
Desertification Reversal Promotes the Complexity of Plant Community by Increasing Plant Species Diversity of Each Plant Functional Type


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Design and Laboratory Analyses
2.3. Statistical Analyses
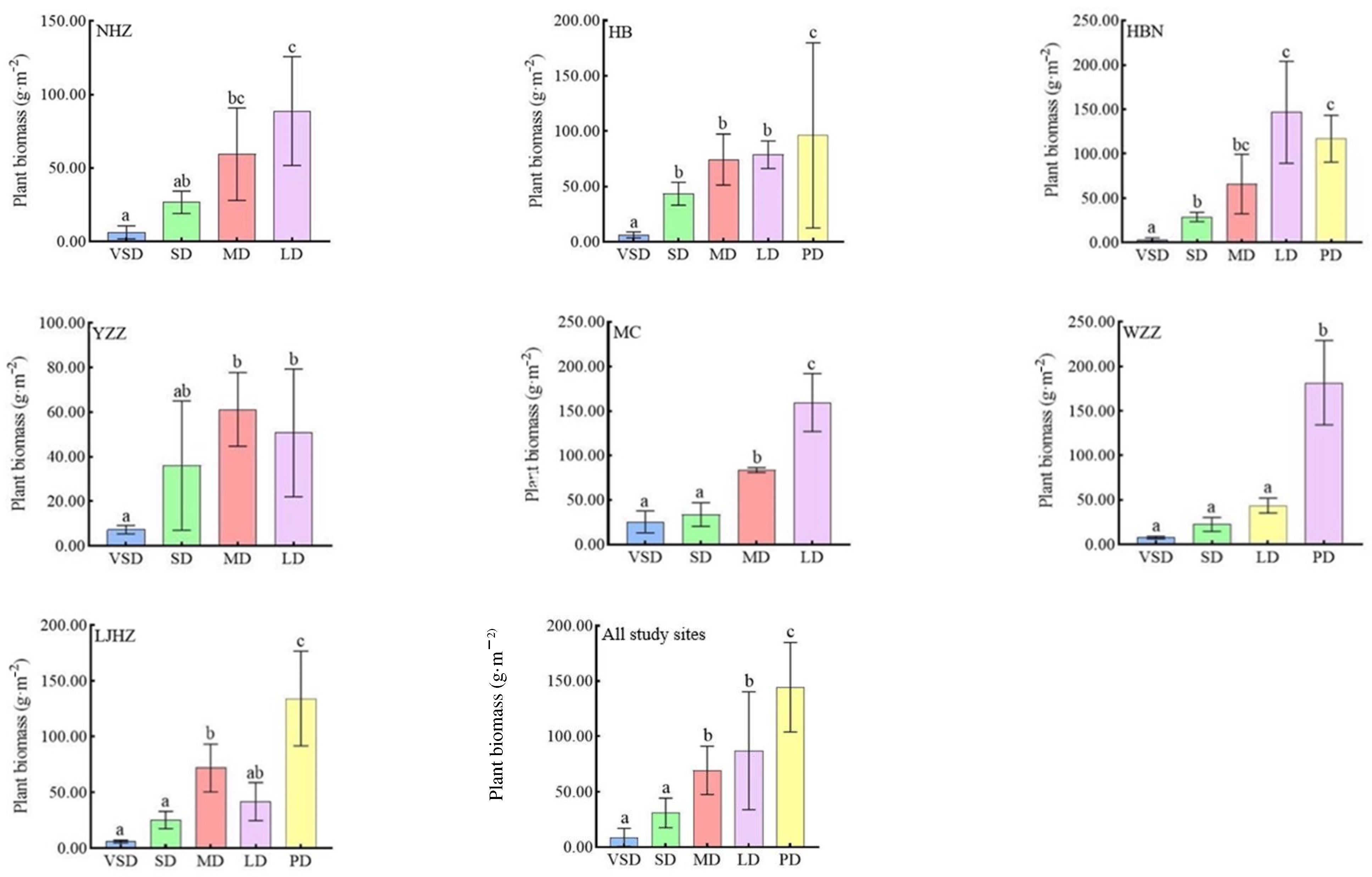
3. Results
3.1. Changes in Vegetation Conditions and Quantitative Plant Characteristics
3.2. Effects of Desertification Reversal on Plant Functional Types
3.3. Responses of Plant Diversity and Biomass to the Reversal of Sandy Desertification
4. Discussion
4.1. Changes in Vegetation Conditions and Plant Community Characteristics with Desertification Reversal
4.2. Relationships between Vegetation Characteristics and Desertification Reversal
4.3. Differences in Changes in Vegetation Characteristics between Early and Later Stages of Desertification Reversal
5. Conclusions and Implications
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| COV | Vegetation cover |
| IV | Importance value |
| DV | Dominance value |
| LD | Light desertification |
| MD | Moderate desertification |
| PD | Potential desertification |
| PFT | Plant functional type |
| RA | Relative abundance |
| RC | Relative cover |
| RF | Relative frequency |
| RH | Relative height |
| S% | Percentages of the number of plant species |
| SAND | Proportion of the bare (mobile) sand area to the total ground area |
| SD | Severe desertification |
| VSD | Very severe desertification |
References
- Adger, W.N.; Benjaminsen, T.A.; Brown, K.; Svarstad, H. Advancing a political ecology of global environmental discourses. Dev. Chang. 2001, 32, 681–715. [Google Scholar] [CrossRef]
- Allington, G.R.H.; Valone, T.J. Reversal of desertification: The role of physical and chemical soil properties. J. Arid. Environ. 2010, 74, 973–977. [Google Scholar] [CrossRef]
- MEA. Millennium Ecosystem Assessment, Ecosystems and Human Well-Being: Desertification Synthesis; World Resources Institute: Washington, DC, USA, 2005. [Google Scholar]
- Li, S.; Xie, Y.C. Investigating coupled impacts of climate change and socioeconomic transformation on desertification by using multitemporal landsat images: A case study in central Xilingol, China. IEEE Geosci. Remote Sens. Lett. 2013, 10, 1244–1248. [Google Scholar] [CrossRef]
- UNCCD. United Nations Convention to Combat Desertification, Elaboration of an International Convention to Combat Desertification in Countries Experiencing Serious Drought and/or Desertification, Particularly in Africa; United Nations: New York, NY, USA, 1994. [Google Scholar]
- Zhou, W.; Sun, Z.G.; Li, J.L.; Gang, C.C.; Zhang, C.B. Desertification dynamic and the relative roles of climate change and human activities in desertification in the Heihe River Basin based on NPP. J. Arid Land 2013, 5, 465–479. [Google Scholar] [CrossRef]
- Johnson, D.; Phoenix, G.K.; Grime, J.P. Plant community composition, not diversity, regulates soil respiration in grasslands. Biol. Lett. 2008, 4, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.P.; Eisenhauer, N. Plant community composition determines the strength of top-down control in a soil food web motif. Sci. Rep. 2015, 5, 9134. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Cabido, M. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci. 1997, 8, 463–474. [Google Scholar] [CrossRef]
- Noble, I.R.; Gitay, H. A functional classification for predicting the dynamics of landscapes. J. Veg. Sci. 1996, 7, 329–336. [Google Scholar] [CrossRef]
- Walker, B.H. Biodiversity and Ecological Redundancy. Conserv. Biol. 1992, 6, 18–23. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Dinerstein, E.; Olson, D.M.; Loucks, C.J.; Eichbaum, W.; DellaSala, D.A.; Kavanagh, K.; Hedao, P.; Hurley, P.; Carney, K.; et al. Terrestrial ecoregions of North America: A Conservation Assessment; Island Press: Washington, DC, USA, 1999. [Google Scholar]
