

Very Early Biomarkers Screening for Water Deficit Tolerance in Commercial Eucalyptus Clones

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Seedling Acquisition and Management
2.2. Control and Water Stress Treatments
2.3. Traits
2.3.1. Morphological
2.3.2. Nutritional Traits
2.3.3. Physiological Traits
2.4. Statistical Analysis
2.4.1. Selection Index
2.4.2. Selection Index Based on Weights
2.4.3. Validation Procedures
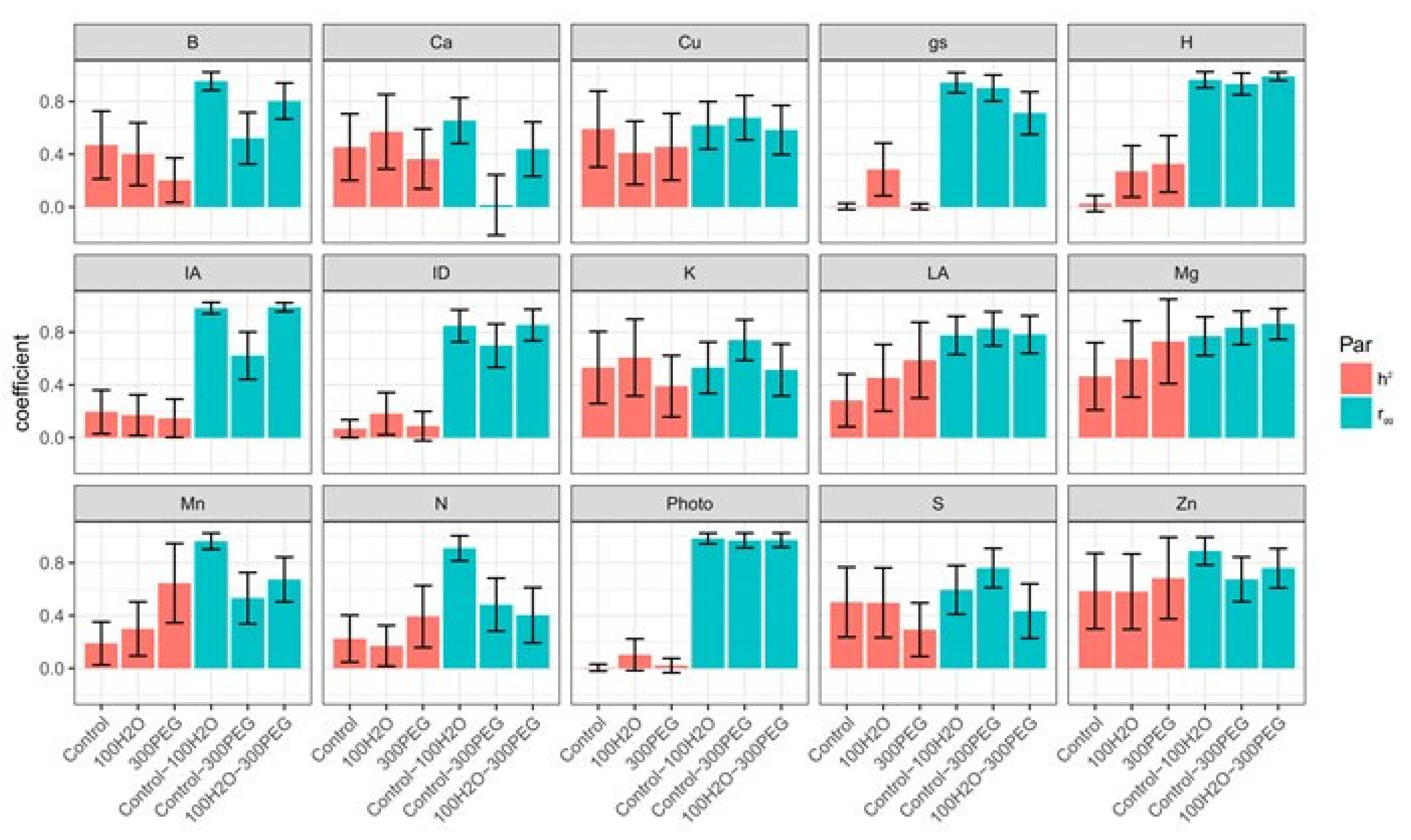
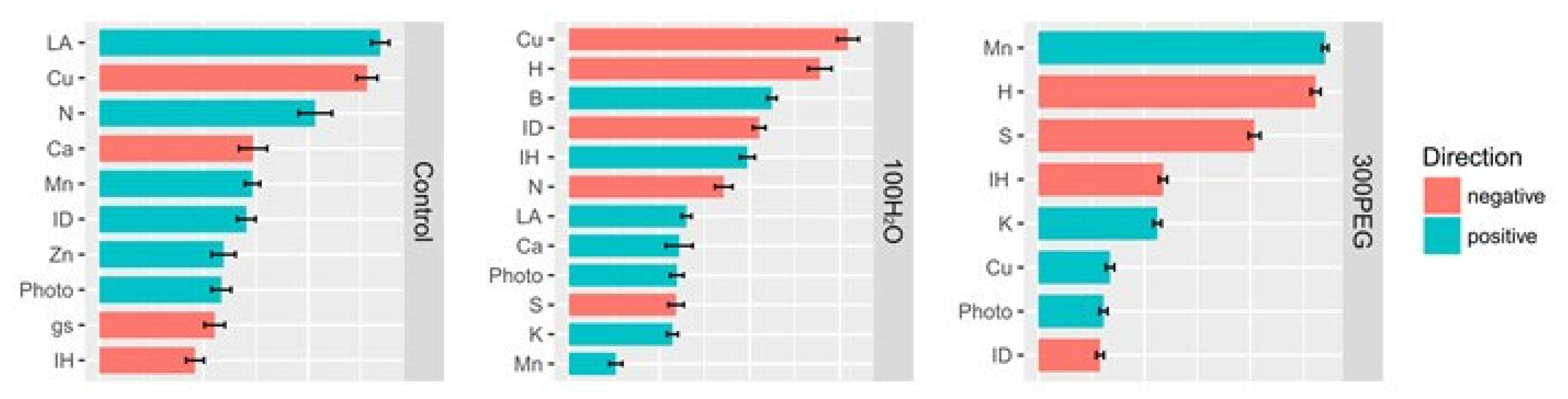
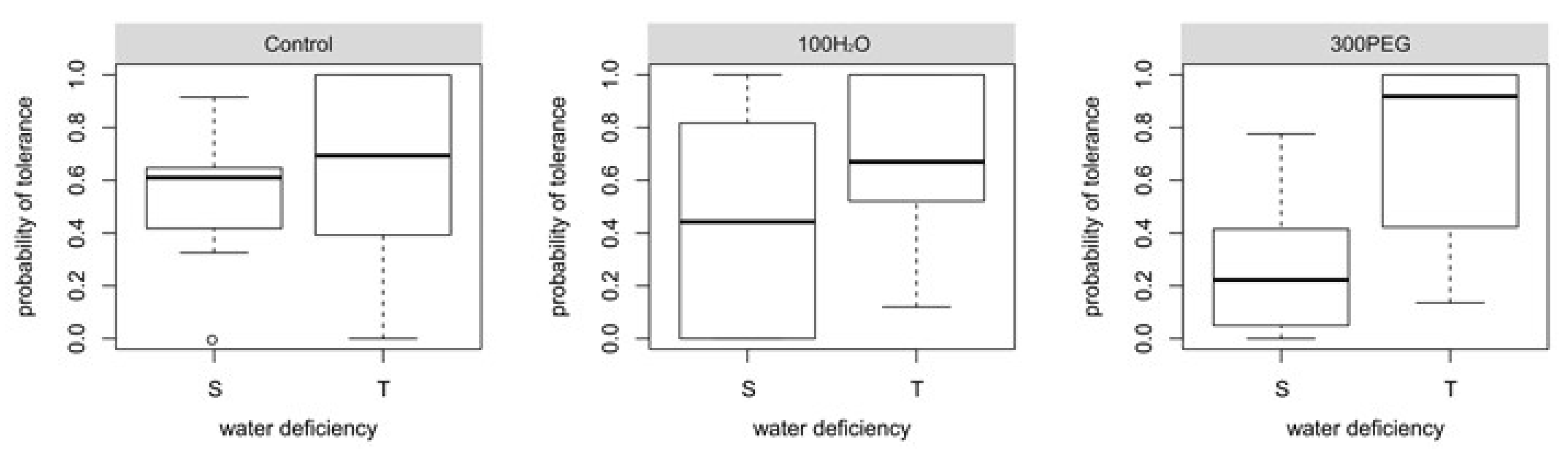
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shanker, A.K.; Maheswari, M.; Yadav, S.K.; Desai, S.; Bhanu, D.; Attal, N.B.; Venkateswarlu, B. Drought stress responses in crops. Funct. Integr. Genom. 2014, 14, 11–22. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Saifullah, U.; Ashraf, M.Y.; Ehsanullah. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop. Sci. 2011, 5, 764–777. [Google Scholar]
- Saadaoui, E.; Yahia, K.B.; Dhahri, S.; Jamaa, M.L.B.; Khouja, M.L. An overview of adaptive responses to drought stress in Eucalyptus spp. For. Stud. 2017, 67, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.R.R.; Souza, G.S.F.; Rodrigues, A.C.P.; Mendonça-Filho, A.L.; Klar, A.E. Análise de crescimento em clones de eucalipto submetidos a estresse hídrico. Irriga 2010, 15, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.D.; Nascimento, J.S.; Scarpinati, E.A.; Paula, R.C. Classification of Eucalyptus urograndis hybrids under different water availability based on biometric traits. For. Syst. 2014, 23, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.S.; Resende, R.T.; Mesquita, A.G.; Rosado, A.M.; Cruz, C.D. Seleção precoce para características de crescimento em testes clonais de Eucalyptus urophylla. Sci. For. 2014, 42, 251–257. [Google Scholar]
- Corrêa, T.R.; Picoli, E.A.T.; Souza, G.A.; Condé, S.A.; Silva, N.M.; Lopes-Mattos, K.L.B.; Resende, M.D.V.; Zauza, E.A.V.; Oda, S. Phenotypic markers in early selection for tolerance to dieback in Eucalyptus. Ind. Crop. Prod. 2017, 107, 130–138. [Google Scholar] [CrossRef]
