
Exogenous Application of Zinc Oxide Nanoparticles Improved Antioxidants, Photosynthetic, and Yield Traits in Salt-Stressed Maize


Abstract
:1. Introduction
2. Materials and Methods
2.1. Nursery Preparation
2.2. Transplanting and Growth Conditions
2.3. Treatments and Experimental Design
2.4. Plant Growth Parameters
2.4.1. Maximum Plant Height, Stem Diameter, and Plant Fresh and Dry Weights
2.4.2. Plant Growth Rate (PGR) and Leaf Growth Rate (LGR)
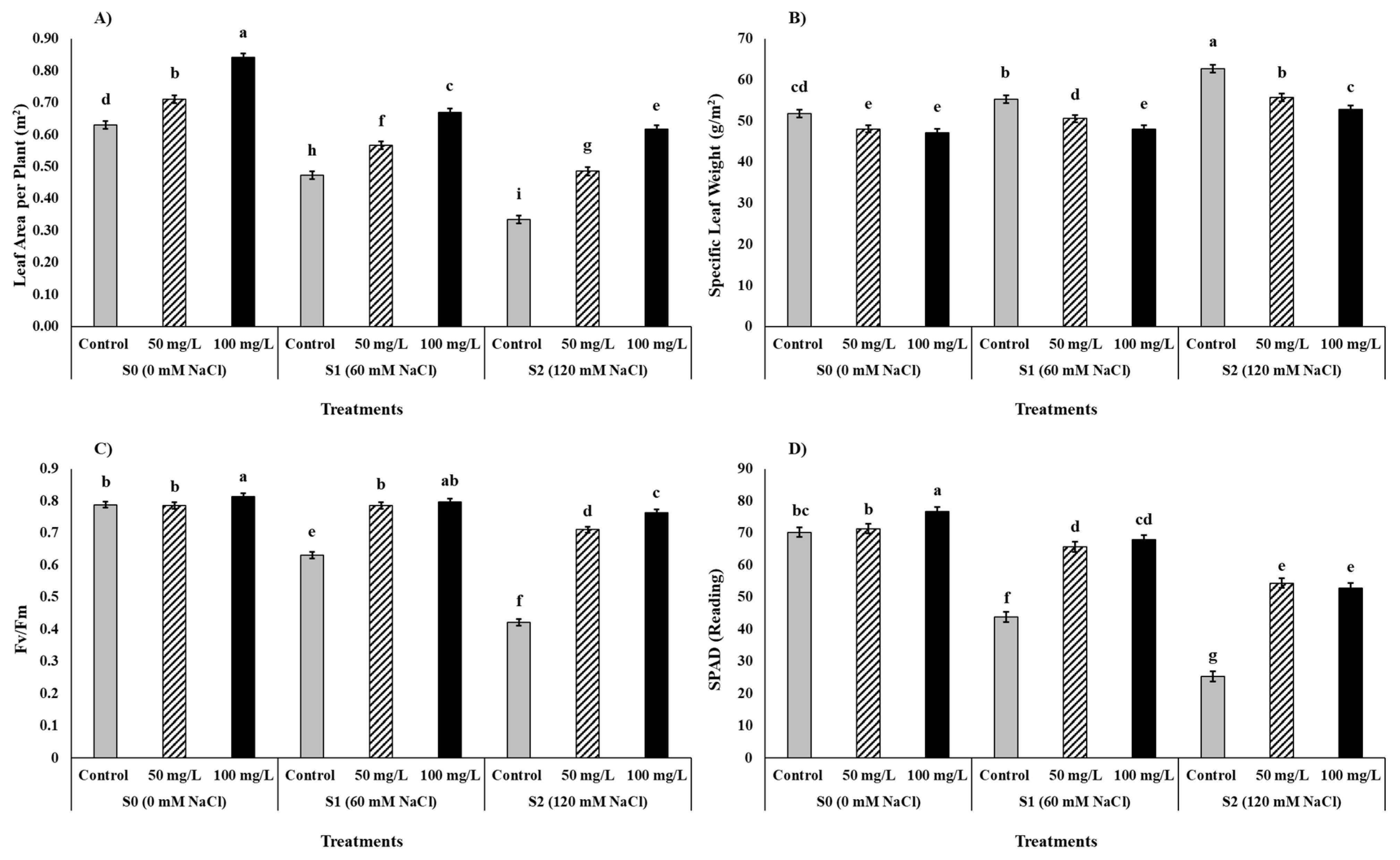
2.4.3. Leaf Area, Relative Water Contents (RWC), and Specific Leaf Weight (SLW)
2.4.4. Chlorophyll Index (SPAD) Maximum Quantum Yield of PS II [Fv/Fm]
2.4.5. Photosynthetic Performance and Gas Exchange
2.4.6. Determination of Na+, K+, Na+/K+, and Zn2+ Concentrations
2.4.7. Proline, Total Phenolic Contents, and Antioxidant
2.4.8. Grain Yield and Yield-Related Attributes
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
6. Recommendations
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hopmans, J.W.; Qureshi, A.S.; Kisekka, I.; Munns, R.; Grattan, S.R.; Rengasamy, P.; Ben-Gal, A.; Assouline, S.; Javaux, M.; Minhas, P.S.; et al. Critical knowledge gaps and research priorities in global soil salinity. Adv. Agron. 2021, 169, 1–191. [Google Scholar] [CrossRef]
- Mukhopadhyay, R.; Sarkar, B.; Jat, H.S.; Sharma, P.C.; Bolan, N.S. Soil salinity under climate change: Challenges for sustainable agriculture and food security. J. Environ. Manag. 2021, 280, 111736. [Google Scholar] [CrossRef] [PubMed]
- Negacz, K.; Vellinga, P.; Barrett-Lennard, E.; Choukr-Allah, R.; Elzenga, T. Future of Sustainable Agriculture in Saline Environments; Taylor & Francis: Abingdon, UK, 2021; p. 541. [Google Scholar] [CrossRef]
- Singh, A. Soil salinization management for sustainable development: A review. J. Environ. Manag. 2021, 277, 111383. [Google Scholar] [CrossRef] [PubMed]
- Eswar, D.; Karuppusamy, R.; Chellamuthu, S. Drivers of soil salinity and their correlation with climate change. Curr. Opin. Environ. Sustain. 2021, 50, 310–318. [Google Scholar] [CrossRef]
- Aksoy, S.; Yildirim, A.; Gorji, T.; Hamzehpour, N.; Tanik, A.; Sertel, E. Assessing the performance of machine learning algorithms for soil salinity mapping in Google Earth Engine platform using Sentinel-2A and Landsat-8 OLI data. Adv. Space Res. 2022, 69, 1072–1086. [Google Scholar] [CrossRef]
- Mokrani, S.; Nabti, E.H.; Cruz, C. Recent trends in microbial approaches for soil desalination. Appl. Sci. 2022, 12, 3586. [Google Scholar] [CrossRef]
- Whitney, K.; Scudiero, E.; El-Askary, H.M.; Skaggs, T.H.; Allali, M.; Corwin, D.L. Validating the use of MODIS time series for salinity assessment over agricultural soils in California, USA. Ecol. Indic. 2018, 93, 889–898. [Google Scholar] [CrossRef]
- Talat, N. Alleviation of soil salinization and the management of saline soils, climate change, and soil interactions. In Climate Change and Soil Interactions; Elsevier: Amsterdam, The Netherlands, 2020; pp. 305–329. [Google Scholar] [CrossRef]
- Ramos, T.B.; Castanheira, N.; Oliveira, A.R.; Paz, A.M.; Darouich, H.; Simionesei, L.; Farzamian, M.; Gonçalves, M.C. Soil salinity assessment using vegetation indices derived from Sentinel-2 multispectral data. application to Lezíria Grande, Portugal. Agric. Water Manag. 2020, 241, 106387. [Google Scholar] [CrossRef]
- Ali, Y.; Aslam, Z.; Ashraf, M.Y.; Tahir, G.R. Effect of salinity on chlorophyll concentration, leaf area, yield and yield components of rice genotypes grown under saline environment. Int. J. Environ. Sci. Technol. 2004, 1, 221–225. [Google Scholar] [CrossRef]
- Singh, A. Soil salinity: A global threat to sustainable development. Soil Use Manag. 2022, 38, 39–67. [Google Scholar] [CrossRef]
