

The Effects of Short-Time Delayed Sealing on Fermentation, Aerobic Stability and Chemical Composition on Maize Silages

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experiment Set-Up
2.2. Ensiling
2.3. Chemical Composition, Gas Production, and Fermentative Profile of Silages
2.4. Microbiological Analyses
2.5. Aerobic Stability Test
2.6. Statistical Analyses
3. Results
3.1. Changes in pH and Chemical Composition of Silages
3.2. Fermentation Profile and Influence of Silage on Gas Dynamics and Effluent Losses
3.3. Microbial Counts and Aerobic Stability
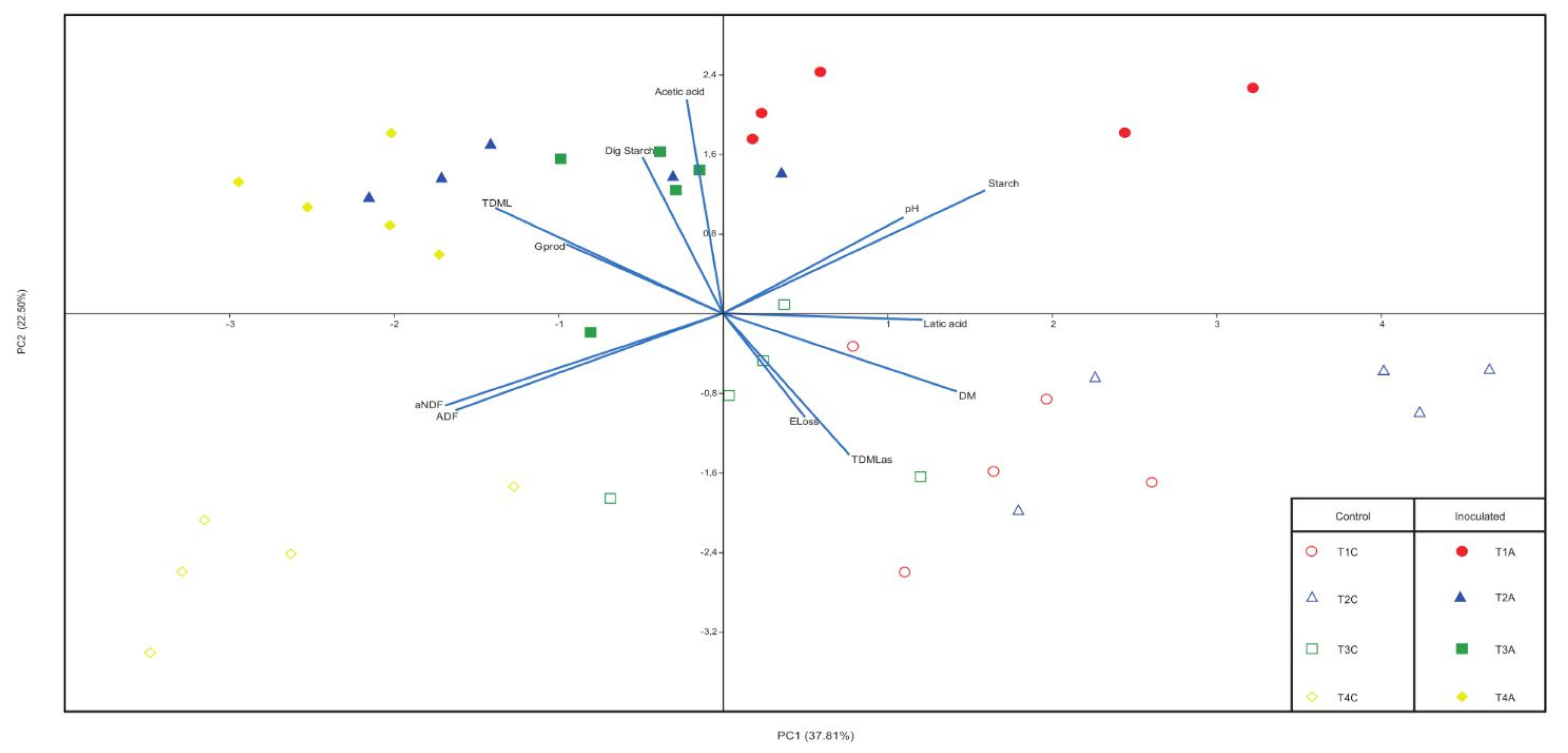
3.4. Association between Chemical and Fermentative Parameters of Silages
4. Discussion
4.1. Changes in pH and Chemical Composition of Silage
4.2. Fermentation Profile and Influence of Silage on Gas Dynamics and Effluent Losses
4.3. Microbial Counts and Aerobic Stability of Silage
4.4. Association between Chemical and Fermentative Parameters of Silage
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Daniel, J.L.P.; Bernardes, T.F.; Jobim, C.C.; Schmidt, P.; Nussio, L.G. Production and Utilization of Silages in Tropical Areas with Focus on Brazil. Grass Forage Sci. 2019, 74, 188–200. [Google Scholar] [CrossRef]
- De Oliveira, I.L.; Lima, L.M.; Casagrande, D.R.; Lara, M.A.S.; Bernardes, T.F. Nutritive Value of Corn Silage from Intensive Dairy Farms in Brazil. Rev. Bras. Zootec. 2017, 46, 494–501. [Google Scholar] [CrossRef] [Green Version]
- Ferraretto, L.F.; Shaver, R.D.; Luck, B.D. Silage Review: Recent Advances and Future Technologies for Whole-Plant and Fractionated Corn Silage Harvesting. J. Dairy Sci. 2018, 101, 3937–3951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ávila, C.L.S.; Carvalho, B.F. Silage Fermentation—Updates Focusing on the Performance of Micro-Organisms. J. Appl. Microbiol. 2019, 128, 966–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, L.