- Picoli, E.A.T.; de Resende, M.D.V.; Oda, S. Come Hell or High Water: Breeding the Profile of Eucalyptus Tolerance to Abiotic Stress Focusing Water Deficit. In Plant Growth and Stress Physiology; Plant in Challenging Environments; Gupta, D.K., Palma, J.M., Eds.; Springer: Cham, Switzerland, 2021; Volume 3, pp. 91–127. [Google Scholar] [CrossRef]
- Caetano, D.D.N. Prospecção De Marcadores Morfológicos, Anatômicos, Fisiológicos E Metabólicos Visando a Seleção Precoce Quanto a Tolerância à Seca De Ponteiros Em Eucalyptus. Dissertation/Master’s Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 16 July 2016; 38p. [Google Scholar]
- Dias, C.N.; Picoli, E.A.T.; Souza, G.A.; Farag, M.A.; Scotti, M.T.; Barbosa Filho, J.M.; da Silva, M.S.; Tavares, J.F. Phenolics metabolismo provides a tool for screening drought tolerant Eucalyptus grandis hybrids. Austr. J. Crop. Sci. 2017, 11, 1016–1025. [Google Scholar] [CrossRef]
- Szijarto, L.; van de Voort, F.R. Determination of added water and bovine milk to caprine milk. J. Dairy Sci. 1983, 66, 620–623. [Google Scholar] [CrossRef]
- Sarruge, J.R.; Haag, H.P. Análise Química em Plantas; Departamento de Química, ESALQ/USP: Piracicaba, Brazil, 1974; 56p. [Google Scholar]
- Braga, J.M.; Defelipo, B.V. Determinação espectrofotométrica do fósforo em extrato de solo e plantas. R. Ceres. 1974, 21, 73–85. [Google Scholar]
- Alvarez, V.V.H.; Dias, L.E.; Ribeiro, E.S., Jr.; Fonseca, C.A. Métodos de Análises de Enxofre em Solos e Plantas; UFV: Viçosa, Brazil, 2001; 131p. [Google Scholar]
- Resende, M.D.V.; Silva, F.F.; Azevedo, C.F. Estatística Matemática, Biométrica e Computacional, 1st ed.; Suprema: Viçosa, Brazil, 2014; 881p. [Google Scholar]
- Resende, M.D.V. Genética Quantitativa e de Populações, 1st ed.; Suprema: Viçosa, Brazil, 2015; 463p. [Google Scholar]
- Resende, M.D.V. Software Selegen-REML/BLUP: A useful tool for plant breeding. CBAB 2016, 16, 330–339. [Google Scholar] [CrossRef]
- Resende, M.D.V. Genética Biométrica e Estatística No Melhoramento de Plantas Perenes; Embrapa, Informação Tecnológica: Brasília, Brazil, 2002; 975p. [Google Scholar]
- Mulamba, N.N.; Mock, J.J. Improvement of yield potential of the Eto Blanco maize (Zea mays L.) population by breeding for plant traits. Egypt J. Genet. Cytol. 1978, 7, 40–51. [Google Scholar]