- Abideen, Z.; Koyro, H.W.; Huchzermeyer, B.; Ahmed, M.Z.; Zulfiqar, F.; Egan, T.; Khan, M.A. Phragmites karka plants adopt different strategies to regulate photosynthesis and ion flux in saline and water deficit conditions. Plant Biosyst. 2020, 155, 524–534. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, C.X.; Li, X.; Liu, A.; Chen, S.; Zhou, J. Overexpression of tomato RING E3 ubiquitin ligase gene SlRING1 confers cadmium tolerance by attenuating cadmium accumulation and oxidative stress. Physiol. Plant. 2020, 173, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Hajihashemi, S.; Skalicky, M.; Brestic, M.; Pavla, V. Cross-talk between nitric oxide, hydrogen peroxide and calcium in salt-stressed Chenopodium quinoa Willd. At seed germination stage. Plant Physiol. Biochem. 2020, 154, 657–664. [Google Scholar] [CrossRef]
- Orooji, Y.; Ghanbari, M.; Amiri, O.; Salavati-Niasari, M. Facile fabrication of silver iodide/graphitic carbon nitride nanocomposites by notable photo catalytic performance through sunlight and antimicrobial activity. J. Hazard. Mater. 2020, 389, 122079. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Maleh, H.; Ayati, A.; Davoodi, R.; Tanhaei, B.; Karimi, F.; Malekmohammadi, S.; Orooji, Y.; Fu, L.; Sillanpaa, M. Recent advances in using of chitosan-based adsorbents for removal of pharmaceutical contaminants: A review. J. Clean. Prod. 2021, 291, 125–880. [Google Scholar] [CrossRef]
- Salam, A.; Afridi, M.S.; Javed, M.A.; Saleem, A.; Hafeez, A.; Khan, A.R.; Zeeshan, M.; Ali, B.; Azhar, W.; Ulhassan, Z.; et al. Nano-priming against abiotic stress: A way forward towards sustainable agriculture. Sustainability 2022, 14, 14880. [Google Scholar] [CrossRef]
- Shahid, M.; Naeem-Ullah, U.; Khan, W.; Saeed, D.S.; Razzaq, K. Application of nanotechnology for insect pests management: A review. J. Innov. Sci. 2021, 7, 28–39. [Google Scholar] [CrossRef]
- Sharma, S.; Singh, S.S.; Bahuguna, A.; Yadav, B.; Barthwal, A.; Nandan, R.; Khatana, R.; Pandey, A.; Thakur, R.; Singh, H. Nanotechnology: An efficient tool in plant nutrition management. In Ecosystem Services: Types, Management and Benefits; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2022. [Google Scholar]
- Singh, A.; Singh, N.Á.; Afzal, S.; Singh, T.; Hussain, I. Zinc oxide nanoparticles: A review of their biological synthesis, antimicrobial activity, uptake, translocation and biotransformation in plants. J. Mater. Sci. 2018, 53, 185–201. [Google Scholar] [CrossRef]
- Mali, S.C.; Raj, S.; Trivedi, R. Nanotechnology a novel approach to enhance crop productivity. Biochem. Biophys. Rep. 2020, 24, 100821. [Google Scholar] [CrossRef]
- Pooja, P.; Nandwal, A.S.; Chand, M.; Pal, A.; Kumari, A.; Rani, B.; Goel, V.; Kulshreshtha, N. Soil Moisture Deficit Induced Changes in Antioxidative Defense Mechanism of Sugarcane (Saccharum officinarum) Varieties Differing in Maturity. 2020. Available online: https://agris.fao.org/agris-search/search.do?recordID=IN2022004578 (accessed on 19 May 2023).
- Bana, R.C.; Gupta, A.K.; Bana, R.S.; Shivay, Y.S.; Bamboriya, S.D.; Thakur, N.P.; Puniya, R.; Gupta, M.; Jakhar, S.R.; Kailash Chaudhary, R.S.; et al. Zinc-coated urea for enhanced zinc biofortification, nitrogen use efficiency and yield of basmati rice under typic fluvents. Sustainability 2021, 14, 104. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Ashraf, M. Nanoparticles potentially mediate salt stress tolerance in plants. Plant Physiol. Biochem. 2021, 160, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Lopez, J.I.; Nino-Medina, G.; Olivares-Saenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vazquez-Alvarado, R.; Rodriguez-Salinas, P.A.; Zavala-Garcia, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef]
