Allelopathy—A Tool to Improve the Weed Competitive Ability of Wheat with Herbicide-Resistant Black-Grass (Alopecurus myosuroides Huds.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
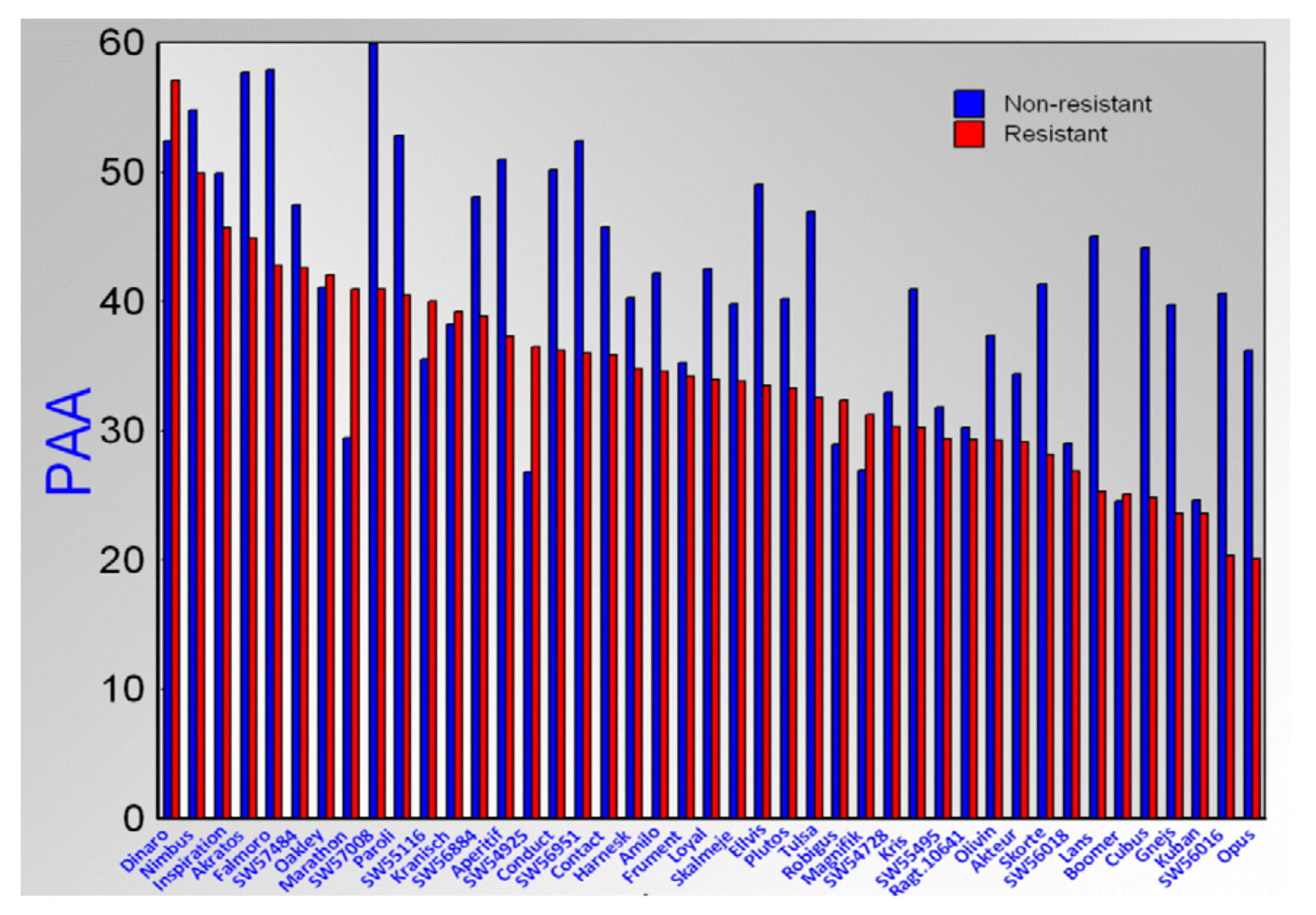
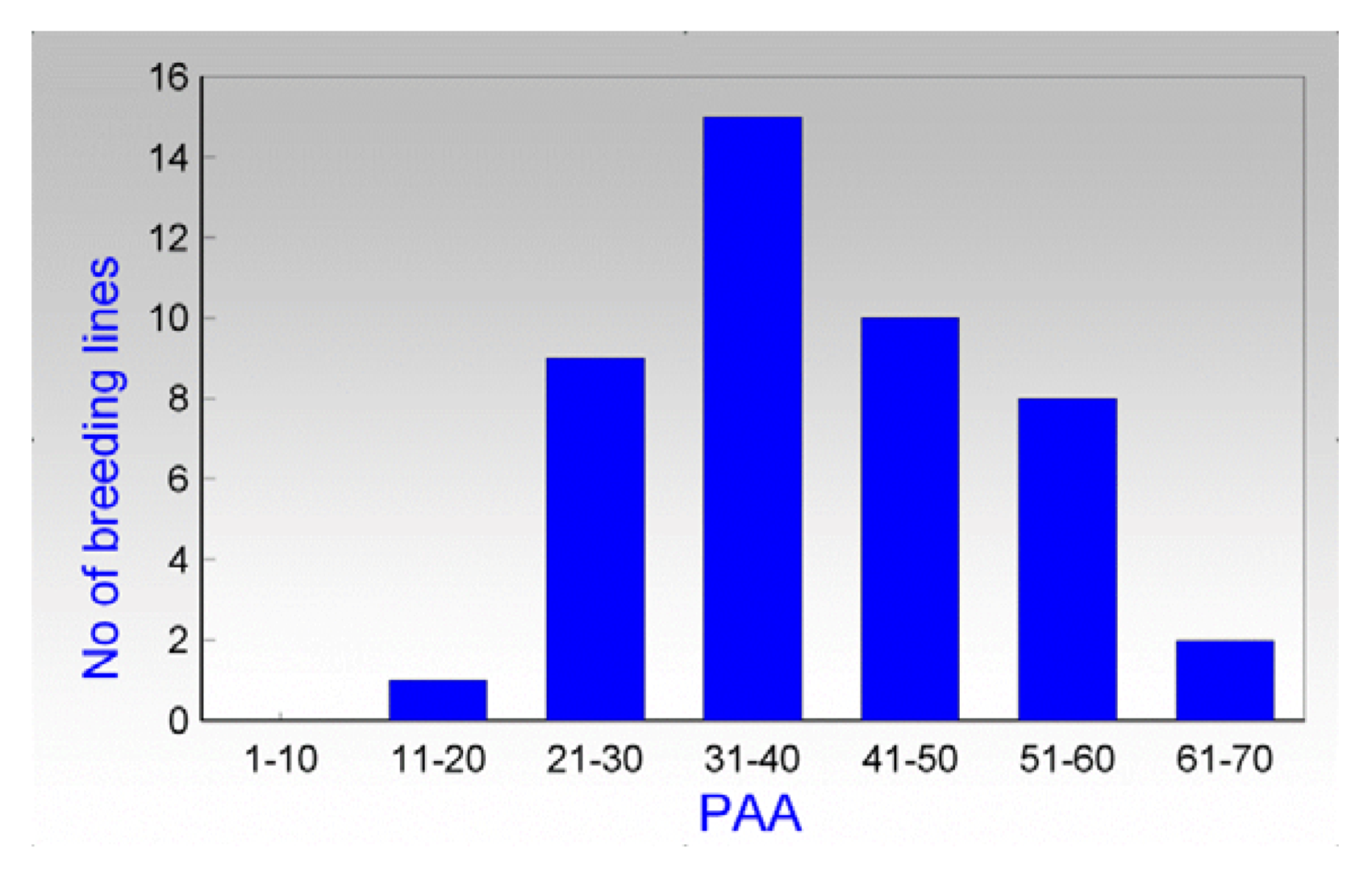
2.1. Allelopathic Potential of Modern Wheat, Triticale, Rye and Wheat Breeding Lines


2.2. Allelopathic Potential of Wheat-Rye Translocation Lines

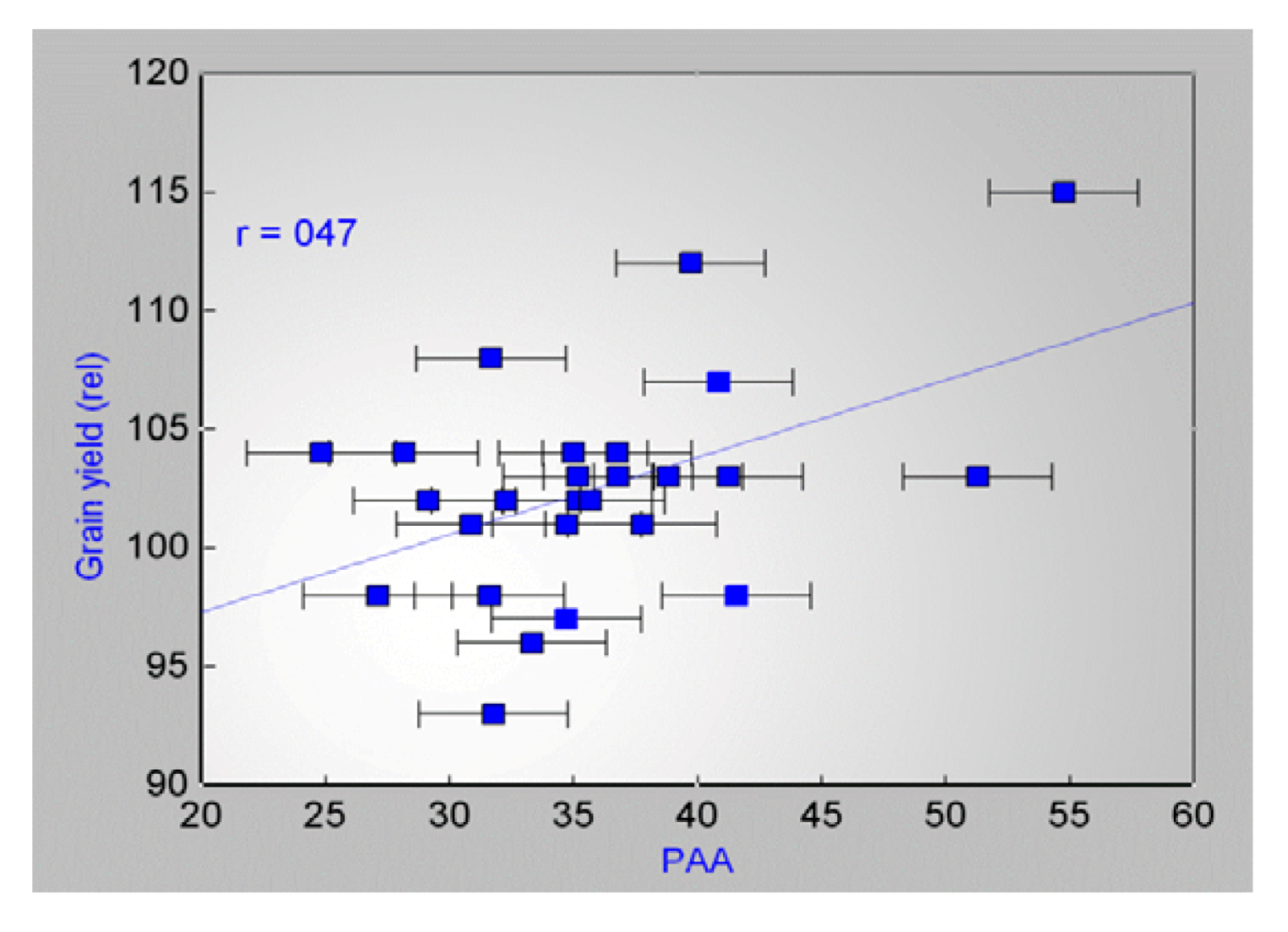
2.3. Field Studies


3. Experimental Section
3.1. Materials
3.2. Methods
3.2.1. Evaluation of Allelopathic Activity
3.2.2. Field Studies
3.2.3. Data analysis
4. Conclusions
Acknowledgments
References
- Moss, S.R.; Perryman, S.A.M.; Tatnell, L.V. Managing herbicide-resistant blackgrass (Alopecurus myosuroides): Theory and practice. Weed Technol. 2007, 21, 300–309. [Google Scholar] [CrossRef]
- Mennan, H.; Isik, D. The competitive ability of Avena spp. and Alopecurus myosuroides Huds. influenced by different wheat (Triticum aestivum) cultivars. Turk. J. Agric. For. 2004, 28, 245–251. [Google Scholar]
- Marczewska-Kolasa, K.; Bortniak, M.; Domaradzki, K. Allelopathy effect of Alopecurus myosuroides on root growth of winter wheat. Progr. Plant Prot. 2010, 50, 820–822. [Google Scholar]
- Kazinczi, G.; Mikulas, J.; Hunyadi, K.; Horvath, J. Allelopathic effects of weeds on growth of wheat, sugarbeet and Brassica napus. Allelopath. J. 1997, 4, 335–339. [Google Scholar]