- Team, R. Core. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Williams, J.S. The evaluation of a selection index. Biometrics 1962, 18, 375–393. [Google Scholar] [CrossRef]
- Kohavi, R. A study of cross-validation and bootstrap for accuracy estimation and model selection. In Proceedings of the 14th International Joint Conference on Artificial Intelligence, Montreal, QC, Canada, 20–25 August 1995. [Google Scholar]
- White, D.A.; Crombie, D.S.; Kinal, J.; Battaglia, M.; McGrath, J.F.; Mendham, D.S.; Walker, S.N. Managing productivity and drought risk in Eucalyptus globulus plantations in south-western Australia. For. Ecol. Manag. 2009, 259, 33–44. [Google Scholar] [CrossRef]
- Barros Filho, N.F. Discriminação Isotópica Do 13C e Nutrição Com Cálcio e Boro em Clones de Eucalipto Submetidos Ao Déficit Hídrico. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, MG, Brazil, 28 March 2014; 59p. [Google Scholar]
- Müller, C.; Hodecker, B.E.R.; Merchant, A.; Barros, N.F. Nutritional Efficiency of Eucalyptus Clones Under Water Stress. Rev. Bras. Ciênc. Solo 2017, 41, e0160528. [Google Scholar] [CrossRef] [Green Version]
- Stape, J.L.; Binkley, D.; Ryan, M.G. Eucalyptus production and the supply, use and efficiency of use of water, light and nitrogen across a geographic gradient in Brazil. For. Ecol. Manag. 2004, 193, 17–31. [Google Scholar] [CrossRef]
- Bedon, F.; Villar, E.; Vincent, D.; Jupuy, J.-W.; Lomenech, A.-M.; Mabialangoma, A.; Chaumeil, P.; Barré, A.; Plomion, C.; Gion, G.-M. Proteomic plasticity of two Eucalyptus genotypes under contrasted water regimes in the field. Plant Cell Environ. 2012, 35, 790–805. [Google Scholar] [CrossRef] [PubMed]
- Coopman, R.E.; Jara, J.C.; Bravo, L.A.; Sáez, K.L.; Mella, G.R.; Escobar, R. Changes in morpho-physiological attributes of Eucalyptus globulus plants in response to different drought hardening treatments. Eletronic. J. Biotechnol. 2008, 11, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S. Introdução À Genética Quantitative; UFV: Viçosa, Brazil, 1987; 279p. [Google Scholar]
- Resende, M.D.V.; Duarte, J.B. Precisão e controle de qualidade em experimentos de avaliação de cultivares. Pesq. Agropec. Trop. 2007, 37, 182–194. [Google Scholar]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-deficit stress-induced anatomical changes in higher plants. Plant Biol. Pathol. 2008, 331, 215–225. [Google Scholar] [CrossRef]
