Dissecting Wheat Grain Yield Drivers in a Mapping Population in the UK

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Field Experiments
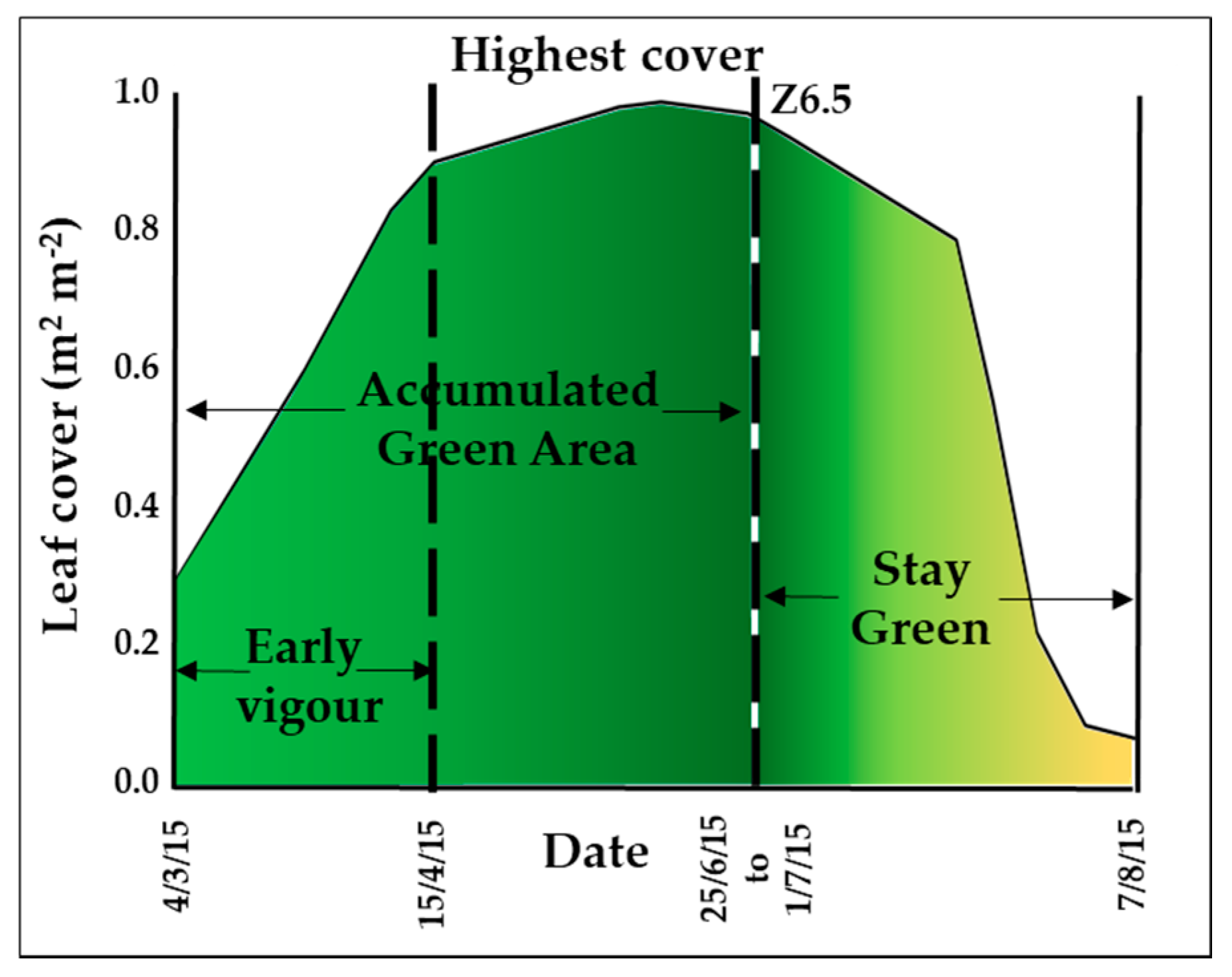
2.2. Crop Development
2.3. Phenotyping
2.4. Statistical Analysis
2.5. Heritability
2.6. Genetic Variation
3. Results
3.1. Traits within the Same Group of Measurements Presented High Collinearity
3.2. Correlation to Grain Yield Varied at Different Developmental Stages
3.2.1. Leaf and Canopy
3.2.2. Biomass
3.2.3. Reflectance Indices
3.2.4. Gas-Exchange
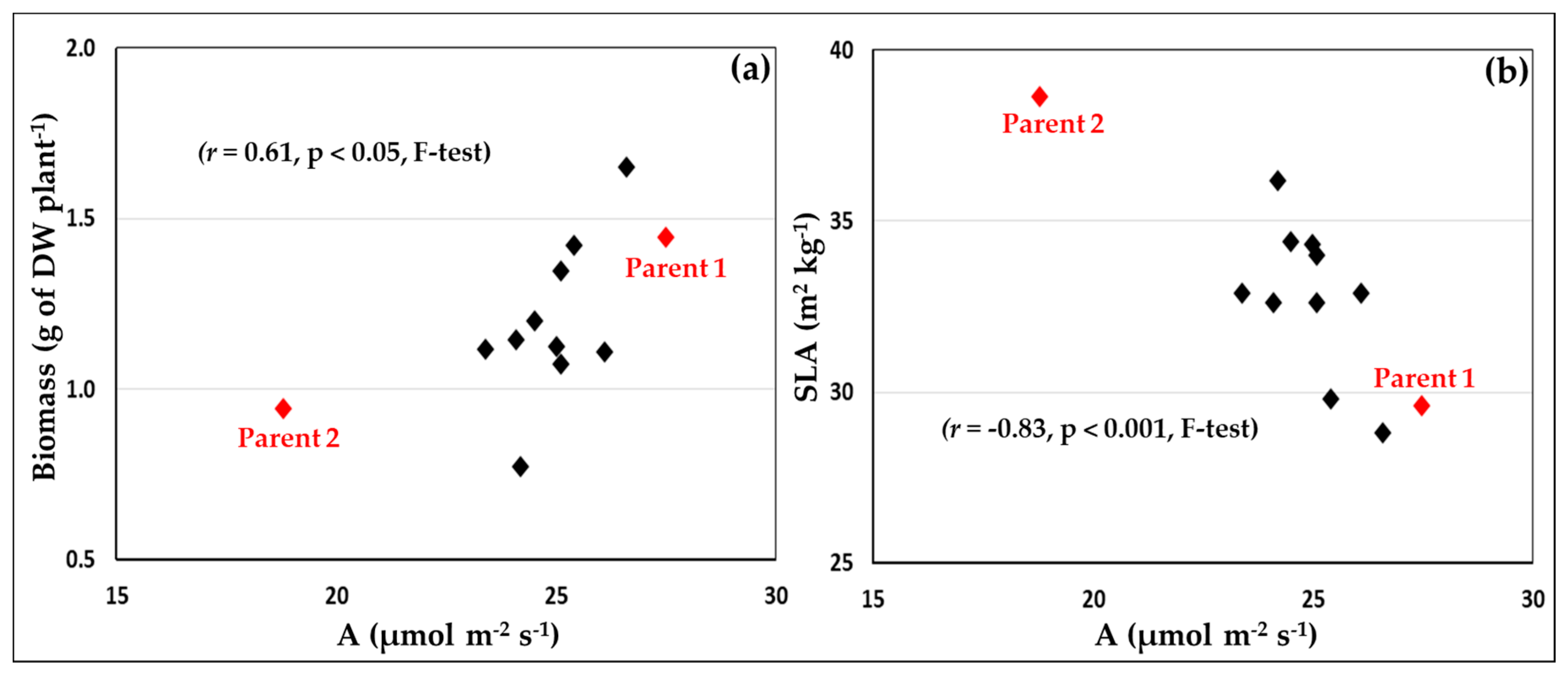
3.3. High Yielding Lines Presented Different Strategies to Build Grain Yield
4. Discussion
4.1. Main Drivers of Yield
4.2. Photosynthesis and Yield
4.3. Canopy vs. Leaf Photosynthesis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- FAO. The State of Food Insecurity in the World; ESA Working Paper; FAO: Rome, Italy, 2002. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Pingali, P.L. Westernization of Asian diets and the transformation of food systems: Implications for research and policy. Food Policy 2006, 32, 281–298. [Google Scholar] [CrossRef]
- Tilman, D.; Clark, M. Food, agriculture & the environment: Can we feed the world & save the earth? Am. Acad. Arts Sci. 2015, 144, 1–23. [Google Scholar]
- Berry, E.M.; Dernini, S.; Burlingame, B.; Meybeck, A.; Conforti, P. Food security and sustainability: Can one exist without the other? Public Health Nutr. 2015, 18, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; ESA Working Paper No. 12-03; Agricultural Development Economics Division, Food and Agriculture Organization of the United Nations: Rome, Italy, 2012. [Google Scholar]
- Grassini, P.; Eskridge, K.M.; Cassman, K.G. Distinguishing between yield advances and yield plateaus in historical crop production trends. Nat. Commun. 2013, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT Food and Agriculture Organization of the United Nations Database. Available online: http://www.fao.org/faostat/en/#home (accessed on 23 December 2016).
