Genetic Diversity of Clinal Freezing Tolerance Variation in Winter Wheat Landraces


 and
and
Abstract
:1. Introduction
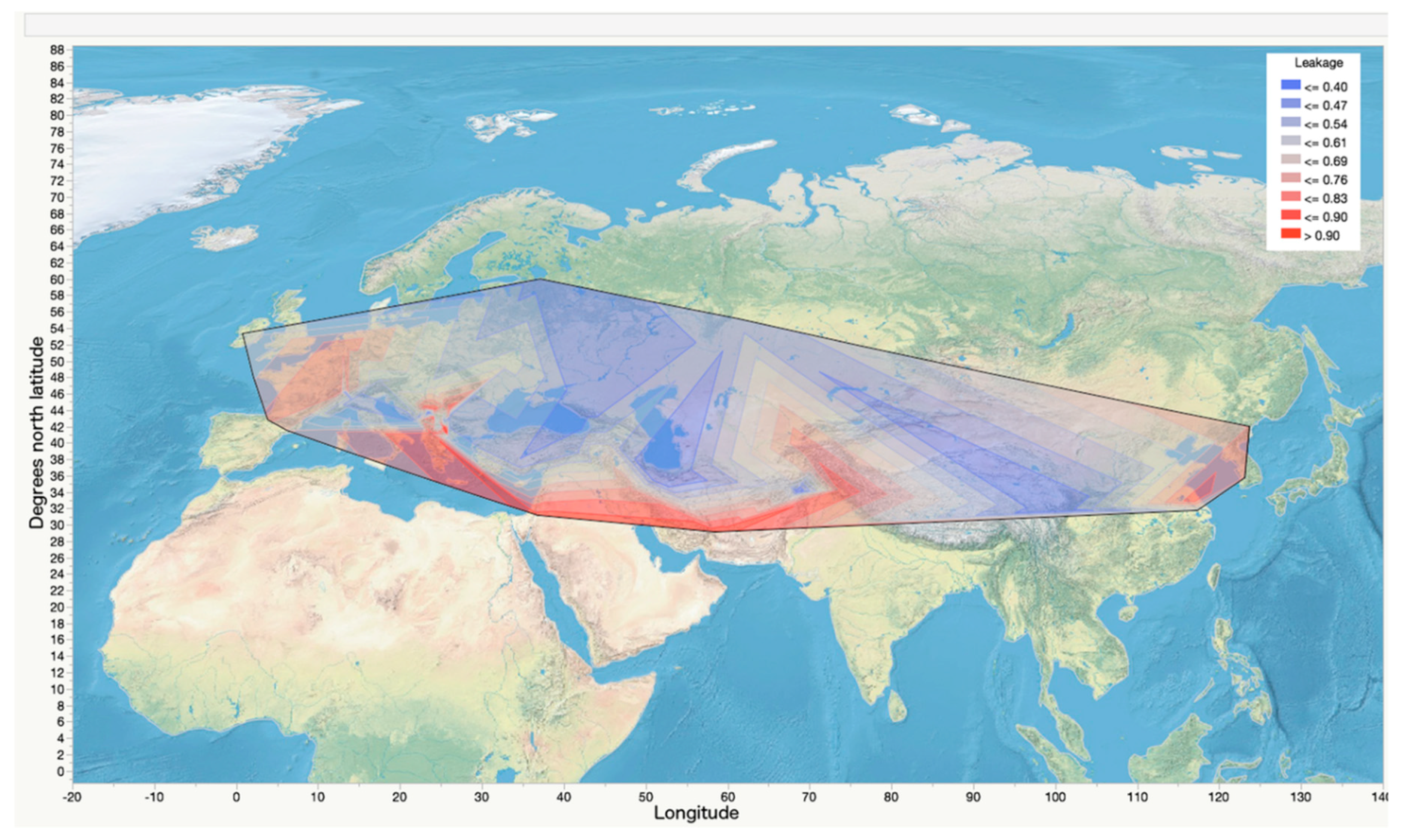
2. Materials and Methods
3. Results
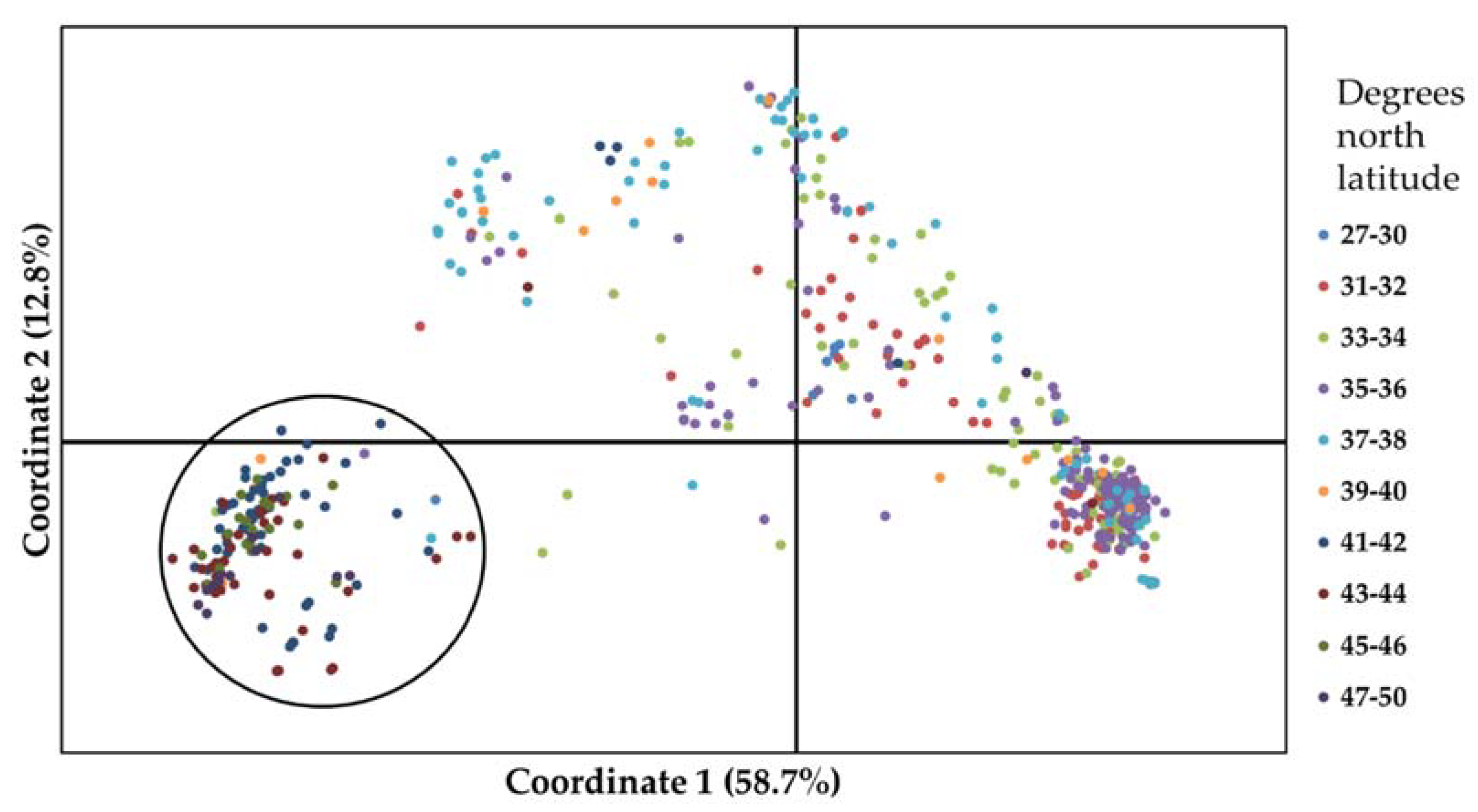
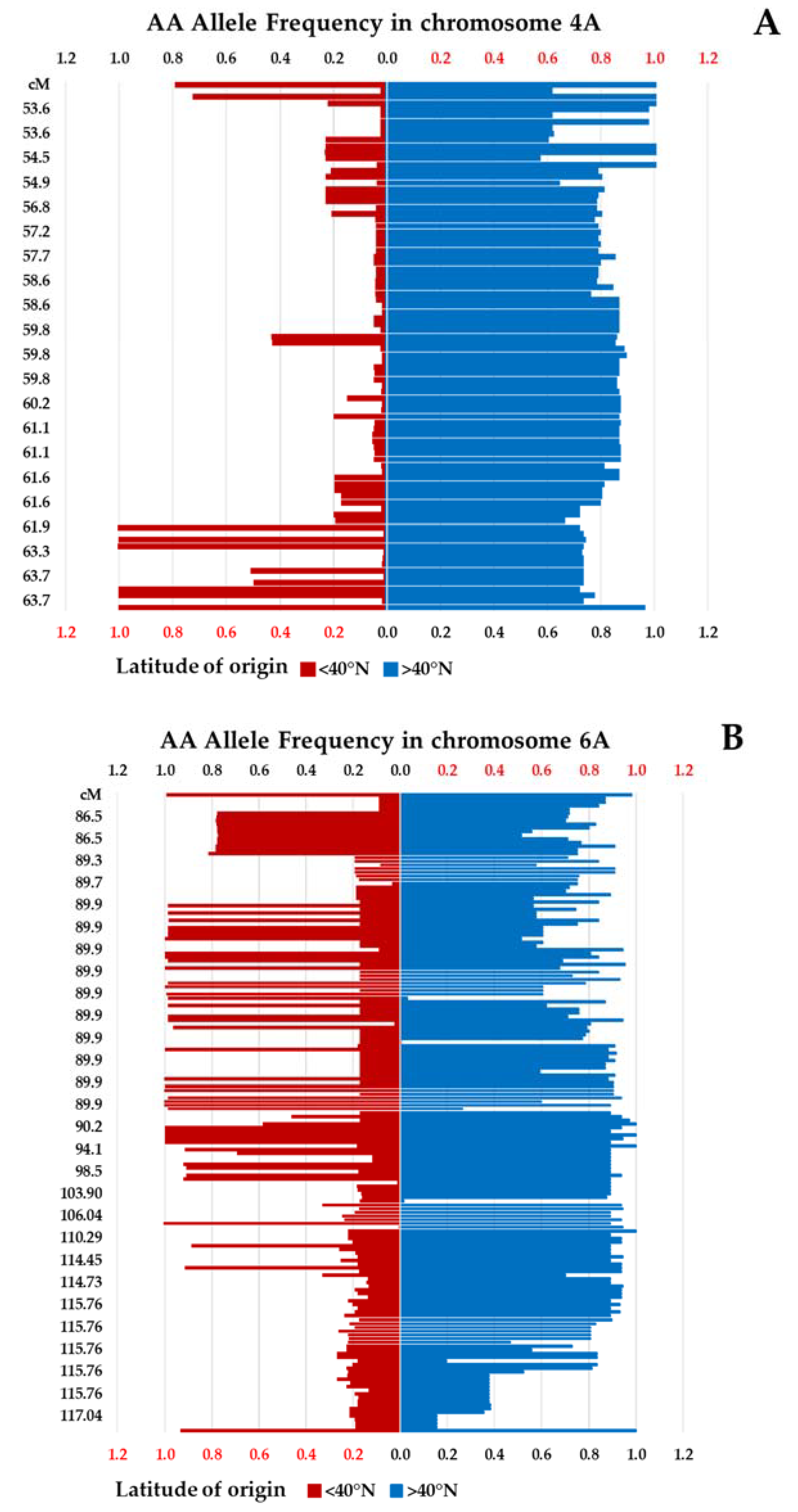
3.1. Genetic Diversity of Winter Wheat Landraces Based on SNPs
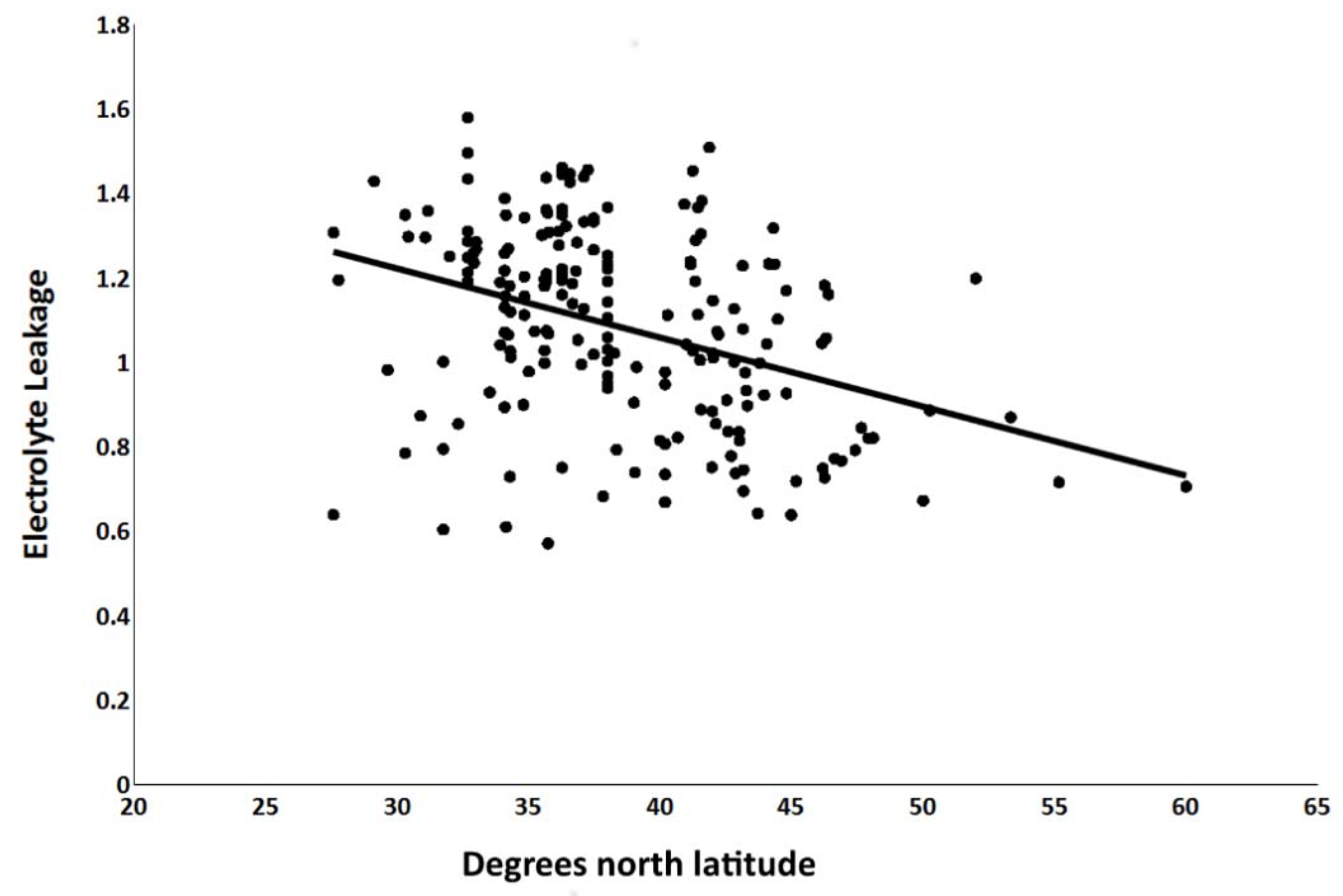
3.2. Electrolyte Leakage Test
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Briggle, L.W.; Curtis, B.C. Wheat worldwide. In Wheat and Wheat Improvement; Agronomy Monograph 13; Heyne, E.G., Ed.; American Society of Agronomy; Crop Science Society of America; Soil Science Society of America: Madison, WI, USA, 1987; pp. 1–31. [Google Scholar]
- Nuttonson, M.Y. Wheat-Climate Relationships and the Use of Phenology in Ascertaining the Thermal and Photo-Thermal Requirements of Wheat: Based on Data of North America and Some Thermally Analogous Areas of North America in the Soviet Union and Finland; American Institute of Crop Ecology: Washington, DC, USA, 1955; p. 34. [Google Scholar]
- Salamini, F.; Ozkan, H.; Brandolini, A.; Schafer-Pregl, R.; Martin, W. Genetics and geography of wild cereal domestication in the near east. Nat. Rev. Genet. 2002, 3, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Gutafson, P.; Raskina, O.; Ma, X.; Nevo, E. Wheat Evolution, Domestication, and Improvement. In Wheat Science and Trade; Carver, B.F., Ed.; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 3–30. [Google Scholar]