- Grassi, G.; Meir, P.; Cromer, R.; Tompkins, D.; Jarvis, P.G. Photosynthetic parameters in seedlings of Eucalyptus grandis as affected by rate of nitrogen supply. Plant Cell Environ. 2002, 5, 1677–1688. [Google Scholar] [CrossRef]
- Rivero, R.M.; Shulaev, V.; Blumwald, E. Cytokinin-Dependent Photorespiration and the Protection of Photosynthesis during Water Deficit. Plant Physiol. 2009, 150, 1530–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wingler, A.; Quick, W.P.; Bungard, R.A.; Bailey, K.J.; Lea, P.J.; Leegood, R.C. The role of photorespiration during drought stress: An analysis utilizing barley mutants with reduced activities of photorespiratory enzymes. Plant Cell Environ. 1999, 22, 361–373. [Google Scholar] [CrossRef]
- Bloom, A.J. Photorespiration and nitrate assimilation: A major intersection between plant carbon and nitrogen. Photosynth. Res. 2014, 123, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Lisar, S.Y.S.; Motafakkerazad, R.; Hossain, M.M.; Rahman, I.M.M. Water Stress in Plants: Causes, Effects and Responses, Water Stress; Rahman, I.M.M., Hasegawa, H., Eds.; InTech: Rijeka, Croatia, 2012; pp. 1–14. [Google Scholar] [CrossRef] [Green Version]
- Battie-Laclau, P.; Laclau, J.P.; Domec, J.C.; Christina, M.; Bouillet, J.P.; Piccolo, M.C.; Gonçalves, J.L.M.; Moreira e Moreira, R.; Krusche, A.V.; Bouvet, J.-M.; et al. Effects of potassium and sodium supply on drought-adaptive mechanisms in Eucalyptus grandis plantations. New Phytol. 2014, 203, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Battie-Laclau, P.; Delgado-Rojas, J.S.; Christina, M.; Nouvellon, Y.; Bouillet, J.-P.; Piccolo, M.C.; Moreira, M.Z.; Gonçalves, J.L.M.; Roupsard, O.; Laclau, J.-P. Potassium fertilization increases water-use efficiency for stem biomass production without affecting intrinsic water-use efficiency in Eucalyptus grandis plantations. For. Ecol. Manag. 2016, 364, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Hepler, P.K. Calcium: A Central Regulator of Plant Growth and Development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- Waters, B.M. Moving magnesium in plant cells. New Phytol. 2011, 190, 510–513. [Google Scholar] [CrossRef] [Green Version]