- Elamawi, R.M.; Bassiouni, S.M.; Elkhoby, W.M.; Zayed, B.A. Effect of zinc oxide nanoparticles on brown spot disease and rice productivity under saline soil. J. Plant Prot. Pathol. 2016, 7, 171–181. [Google Scholar] [CrossRef]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, zinc nanoparticles and plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, F.; Azhar, M.; Anwar-ul-Haq, M.; Sabir, M.; Samreen, T.; Tufail, A.; Awan, H.U.M.; Juan, W. Comparative response of two rice (Oryza sativa L.) cultivars to applied zinc and manganese for mitigation of salt stress. J. Soil Sci. Plant Nutr. 2020, 20, 2059–2072. [Google Scholar] [CrossRef]
- Tolay, I. The impact of different Zinc (Zn) levels on growth and nutrient uptake of Basil (Ocimum basilicum L.) grown under salinity stress. PLoS ONE 2021, 16, 0246493. [Google Scholar] [CrossRef]
- Cicek, S.; Nadaroglu, H. The use of nanotechnology in the agriculture. Adv. Nano Res. 2015, 3, 207–223. [Google Scholar] [CrossRef]
- Shang, Y.; Hasan, M.K.; Ahammed, G.J.; Li, M.; Yin, H.; Zhou, J. Applications of nanotechnology in plant growth and crop protection: A review. Molecules 2019, 24, 2558. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Navarro, M.; Ashraf, M.; Akram, N.A.; Munn’e-Bosch, S. Nanofertilizer use for sustainable agriculture: Advantages and limitations. Plant Sci. 2019, 289, 110270. [Google Scholar] [CrossRef]
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global maize production, consumption and trade: Trends and R&D implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar] [CrossRef]
- Erenstein, O. The evolving maize sector in Asia: Challenges and opportunities. J. New Seeds 2010, 11, 1–15. [Google Scholar] [CrossRef]
- Grote, U.; Fasse, A.; Nguyen, T.T.; Erenstein, O. Food security and the dynamics of wheat and maize value chains in Africa and Asia. Front. Sustain. Food Syst. 2021, 4, 617009. [Google Scholar] [CrossRef]
- Bhusal, B.; Poudel, M.R.; Rishav, P.; Regmi, R.; Neupane, P.; Bhattarai, K.; Maharjan, B.; Bigyan, K.C.; Acharya, S. A review on abiotic stress resistance in maize (Zea mays L.): Effects, resistance mechanisms and management. J. Biol. Today’s World 2021, 10, 1–3. [Google Scholar]
- Sabagh, A.E.; Çiğ, F.; Seydoşoğlu, S.; Battaglia, M.L.; Javed, T.; Iqbal, M.A.; Awad, M. Salinity stress in maize: Effects of stress and recent developments of tolerance for improvement. Cereal Grains 2021, 1, 213. [Google Scholar]
- Prasanna, B.M.; Cairns, J.E.; Zaidi, P.H.; Beyene, Y.; Makumbi, D.; Gowda, M.; Magorokosho, C.; Zaman-Allah, M.; Olsen, M.; Das, A.; et al. Beat the stress: Breeding for climate resilience in maize for the tropical rainfed environments. Theor. Appl. Genet. 2021, 134, 1729–1752. [Google Scholar] [CrossRef]
- Basit, F.; Chen, M.; Ahmed, T.; Shahid, M.; Noman, M.; Liu, J.; An, J.; Hashem, A.; Fahad Al-Arjani, A.B.; Alqarawi, A.A.; et al. Seed priming with brassinosteroids alleviates chromium stress in rice cultivars via improving ROS metabolism and antioxidant defense response at biochemical and molecular levels. Antioxidants 2021, 10, 1089. [Google Scholar] [CrossRef]