- Knight, S.; Kightley, S.; Bingham, I.; Hoad, S.; Lang, B.; Philpott, H.; Stobart, R.; Thomas, J.; Barnes, A.; Ball, B. Desk Study to Evaluate Contributory Causes of the Current ‘Yield Plateau’ in Wheat and Oilseed Rape; HGCA Project Report No. 502; AHDB: Kenilworth, UK, 2012. [Google Scholar]
- Reynolds, M.; Langridge, P. Physiological breeding. Curr. Opin. Plant Biol. 2016, 31, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Monteith, J.L.; Moss, C.J. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. Lond. B 1977, 281, 277–294. [Google Scholar] [CrossRef]
- Long, S.P.; Marshall-Colon, A.; Zhu, X.G. Meeting the global food demand of the future by engineering crop photosynthesis and yield potential. Cell 2015, 161, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Slafer, G.A.; Royo, C.; Serret, M.D. Breeding for yield potential and stress adaptation in cereals. Crit. Rev. Plant Sci. 2008, 27, 377–412. [Google Scholar] [CrossRef]
- Richards, R.A. Selectable traits to increase crop photosynthesis and yield of grain crops. J. Exp. Bot. 2000, 51, 447–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foulkes, M.J.; Slafer, G.A.; Davies, W.J.; Berry, P.M.; Sylvester-Bradley, R.; Martre, P.; Calderini, D.F.; Griffiths, S.; Reynolds, M.P. Raising yield potential of wheat. III. Optimizing partitioning to grain while maintaining lodging resistance. J. Exp. Bot. 2011, 62, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Medlyn, B.E. Physiological basis of the light use efficiency model. Tree Physiol. 1998, 18, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araus, J.L.; Slafer, G.A.; Reynolds, M.P.; Royo, C. Plant breeding and drought in C3 cereals: What should we breed for? Ann. Bot. 2002, 89, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Long, S.P.; Ort, D.R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr. Opin. Biotechnol. 2008, 19, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Raines, C.A. Increasing photosynthetic carbon assimilation in C3 plants to improve crop yield: Current and future strategies. Plant Physiol. 2011, 155, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Mir, R.R.; Zaman-Allah, M.; Sreenivasulu, N.; Trethowan, R.; Varshney, R.K. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor. Appl. Genet. 2012, 125, 625–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avery, B.W.; Catt, J.A. The Soil at Rothamsted; Map prepared by E M Thompson and the Soil Survey and Land Research Centre; Cranfield University, Lawes Agricultural Trust: Harpenden, UK, 1995. [Google Scholar]
- Zadoks, I.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Pask, A.; Pietragalla, J. Leaf area, green crop area and senescence. In Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Mexico City, Mexico, 2012. [Google Scholar]
- Casadesus, J.; Kaya, Y.; Bort, J.; Nachit, M.M.; Araus, J.L.; Amor, S.; Ferrazzano, G.; Maalouf, F.; Maccaferri, M.; Martos, V.; et al. Using vegetation indices derived from conventional digital cameras as selection criteria for wheat breeding in water-limited environments. Ann. Appl. Biol. 2007, 150, 227–236. [Google Scholar] [CrossRef]