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage Review: Interpretation of Chemical, Microbial, and Organoleptic Components of Silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L. Silage Review: Recent Advances and Future Uses of Silage Additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.M.; Rinne, M. Highlights of Progress in Silage Conservation and Future Perspectives. Grass Forage Sci. 2018, 73, 40–52. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Davies, D.R. The Aerobic Stability of Silage: Key Findings and Recent Developments. Grass Forage Sci. 2013, 68, 1–19. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe Publications: Marlow, UK, 1991; p. 340. [Google Scholar]
- Brüning, D.; Gerlach, K.; Weiß, K.; Südekum, K.-H. Effect of Compaction, Delayed Sealing and Aerobic Exposure on Maize Silage Quality and on Formation of Volatile Organic Compounds. Grass Forage Sci. 2018, 73, 53–66. [Google Scholar] [CrossRef]
- Kim, S.C.; Adesogan, A.T. Influence of Ensiling Temperature, Simulated Rainfall, and Delayed Sealing on Fermentation Characteristics and Aerobic Stability of Corn Silage. J. Dairy Sci. 2006, 89, 3122–3132. [Google Scholar] [CrossRef]
- Mills, J.A.; Kung, L., Jr. The Effect of Delayed Ensiling and Application of a Propionic Acid-Based Additive on the Fermentation of Barley Silage. J. Dairy Sci. 2002, 85, 1969–1975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, K.; Kroschewski, B.; Auerbach, H. Effects of Air Exposure, Temperature and Additives on Fermentation Characteristics, Yeast Count, Aerobic Stability and Volatile Organic Compounds in Corn Silage. J. Dairy Sci. 2016, 99, 8053–8069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage Review: Factors Affecting Dry Matter and Quality Losses in Silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [Green Version]
- Bernardi Zardin, P.; Pedro Velho, J.; Cabreira Jobim, C.; Moro Alessio, D.R.; Pereira Haygert-Velho, I.M.; Menegazzi da Conceição, G.; Gois Almeida, P.S. Chemical composition of corn silage produced by scientific studies in Brazil—A meta-analysis. Semin. Ciênc. Agrár. 2017, 38, 503–511. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Soil Survey Staff. Keys to Soil Taxonomy, 11th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2010; p. 338.