- Wolf, B. The comprehensive system of leaf analysis and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 3, 1035–1059. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.A.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Tawaha, K.; Alali, F.Q.; Gharaibeh, M.; Mohammad, M.; El-Elimat, T. Antioxidant activity and total phenolic content of selected Jordanian plant species. Food Chem. 2017, 104, 1372–1378. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Wang, M.; Bi, D. Selenium modulates the activities of antioxidant enzymes, osmotic homeostasis and promotes the growth of sorrel seedlings under salt stress. Plant Growth Regul. 2005, 45, 155–163. [Google Scholar] [CrossRef]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity stress in potato: Understanding physiological, biochemical and molecular responses. Life 2021, 11, 545. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Hossain, A.; Barutçular, C.; Iqbal, M.A.; Islam, M.S.; Fahad, S.; Sytar, O.; Çiğ, F.; Meena, R.S.; Erman, M. Consequences of salinity stress on the quality of crops and its mitigation strategies for sustainable crop production: An outlook of arid and semi-arid regions. In Environment, Climate, Plant and Vegetation Growth; Springer: Berlin/Heidelberg, Germany, 2020; pp. 503–533. [Google Scholar]
- Gerona, M.E.B.; Deocampo, M.P.; Egdane, J.A.; Ismail, A.M.; Dionisio-Sese, M.L. Physiological responses of contrasting rice genotypes to salt stress at reproductive stage. Rice Sci. 2019, 26, 207–219. [Google Scholar] [CrossRef]
- Yang, Z.; Li, J.L.; Liu, L.N.; Xie, Q.; Sui, N. Photosynthetic regulation under salt stress and salt-tolerance mechanism of sweet sorghum. Front. Plant Sci. 2020, 10, 1722. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, L.; Barkla, B.J. Membrane lipid remodeling in response to salinity. Int. J. Mol. Sci. 2019, 20, 4264. [Google Scholar] [CrossRef]
- Shahzad, R.; Harlina, P.W.; Ewas, M.; Zhenyuan, P.; Nie, X.; Gallego, P.P.; Ullah Khan, S.; Nishawy, E.; Khan, A.H.; Jia, H. Foliar applied 24-epibrassinolide alleviates salt stress in rice (Oryza sativa L.) by suppression of ABA levels and upregulation of secondary metabolites. J. Plant Interact. 2021, 16, 533–549. [Google Scholar] [CrossRef]
- Shabala, S.; Shabala, S.; Cuin, T.A.; Pang, J.; Percey, W.; Chen, Z.; Conn, S.; Eing, C.; Wegner, L.H. Xylem ionic relations and salinity tolerance in barley. Plant J. 2010, 61, 839–853. [Google Scholar] [CrossRef]
- Yun, P.; Xu, L.; Wang, S.S.; Shabala, L.; Shabala, S.; Zhang, W.Y. Piriformospora indica improves salinity stress tolerance in Zea mays L. plants by regulating Na+ and K+ loading in root and allocating K+ in shoot. Plant Growth Regul. 2018, 86, 323–331. [Google Scholar] [CrossRef]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- Zafar, S.; Perveen, S.; Kamran Khan, M.; Shaheen, M.R.; Hussain, R.; Sarwar, N.; Rashid, S.; Nafees, M.; Farid, G.; Alamri, S.; et al. Effect of zinc nanoparticles seed priming and foliar application on the growth and physio-biochemical indices of spinach (Spinacia oleracea L.) under salt stress. PLoS ONE 2022, 17, e0263194. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Mohamed, I.A.; Khatab, A.; Sherif, A.; Wang, Z.; Salah, A.; Nishawy, E.; Ayaad, M.; Kuai, J.; et al. Modulation of salinity impact on early seedling stage via nano-priming application of zinc oxide on rapeseed (Brassica napus L.). Plant Physiol. Biochem. 2021, 166, 376–392. [Google Scholar] [CrossRef]
- Miller, G.A.D.; Suzuki, N.; Ciftci-Yilmaz, S.U.L.T.A.N.; Mittler, R.O.N. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; Fu, Z.; Zhang, L.; Zhang, Z.; Zhang, J. The positive effect of different 24-epiBL pretreatments on salinity tolerance in Robinia pseudoacacia L. seedlings. Forests 2018, 10, 4. [Google Scholar] [CrossRef]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Cho, M.C.; Yang, E.Y.; Lee, J.G. Response to salt stress in lettuce: Changes in chlorophyll fluorescence parameters, phytochemical contents, and antioxidant activities. Agronomy 2020, 10, 1627. [Google Scholar] [CrossRef]