- Pietragalla, J.; Mullan, D.; Mendoza, R.S. Spectral reflectance. In Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Mexico City, Mexico, 2012. [Google Scholar]
- Loriaux, S.D.; Avenson, T.J.; Welles, J.M.; McDermitt, D.K.; Eckles, R.D.; Riensche, B.; Genty, B. Closing in on maximum yield of chlorophyll fluorescence using a single multiphase flash of sub-saturating intensity. Plant Cell Environ. 2013, 36, 1755–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driever, S.M.; Lawson, T.; Andralojc, P.J.; Raines, C.A.; Parry, M.A.J. Natural variation in photosynthetic capacity, growth, and yield in 64 field-grown wheat genotypes. J. Exp. Bot. 2014, 65, 4959–4973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullis, B.R.; Smith, A.B.; Coombes, N.E. On the design of early generation variety trials with correlated data. J. Agric. Biol. Environ. Stat. 2006, 11, 381–393. [Google Scholar] [CrossRef]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, A.; Sinmena, B.; Mayer, J.; Golan, G.; Shpiler, L. Stem reserve mobilisation supports wheat grain filling under heat stress. Aust. J. Plant Physiol. 1994, 21, 771–781. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Sayre, K.D.; Rajaram, S.; Fischer, R.A. Yield potential progress in short bread wheats in northwest Mexico. Crop Sci. 1997, 37, 36–42. [Google Scholar] [CrossRef]
- Gutierrez-Rodriguez, M.; Reynolds, M.P.; Larque-Saavedra, A. Photosynthesis of wheat in a warm, irrigated environment II. Traits associated with genetic gains in yield. Field Crops Res. 2000, 66, 51–62. [Google Scholar] [CrossRef]
- Condon, A.G.; Reynolds, M.P.; Brennan, J.; van Ginkel, M.; Trethowan, R.; Rebetzke, G.J.; Bonnett, D.G.; Richards, R.A.; Farquhar, G.D. Stomatal aperture related traits and yield potential in bread wheat. In International Symposium on Wheat Yield Potential: Challenges to International Wheat Breeding; CIMMYT: Mexico City, Mexico, 2008. [Google Scholar]
- Gaju, O.; DeSilva, J.; Carvalho, P.; Hawkesford, M.J.; Griffiths, S.; Greenland, A.; Foulkes, M.J. Leaf photosynthesis and associations with grain yield, biomass and nitrogen-use efficiency in landraces, synthetic-derived lines and cultivars in wheat. Field Crops Res. 2016, 193, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Carmo-Silva, E.; Andralojc, P.J.; Scales, J.C.; Driever, S.M.; Mead, A.; Lawson, T.; Raines, C.A.; Parry, M.A.J. Phenotyping of field-grown wheat in the UK highlights contribution of light response of photosynthesis and flag leaf longevity to grain yield. J. Exp. Bot. 2017, 68, 3473–3486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, T.; Kramer, D.M.; Raines, C.A. Improving yield by exploiting mechanisms underlying natural variation of photosynthesis. Curr. Opin. Biotechnol. 2012, 23, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, D.; Górny, A.G. Water- and nitrogen-dependent alterations in the inheritance mode of transpiration efficiency in winter wheat at the leaf and whole-plant level. J. Appl. Genet. 2012, 53, 377–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, M.A.J.; Hawkesford, M.J. An integrated approach to crop genetic improvement. J. Integr. Plant Biol. 2012, 54, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Peñuelas, J.; Garbulsky, M.F.; Filella, I. Photochemical reflectance index (PRI) and remote sensing of plant CO2 uptake. New Phytol. 