- Bueno, A.V.I.; Vigne, G.L.D.; Novinski, C.O.; Bayer, C.; Jobim, C.C.; Schmidt, P. Natamycin as a Potential Silage Additive: A Lab Trial Using Sugarcane to Assess Greenhouse Gas Emissions. Rev. Bras. Zootec. 2020, 49, e20200017. [Google Scholar] [CrossRef]
- Jobim, C.C.; Nussio, L.G.; Reis, R.A.; Schmidt, P. Methodological Advances in Evaluation of Preserved Forage Quality. Rev. Bras. Zootec. 2007, 36, 101–119. [Google Scholar] [CrossRef] [Green Version]
- Restelatto, R.; Novinski, C.O.; Pereira, L.M.; Silva, E.P.A.; Volpi, D.; Zopollatto, M.; Schmidt, P.; Faciola, A.P. Chemical Composition, Fermentative Losses, and Microbial Counts of Total Mixed Ration Silages Inoculated with Different Lactobacillus Species. J. Anim. Sci. 2019, 97, 1634–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, L., Jr.; Grieve, D.B.; Thomas, J.W.; Huber, J.T. Added Ammonia or Microbial Inocula for Fermentation and Nitrogenous Compounds of Alfalfa Ensiled at Various Percents of Dry Matter. J. Dairy Sci. 1984, 67, 299–306. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis: Volume 1. Agricultural Chemicals; Contaminants; Drugs, 15th ed.; Association of Official Analytical Chemists: Arlington, TX, USA, 1990; p. 69. [Google Scholar]
- Hall, M.B. Neutral Detergent Soluble Carbohydrates Nutritional Relevance and Analysis: A Laboratory Manual; University of Florida: Gainesville, FL, USA, 2000; pp. 1–34. [Google Scholar]
- Kung, L., Jr.; Ranjit, N.K. The Effect of Lactobacillus buchneri and Other Additives on the Fermentation and Aerobic Stability of Barley Silage. J. Dairy Sci. 2001, 84, 1149–1155. [Google Scholar] [CrossRef]
- Kung, L., Jr.; Robinson, J.R.; Ranjit, N.K.; Chen, J.H.; Golt, C.M.; Pesek, J.D. Microbial Populations, Fermentation End-Products, and Aerobic Stability of Corn Silage Treated with Ammonia or a Propionic Acid-Based Preservative. J. Dairy Sci. 2000, 83, 1479–1486. [Google Scholar] [CrossRef]
- Hu, W.; Schmidt, R.J.; McDonell, E.E.; Klingerman, C.M.; Kung, L. The Effect of Lactobacillus buchneri 40788 or Lactobacillus plantarum MTD-1 on the Fermentation and Aerobic Stability of Corn Silages Ensiled at Two Dry Matter Contents. J. Dairy Sci. 2009, 92, 3907–3914. [Google Scholar] [CrossRef] [Green Version]
- Ranjit, N.K.; Kung, L., Jr. The Effect of Lactobacillus buchneri, Lactobacillus plantarum, or a Chemical Preservative on the Fermentation and Aerobic Stability of Corn Silage. J. Dairy Sci. 2000, 83, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- de Almeida Carvalho-Estrada, P.; Fernandes, J.; da Silva, É.B.; Tizioto, P.; de Fátima Paziani, S.; Duarte, A.P.; Coutinho, L.L.; Verdi, M.C.Q.; Nussio, L.G. Effects of Hybrid, Kernel Maturity, and Storage Period on the Bacterial Community in High-Moisture and Rehydrated Corn Grain Silages. Syst. Appl. Microbiol. 2020, 43, 126131. [Google Scholar] [CrossRef]