- Naylor, R.L. The nature and consequences of interference by Alopecurus myosuroides Huds. on the growth of winter wheat. Weed Res. 1972, 12, 137–143. [Google Scholar] [CrossRef]
- Hall, L.M.; Moss, S.R.; Powles, B. Mechanisms of resistance to Aryloxyphenoxypropionate herbicides in two resistant biotypes of Alopecurus myosuroides (blackgrass): Herbicide metabolism as a cross-resistance mechanism. Pestic. Biochem. Physiol. 1997, 57, 87–98. [Google Scholar] [CrossRef]
- Moss, R.S.; Cocker, K.M.; Brown, A.C.; Hall, L.; Field, L.M. Characterisation of target-site resistance to ACCase-inhibiting herbicides in the weed Alopecurus myosuroides (black-grass). Pest Manag. Sci. 2003, 59, 190–201. [Google Scholar] [CrossRef]
- Marshall, R.; Moss, S.R. Characterisation and molecular basis of ALS inhibitor resistance in the grass weed Alopecurus myosuroides. Weed Res. 2007, 48, 439–447. [Google Scholar] [CrossRef]
- Beckie, H.J. Herbicide-resistant weeds: Management tactics and practices. Weed Technol. 2006, 20, 793–814. [Google Scholar]
- Llewellyn, R.S.; Lindner, R.K.; Pannell, D.J.; Powles, S.B. Herbicide resistance and adoption of integrated weed management by Western Australian grain growers. Agric. Econ. 2007, 36, 123–130. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T. Evaluation of seedling allelopathy in 453 wheat (Triticum aestivum) accessions against annual ryegrass (Lolium rigidum) by the equal-compartment-agar method. Aust. J. Agric. Res. 2000, 51, 937–944. [Google Scholar] [CrossRef]
- Lemerle, D.; Gill, G.S.; Murphy, C.E.; Walker, S.R.; Cousens, R.D.; Mokhtari, S.; Peltzer, D.J.; Coleman, R.; Luckett, D. Genetic improvement and agronomy for enhanced wheat competitiveness with weeds. Aust. J. Agric. Res. 2001, 152, 527–548. [Google Scholar]
- Bertholdsson, N.-O. Use of multivariate statistics to separate allelopathic and competitive factors influencing weed suppression ability in winter wheat. Weed Res. 2011, 51, 273–283. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; An, M.; Liu, D.L. Autotoxicity of wheat (Triticum aestivum L.) as determined by laboratory bioassays. Plant Soil 2007, 296, 85–96. [Google Scholar] [CrossRef]
- Belz, R.G. Allelopathy in crop/weed interactions—An update. Pest Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef]
- Przepiokowski, T.; Gorski, S.F. Influence of rye (Secale cereale) plant residues on germination and growth of three triazine-resistant and susceptible weeds. Weed Technol. 1994, 8, 744–747. [Google Scholar]
- Wu, H.; Pratley, J.; Haig, T. Phytotoxic effects of wheat extracts on a herbicide-resistant biotype of annual ryegrass (Lolium rigidum). J. Agric. Food Chem. 2003, 51, 4610–4616. [Google Scholar] [CrossRef]