- Gopher, A.; Abbo, S.; Lev-Yadun, S. The “when”, the “where” and the “why” of the Neolithic revolution in the Levant. Doc. Prehist. 2002, 28, 49–62. [Google Scholar] [CrossRef]
- Lev-Yadun, S.; Gopher, A.; Abbo, S. The cradle of agriculture. Science 2000, 288, 1602–1603. [Google Scholar] [CrossRef] [PubMed]
- Harlan, J.R.; Zohary, D. Distribution of wild wheats and barley. Science 1966, 153, 1074–1080. [Google Scholar] [CrossRef] [PubMed]
- Heun, M.; Schäfer-Pregl, R.; Klawan, D.; Castagna, R.; Accerbi, M.; Borghi, B.; Salamini, F. Site of einkorn wheat domestication identified by DNA fingerprinting. Science 1997, 278, 1312–1314. [Google Scholar] [CrossRef]
- Luo, M.-C.; Yang, Z.-L.; You, F.M.; Kawahara, T.; Waines, J.G.; Dvorak, J. The structure of wild and domesticated emmer wheat populations, gene flow between them, and the site of emmer domestication. Theor. Appl. Genet. 2007, 114, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Diamond, J. Location, location, location: The first farmers. Science 1997, 278, 1243–1244. [Google Scholar] [CrossRef]
- Chapman, V.; Miller, T.E.; Riley, R. Equivalence of the A genome of bread wheat and that of Triticum urartu. Genet. Res. 1976, 27, 69–76. [Google Scholar] [CrossRef]
- Dvorak, J.; di Terlizzi, P.; Zhang, H.-B.; Resta, P. The evolution of polyploid wheats: Identification of the A genome donor species. Genome 1993, 36, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.; Unrau, J.; Chapman, V. Evidence on the origin of the B. genome of wheat. J. Hered. 1958, 49, 91–98. [Google Scholar] [CrossRef]