- Massé, D.I.; Jarret, G.; Hassanat, F.; Benchaar, C.; Saady, N.M.C. Effect of Increasing Levels of Corn Silage in an Alfalfa-Based Dairy Cow Diet and of Manure Management Practices on Manure Fugitive Methane Emissions. Agric. Ecosyst. Environ. 2016, 221, 109–114. [Google Scholar] [CrossRef]
- de Lima, E.M.; Gonçalves, L.C.; Keller, K.M.; Rodrigues, J.A.D.S.; Santos, F.P.C.; Michel, P.H.F.; Raposo, V.S.; Jayme, D.G. Re-Ensiling and Its Effects on Chemical Composition, in Vitro Digestibility, and Quality of Corn Silage after Different Lengths of Exposure to Air. Can. J. Anim. Sci. 2016, 97, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.M.; Gonçalves, L.C.; Rodrigues, J.A.S.; Keller, K.M.; Anjos, G.V.D.S.D.; Ottoni, D.; Michel, P.H.F.; Jayme, D.G. Chemical Characteristics, Aerobic Stability, and Microbiological Counts in Corn Silage Re-Ensiled with Bacterial Inoculant. Pesqui. Agropecu. Bras. 2018, 53, 1045–1052. [Google Scholar] [CrossRef]
- Rooke, J.A.; Hatfield, R.D. Biochemistry of Ensiling. In Silage Science and Technology; Buxton, D.R., Muck, R.E., Harrison, J.H., Eds.; American Society of Agronomy Inc., Crop Science Society of America Inc., Soil Science Society of America Inc.: Madison, WI, USA, 2003; pp. 95–139. [Google Scholar]
- Ning, T.; Wang, H.; Zheng, M.; Niu, D.; Zuo, S.; Xu, C. Effects of Microbial Enzymes on Starch and Hemicellulose Degradation in Total Mixed Ration Silages. Asian-Australas. J. Anim. Sci. 2017, 30, 171–180. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Risu, N.; Gentu, G.; Jia, Y.; Cai, Y. Dynamic Changes and Characterization of the Protein and Carbohydrate Fractions of Native Grass Grown in Inner Mongolia during Ensiling and the Aerobic Stage. Asian-Australas. J. Anim. Sci. 2020, 33, 556–567. [Google Scholar] [CrossRef] [Green Version]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Hussin, G.; Ramli, A.; Miah, G. Fermentation Quality and Additives: A Case of Rice Straw Silage. BioMed Res. Int. 2016, 2016, 7985167. [Google Scholar] [CrossRef]
- Hao, W.; Tian, P.; Zheng, M.; Wang, H.; Xu, C. Characteristics of Proteolytic Microorganisms and Their Effects on Proteolysis in Total Mixed Ration Silages of Soybean Curd Residue. Asian-Australas. J. Anim. Sci. 2019, 33, 100–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Givens, D.I.; Rulquin, H. Utilisation by Ruminants of Nitrogen Compounds in Silage-Based Diets. Anim. Feed Sci. Technol. 2004, 114, 1–18. [Google Scholar] [CrossRef]