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants: Principles and Perspectives; Sinauer Associates: Sunderland, UK, 2004; 380p. [Google Scholar]
- Tezara, W.; Mitchell, V.J.; Driscoll, S.D.; Lawlor, D.W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 1999, 401, 914–917. [Google Scholar] [CrossRef]
- Ma, D.; Sun, D.; Wang, C.; Ding, H.; Qin, H.; Hou, J.; Huang, X.; Xie, Y.; Guo, T. Physiological Responses and Yield of Wheat Plants in Zinc-Mediated Alleviation of Drought Stress. Front. Plant Sci. 2017, 8, 914–917. [Google Scholar] [CrossRef] [Green Version]
- Burnell, J.N. The Biochemistry of Manganese in Plants. In Developments in Plant and Soil Sicences; Grahan, R.D., Hannam, R.J., Uren, N.C., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 125–137. [Google Scholar] [CrossRef]
- Leite, F.P.; Novais, R.F.; Silva, I.R.; Barros, N.F.; Neves, J.C.L.; Medeiros, A.G.B.; Ventrella, M.C.; Villani, E.M.A. Manganese accumulation and its relation to “eucalyptus shoot blight in the Vale do Rio Doce”. Rev. Bras. Ciênc. Solo 2014, 38, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Mou, D.; Yao, Y.; Yang, Y.; Zhang, Y.; Tian, C.; Achal, V. Plant high tolerance to excess manganese related with root growth, manganese distribution and antioxidative enzyme activity in three grape cultivars. Ecotoxicol. Environ. Saf. 2011, 74, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Christeller, J.T. The effects of bivalent cations on ribulose bisphosphate carboxylase/oxygenase. Biochem. J. 1981, 193, 839–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattiello, E.M.; Ruiz, H.A.; Silva, I.R.; Barros, N.F.; Neves, J.C.L.; Behling, M. Transporte de boro no solo e sua absorção por eucalipto. Rev. Bras. Ciênc. Solo 2009, 33, 1281–1290. [Google Scholar] [CrossRef] [Green Version]
- Hajiboland, R.; Bastani, S. Tolerance to water stress in boron deficient tea (Camellia sinensis) plants. Folia. Hort. 2012, 24, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Z.; Zhao, P.; Oren, R.; McCarthy, H.R.; Niu, J.F.; Zhu, L.W.; Ni, G.Y.; Huang, Y.Q. Water use strategies of a young Eucalyptus. urophylla forest in response to seasonal change of climatic factors in South China. Biogeosciences 2015, 12, 10469–10510. [Google Scholar] [CrossRef] [Green Version]