- Maestre, F.T.; Bowker, M.A.; Puche, M.D.; Belen Hinojosa, M.; Martinez, I.; Garcia-Palacios, P.; Castillo, A.P.; Soliveres, S.; Luzuriaga, A.L.; Sanchez, A.M.; et al. Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands. Ecol. Lett. 2009, 12, 930–941. [Google Scholar] [CrossRef]
- Zhou, X.J.; An, X.L.; De Philippis, R.; Ye, C.R.; Ke, T.; Zhang, Y.R.; Chen, L.Z. The facilitative effects of shrub on induced biological soil crust development and soil properties. Appl. Soil Ecol. 2019, 137, 129–138. [Google Scholar] [CrossRef]
- Liu, W.S.; Qiu, K.Y.; Xie, Y.Z.; Huang, Y.Y.; Wang, R.X.; Li, H.C.; Meng, W.F.; He, Y.; Li, Y.Y.; Li, H.Q.; et al. High-Throughput absolute quantification sequencing reveals that a combination of leguminous shrubs is effective in driving soil bacterial diversity during the process of desertification reversal. Microb. Ecol. 2023, 86, 1145–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Y.; Wang, G.Z.; Yu, M.H.; Gao, G.L.; Ding, G.D. The leguminous Hedysarum shrubs effectively drive the diversity and structural composition of soil bacterial community through rhizocompartments in the process of desertification reversal. Land Degrad. Dev. 2023, 34, 4833–4846. [Google Scholar] [CrossRef]
- Zhao, X.F.; Xu, H.L.; Zhang, P.; Fu, J.Y.; Tu, W.X.; Zhang, Q.Q. The effects of nutrient addition on plant species diversity in desert grassland, Xinjiang, northwest China. Quat. Int. 2013, 298, 152–160. [Google Scholar] [CrossRef]
- Böcük, H.; Türe, C.; Ketenoğlu, O. Plant diversity and conservation of the Northeast Phrygia region under the impact of land degradation and desertification (Central Anatolia, Turkey). Pak. J. Bot. 2009, 41, 2305–2321. [Google Scholar]
- Elmberg, J.; Nummi, P.; Pöysä, H.; Sjöberg, K. Factors affecting species number and density of dabbling duck guilds in North Europe. Ecography 1993, 16, 251–260. [Google Scholar] [CrossRef]
- Gill, A.H.; Ahmad, K.S.; Habib, S.; Hameed, M.; Ahmad, M.S.A.; Nawaz, T.; Ahmad, F.; Batool, R. Impact of highly saline wetland ecosystem on floral diversity of the Cholistan Desert. Pak. J. Bot. 2012, 44, 107–112. [Google Scholar]
- Yang, X.D.; Zhang, X.N.; Lv, G.H.; Ali, A. Linking Populus euphratica hydraulic redistribution to diversity assembly in the arid desert zone of Xinjiang, China. PLoS ONE 2014, 9, e109071. [Google Scholar] [CrossRef]
- Fang, W.; Peng, S.L. Development of species diversity in the restoration process of establishing a tropical man-made forest ecosystem in China. For. Ecol. Manag. 1997, 99, 185–196. [Google Scholar] [CrossRef]
- Xie, Y.Z.; Qiu, K.Y.; Xu, D.M.; Shi, X.F.; Qi, T.Y.; Pott, R. Spatial heterogeneity of soil and vegetation characteristics and soil-vegetation relationships along an ecotone in Southern Mu Us Sandy Land, China. J. Soils Sediments 2015, 15, 1584–1601. [Google Scholar] [CrossRef]
- Chen, X.H.; Duan, Z.H. Changes in soil physical and chemical properties during reversal of desertification in Yanchi County of Ningxia Hui autonomous region, China. Environ. Geol. 2009, 57, 975–985. [Google Scholar] [CrossRef]
- Qiao, F.; Zhang, S.Y.; Zhang, K.B.; Yang, J.J.; Li, R.; Liu, G. The dynamic change of vegetation coverage in the Yanchi County monitoring based on remote sensing. Res. Soil Water Conserv. 2006, 13, 181–183, 186, (In Chinese with English Abstract). [Google Scholar]
- Li, J.Y.; Xu, B.; Yang, X.C.; Jin, Y.X.; Gao, T.; Yu, H.D.; Ma, H.L.; Qin, Z.H.; Zhao, L.N. Temporal and Spatial Variations of Grassland Desertification in Lingwu and Yanchi of Ningxia, China). In Proceedings of the Second International Conference on Agro-Geoinformatics, Fairfax, VA, USA, 12–16 August 2013; pp. 356–360. [Google Scholar]
- Institute of Soil Science; Chinese Academy of Sciences. Chinese Soil Taxonomy; USTC Press: Hefei, China, 2001. (In Chinese) [Google Scholar]
- Zhou, T.J.; Zhao, T.N. Study on analysis of land use change in Yanchi County. Res. Soil Water Conserv. 2005, 12, 116–118, (In Chinese with English Abstract). [Google Scholar]