- Hristov, A.N.; Bannink, A.; Crompton, L.A.; Huhtanen, P.; Kreuzer, M.; McGee, M.; Nozière, P.; Reynolds, C.K.; Bayat, A.R.; Yáñez-Ruiz, D.R.; et al. Invited Review: Nitrogen in Ruminant Nutrition: A Review of Measurement Techniques. J. Dairy Sci. 2019, 102, 5811–5852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, P.C.; Esser, N.M.; Shaver, R.D.; Coblentz, W.K.; Scott, M.P.; Bodnar, A.L.; Schmidt, R.J.; Charley, R.C. Influence of Ensiling Time and Inoculation on Alteration of the Starch-Protein Matrix in High-Moisture Corn. J. Dairy Sci. 2011, 94, 2465–2474. [Google Scholar] [CrossRef] [Green Version]
- de Almeida Araújo, C.; dos Santos, A.P.M.; de Figueiredo Monteiro, C.C.; Lima, D.O.; Torres, A.M.; dos Santos, C.V.S.; dos Santos Monteiro, S.E.; da Silva, J.J. Efeito Do Tempo de Ensilagem Sobre a Composição Química, Perfil Fermentativo e Estabilidade Aeróbia de Silagens de Milho (Zea mays). Divers. J. 2020, 5, 547–561. [Google Scholar] [CrossRef] [Green Version]
- Nutcher, K.; Salacci, R.; Kuber, C.P.; Kuber, R.; Uriarte, M.E.; Bolsen, K.K. Effects of Sealing Time Post-Filling and Sealing Material on Fermentation, Nutritional Quality, and Organic Matter Loss of Whole-Plant Maize Ensiled in a Drive-over Pile. In Proceedings of the XVII International Silage Conference, Piracicaba, SP, Brazil, 1–3 July 2015; pp. 320–321. [Google Scholar]
- Yang, C.-M.J. Proteolysis, Fermentation Efficiency, and in vitro Ruminal Digestion of Peanut Stover Ensiled with Raw or Heated Corn. J. Dairy Sci. 2005, 88, 2903–2910. [Google Scholar] [CrossRef] [Green Version]
- Miyaji, M.; Matsuyama, H.; Nonaka, K. Effect of Ensiling Process of Total Mixed Ration on Fermentation Profile, Nutrient Loss and in Situ Ruminal Degradation Characteristics of Diet. Anim. Sci. J. 2017, 88, 134–139. [Google Scholar] [CrossRef]
- Junges, D.; Morais, G.; Spoto, M.H.F.; Santos, P.S.; Adesogan, A.T.; Nussio, L.G.; Daniel, J.L.P. Short Communication: Influence of Various Proteolytic Sources during Fermentation of Reconstituted Corn Grain Silages. J. Dairy Sci. 2017, 100, 9048–9051. [Google Scholar] [CrossRef] [Green Version]
- Kang, T.S.; Korber, D.R.; Tanaka, T. Influence of Oxygen on NADH Recycling and Oxidative Stress Resistance Systems in Lactobacillus panis PM1. AMB Express 2013, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Maresca, D.; Zotta, T.; Mauriello, G. Adaptation to Aerobic Environment of Lactobacillus johnsonii/gasseri Strains. Front. Microbiol. 2018, 9, 157. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic Metabolism in the Genus Lactobacillus: Impact on Stress Response and Potential Applications in the Food Industry. J. Appl. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, H.; Ran, Q.; Li, H.; Zhang, X. Succession of Microbial Communities of Corn Silage Inoculated with Heterofermentative Lactic Acid Bacteria from Ensiling to Aerobic Exposure. Fermentation 2021, 7, 258. [Google Scholar] [CrossRef]