- Nevo, E.; Korol, A.B.; Beiles, A.; Fahima, T. Evolution of Wild Emmer and Wheat Improvement: Population Genetics, Genetic Resources, and Genome Organization of Wheat’s Progenitor, Triticum dicoccoides; Springer: Heidelberg, Germany, 2002. [Google Scholar]
- Feldman, M.; Kislev, M.E. Domestication of emmer wheat and evolution of free-threshing tetraploid wheat. Isr. J. Plant Sci. 2007, 55, 207–221. [Google Scholar] [CrossRef]
- Gill, B.S.; Friebe, B. Cytogenetics, phylogeny and evolution of cultivated wheats. In Bread Wheat—Improvement and Production; FAO Plant Production and Protection Series; Curtis, B.C., Rajaram, S., Gomez Macpherson, H., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; Volume 30. [Google Scholar]
- McFadden, E.S.; Sears, E.R. The origin of Triticum spelta and its free-threshing hexaploid relatives. J. Hered. 1946, 37, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Cox, T.S. Deepening the wheat gene pool. J. Crop Prod. 1997, 1, 1–25. [Google Scholar] [CrossRef]
- Dvorak, J.; Luo, M.-C.; Yang, Z.-L.; Zhang, H.-B. The structure of the Aegilops tauschii genepool and the evolution of hexaploid wheat. Theor. Appl. Genet. 1998, 97, 657–670. [Google Scholar] [CrossRef]
- Talbert, L.E.; Smith, L.Y.; Blake, N.K. More than one origin of hexaploid wheat is indicated by sequence comparison of low-copy DNA. Genome 1998, 41, 402–407. [Google Scholar] [CrossRef]