- Carnicer, J.; Coll, M.; Ninyerola, M.; Pons, X.; Sánchez, G.; Peñuelas, J. Widespread crown condition decline, food web disruption, and amplified tree mortality with increase climate-type drought. Proc. Natl. Acad. Sci. USA 2011, 108, 1474–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | Genetic Background | Phenotype Associated with Water Deficit |
|---|---|---|
| E-01 | E. grandis × E. urophylla | Susceptible |
| E-02 | E. grandis × E. pellita | Tolerant |
| E-03 | E. grandis × E. urophylla | Tolerant |
| E-04 | E. grandis × E. pellita | Susceptible |
| E-05 | E. grandis × E. urophylla | Susceptible |
| E-06 | E. platyphylla | Tolerant |
| E-07 | E. grandis × E. urophylla | Susceptible |
| E-08 | E. grandis × E. urophylla | Tolerant |
| E-09 | E. grandis × E. urophylla | Susceptible |
| E-10 | E. grandis | Tolerant |
| E-11 | E. grandis × E. urophylla | Tolerant |
| E-12 | E. urophylla | Tolerant |
| E-13 | E. grandis | Susceptible |
| E-14 | E. grandis × E. urophylla | Tolerant |
| E-15 | (GG 100) | Susceptible |
| E-16 | Mercado I144 | Tolerant |
| E-17 | E. grandis × E. urophylla | Tolerant |
| E-18 | E. grandis × E. urophylla | Tolerant |
| E-19 | E. grandis × E. urophylla | Susceptible |
| Parameters | ||||||||
|---|---|---|---|---|---|---|---|---|
| Traits | Acgen | % | % | |||||
| PH | 0.47 | 0.84 | 0.91 | 0.00 | 0.99 | 9.70 | 10.35 | 61.07 |
| LA | 0.40 | 0.74 | 0.86 | 0.11 | 0.28 | 20.89 | 23.09 | 177.24 |
| ID | 0.27 | 0.66 | 0.81 | 0.04 | 0.85 | 11.34 | 18.07 | 3.22 |
| IH | 0.31 | 0.73 | 0.85 | 0.00 | 0.99 | 17.13 | 25.67 | 31.70 |
| N | 0.11 | 0.36 | 0.70 | 0.17 | 0.40 | 6.17 | 15.39 | 1.68 |
| K | 0.29 | 0.58 | 0.76 | 0.28 | 0.51 | 7.48 | 9.01 | 1.08 |
| Ca | 0.19 | 0.54 | 0.73 | 0.32 | 0.37 | 11.66 | 18.72 | 0.79 |
| Mg | 0.54 | 0.84 | 0.91 | 0.08 | 0.86 | 13.38 | 10.86 | 0.42 |
| S | 0.29 | 0.69 | 0.83 | 0.21 | 0.57 | 17.64 | 23.40 | 0.23 |
| Cu | 0.32 | 0.74 | 0.86 | 0.19 | 0.63 | 16.72 | 20.05 | 5.90 |
| Zn | 0.48 | 0.78 | 0.88 | 0.15 | 0.76 | 16.76 | 14.75 | 29.58 |
| Mn | 0.37 | 0.69 | 0.83 | 0.18 | 0.67 | 14.54 | 16.05 | 678.86 |