- Filannino, P.; Di Cagno, R.; Gobbetti, M. Metabolic and Functional Paths of Lactic Acid Bacteria in Plant Foods: Get out of the Labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Amer, S.; Hassanat, F.; Berthiaume, R.; Seguin, P.; Mustafa, A.F. Effects of Water Soluble Carbohydrate Content on Ensiling Characteristics, Chemical Composition and in Vitro Gas Production of Forage Millet and Forage Sorghum Silages. Anim. Feed Sci. Technol. 2012, 177, 23–29. [Google Scholar] [CrossRef]
- Schmidt, P.; Novinski, C.O.; Zopollatto, M. Carbon Absorption in Silages: A Novel Approach in Silage Microbiology. In Proceedings of the XVIII International Silage Conference, Bonn, Germany, 24–26 July 2018; Gerlach, K., Südekum, K.-H., Eds.; The University of Bonn: Bonn, Germany, 2018; pp. 20–21. [Google Scholar]
- Schmidt, P.; Novinski, C.O.; Carneiro, E.W.; Bayer, C. Greenhouse Gas Emissions from Fermentation of Corn Silage. In Proceedings of the XVI International Silage Conference, Hämeenlinna, Finland, 2–4 July 2004; Kuoppala, K., Rinne, M., Vanhatalo, A., Eds.; MTT Agrifood Research Finland, University of Helsinki: Hämeenlinna, Finland, 2012; pp. 448–449. [Google Scholar]
- Puntillo, M.; Gaggiotti, M.; Oteiza, J.M.; Binetti, A.; Massera, A.; Vinderola, G. Potential of Lactic Acid Bacteria Isolated From Different Forages as Silage Inoculants for Improving Fermentation Quality and Aerobic Stability. Front. Microbiol. 2020, 11, 586716. [Google Scholar] [CrossRef]
- Hu, Z.; Chang, J.; Yu, J.; Li, S.; Niu, H. Diversity of Bacterial Community during Ensiling and Subsequent Exposure to Air in Whole-Plant Maize Silage. Asian-Australas. J. Anim. Sci. 2018, 31, 1464–1473. [Google Scholar] [CrossRef]
- Sieuwerts, S.; Bron, P.A.; Smid, E.J. Mutually Stimulating Interactions between Lactic Acid Bacteria and Saccharomyces cerevisiae in Sourdough Fermentation. LWT 2018, 90, 201–206. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Bacterial Dynamics of Wheat Silage. Front. Microbiol. 2019, 10, 1532. [Google Scholar] [CrossRef]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial Communities and Natural Fermentation of Corn Silages Prepared with Farm Bunker-Silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]


{kind=link}
| Item 2 | Additive (A) | Delay Period (T) 1 | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 | 90 | 150 | 210 | T | A | T × A | ||||
| DM | Control | 28.61 a | 29.16 bA | 28.51 b | 28.51 bA | 28.70 A | 0.0370 | 0.0016 | <0.001 | <0.001 |
| Inoculated | 28.78 a | 28.25 bB | 28.32 b | 28.16 bB | 28.38 B | |||||
| Mean | 28.70 a | 28.71 a | 28.41 b | 28.33 b | ||||||
| pH | Control | 3.77 bc | 3.88 aA | 3.85 ab | 3.68 cB | 3.79 | 0.0085 | 0.0019 | 0.3756 | 0.001 |
| Inoculated | 3.83 | 3.81 B | 3.79 | 3.80 A | 3.81 | |||||
| Mean | 3.80 ab | 3.85 a | 3.82 a | 3.74 b | ||||||
| CP | Control | 8.08 a | 8.11 a | 8.13 a | 7.66 bB | 8.21 A | 0.0267 | 0.0120 | <0.001 | 0.0371 |