- Zhou, L.H.; Yang, G.J.; Wang, T. Analysis on driving factors of desertification in northern China: A case study of Yanchi County). In Proceedings of the International Geoscience and Remote Sensing Symposium, Honolulu, HI, USA, 25–30 July 2010; pp. 2127–2130. [Google Scholar]
- Zhou, L.; Zhu, Y.; Yang, G.; Luo, Y. Quantitative evaluation of the effect of prohibiting grazing policy on grassland desertification reversal in northern China. Environ. Earth Sci. 2013, 68, 2181–2188. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, H.; Zhao, X.; Zhang, T.; Chen, Y. Biomass energy, carbon and nitrogen stores in different habitats along a desertification gradient in the semiarid Horqin Sandy Land. Arid Land Res. Manag. 2006, 20, 43–60. [Google Scholar] [CrossRef]
- Zhao, H.L.; Zhou, R.L.; Zhao, X.Y.; Zhang, T.H. Ground Discriminance on positive and negative processes of land desertification in Horqin Sand Land. J. Desert Res. 2008, 28, 8–15, (In Chinese with English Abstract). [Google Scholar]
- Li, S.; Yang, P.; Gao, S.; Chen, H.; Yao, F. Dynamic changes and developmental trends of the land desertification in Tibetan Plateau over the past 10 years. Adv. Earth Sci. 2004, 19, 63–70, (In Chinese with English Abstract). [Google Scholar]
- Li, X.R.; Jia, X.H.; Dong, G.R. Influence of desertification on vegetation pattern variations in the cold semi-arid grasslands of Qinghai-Tibet Plateau, North-west China. J. Arid Environ. 2006, 64, 505–522. [Google Scholar] [CrossRef]
- Verón, S.R.; Paruelo, J.M.; Oesterheld, M. Assessing desertification. J. Arid Environ. 2006, 66, 751–763. [Google Scholar] [CrossRef]
- Zhu, Z.D. Concept, cause and control of desertification in China. Quat. Sci. 1998, 18, 145–155, (In Chinese with English Abstract). [Google Scholar]
- Wang, H.Z.; Liu, C.Q.; Zhang, Y.L. Grassland Analysis and Production Design; China Agriculture Press: Beijing, China, 1997. (In Chinese) [Google Scholar]
- Song, Y.C. Vegetation Ecology; East China Normal University Press: Shanghai, China, 2001. (In Chinese) [Google Scholar]
- An, H.; Li, G.Q. Effects of grazing on carbon and nitrogen in plants and soils in a semiarid desert grassland, China. J. Arid Land 2014, 7, 341–349. [Google Scholar] [CrossRef]
- Cheng, J.M.; Jing, Z.B.; Jin, J.W.; Gao, Y. Restoration and utilization mechanism of degraded grassland in the semi-arid region of Loess Plateau. Sci. Sin. Vitae 2014, 44, 267–279, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Cui, Y.Q.; Ma, J.Y.; Sun, W.; Sun, J.H.; Duan, Z.H. A preliminary study of water use strategy of desert plants in Dunhuang, China. J. Arid Land 2015, 7, 73–81. [Google Scholar] [CrossRef]
- Feng, Y.; Duan, S.M.; Mu, S.Y.; Zhao, L.; Zhao, X.H. Geographic distribution and ecology of C4 plants in Xinjiang. Arid Land Geogr. 2012, 35, 145–153, (In Chinese with English Abstract). [Google Scholar]
- Jiang, G.M.; He, W.M. Species- and habitat-variability of photosynthesis, transpiration and water use efficiency of different plant species in Maowusu Sand Area. Acta Bot. Sin. 1999, 41, 1114–1124. [Google Scholar]
- Li, M.C.; Yi, X.F.; Zhang, X.A.; Li, L.X. The list of C4 plants in alpine locality of Qinghai Plateau. Acta Bot. Boreali-Occident. Sin. 2005, 25, 1046–1050, (In Chinese with English Abstract). [Google Scholar]
- Liu, X.Q.; Wang, R.Z.; Li, Y.Z. Photosynthetic pathway types in rangeland plant species from Inner Mongolia, North China. Photosynthetica 2004, 42, 339–344. [Google Scholar] [CrossRef]
- Ma, J.Y.; Sun, W.; Liu, X.N.; Chen, F.H. Variation in the stable carbon and nitrogen isotope composition of plants and soil along a precipitation gradient in northern China. PLoS ONE 2012, 7, e51894. [Google Scholar] [CrossRef]
- Niu, S.L.; Jiang, G.M.; Gao, L.M.; Li, Y.G.; Liu, M.Z. Comparison of gas exchange traits of different plant species in Hunshandak Sand Area. Acta Phytoecol. Sin. 2003, 27, 318–324. [Google Scholar]