- Liu, C.; Mao, B.; Yuan, D.; Chu, C.; Duan, M. Salt tolerance in rice: Physiological responses and molecular mechanisms. Crop J. 2022, 10, 13–25. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Razzaq, A.; Ali, A.; Safdar, L.B.; Zafar, M.M.; Rui, Y.; Shakeel, A.; Shaukat, A.; Ashraf, M.; Gong, W.; Yuan, Y. Salt stress induces physiochemical alterations in rice grain composition and quality. J. Food Sci. 2020, 85, 14–20. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; Zia ur Rehman, M.; Waris, A.A. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Faizan, M.; Hayat, S.; Pichtel, J. Effects of zinc oxide nanoparticles on crop plants: A perspective analysis. In Sustainable Agriculture Reviews 41; Springer: Cham, Switzerland, 2020; pp. 83–99. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Hessini, K.; Yu, F.; Ahmad, P. Zinc oxide nanoparticles alleviates the adverse effects of cadmium stress on Oryza sativa via modulation of the photosynthesis and antioxidant defense system. Ecotoxicol. Environ. Saf. 2021, 220, 112401. [Google Scholar] [CrossRef] [PubMed]
- Abou-Zeid, H.M.; Ismail, G.S.M.; Abdel-Latif, S.A. Influence of seed priming with ZnO nanoparticles on the salt-induced damages in wheat (Triticum aestivum L.) plants. J. Plant Nutr. 2021, 44, 629–643. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651. [Google Scholar] [CrossRef]
- Babitha, K.C.; Vemanna, R.S.; Nataraja, K.N.; Udayakumar, M. Overexpression of EcbHLH57 transcription factor from Eleusine coracana L. in tobacco confers tolerance to salt, oxidative and drought stress. PLoS ONE 2015, 10, e0137098. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef]
- Singh, A.; Roychoudhury, A. Gene regulation at transcriptional and post-transcriptional levels to combat salt stress in plants. Physiol. Plant. 2021, 173, 1556–1572. [Google Scholar] [CrossRef]
- Van Raamsdonk, J.M.; Hekimi, S. Superoxide dismutase is dispensable for normal animal lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 5785–5790. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Tanveer, M. Role of 24-Epibrassinolide in inducing thermo-tolerance in plants. J. Plant Growth Regul. 2019, 38, 945–955. [Google Scholar] [CrossRef]
- Singh, R.P.; Handa, R.; Manchanda, G. Nanoparticles in sustainable agriculture: An emerging opportunity. J. Control. Release 2021, 329, 1234–1248. [Google Scholar] [CrossRef]
- Adrees, M.; Khan, Z.S.; Hafeez, M.; Rizwan, M.; Hussain, K.; Asrar, M.; Alyemeni, M.N.; Wijaya, L.; Ali, S. Foliar exposure of zinc oxide nanoparticles improved the growth of wheat (Triticum aestivum L.) and decreased cadmium concentration in grains under simultaneous Cd and water deficient stress. Ecotoxicol. Environ. Saf. 2021, 208, 111627. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Chen, C.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P.; Yu, F. Zinc oxide nanoparticles (ZnO-NPs) induce salt tolerance by improving the antioxidant system and photosynthetic machinery in tomato. Plant Physiol. Biochem. 2021, 161, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [Google Scholar] [CrossRef] [PubMed]
- Aqeel, U.; Aftab, T.; Khan, M.M.A.; Naeem, M.; Khan, M.N. A comprehensive review of impacts of diverse nanoparticles on growth, development and physiological adjustments in plants under changing environment. Chemosphere 2022, 291, 132672. [Google Scholar] [CrossRef]
- Ali, B.; Saleem, M.H.; Ali, S.; Shahid, M.; Sagir, M.; Tahir, M.B.; Qureshi, K.A.; Jaremko, M.; Selim, S.; Hussain, A.; et al. Mitigation of salinity stress in barley genotypes with variable salt tolerance by application of zinc oxide nanoparticles. Front. Plant Sci. 2022, 13, 973782. [Google Scholar] [CrossRef]