- Snape, J.W.; Law, C.N.; Parker, B.B.; Worland, A.J. Genetical analysis of chromosome 5A of wheat and its influence on important agronomic characters. Theor. Appl. Genet. 1985, 71, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.J.; Fellers, J.P.; Trick, H.N.; Zhang, Z.; Tai, Y.-S.; Gill, B.S.; Faris, J.D. Molecular characterization of the major wheat domestication gene Q. Genetics 2005, 172, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Jones, H.; Charles, M.P.; Jones, M.K.; Colledge, S.; Leigh, F.J.; Lister, D.A.; Smith, L.M.J.; Powell, W.; Brown, T.A. Phylogeographic analysis of barley DNA as evidence for the spread of Neolithic agriculture through Europe. J. Archaeol. Sci. 2012, 39, 3230–3238. [Google Scholar] [CrossRef]
- Colledge, S.; Conolly, J.; Shennan, S. The evolution of Neolithic farming from SW Asian origins to NW European limits. Eur. J. Archaeol. 2005, 8, 137–156. [Google Scholar] [CrossRef]
- Bogucki, P. How agriculture came to North-Central Europe. In Europe’s First Farmers; Price, T.D., Ed.; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Ladizinsky, G. Founder effect in crop-plant evolution. Econ. Bot. 1985, 39, 191–199. [Google Scholar] [CrossRef]
- Santamaria, L.; Figuerola, J.; Pilon, J.J.; Mjelde, M.; Green, A.J.; De Boer, T.; King, R.A.; Gornall, R.J. Plant performance across latitude: The role of plasticity and local adaptation in an aquatic plant. Ecology 2003, 84, 2454–2461. [Google Scholar] [CrossRef] [Green Version]
- Sakai, A. Freezing resistance in willows from different climates. Ecology 1970, 51, 485–491. [Google Scholar] [CrossRef]
- Ying, C.C.; Bagley, W.T. Genetic variation of eastern cottonwood in an eastern Nebraska provenance study. Silvae Genet. 1976, 25, 67–73. [Google Scholar]