| Inoculated | 8.17 | 8.21 | 8.26 | 8.19 A | 7.99 B | |||||
| Mean | 8.12 ab | 8.16 a | 8.20 a | 7.93 b | ||||||
| NDF | Control | 47.94 bc | 46.64 cB | 49.27 b | 52.64 aA | 49.12 | 0.2307 | <0.001 | 0.7963 | 0.0047 |
| Inoculated | 46.99 b | 49.54 aA | 48.97 ab | 50.48 aB | 49.00 | |||||
| Mean | 47.47 b | 48.09 b | 49.13 b | 51.56 a | ||||||
| ADF | Control | 27.78 b | 27.67 bB | 28.88 b | 30.97 a | 28.83 | 0.1518 | <0.001 | 0.5470 | 0.0138 |
| Inoculated | 27.20 b | 29.25 aA | 28.51 ab | 29.59 a | 28.64 | |||||
| Mean | 27.49 c | 28.46 bc | 28.70 b | 30.28 a | ||||||
| Ash | Control | 3.07 | 3.36 | 3.28 | 3.12 | 3.21 A | 0.0441 | 0.6546 | 0.0336 | 0.4386 |
| Inoculated | 2.97 | 2.97 | 2.98 | 3.10 | 3.01 B | |||||
| Mean | 3.02 | 3.17 | 3.13 | 3.11 | ||||||
| Lignin | Control | 3.90 | 3.92 | 4.05 | 4.32 | 4.05 | 0.0244 | <0.001 | 0.4540 | 0.1248 |
| Inoculated | 3.86 | 4.15 | 4.12 | 4.22 | 4.09 | |||||
| Mean | 3.88 c | 4.04 bc | 4.09 ab | 4.27 a | ||||||
| Starch | Control | 20.83 abB | 22.53 aA | 19.00 bB | 15.85 cB | 19.55 B | 0.2089 | <0.001 | 0.0320 | <0.001 |
| Inoculated | 22.68 aA | 20.08 bB | 20.67 abA | 18.59 bA | 20.51 A | |||||
| Mean | 21.76 a | 21.30 ab | 19.83 b | 17.22 c | ||||||
| Item 2 | Additive (A) | Delay Period (T) 1 | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 | 90 | 150 | 210 | T | A | T × A | ||||
| Lactic acid | Control | 9.29 | 9.29 | 8.51 | 7.48 | 8.65 | 0.1316 | 0.0046 | 0.0613 | 0.5445 |
| Inoculated | 8.66 | 8.22 | 8.06 | 7.55 | 8.13 | |||||
| Mean | 8.98 a | 8.76 a | 8.29 ab | 7.52 b | ||||||
| Acetic acid | Control | 1.68 | 1.64 | 1.58 | 1.47 | 1.59 B | 0.0360 | 0.1389 | <0.001 | 0.9815 |
| Inoculated | 2.62 | 2.54 | 2.47 | 2.32 | 2.49 A | |||||
| Mean | 2.15 | 2.09 | 2.03 | 1.90 | ||||||
| Ethanol | Control | 0.26 | 0.28 | 0.29 | 0.29 | 0.28 B | 0.0244 | 0.6339 | <0.001 | 0.3461 |
| Inoculated | 0.65 | 0.59 | 0.47 | 0.47 | 0.55 A | |||||
| Mean | 0.45 | 0.44 | 0.38 | 0.38 | ||||||
| TDML | Control | 3.05 a | 1.18 bB | 3.28 a | 3.36 aB | 2.71 B | 0.1343 | 0.0123 | <0.001 | 0.0014 |
| Inoculated | 3.08 b | 4.41 abA | 4.15 ab | 4.73 aA | 4.09 A | |||||
| Mean | 3.06 ab | 2.79 b | 3.71 ab | 4.04 a | ||||||
| GProd (L−1 kg−1 DM) | Control | 5.45 | 5.70 | 5.68 | 6.17 | 5.75 B | 0.0565 | 0.0174 | 0.0241 | 0.6664 |
| Inoculated | 5.97 | 5.91 | 5.89 | 6.32 | 6.02 A | |||||
| Mean | 5.71 b | 5.81 ab | 5.78 b | 6.25 a | ||||||
| Eloss | Control | 2.49 | 2.46 | 1.71 | 1.91 | 2.14 A | 0.1271 | 0.1862 | 0.0029 | 0.2638 |
| Inoculated | 0.76 | 1.91 | 1.14 | 1.40 | 1.30 B | |||||
| Mean | 1.63 | 2.19 | 1.43 | 1.65 | ||||||
| Gloss | Control | 2.81 a | 0.98 bB | 3.13 a | 3.17 aB | 2.52 B | 0.1320 | 0.0073 | <0.001 | 0.0018 |
| Inoculated | 3.00 b | 4.22 abA | 4.03 ab | 4.63 aA | 3.97 A | |||||