- Su, P.X.; Xie, T.T.; Zhou, Z.J. Geographical distribution of C4 plant species in desert regions of China and its relation with climate factors. J. Desert Res. 2011, 31, 267–276, (In Chinese with English Abstract). [Google Scholar]
- Wang, G.A.; Mao, H.J. δ13C variations of C3 plants in dry and rainy seasons. Mar. Geol. Quat. Geol. 2001, 21, 43–47, (In Chinese with English Abstract). [Google Scholar]
- Wang, J. Along the 400 mm isopleth of mean annual precipitation in China. Mendeley Data 2017. [Google Scholar] [CrossRef]
- Wang, R.Z. Photosynthetic pathways and life forms in different grassland types from North China. Photosynthetica 2002, 40, 243–250. [Google Scholar] [CrossRef]
- Wang, R.Z. Photosynthetic pathways, life forms, and reproductive types for forage species along the desertification gradient on Hunshandake desert, North China. Photosynthetica 2002, 40, 321–329. [Google Scholar] [CrossRef]
- Wang, R.Z. Photosynthetic pathway and morphological functional types in the steppe vegetation from Inner Mongolia, North China. Photosynthetica 2003, 41, 143–150. [Google Scholar] [CrossRef]
- Wang, R.Z. Photosynthetic and morphological functional types from different steppe communities in Inner Mongolia, North China. Photosynthetica 2004, 42, 493–503. [Google Scholar] [CrossRef]
- Wang, R.Z. Photosynthetic pathways and life form types for native plant species from Hulunbeier Rangelands, Inner Mongolia, North China. Photosynthetica 2004, 42, 219–227. [Google Scholar] [CrossRef]
- Werger, M.J.A.; van Staalduinen, M.A. Eurasian steppes. In Ecological Problems and Livelihoods in a Changing World; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Yin, L.J.; Zhu, L. A preliminary study on C3 and C4 plants in the Northeast steppes and their ecological distribution. J. Appl. Ecol. 1990, 1, 237–242, (In Chinese with English Abstract). [Google Scholar]
- Zhang, B.; Ning, Y.F.; An, Z.S.; Liu, W.G. Abundance of C4/C3 plants in the Chinese Loess Plateau and their response to plant growing environment. Quat. Sci. 2015, 35, 801–808, (In Chinese with English Abstract). [Google Scholar]
- Zhang, C.J.; Chen, F.H.; Ming, J. Study on modern plant C-13 in western China and its Significance. Chin. J. Geochem. 2003, 22, 97–106. [Google Scholar]
- Zheng, S.X.; Shangguan, Z.P. Spatial patterns of foliar stable carbon isotope compositions of C3 plant species in the Loess Plateau of China. Ecol. Res. 2007, 22, 342–353. [Google Scholar] [CrossRef]
- Ziska, L.H. Evaluation of the growth response of six invasive species to past, present and future atmospheric carbon dioxide. J. Exp. Bot. 2003, 54, 395–404. [Google Scholar] [CrossRef]
- Myers, J.H.; Bazely, D.R. Ecology and Control of Introduced Plants; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Zhang, J.; Zhao, H.; Zhang, T.; Zhao, X.; Drake, S. Community succession along a chronosequence of vegetation restoration on sand dunes in Horqin Sandy Land. J. Arid Environ. 2005, 62, 555–566. [Google Scholar] [CrossRef]
- Belsky, A.J. Effects of grazing, competition, disturbance and fire on species composition and diversity in grassland communities. J. Veg. Sci. 1992, 3, 187–200. [Google Scholar] [CrossRef]
- Chawla, A.; Kumar, A.; Rajkumar, S.; Singh, R.D.; Thukral, A.K.; Ahuja, P.S. Correlation of multispectral satellite data with plant species diversity vis-à-vis soil characteristics in a landscape of Western Himalayan Region, India. Appl. Remote Sens. J. 2010, 1, 1–13. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Management; Princeton Univ. Press: Princeton, NJ, USA, 1998. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423, 623–656. [Google Scholar] [CrossRef]
- Lu, W.D. SPSS for Windows Statistics Analysis, 3rd ed.; Press of Electronics Industry: Beijing, China, 2003. (In Chinese) [Google Scholar]
- Du, F.; Sui, Y.Y.; Xu, X.X.; Zhang, X.C.; Hao, W.F.; Shao, M.G.; Hu, L.J.; Shan, L. Morphological responses of six plant species to patchy habitat with different nitrogen concentrations using in-growth core method. Pol. J. Ecol. 2013, 61, 257–269. [Google Scholar]