- Hussein, M.M.; Abou-Baker, N.H. The contribution of nano-zinc to alleviate salinity stress on cotton plants. R. Soc. Open Sci. 2018, 5, 171809. [Google Scholar] [CrossRef]
- Qiao, T.; Zhao, Y.; Zhong, D.B.; Yu, X. Hydrogen peroxide and salinity stress act synergistically to enhance lipids production in microalga by regulating reactive oxygen species and calcium. Algal Res. 2021, 53, 102017. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Ahmad, A.; Battaglia, M.L.; Bilal, H.M.; Alhammad, B.A.; Khan, N. Zinc oxide nanoparticles: A unique salinity mitigator with the potential to increase future crop production. S. Afr. J. Bot. 2023, 159, 208–218. [Google Scholar] [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Abdelaziz, M.E.; Kim, D.; Ali, S.; Fedoroff, N.V.; Al-Babili, S. The endophytic fungus Piriformospora indica enhances Arabidopsis thaliana growth and modulates Na+/K+ homeostasis under salt stress conditions. Plant Sci. 2017, 263, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Heikal, Y.M.; El-Esawi, M.A.; El-Ballat, E.M.; Abdel-Aziz, H.M. Applications of nanoparticles for mitigating salinity and drought stress in plants: An overview on the physiological, biochemical and molecular genetic aspects. N. Z. J. Crop Hortic. Sci. 2022, 51, 297–327. [Google Scholar] [CrossRef]
- Noohpisheh, Z.; Amiri, H.; Mohammadi, A.; Farhadi, S. Effect of the foliar application of zinc oxide nanoparticles on some biochemical and physiological parameters of Trigonella foenum-graecum under salinity stress. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 155, 267–280. [Google Scholar] [CrossRef]
- Dey, A.; Somaiah, S. Green synthesis and characterization of zinc oxide nanoparticles using leaf extract of Thryallis glauca (Cav.) Kuntze and their role as antioxidant and antibacterial. Microsc. Res. Tech. 2022, 85, 2835–2847. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Selwey, W.A.; Ibrahim, A.A.; Shady, M.; Alsadon, A.A. Foliar applications of ZnO and SiO2 nanoparticles mitigate water deficit and enhance potato yield and quality traits. Agronomy 2023, 13, 466. [Google Scholar] [CrossRef]
- Das, P.; Nutan, K.K.; Singla-Pareek, S.L.; Pareek, A. Understanding salinity responses and adopting ‘omics-based’approaches to generate salinity tolerant cultivars of rice. Front. Plant Sci. 2015, 6, 712. [Google Scholar] [CrossRef]
- Wegner, L.H.; Stefano, G.; Shabala, L.; Rossi, M.; Mancuso, S.; Shabala, S. Sequential depolarization of root cortical and stelar cells induced by an acute salt shock–implications for Na+ and K+ transport into xylem vessels. Plant Cell Environ. 2011, 34, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Kumawat, K.C.; Nagpal, S.; Sharma, P. Potential of plant growth-promoting rhizobacteria-plant interactions in mitigating salt stress for sustainable agriculture: A review. Pedosphere 2022, 32, 223–245. [Google Scholar] [CrossRef]
- Costan, A.; Stamatakis, A.; Chrysargyris, A.; Petropoulos, S.A.; Tzortzakis, N. Interactive effects of salinity and silicon application on Solanum lycopersicum growth, physiology and shelf-life of fruit produced hydroponically. J. Sci. Food Agric. 2020, 100, 732–743. [Google Scholar] [CrossRef] [PubMed]
- Loudari, A.; Benadis, C.; Naciri, R.; Soulaimani, A.; Zeroual, Y.; Gharous, M.E.; Kalaji, H.M.; Oukarroum, A. Salt stress affects mineral nutrition in shoots and roots and chlorophyll a fluorescence of tomato plants grown in hydroponic culture. J. Plant Interact. 2020, 15, 398–405. [Google Scholar] [CrossRef]