| Mean | 2.90 ab | 2.60 b | 3.58 ab | 3.90 a | ||||||
| Item 2 | Additive (A) | Delay Period (T) 1 | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 | 90 | 150 | 210 | T | A | T × A | ||||
| LAB | Control | 7.68 | 7.59 | >8.0 | >8.0 | >8.0 | 0.57 | 0.2722 | 0.0614 | - |
| Inoculated | >8.0 | >8.0 | >8.0 | - | >8.0 | |||||
| Mean | >8.0 | >8.0 | >8.0 | >8.0 | ||||||
| Yeasts | Control | 4.86 | 5.15 | 5.38 | 4.52 | 4.98 | 0.09 | 0.5950 | 0.1049 | 0.7352 |
| Inoculated | 3.30 | 3.78 | 3.90 | 3.78 | 3.69 | |||||
| Mean | 4.08 | 4.47 | 4.64 | 4.15 | ||||||
| Molds | Control | 3.78 | 3.90 | 3.30 | 3.48 | 3.62 | 0.03 | 0.8547 | 0.6211 | 0.6211 |
| Inoculated | 3.78 | 3.30 | 3.30 | 3.90 | 3.57 | |||||
| Mean | 3.78 | 3.60 | 3.30 | 3.69 | ||||||
| Item 2 | Additive (A) | Delay Period (T) 1 | Mean | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 30 | 90 | 150 | 210 | T | A | T × A | ||||
| AS | Control | 62.03 cB | 80.35 bB | 90.20 aB | 89.13 aB | 80.45 B | 0.1662 | <0.001 | <0.001 | <0.001 |
| Inoculated | 213.00 A | 213.00 A | 213.00 A | 213.00 A | 213.00 A | |||||
| Mean | 137.51 c | 146.68 b | 151.65 a | 151.07 a | ||||||
| TDMLAS | Control | 7.84 abA | 8.71 aA | 6.19 b | 6.56 ab | 7.33 A | 0.2363 | 0.2530 | 0.0011 | 0.0433 |
| Inoculated | 5.47 B | 5.46 B | 6.71 | 4.75 | 5.60 B | |||||
| Mean | 6.66 | 7.09 | 6.45 | 5.65 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Melo, N.N.; Carvalho-Estrada, P.d.A.; Tavares, Q.G.; Pereira, L.d.M.; Delai Vigne, G.L.; Camargo Rezende, D.M.L.; Schmidt, P. The Effects of Short-Time Delayed Sealing on Fermentation, Aerobic Stability and Chemical Composition on Maize Silages. Agronomy 2023, 13, 223. https://doi.org/10.3390/agronomy13010223
de Melo NN, Carvalho-Estrada PdA, Tavares QG, Pereira LdM, Delai Vigne GL, Camargo Rezende DML, Schmidt P. The Effects of Short-Time Delayed Sealing on Fermentation, Aerobic Stability and Chemical Composition on Maize Silages. Agronomy. 2023; 13(1):223. https://doi.org/10.3390/agronomy13010223
Chicago/Turabian Stylede Melo, Natália Nunes, Paula de Almeida Carvalho-Estrada, Queila Gouveia Tavares, Lucelia de Moura Pereira, Gabriela Leticia Delai Vigne, Douglas Messias Lamounier Camargo Rezende, and Patrick Schmidt. 2023. "The Effects of Short-Time Delayed Sealing on Fermentation, Aerobic Stability and Chemical Composition on Maize Silages" Agronomy 13, no. 1: 223. https://doi.org/10.3390/agronomy13010223
APA Stylede Melo, N. N., Carvalho-Estrada, P. d. A., Tavares, Q. G., Pereira, L. d. M., Delai Vigne, G. L., Camargo Rezende, D. M. L., & Schmidt, P. (2023). The Effects of Short-Time Delayed Sealing on Fermentation, Aerobic Stability and Chemical Composition on Maize Silages. Agronomy, 13(1), 223. https://doi.org/10.3390/agronomy13010223