| B | 0.34 | 0.77 | 0.88 | 0.10 | 0.76 | 13.13 | 16.78 | 85.92 |
| A | 0.22 | 0.62 | 0.79 | 0.00 | 0.97 | 20.81 | 39.41 | 6.50 |
| gs | 0.15 | 0.48 | 0.70 | 0.06 | 0.71 | 20.19 | 46.58 | 0.08 |
| Clone | Rank PH | Rank LA | Rank ID | Rank IH | Rank N | Rank K | Rank Ca | Rank Mg | Rank S | Rank Cu | Rank Zn | Rank Mn | Rank B | Rank A | Rank gs(1) | Rank Average |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-18 T | 12-7 | 1-3 | 14-4 | 4-12 | 7-1 | 7-14 | 1-5 | 1-1 | 19-2 | 5-16 | 11-7 | 18-14 | 17-13 | 6-1 | 5-7 | 8.03 |
| E-08 T | 1-4 | 7-5 | 4-11 | 16-1 | 8-2 | 4-3 | 3-3 | 8-5 | 1-12 | 9-18 | 18-4 | 11-16 | 7-18 | 16-14 | 14-9 | 8.40 |
| E-02 T | 2-1 | 2-1 | 2-1 | 15-4 | 4-6 | 19-1 | 4-15 | 4-13 | 15-7 | 12-12 | 5-9 | 19-13 | 18-12 | 17-8 | 8-19 | 8.93 |
| E-17 T | 19-17 | 17-6 | 11-2 | 1-18 | 11-5 | 1-19 | 9-6 | 2-2 | 17-3 | 14-9 | 17-1 | 14-15 | 4-7 | 2-2 | 13-5 | 8.96 |
| E-11 T | 9-15 | 15-9 | 1-6 | 10-14 | 5-13 | 18-6 | 8-14 | 12-19 | 6-9 | 10-2 | 14-3 | 9-4 | 15-5 | 4-5 | 10-6 | 9.20 |
| E-01 S | 16-16 | 12-7 | 7-3 | 17-8 | 16-3 | 2-9 | 17-7 | 17-8 | 5-6 | 13-17 | 8-13 | 5-9 | 14-11 | 3-1 | 3-8 | 9.37 |
| E-12 T | 3-6 | 5-11 | 6-13 | 12-7 | 1-11 | 3-13 | 14-12 | 18-12 | 7-19 | 18-4 | 9-17 | 16-3 | 1-3 | 8-11 | 15-4 | 9.40 |
| E-10 T | 7-3 | 19-18 | 19-12 | 9-11 | 9-18 | 13-11 | 12-2 | 16-15 | 2-15 | 7-5 | 4-15 | 3-6 | 1-2 | 5-18 | 9-1 | 9.57 |
| E-03 T | 8-5 | 16-10 | 9-14 | 8-5 | 10-8 | 12-8 | 2-11 | 6-7 | 14-17 | 2-13 | 1-18 | 15-8 | 10-14 | 18-4 | 1-16 | 9.67 |
| E-06 T | 18-18 | 11-13 | 10-7 | 13-3 | 18-10 | 9-5 | 11-8 | 10-4 | 11-1 | 3-19 | 3-19 | 2-19 | 5-5 | 7-16 | 11-14 | 10.03 |
| E-04 S | 4-8 | 6-2 | 8-16 | 14-10 | 3-14 | 6-16 | 5-4 | 3-3 | 18-4 | 16-7 | 15-11 | 16-19 | 19-19 | 10-9 | 2-17 | 10.13 |
| E-16 T | 11-9 | 3-12 | 18-17 | 10-16 | 14-17 | 17-12 | 7-9 | 14-10 | 4-16 | 1-8 | 10-2 | 10-2 | 12-8 | 14-12 | 7-11 | 10.20 |
| E-14 T | 6-10 | 10-14 | 13-15 | 18-2 | 12-16 | 5-17 | 16-17 | 5-9 | 16-10 | 17-11 | 12-6 | 8-10 | 2-1 | 11-10 | 17-2 | 10.27 |
| E-15 S | 13-13 | 4-17 | 15-10 | 6-17 | 13-19 | 16-18 | 6-1 | 11-6 | 10-11 | 8-6 | 2-16 | 7-12 | 9-4 | 12-17 | 12-10 | 10.70 |
| E-05 S | 15-19 | 13-15 | 3-5 | 19-6 | 2-4 | 15-10 | 10-18 | 7-11 | 8-13 | 3-15 | 6-10 | 6-1 | 13-10 | 15-15 | 19-18 | 10.80 |