- Fu, Y.-B.; Somers, D.J. Genome-wide reduction of genetic diversity in wheat breeding. Crop Sci. 2009, 49, 161–168. [Google Scholar] [CrossRef]
- Zamir, D. Improving plant breeding with exotic genetic libraries. Nat. Rev. Genet. 2001, 2, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 2009 457, 843–848. [Google Scholar] [CrossRef]
- Patterson, F.L.; Shaner, G.E.; Ohm, H.W.; Foster, J.E. A historical perspective for the establishment of research goals for wheat improvement. J. Prod. Agric. 1990, 3, 30–38. [Google Scholar] [CrossRef]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Fowler, D.B. Wheat production in the high winter stress climate of the Great Plains of North America—An experiment in crop adaptation. Crop Sci. 2012, 52, 11–20. [Google Scholar] [CrossRef]
- Law, C.N.; Jenkins, G. A genetic study of cold resistance in wheat. Genet. Res. 1970, 15, 197–308. [Google Scholar] [CrossRef]
- Limin, A.E.; Danyluk, J.; Chauvin, L.-P.; Fowler, D.B.; Sarhan, F. Chromosome mapping of low-temperature induced WCS120 family genes and regulation of cold-tolerance expression in wheat. Mol. Gen. Genet. 1997, 253, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Baga, M.; Chodaparambil, S.V.; Limin, A.E.; Pecar, M.; Fowler, D.B.; Chibbar, R.N. Identification of quantitative trait loci and associated candidate genes for low-temperature tolerance in cold-hardy winter wheat. Funct. Integr. Genom. 2007, 7, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Case, A.J.; Skinner, D.Z.; Garland-Campbell, K.A.; Carter, A.H. Freezing tolerance-associated quantitative trait loci in the Brundage × Coda wheat recombinant inbred line population. Crop Sci. 2014, 54, 982–992. [Google Scholar] [CrossRef]