- Tola, E.; Al-Gaadi, K.A.; Madugundu, R.; Patil, V.C.; Sygrimis, N. Impact of water salinity levels on the spectral behavior and yield of tomatoes in hydroponics. J. King Saud Univ. Sci. 2023, 35, 102515. [Google Scholar] [CrossRef]
- Harizanova, A.; Koleva-Valkova, L. Effect of silicon on photosynthetic rate and the chlorophyll fluorescence parameters at hydroponically grown cucumber plants under salinity stress. J. Cent. Eur. Agric. 2019, 20, 953–960. [Google Scholar] [CrossRef]
- Moncada, A.; Vetrano, F.; Miceli, A. Alleviation of salt stress by plant growth-promoting bacteria in hydroponic leaf lettuce. Agronomy 2020, 10, 1523. [Google Scholar] [CrossRef]
- Kaloterakis, N.; van Delden, S.H.; Hartley, S.; De Deyn, G.B. Silicon application and plant growth promoting rhizobacteria consisting of six pure Bacillus species alleviate salinity stress in cucumber (Cucumis sativus L.). Sci. Hortic. 2021, 288, 110383. [Google Scholar] [CrossRef]
- Sneath, P.H.; Sokal, R.R. Numerical Taxonomy; Freeman: San Francisco, CA, USA, 1973. [Google Scholar]
- Afzal, M.; Alghamdi, S.S.; Migdadi, H.H.; El-Harty, E.; Al-Faifi, S.A. Agronomical and physiological responses of faba bean genotypes to salt stress. Agriculture 2022, 12, 235. [Google Scholar] [CrossRef]
- Saed-Moucheshi, A.; Pessarakli, M.; Heidari, B. Comparing relationships among yield and its related traits in mycorrhizal and nonmycorrhizal inoculated wheat cultivars under different water regimes using multivariate statistics. Int. J. Agron. 2013, 2013, 682781. [Google Scholar] [CrossRef]
- Otie, V.; Ibrahim, A.; Udo, I.; Kashiwagi, J.; Matsuura, A.; Shao, Y.; Itam, M.; An, P.; Eneji, A.E. Foliarly applied 24-Epibrassinolide modulates the electrical conductivity of the saturated rhizospheric soil extracts of soybean under salinity stress. Plants 2022, 11, 2330. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments/Parameters | Maximum Plant Height (cm) | Stem Diameter (mm) | Fresh Weight (g/Plant) | Dry Weight (g/Plant) | RWC (%) | |
|---|---|---|---|---|---|---|
| Salinity | ||||||
| S0 (0 mM NaCl) | 249.67 A | 18.31 A | 526.23 A | 117.06 A | 82.36 A | |
| S1 (60 mM NaCl) | 223.33 B | 17.82 B | 461.12 B | 102.32 B | 74.73 B | |
| S2 (120 mM NaCl) | 202.44 C | 16.70 C | 412.09 C | 91.58 C | 68.37 C | |
| LSD0.05 | 2.67 | 0.28 | 6.37 | 2.23 | 0.92 | |
| ZnO-NPs | ||||||
| Control (0 mg/L) | 191.78 C | 15.79 C | 418.26 C | 92.96 C | 72.26 C | |
| 50 mg/L | 232.01 B | 18.02 B | 450.61 B | 100.08 B | 75.03 B | |
| 100 mg/L | 251.67 A | 18.99 A | 530.58 A | 117.92 A | 78.17 A | |
| LSD0.05 | 4.70 | 0.41 | 10.76 | 3.96 | 1.10 | |
| Salinity × ZnO-NPs | ||||||
| Salinity | ZnO-NPs | |||||
| S0 (0 mM NaCl) | Control | 209.67 e | 16.37 | 486.97 c | 108.21 | 81.77 a |
| 50 mg/L | 255.00 b | 18.63 | 508.63 b | 113.52 | 81.83 a | |
| 100 mg/L | 284.33 a | 19.90 | 583.10 a | 129.47 | 83.47 a | |
| S1 (60 mM NaCl) | Control | 197.33 f | 15.93 | 420.37 e | 93.43 | 71.73 d |
| 50 mg/L | 226.01 d | 18.28 | 446.07 d | 98.53 | 74.77 c | |
| 100 mg/L | 246.67 c | 19.23 | 516.93 b | 115.01 | 77.71 b | |
| S2 (120 mM NaCl) | Control | 168.33 g | 15.07 | 347.43 g | 77.23 | 63.27 f |
| 50 mg/L | 215.03 e | 17.18 | 397.13 f | 88.23 | 68.52 e | |
| 100 mg/L | 224.00 d | 17.83 | 491.70 c | 109.31 | 73.33 cd | |
| LSD0.05 | 8.14 | 0.71 | 18.63 | 6.86 | 1.90 | |
| Treatments/Parameters | Proline (mg/gFW) | Phenolic (mgGAE/gFM) | SOD (U/gFW) | CAT (U/gFW/min) | APX (µmol/gFW/min) | |
|---|---|---|---|---|---|---|
| Salinity | ||||||
| S0 (0 mM NaCl) | 0.48 C | 47.25 B | 98.67 C | 31.56 C | 19.56 C | |
| S1 (60 mM NaCl) | 1.65 B | 61.40 A | 137.11 B | 47.56 B | 29.11 B | |
| S2 (120 mM NaCl) | 2.15 A | 35.11 C | 175.11 A | 61.44 A | 39.56 A | |