- Berg, N.; Steinberger, Y. Are biological effects of desert shrubs more important than physical effects on soil microorganisms? Microb. Ecol. 2010, 59, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Diedhiou, S.; Dossa, E.L.; Badiane, A.N.; Diedhiou, I.; Sène, M.; Dick, R.P. Decomposition and spatial microbial heterogeneity associated with native shrubs in soils of agroecosystems in semi-arid Senegal. Pedobiologia 2009, 52, 273–286. [Google Scholar] [CrossRef]
- Su, Y.Z.; Zhao, H.L. Soil properties and plant species in an age sequence of Caragana microphylla plantations in the Horqin Sandy Land, north China. Ecol. Eng. 2003, 20, 223–235. [Google Scholar] [CrossRef]
- Su, Y.; Zhang, T.; Li, Y.; Wang, F. Changes in soil properties after establishment of Artemisia halodendron and Caragana microphylla on shifting sand dunes in semiarid Horqin Sandy Land, northern China. Environ. Manag. 2005, 36, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.A.; Zhao, X.Y.; Zhao, H.L.; Zhang, T.H.; Guo, Y.R.; Li, Y.Q.; Huang, Y.X. Spatial heterogeneity of soil properties and vegetation-soil relationships following vegetation restoration of mobile dunes in Horqin Sandy Land, Northern China. Plant Soil 2009, 318, 153–167. [Google Scholar] [CrossRef]
- Alakukku, L. Soil compaction. In Ecosystem Health & Sustainable Agriculture, Book 1. Sustainable Agriculture; Jakobsson, C., Ed.; The Baltic University Programme, Uppsala University: Uppsala, Sweden, 2012. [Google Scholar]
- Valone, T.J. Effects of long-term livestock removal on water infiltration rate in arid grasslands. In Grasslands: Ecology, Management and Restoration; Schroder, H.G., Ed.; Nova Science Publishers: New York, NY, USA, 2008; pp. 81–91. [Google Scholar]
- Steffens, M.; Kölbl, A.; Totsche, K.U.; Kögel-Knabner, I. Grazing effects on soil chemical and physical properties in a semiarid steppe of Inner Mongolia (P.R. China). Geoderma 2008, 143, 63–72. [Google Scholar] [CrossRef]
- Pyankov, V.I.; Gunin, P.D.; Tsoog, S.; Black, C.C. C4 plants in the vegetation of Mongolia: Their natural occurrence and geographical distribution in relation to climate. Oecologia 2000, 123, 15–31. [Google Scholar] [CrossRef]
- Zhang, J.W. C4 plants, one of the ways of desertification control. Arid Zone Res. 2006, 23, 384–387, (In Chinese with English Abstract). [Google Scholar]
- de Carvalho, F.; Godoy, E.L.; Lisboa, F.J.G.; Moreira, F.M.D.; de Souza, F.A.; Berbara, R.L.L.; Fernandes, G.W. Relationship between physical and chemical soil attributes and plant species diversity in tropical mountain ecosystems from Brazil. J. Mt. Sci. 2014, 11, 875–883. [Google Scholar] [CrossRef]
- Loranger-Merciris, G.; Barthes, L.; Gastine, A.; Leadley, P. Rapid effects of plant species diversity and identity on soil microbial communities in experimental grassland ecosystems. Soil Biol. Biochem. 2006, 38, 2336–2343. [Google Scholar] [CrossRef]
- Guo, Y.R.; Zhao, H.L.; Zuo, X.A.; Drake, S.; Zhao, X.Y. Biological soil crust development and its topsoil properties in the process of dune stabilization, Inner Mongolia, China. Environ. Geol. 2007, 54, 653–662. [Google Scholar] [CrossRef]
- Su, Y.Z.; Zhao, W.Z.; Su, P.X.; Zhang, Z.H.; Wang, T.; Ram, R. Ecological effects of desertification control and desertified land reclamation in an oasis–desert ecotone in an arid region: A case study in Hexi Corridor, northwest China. Ecol. Eng. 2007, 29, 117–124. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhao, H.L.; Zhao, X.Y.; Zhang, T.H.; Li, Y.L.; Cui, J.Y. Effects of grazing and livestock exclusion on soil physical and chemical properties in desertified sandy grassland, Inner Mongolia, northern China. Environ. Earth Sci. 2010, 63, 771–783. [Google Scholar] [CrossRef]


{kind=link}
| Desertification Reversal Stage | ||||||
|---|---|---|---|---|---|---|
| Species | Life Form | VSD | SD | MD | LD | PD |