- Kruse, E.B.; Carle, S.W.; Wen, N.; Skinner, D.Z.; Murray, T.D.; Garland-Campbell, K.A. Genomic regions associated with tolerance to freezing stress and snow mold in winter wheat. Genes Genomes Genet. 2017, 7, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Vagujfalvi, A.; Galiba, G.; Cattivelli, L.; Dubcovsky, J. The cold-regulated transcriptional activator CBF3 is linked to the frost-tolerance locus Fr-A2 on wheat chromosome 5A. Mol. Genet. Genom. 2003, 269, 60–67. [Google Scholar]
- Motomura, Y.; Kobayashi, F.; Iehisa, J.; Takumi, S. A major quantitative trait locus for cold-responsive gene expression is linked to frost-resistance gene Fr-A2 in common wheat. Breed. Sci. 2013, 63, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Pearce, S.; Zhu, J.; Boldizsar, A.; Vagujfalvi, A.; Burke, A.; Garland-Campbell, K.; Galiba, G.; Dubcovsky, J. Large deletions in the CBF gene cluster at the Fr-B2 locus are associated with reduced frost tolerance in wheat. Theor. Appl. Genet. 2013, 126, 2683–2697. [Google Scholar] [CrossRef] [PubMed]
- Todorovska, E.; Kolev, S.; Christov, N.; Balint, A.; Kocsy, G.; Vagujfalvi, A.; Galiba, G. The expression of CBF genes at Fr-2 locus is associated with the level of frost tolerance in Bulgarian winter wheat cultivars. Biotechnol. Biotechnol. Equip. 