- Bertholdsson, N.-O.; Andersson, S.C.; Merker, A. Allelopathic potential of Triticum spp., Secale spp. and Triticosecale and use of chromosome substitutions and translocations to improve weed suppression ability in winter wheat. Plant Breed. 2012, 131, 75–80. [Google Scholar]
- Barnes, L.P.; Putnam, A.R. Evidence for allelopathy by residues and aqueous extract of rye (Secale cereal L.). Weed Sci. 1986, 34, 384–390. [Google Scholar]
- Burgos, N.R.; Talbert, R.E. Differential activity of allelochemicals from Secale cereal in seedling bioassays. Weed Sci. 2000, 48, 302–310. [Google Scholar] [CrossRef]
- Reberg-Horton, S.C.; Burton, J.D.; Danehower, D.A.; Ma, G.; Monks, D.W.; Murphy, J.P.; Ranells, N.N.; Williamson, J.D.; Creamer, N.G. Changes over time in the allelochemicals content of ten cultivars of rye (Secale cereale L.). J. Chem. Ecol. 2005, 31, 179–193. [Google Scholar] [CrossRef]
- Macias, F.A.; Molinillo, J.M.G.; Varela, R.M.; Galindo, C.G. Allelopathy—A natural alternative for weed control. Pest Manag. Sci. 2007, 63, 327–348. [Google Scholar]
- Gressel, J. Evolving understanding of the evolution of herbicide resistance. Pest Manag. Sci. 2009, 64, 1164–1173. [Google Scholar] [CrossRef]