| Artemisia ordosica Krasch. | PS | 32.89 | 12.25 | 14.72 | 10.54 | 4.31 |
| Agriophyllum squarrosum (L.) Moq. | AF | 29.27 | 16.35 | 0.47 | ||
| Leymus secalinus (Georgi) Tzvel. | PG | 9.52 | 21.30 | 4.96 | 11.85 | 26.46 |
| Salsola beticolor Iljin | AF | 9.42 | 5.99 | 1.46 | 4.30 | 2.26 |
| Setaria viridis (L.) Beauv. | AG | 8.46 | 5.38 | 1.26 | 5.88 | 2.85 |
| Pennisetum flaccidum Griseb. | PG | 3.74 | 0.69 | 12.58 | 4.34 | 9.26 |
| Bassia scoparia (L.) A.J.Scott | AF | 2.85 | 3.43 | 0.81 | 2.29 | |
| Ixeris chinensis subsp. versicolor (Fisch. ex Link) Kitam. | PF | 2.18 | 3.68 | 6.62 | 9.94 | 4.11 |
| Panzerina lanata var. alaschanica (Kuprian.) H. W. Li | PF | 0.53 | 0.09 | 0.09 | ||
| Artemisia scoparia Waldst. et Kit. | AF | 0.41 | 1.01 | 19.42 | 18.98 | 15.80 |
| Euphorbia humifusa Willd. | AF | 0.30 | 1.32 | 0.75 | 0.18 | 0.82 |
| Chenopodium acuminatum Willd. | AF | 0.16 | 5.21 | 1.84 | 0.34 | |
| Tribulus terrestris L. | AF | 0.11 | 0.60 | |||
| Lespedeza potaninii Vass. | PS | 0.09 | 1.81 | 1.37 | 2.32 | 0.10 |
| Grubovia dasyphylla (Fisch. & C. A. Mey.) Freitag & G. Kadereit | AF | 0.07 | 2.06 | 0.48 | 0.46 | 0.88 |
| Oxytropis racemosa Turcz. | PL | 8.06 | 0.25 | 0.31 | ||
| Sophora alopecuroides L. | PS | 6.77 | 1.65 | 1.19 | 1.04 | |
| Cleistogenes squarrosa (Trin.) Keng | PG | 1.89 | 5.20 | 2.13 | 7.09 | |
| Astragalus melilotoides Pall. | PL | 1.85 | 0.98 | 2.82 | 0.58 | |
| Salix psammophila C.Wang et Ch.Y. Yang | SH | 0.96 | ||||
| Corethrodendron fruticosum var. mongolicum (Turczaninow) Turczaninow ex Kitagawa | PS | 0.74 | ||||
| Agropyron mongolicum Keng | PG | 1.03 | 0.66 | 2.91 | 8.44 | |
| Oxytropis aciphylla Ledeb. | PS | 0.69 | ||||
| Cynanchum thesioides (Freyn) K. Schum | PF | 0.54 | 0.13 | |||
| Gueldenstaedtia stenophylla Bge. | PL | 0.53 | 0.72 | 0.19 | ||
| Inula salsoloides (Turcz.) Ostenf. | PF | 0.32 | ||||
| Vincetoxicum mongolicum Maxim. | PF | 0.24 | 0.27 | 0.34 | ||
| Olgaea leucophylla (Turcz.) Iljin | PF | 0.13 | ||||
| Euphorbia esula L. | PF | 0.13 | 0.81 | 0.09 | ||
| Polygala tenuifolia Willd. | PF | 0.09 | ||||
| Echinops gmelinii Turcz. | AF | 0.06 | 0.32 | 0.29 | 0.59 | |
| Chenopodium album L. | AF | 4.13 | 1.14 | 0.94 | ||
| Stipa breviflora Griseb. | PG | 3.62 | 4.46 | 0.98 | ||
| Thermopsis lanceolata R. Br. | PL | 3.57 | 1.79 | 0.20 | ||
| Aster altaicus Willd. | PF | 2.25 | 1.88 | 4.66 | ||
| Setaria arenaria Kitag. | AG | 1.46 | 0.08 | 1.13 | ||
| Cuscuta chinensis Lam. | AF | 1.36 | ||||
| Glycyrrhiza uralensis Fisch. | PL | 1.14 | 1.74 | 1.93 | ||
| Equisetum ramosissimum Desf. | PF | 0.94 | ||||
| Polygonum aviculare L. | AF | 0.40 | ||||
| Stipa bungeana Trin. | PG | 0.16 | 1.71 | |||
| Plantago asiatica L. | PF | 0.12 | 0.11 | |||
| Incarvillea sinensis Lam. | AF | 0.11 | 0.19 | |||
| Kalidium foliatum (Pall.) Moq. | SH | 1.27 | ||||
| Artemisia sacrorum Ledeb. | PF | 1.24 | ||||
| Limonium aureum (L.) Hill. ex Kuntze | PF | 1.00 | ||||
| Peganum nigellastrum Bge. | PF | 0.91 | ||||
| Typha minima Funk. | PF | 0.54 | 0.88 | |||
| Sonchus arvensis L. | PF | 0.34 | ||||
| Lactuca tatarica (L.) C. A. Mey. | PF | 0.09 | ||||
| Artemisia dubia Wall. ex Bess. var. subdigitata (Mattf.) Y. R. Ling | PF | 1.37 | ||||
| Artemisia mongolica Fisch. ex Bess. | PF | 0.32 | ||||
| Silene conoidea L. | AF | 0.27 | ||||
| Oxybasis glauca (L.) S. Fuentes, Uotila & Borsch | AF | 0.23 | ||||
| Desertification Reversal Stage | ||||||
|---|---|---|---|---|---|---|
| Life Form | VSD | SD | MD | LD | PD | |
| AF | S | 8 | 8 | 11 | 10 | 9 |
| IV | 42.59 | 30.82 | 34.45 | 30.14 | 22.13 | |
| AG | S | 1 | 1 | 2 | 2 | 2 |
| IV | 8.46 | 5.38 | 2.72 | 5.96 | 3.98 | |
| PF | S | 2 | 8 | 8 | 9 | 7 |
| IV | 2.71 | 5.22 | 11.23 | 16.03 | 11.79 | |
| PG | S | 2 | 4 | 6 | 5 | 6 |
| IV | 13.26 | 24.91 | 27.18 | 25.69 | 53.94 | |
| PL | S | 0 | 3 | 5 | 5 | 3 |
| IV | 0.00 | 10.44 | 6.66 | 6.85 | 2.71 | |
| PS | S | 2 | 5 | 3 | 3 | 3 |