2014, 28, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Pearce, S.; Burke, A.; See, D.; Skinner, D.Z.; Dubcovsky, J.; Garland-Campbell, K. Copy number and haplotype variation at the VRN-A1 and central FR-A2 loci are associated with frost tolerance in hexaploid wheat. Theor. Appl. Genet. 2014, 127, 1183–1197. [Google Scholar] [CrossRef] [PubMed]
- Würschum, T.; Longin, C.F.H.; Hahn, V.; Tucker, M.R.; Leiser, W.L. Copy number variations of CBF genes at the Fr-A2 locus are essential components of winter hardiness in wheat. Plant J. 2017, 89, 764–773. [Google Scholar] [CrossRef] [PubMed]
- Brule-Babel, A.L.; Fowler, D.B. Genetic control of cold hardiness and vernalization requirement in winter wheat. Crop Sci. 1988, 28, 879–884. [Google Scholar] [CrossRef]
- Mahfoozi, S.; Limin, A.E.; Fowler, D.B. Influence of vernalization and photoperiod responses on cold hardiness in winter cereals. Crop Sci. 2001, 41, 1006–1011. [Google Scholar] [CrossRef]
- Snape, J.W.; Butterworth, K.; Whitechurch, E.; Worland, A.J. Waiting for fine times: Genetics of flowering time in wheat. In Wheat in a Global Environment. Developments in Plant Breeding; Bedo, Z., Lang, L., Eds.; Springer: Dordrecht, The Netherlands, 2001; Volume 9, pp. 67–74. [Google Scholar]
- McIntosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Appels, R.; Xia, X.C. Catalogue of Gene Symbols for wheat: 2012 Supplement. KOMUGI Wheat Genetic Resources Database. 2013. Available online: https://Shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2012.pdf (accessed on 27 March 2018).