| E-09 S | 14-11 | 9-8 | 5-8 | 7-13 | 19-15 | 14-7 | 18-16 | 13-16 | 13-5 | 11-11 | 13-14 | 13-18 | 8-16 | 1-3 | 6-3 | 10.93 |
| E-13 S | 17-14 | 14-16 | 13-18 | 2-19 | 15-9 | 11-2 | 13-10 | 9-14 | 12-8 | 15-10 | 7-11 | 4-7 | 11-15 | 9-6 | 16-13 | 11.33 |
| E-19 S | 10-12 | 8-4 | 17-8 | 5-15 | 17-7 | 8-15 | 15-19 | 15-17 | 9-14 | 6-14 | 16-5 | 17-11 | 6-6 | 19-19 | 4-12 | 11.70 |
| E-07 S | 5-21 | 18-19 | 16-19 | 11-9 | 6-12 | 10-4 | 19-13 | 10-18 | 3-18 | 19-16 | 19-8 | 12-17 | 3-9 | 13-13 | 18-15 | 12.33 |
| Control | 100H2O | 300PEG | ||
|---|---|---|---|---|
| Average of all blocks | r | 0.17 | 0.37 | 0.56 |
| RMSE | 0.55 | 0.49 | 0.42 | |
| Observed | Success rate | |||
| Control | 100H2O | 300PEG | ||
| Average of all blocks | Susceptible (8) | 25.00% | 50.00% | 87.50% |
| Tolerant (11) | 72.73% | 72.73% | 63.54% | |
| Total (19) | 52.63% | 63.16% | 73.68% | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corrêa, T.R.; Picoli, E.A.d.T.; Pereira, W.L.; Condé, S.A.; Resende, R.T.; de Resende, M.D.V.; da Costa, W.G.; Cruz, C.D.; Zauza, E.A.V. Very Early Biomarkers Screening for Water Deficit Tolerance in Commercial Eucalyptus Clones. Agronomy 2023, 13, 937. https://doi.org/10.3390/agronomy13030937
Corrêa TR, Picoli EAdT, Pereira WL, Condé SA, Resende RT, de Resende MDV, da Costa WG, Cruz CD, Zauza EAV. Very Early Biomarkers Screening for Water Deficit Tolerance in Commercial Eucalyptus Clones. Agronomy. 2023; 13(3):937. https://doi.org/10.3390/agronomy13030937
Chicago/Turabian StyleCorrêa, Thais R., Edgard Augusto de T. Picoli, Washington Luiz Pereira, Samyra A. Condé, Rafael T. Resende, Marcos Deon V. de Resende, Weverton Gomes da Costa, Cosme Damião Cruz, and Edival Angelo V. Zauza. 2023. "Very Early Biomarkers Screening for Water Deficit Tolerance in Commercial Eucalyptus Clones" Agronomy 13, no. 3: 937. https://doi.org/10.3390/agronomy13030937
APA StyleCorrêa, T. R., Picoli, E. A. d. T., Pereira, W. L., Condé, S. A., Resende, R. T., de Resende, M. D. V., da Costa, W. G., Cruz, C. D., & Zauza, E. A. V. (2023). Very Early Biomarkers Screening for Water Deficit Tolerance in Commercial Eucalyptus Clones. Agronomy, 13(3), 937. https://doi.org/10.3390/agronomy13030937