| IV | 32.98 | 22.26 | 17.74 | 14.05 | 5.54 | |
| SH | S | 0 | 1 | 0 | 1 | 0 |
| IV | 0.00 | 0.96 | 0.00 | 1.27 | 0.00 | |
| Desertification Reversal Stage | Species | Family | C3/C4 | PFTs | |
|---|---|---|---|---|---|
| VSD and SD | Early stages | Tribulus terrestris L. | Zygophyllaceae | C4 | Forb |
| SD | Salix psammophila C.Wang et Ch.Y. Yang | Salicaceae | C3 | Shrub | |
| SD | Corethrodendron fruticosum var. mongolicum (Turczaninow) Turczaninow ex Kitagawa | Fabaceae | C3 | Legume/sub-shrub | |
| SD | Oxytropis aciphylla Ledeb. | Fabaceae | C3 | Legume/sub-shrub | |
| SD | Inula salsoloides (Turcz.) Ostenf. | Asteraceae | C3 | Forb | |
| SD | Olgaea leucophylla (Turcz.) Iljin | Asteraceae | C3 | Forb | |
| SD | Polygala tenuifolia Willd. | Polygalaceae | C4 | Forb | |
| SD MD | Cynanchum thesioides (Freyn) K. Schum | Asclepiadaceae | C3 | Forb | |
| VSD SD MD | Panzerina lanata var. alaschanica (Kuprian.) H. W. Li | Labiatae | C3 | Forb | |
| VSD SD LD | Agriophyllum squarrosum (L.) Moq. | Amaranthaceae | C4 | Forb | |
| MD | Middle stages | Cuscuta chinensis Lam. | Convolvulaceae | C4 | Forb |
| MD | Equisetum ramosissimum Desf. | Equisetaceae | C3 | Forb | |
| MD | Polygonum aviculare L. | Polygonaceae | C3 | Forb | |
| SD MD LD | Oxytropis racemosa Turcz. | Fabaceae | C3 | Legume | |
| SD MD LD | Gueldenstaedtia stenophylla Bge. | Fabaceae | C3 | Legume | |
| SD MD LD | Euphorbia esula L. | Euphorbiaceae | C3 | Forb | |
| MD LD | Later stages | Incarvillea sinensis Lam. | Bignoniaceae | C3 | Forb |
| LD | Kalidium foliatum (Pall.) Moq. | Amaranthaceae | C4 | Shrub | |
| LD | Artemisia sacrorum Ledeb. | Asteraceae | C3 | Forb | |
| LD | Limonium aureum (L.) Hill. ex Kuntze | Plumbaginaceae | C3 | Forb | |
| LD | Peganum nigellastrum Bge. | Zygophyllaceae | C3 | Forb | |
| LD | Sonchus arvensis L. | Asteraceae | C3 | Forb | |
| LD | Lactuca tatarica (L.) C. A. Mey. | Asteraceae | C3 | Forb | |
| LD PD | Typha minima Funk. | Typhaceae | C3 | Forb | |
| PD | Artemisia dubia Wall. ex Bess. var. subdigitata (Mattf.) Y. R. Ling | Asteraceae | — | Forb | |
| PD | Artemisia mongolica Fisch. ex Bess. | Asteraceae | C3 | Forb | |
| PD | Silene conoidea L. | Caryophyllaceae | C3 | Forb | |
| PD | Oxybasis glauca (L.) S. Fuentes, Uotila & Borsch | Amaranthaceae | C4 | Forb | |
| SD MD PD | Vincetoxicum mongolicum Maxim. | Asclepiadaceae | C3 | Forb | |
| MD PD | Stipa bungeana Trin. | Poaceae | C3 | Grass | |
| MD PD | Plantago asiatica L. | Plantaginaceae | C3 | Forb | |
| MD LD PD | Chenopodium album L. | Amaranthaceae | C4 | Forb | |
| MD LD PD | Stipa breviflora Griseb. | Poaceae | C3 | Grass | |
| MD LD PD | Thermopsis lanceolata R. Br. | Fabaceae | C3 | Legume | |
| MD LD PD | Aster altaicus Willd. | Asteraceae | C3 | Forb | |
| MD LD PD | Setaria arenaria Kitag. | Poaceae | C4 | Grass | |
| MD LD PD | Glycyrrhiza uralensis Fisch. | Fabaceae | C3 | Legume | |
| VSD SD MD LD | Common stages | Bassia scoparia (L.) A.J.Scott | Amaranthaceae | C4 | Forb |
| SD MD LD PD | Sophora alopecuroides L. | Fabaceae | C3 | Legume/Sub-shrub | |
| SD MD LD PD | Cleistogenes squarrosa (Trin.) Keng | Poaceae | C4 | Grass | |
| SD MD LD PD | Astragalus melilotoides Pall. | Fabaceae | C3 | Legume | |
| SD MD LD PD | Agropyron mongolicum Keng | Poaceae | C3 | Grass | |
| SD MD LD PD | Echinops gmelinii Turcz. | Asteraceae | C3 | Forb | |
| VSD MD LD PD | Chenopodium acuminatum Willd. | Amaranthaceae | C4 | Forb | |
| VSD SD MD LD PD | Artemisia ordosica Krasch. | Asteraceae | C4 | Sub-shrub | |
| VSD SD MD LD PD | Leymus secalinus (Georgi) Tzvel. | Poaceae | C3 | Grass | |
| VSD SD MD LD PD | Salsola beticolor Iljin | Amaranthaceae | C4 | Forb | |
| VSD SD MD LD PD | Setaria viridis (L.) Beauv. | Poaceae | C4 | Grass | |
| VSD SD MD LD PD | Pennisetum flaccidum Griseb. | Poaceae | C4 | Grass | |
| VSD SD MD LD PD | Ixeris chinensis subsp. versicolor (Fisch. ex Link) Kitam. | Asteraceae | C3 | Forb | |