| LSD0.05 | 0.05 | 2.26 | 2.15 | 0.80 | 1.53 | |
| ZnO-NPs | ||||||
| Control (0 mg/L) | 2.38 A | 41.06 C | 126.56 C | 42.56 C | 25.33 C | |
| 50 mg/L | 1.02 B | 49.21 B | 137.89 B | 46.01 B | 28.02 B | |
| 100 mg/L | 0.88 C | 53.49 A | 146.44 A | 52.00 A | 31.89 C | |
| LSD0.05 | 0.06 | 1.33 | 1.90 | 1.61 | 1.21 | |
| Salinity × ZnO-NPs | ||||||
| Salinity | ZnO-NPs | |||||
| S0 (0 mM NaCl) | Control | 0.49 f | 44.32 e | 93.33 i | 23.99 g | 18.67 g |
| 50 mg/L | 0.48 f | 47.39 d | 98.33 h | 31.01 g | 19.00 fg | |
| 100 mg/L | 0.46 f | 50.03 c | 104.33 g | 34.33 f | 21.01 f | |
| S1 (60 mM NaCl) | Control | 2.83 b | 52.06 c | 126.33 f | 41.67 e | 25.67 e |
| 50 mg/L | 1.17 d | 61.52 b | 138.67 e | 45.67 d | 28.67 d | |
| 100 mg/L | 0.96 e | 70.62 a | 146.33 d | 55.33 c | 33.00 c | |
| S2 (120 mM NaCl) | Control | 3.83 a | 26.79 g | 160.01 c | 56.67 c | 31.67 c |
| 50 mg/L | 1.39 c | 38.71 f | 176.67 b | 61.33 b | 36.33 b | |
| 100 mg/L | 1.21 d | 39.83 f | 188.67 a | 66.33 a | 41.67 a | |
| LSD0.05 | 0.11 | 2.31 | 3.28 | 2.78 | 2.10 | |
| Variables | DW | LA | RWC | Pn | Sc | iWUE | SOD | CAT | APX | Na+/K+ | Zn++ | GY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DW | - | *** | *** | * | ** | ns | * | ns | ns | ** | *** | ** |
| LA | 0.986 | - | *** | ** | ** | ns | * | ns | ns | ** | *** | *** |
| RWC | 0.914 | 0.929 | - | *** | *** | ns | ** | * | ns | *** | ** | *** |
| Pn | 0.789 | 0.804 | 0.931 | - | *** | ns | *** | ** | * | *** | * | *** |
| Sc | 0.813 | 0.832 | 0.953 | 0.993 | - | ns | *** | ** | ** | *** | * | *** |
| iWUE | −0.562 | −0.581 | −0.596 | −0.366 | −0.468 | - | ns | ns | ns | ns | ns | ns |
| SOD | −0.693 | −0.728 | −0.865 | −0.912 | −0.940 | 0.570 | - | *** | *** | ** | ns | *** |
| CAT | −0.432 | −0.470 | −0.721 | −0.803 | −0.832 | 0.533 | 0.918 | - | *** | ** | ns | ** |
| APX | −0.376 | −0.422 | −0.654 | −0.776 | −0.802 | 0.469 | 0.904 | 0.981 | - | ns | ns | ** |
| Na+/K+ | −0.838 | −0.858 | −0.960 | −0.936 | −0.943 | 0.493 | 0.867 | 0.724 | 0.650 | - | ** | *** |
| Zn++ | 0.935 | 0.963 | 0.854 | 0.777 | 0.784 | −0.393 | −0.638 | −0.351 | −0.330 | −0.803 | - | ** |
| GY | 0.881 | 0.910 | 0.967 | 0.945 | 0.972 | −0.594 | −0.929 | −0.777 | −0.755 | −0.918 | 0.851 | - |
| PC | Eigenvalue | Variance (%) | Cumulative Eigenvalue (%) | Cumulative Variance (%) |
|---|---|---|---|---|
| 1 | 21,075.1 | 89.786 | 21,075.1 | 89.786 |
| 2 | 1700.78 | 7.2458 | 22,775.88 | 97.0318 |
| 3 | 333.833 | 1.4222 | 23,109.713 | 98.454 |
| 4 | 219.707 | 0.93601 | 23,329.42 | 99.39001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seleiman, M.F.; Ahmad, A.; Alhammad, B.A.; Tola, E. Exogenous Application of Zinc Oxide Nanoparticles Improved Antioxidants, Photosynthetic, and Yield Traits in Salt-Stressed Maize. Agronomy 2023, 13, 2645. https://doi.org/10.3390/agronomy13102645
Seleiman MF, Ahmad A, Alhammad BA, Tola E. Exogenous Application of Zinc Oxide Nanoparticles Improved Antioxidants, Photosynthetic, and Yield Traits in Salt-Stressed Maize. Agronomy. 2023; 13(10):2645. https://doi.org/10.3390/agronomy13102645
Chicago/Turabian StyleSeleiman, Mahmoud F., Awais Ahmad, Bushra Ahmed Alhammad, and ElKamil Tola. 2023. "Exogenous Application of Zinc Oxide Nanoparticles Improved Antioxidants, Photosynthetic, and Yield Traits in Salt-Stressed Maize" Agronomy 13, no. 10: 2645. https://doi.org/10.3390/agronomy13102645
APA StyleSeleiman, M. F., Ahmad, A., Alhammad, B. A., & Tola, E. (2023). Exogenous Application of Zinc Oxide Nanoparticles Improved Antioxidants, Photosynthetic, and Yield Traits in Salt-Stressed Maize. Agronomy, 13(10), 2645. https://doi.org/10.3390/agronomy13102645