2011, 191, 596–599. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Huang, W.; Yang, Q.; Xie, Q. Improved estimation of light use efficiency by removal of canopy structural effect from the photochemical reflectance index (PRI). Agric. Ecosyst. Environ. 2015, 199, 333–338. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Leaf and Canopy | Biomass | |||||||
| Early Vigour | Acc. Green Area | Highest Cover | Stay Green | Leaves Weight (Post-A) | Stem Weight (Post-A) | Total Biomass (Post-A) | ||
| Correlation (r) | 0.27 ** | 0.43 *** | 0.42 *** | 0.26 ** | 0.25 ** | 0.23 * | 0.24 * | |
| Variation (σ/) | 0.09 | 0.03 | 0.01 | 0.09 | 0.10 | 0.09 | 0.08 | |
| Heritability (H2) | 0.50 | 0.77 | 0.77 | 0.78 | 0.42 | 0.42 | 0.24 | |
| Reflectance Indices | ||||||||
| NDVI (Veg) | PRI (Veg) | WI (Veg) | NDVI (Pre-A) | PRI (Pre-A) | WI (Pre-A) | PRI (Post-A) | WI (Post-A) | |
| Correlation (r) | 0.49 *** | 0.48 *** | −0.36 *** | 0.28 ** | 0.44 *** | −0.21 * | 0.35 *** | −0.42 *** |
| Variation (σ/) | 0.02 | 0.70 | 0.01 | 0.01 | 1.98 | 0.02 | 0.33 | 0.01 |
| Heritability (H2) | 0.71 | 0.89 | 0.57 | 0.79 | 0.87 | 0.70 | 0.93 | 0.79 |
| Gas-Exchange | ||||||||
| AQ1800 (Pre-A) | AQ1000 (Pre-A) | AQ500 (Pre-A) | AQ250 (Pre-A) | AQ100 (Pre-A) | Amax (Pre-A) | AQ1800 (Post-A) | AQ1000 (Post-A) | |
| Correlation (r) | 0.28 ** | 0.29 ** | 0.28 ** | 0.28 ** | 0.23 * | 0.21 * | 0.18 * | 0.19 * |
| Variation (σ/) | 0.07 | 0.06 | 0.05 | 0.04 | 0.05 | 0.05 | 0.07 | 0.06 |
| Heritability (H2) | 0.59 | 0.61 | 0.61 | 0.47 | 0.28 | 0.54 | 0.59 | 0.61 |
| Light Levels (µmol Photons m−2 s−1) | |||||
|---|---|---|---|---|---|
| 1800 | 1000 | 500 | 250 | 100 | |
| Pre-A | |||||
| A-gs | 0.92 ** | 0.88 ** | 0.82 ** | 0.76 ** | 0.55 ** |
| A-Yield | 0.28 ** | 0.29 ** | 0.28 ** | 0.28 ** | 0.23 * |
| gs-Yield | 0.22 * | 0.21 * | 0.21 * | 0.21 * | 0.21 * |
| Post-A | |||||
| A-gs | 0.80 ** | 0.85 ** | 0.77 ** | 0.73 ** | 0.49 ** |
| A-Yield | 0.18 * | 0.19 * | 0.17 | 0.12 | 0.08 |
| gs-Yield | 0.16 | 0.17 | 0.18 | 0.16 | 0.16 |

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pennacchi, J.P.; Carmo-Silva, E.; Andralojc, P.J.; Feuerhelm, D.; Powers, S.J.; Parry, M.A.J. Dissecting Wheat Grain Yield Drivers in a Mapping Population in the UK. Agronomy 2018, 8, 94. https://doi.org/10.3390/agronomy8060094
Pennacchi JP, Carmo-Silva E, Andralojc PJ, Feuerhelm D, Powers SJ, Parry MAJ. Dissecting Wheat Grain Yield Drivers in a Mapping Population in the UK. Agronomy. 2018; 8(6):94. https://doi.org/10.3390/agronomy8060094
Chicago/Turabian StylePennacchi, João P., Elizabete Carmo-Silva, P. John Andralojc, David Feuerhelm, Stephen J. Powers, and Martin A. J. Parry. 2018. "Dissecting Wheat Grain Yield Drivers in a Mapping Population in the UK" Agronomy 8, no. 6: 94. https://doi.org/10.3390/agronomy8060094
APA StylePennacchi, J. P., Carmo-Silva, E., Andralojc, P. J., Feuerhelm, D., Powers, S. J., & Parry, M. A. J. (2018). Dissecting Wheat Grain Yield Drivers in a Mapping Population in the UK. Agronomy, 8(6), 94. https://doi.org/10.3390/agronomy8060094