| VSD SD MD LD PD | Artemisia scoparia Waldst. et Kit. | Asteraceae | C3 | Forb | |
| VSD SD MD LD PD | Euphorbia humifusa Willd. | Euphorbiaceae | C4 | Forb | |
| VSD SD MD LD PD | Lespedeza potaninii Vass. | Fabaceae | C3 | Legume/sub-shrub | |
| VSD SD MD LD PD | Grubovia dasyphylla (Fisch. & C. A. Mey.) Freitag & G. Kadereit | Amaranthaceae | C4 | Forb | |
| Desertification Reversal Stage | ||||||
|---|---|---|---|---|---|---|
| Plant Functional Type | VSD | SD | MD | LD | PD | |
| C3 plant | S | 4 | 18 | 22 | 22 | 18 |
| S% | 26.67 | 60.00 | 62.86 | 62.86 | 60.00 | |
| IV | 12.20 | 49.85 | 50.47 | 65.07 | 68.52 | |
| C4 plant | S | 10 | 11 | 12 | 13 | 11 |
| S% | 66.67 | 36.67 | 34.29 | 37.14 | 36.67 | |
| IV | 87.27 | 50.05 | 49.42 | 34.92 | 30.11 | |
| Legume a | S | 1 | 7 | 7 | 7 | 5 |
| S% | 6.67 | 23.33 | 20.00 | 20.00 | 16.67 | |
| IV | 0.09 | 20.45 | 9.68 | 10.36 | 3.85 | |
| Grass | S | 3 | 5 | 8 | 7 | 8 |
| S% | 20.00 | 16.67 | 22.86 | 20.00 | 26.67 | |
| IV | 21.72 | 30.29 | 29.90 | 31.65 | 57.92 | |
| Forb | S | 10 | 16 | 19 | 19 | 16 |
| S% | 66.67 | 53.33 | 54.29 | 54.29 | 53.33 | |
| IV | 45.30 | 36.04 | 45.68 | 46.17 | 33.92 | |
| Sub-shrub | S | 2 | 6 | 3 | 5 | 3 |
| S% | 13.33 | 20.00 | 8.57 | 14.29 | 10.00 | |
| IV | 32.98 | 23.22 | 17.74 | 16.51 | 5.45 | |
| Study Site | Reversal Stage | Species Richness | Species Diversity |
|---|---|---|---|
| NHZ | 1 | 2.3 ± 0.6 a | 0.67 ± 0.04 a |
| 2 | 7.0 ± 2.0 b | 1.08 ± 0.51 ab | |
| 3 | 9.3 ± 0.6 b | 1.05 ± 0.16 ab | |
| 4 | 8.0 ± 2.6 b | 1.51 ± 0.13 b | |
| HB | 1 | 2.3 ± 0.6 a | 0.56 ± 0.26 a |
| 2 | 4.0 ± 1.0 b | 1.02 ± 0.19 ab | |
| 3 | 7.7 ± 1.2 c | 1.02 ± 0.59 ab | |
| 4 | 7.7 ± 1.5 c | 1.19 ± 0.21 b | |
| 5 | 8.7 ± 2.9 c | 1.01 ± 0.31 ab | |
| HBN | 1 | 2.7 ± 0.6 a | 0.92 ± 0.25 ab |
| 2 | 4.3 ± 2.5 ac | 0.82 ± 0.81 a | |
| 3 | 9.7 ± 1.5 bd | 2.01 ± 0.10 b | |
| 4 | 5.7 ± 1.5 c | 1.13 ± 0.28 ab | |
| 5 | 11.3 ± 1.2 d | 1.62 ± 0.66 ab | |
| YZZ | 1 | 6.3 ± 1.2 a | 0.88 ± 0.25 a |
| 2 | 8.0 ± 1.0 a | 0.50 ± 0.10 a | |
| 3 | 7.7 ± 1.5 a | 0.65 ± 0.33 a | |
| 4 | 10.3 ± 1.2 b | 1.66 ± 0.13 b | |
| MC | 1 | 4.3 ± 1.5 a | 0.90 ± 0.23 a |
| 2 | 6.0 ± 1.0 ac | 1.13 ± 0.29 a | |
| 3 | 10.3 ± 1.2 b | 1.28 ± 0.06 a | |
| 4 | 7.3 ± 1.5 c | 0.90 ± 0.33 a | |
| WZZ | 1 | 1.0 ± 0.0 a | 0.00 ± 0.00 a |
| 2 | 5.7 ± 2.1 b | 0.89 ± 0.37 b | |
| 4 | 5.7 ± 2.1 b | 0.71 ± 0.13 b | |
| 5 | 9.0 ± 1.0 c | 1.47 ± 0.30 c | |
| LJHZ | 1 | 2.3 ± 1.2 a | 0.07 ± 0.07 a |
| 2 | 5.0 ± 1.0 bc | 0.68 ± 0.24 b | |
| 3 | 3.7 ± 1.5 ab | 0.22 ± 0.26 ac | |
| 4 | 5.3 ± 0.6 bc | 1.42 ± 0.14 bd | |
| 5 | 7.0 ± 1.7 c | 0.80 ± 0.52 bd | |
| All study sites | 1 | 3.0 ± 1.8 a | 0.57 ± 0.40 a |
| 2 | 5.7 ± 1.9 b | 0.87 ± 0.41 b | |
| 3 | 8.1 ± 2.5 cd | 1.04 ± 0.62 bc | |
| 4 | 7.1 ± 2.2 c | 1.22 ± 0.37 c | |
| 5 | 9.0 ± 2.3 d | 1.22 ± 0.53 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, K.; Li, Z.; Xie, Y.; Xu, D.; He, C.; Pott, R. Desertification Reversal Promotes the Complexity of Plant Community by Increasing Plant Species Diversity of Each Plant Functional Type. Agronomy 2024, 14, 96. https://doi.org/10.3390/agronomy14010096
Qiu K, Li Z, Xie Y, Xu D, He C, Pott R. Desertification Reversal Promotes the Complexity of Plant Community by Increasing Plant Species Diversity of Each Plant Functional Type. Agronomy. 2024; 14(1):96. https://doi.org/10.3390/agronomy14010096
Chicago/Turabian StyleQiu, Kaiyang, Zhigang Li, Yingzhong Xie, Dongmei Xu, Chen He, and Richard Pott. 2024. "Desertification Reversal Promotes the Complexity of Plant Community by Increasing Plant Species Diversity of Each Plant Functional Type" Agronomy 14, no. 1: 96. https://doi.org/10.3390/agronomy14010096
APA StyleQiu, K., Li, Z., Xie, Y., Xu, D., He, C., & Pott, R. (2024). Desertification Reversal Promotes the Complexity of Plant Community by Increasing Plant Species Diversity of Each Plant Functional Type. Agronomy, 14(1), 96. https://doi.org/10.3390/agronomy14010096