 0.1 | 0.154 | 1 | 0.977 | −0.758 |
| nitrate nitrogen | −0.059 | −0.168 | −0.006 | −0.064 | 0.021 | −0.071 | 0.741 ** | 1 | −0.601 |
| TOC | −0.291 | −0.479 | −0.077 | −0.335 | 0.119 | 0.092 | 0.788 ** | 0.592 * | 1 |
| Correlation | TN | Nitrate Nitrogen | TOC | Soil Moisture | Soil Conductivity | Ascomycota | Basidiomycota | Proteobacteria | Actinobacteria | Firmicutes |
|---|---|---|---|---|---|---|---|---|---|---|
| TN | 1 | 0.977 | −0.758 | −0.342 | −0.521 | 0.008 | −0.001 | −0.120 | 0.735 | −0.732 |
| nitrate nitrogen | 0.741 ** | 1 | −0.601 | −0.134 | −0.327 | 0.221 | −0.214 | −0.329 | 0.863 | −0.569 |
| TOC | 0.788 ** | 0.592 * | 1 | 0.873 | 0.952 | 0.647 | −0.652 | −0.557 | −0.115 | 0.999 * |
| Soil Moisture | 0.054 | −0.197 | 0.100 | 1 | 0.980 | 0.937 | −0.939 | −0.892 | 0.385 | 0.891 |
| Soil conductivity | 0.265 | 0.038 | 0.238 | 0.356 | 1 | 0.850 | −0.853 | −0.785 | 0.195 | 0.963 |
| Ascomycota | −0.266 | −0.128 | −0.427 | 0.121 | 0.006 | 1 | −1.000 ** | −0.994 | 0.683 | 0.676 |
| Basidiomycota | 0.280 | 0.075 | 0.407 | −0.040 | −0.018 | −0.982 ** | 1 | 0.993 | −0.678 | −0.681 |
| Proteobacteria | −0.381 | −0.609 * | −0.506 | −0.257 | −0.431 | 0.276 | −0.179 | 1 | −0.761 | −0.589 |
| Actinobacteria | 0.718 ** | 0.855 ** | 0.737 ** | 0.217 | 0.224 | −0.080 | 0.055 | −0.703 * | 1 | −0.076 |
| Firmicutes | 0.118 | −0.233 | 0.050 | 0.620 * | 0.299 | −0.362 | 0.477 | −0.110 | −0.072 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Lu, G.; Jin, X.; Wang, Y.; Ma, K.; Zhang, H.; Yan, H.; Zhou, X. Effects of Microbial Fertilizer on Soil Fertility and Alfalfa Rhizosphere Microbiota in Alpine Grassland. Agronomy 2022, 12, 1722. https://doi.org/10.3390/agronomy12071722
Zhao Y, Lu G, Jin X, Wang Y, Ma K, Zhang H, Yan H, Zhou X. Effects of Microbial Fertilizer on Soil Fertility and Alfalfa Rhizosphere Microbiota in Alpine Grassland. Agronomy. 2022; 12(7):1722. https://doi.org/10.3390/agronomy12071722
Chicago/Turabian StyleZhao, Yangan, Guangxin Lu, Xin Jin, Yingcheng Wang, Kun Ma, Haijuan Zhang, Huilin Yan, and Xueli Zhou. 2022. "Effects of Microbial Fertilizer on Soil Fertility and Alfalfa Rhizosphere Microbiota in Alpine Grassland" Agronomy 12, no. 7: 1722. https://doi.org/10.3390/agronomy12071722
APA StyleZhao, Y., Lu, G., Jin, X., Wang, Y., Ma, K., Zhang, H., Yan, H., & Zhou, X. (2022). Effects of Microbial Fertilizer on Soil Fertility and Alfalfa Rhizosphere Microbiota in Alpine Grassland. Agronomy, 12(7), 1722. https://doi.org/10.3390/agronomy12071722