- Seal, A.N.; Pratley, J.E.; Haig, T. Can results from a laboratory bioassay be used as an indicator of field performance of rice cultivars with allelopathic potential against Damasonium minus (starfruit). Aust. J. Agric. Res. 2008, 59, 183–188. [Google Scholar] [CrossRef]
- Bertholdsson, N.-O. Early vigour and allelopathy—Two useful traits for enhanced barley and wheat competitiveness with weeds. Weed Res. 2005, 45, 94–102. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bertholdsson, N.-O. Allelopathy—A Tool to Improve the Weed Competitive Ability of Wheat with Herbicide-Resistant Black-Grass (Alopecurus myosuroides Huds.). Agronomy 2012, 2, 284-294. https://doi.org/10.3390/agronomy2040284
Bertholdsson N-O. Allelopathy—A Tool to Improve the Weed Competitive Ability of Wheat with Herbicide-Resistant Black-Grass (Alopecurus myosuroides Huds.). Agronomy. 2012; 2(4):284-294. https://doi.org/10.3390/agronomy2040284
Chicago/Turabian StyleBertholdsson, Nils-Ove. 2012. "Allelopathy—A Tool to Improve the Weed Competitive Ability of Wheat with Herbicide-Resistant Black-Grass (Alopecurus myosuroides Huds.)" Agronomy 2, no. 4: 284-294. https://doi.org/10.3390/agronomy2040284
APA StyleBertholdsson, N. -O. (2012). Allelopathy—A Tool to Improve the Weed Competitive Ability of Wheat with Herbicide-Resistant Black-Grass (Alopecurus myosuroides Huds.). Agronomy, 2(4), 284-294. https://doi.org/10.3390/agronomy2040284