- Peakall, R.; Smouse, P.E. Genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Resour. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [PubMed]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Res. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- McKay, H.M. Electrolyte leakage from fine roots of conifer seedlings: A rapid index of plant vitality following cold storage. Can. J. For. Res. 1992, 22, 1371–1377. [Google Scholar] [CrossRef]
- Dexter, S.T.; Tottingham, W.E.; Graber, L.F. Investigations of the hardiness of plants by measurement of electrical conductivity. Plant Physiol. 1932, 7, 63–78. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.B.; Cape, J.N.; Fowler, D. Quantification of frost damage in plant tissues by rates of electrolyte leakage. New Phytol. 1989, 113, 307–311. [Google Scholar] [CrossRef] [Green Version]
- Bajji, M.; Kinet, J.-M.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Cuevas, C.; Bellinger, B.S.; Skinner, D.Z. Membrane stability of winter wheat plants exposed to subzero temperatures for variable lengths of time. Commun. Plant Sci. 2015, 5, 9–14. [Google Scholar]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 2016, 12, E1005767. [Google Scholar] [CrossRef] [PubMed]
- Endler, J.A. Geographic Variation, Speciation, and Clines; Princeton University Press: Princeton, NJ, USA, 1977; p. 246. [Google Scholar]
- Skinner, D.Z. Post-acclimation transcriptome adjustment is a major factor in freezing tolerance of winter wheat. Funct. Integr. Genom. 2009, 9, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Skinner, D.Z. Genes upregulated in winter wheat (Triticum aestivum L.) during mild freezing and subsequent thawing suggest sequential activation of multiple response mechanisms. PLoS ONE 2015, 10, E0133166. [Google Scholar] [CrossRef] [PubMed]
- Churin, Y.; Adam, E.; Kozma-Bognar, L.; Nagy, F.; Borner, T. Characterization of two Myb-like transcription factors binding to CAB promoters in wheat and barley. Plant Mol. Biol. 2003, 52, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cai, J.; Jiang, D.; Liu, F.; Dai, T.; Cao, W. Pre-anthesis high-temperature acclimation alleviates damage to the flag leaf caused by post-anthesis heat stress in wheat. J. Plant Physiol. 2011, 168, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Wollenweber, B.; Jiang, D. Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings. Plant Physiol. Biochem. 2014, 74, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Ning, Z.; Bai, G.; Li, R.; Yan, G.; Siddique, K.H.M.; Baum, M.; Guo, P. Allelic variations of a light harvesting chlorophyll a/b-binding protein gene (Lhcb1) associated with agronomic traits in barley. PLoS ONE 2012, 7, e37573. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP a | CHR b | cM c | MAF d | Obs e | p-Value | Trait | Possible Gene Identification f |
|---|---|---|---|---|---|---|---|
| IWA3431 | 2A | 117.57 | 0.37 | 553 | 3.02 × 10−124 | Population structure | Translation initiation factor |
| IWA6597 | 4A | 57.56 | 0.26 | 553 | 1.11 × 10−107 | Population structure | Unknown function, transmembrane domains |
| IWA7271 | 4A | 46.27 | 0.23 | 553 | 1.02 × 10−50 | Latitude | phosphatidylinositol N-acetyglucosaminyltransferase |
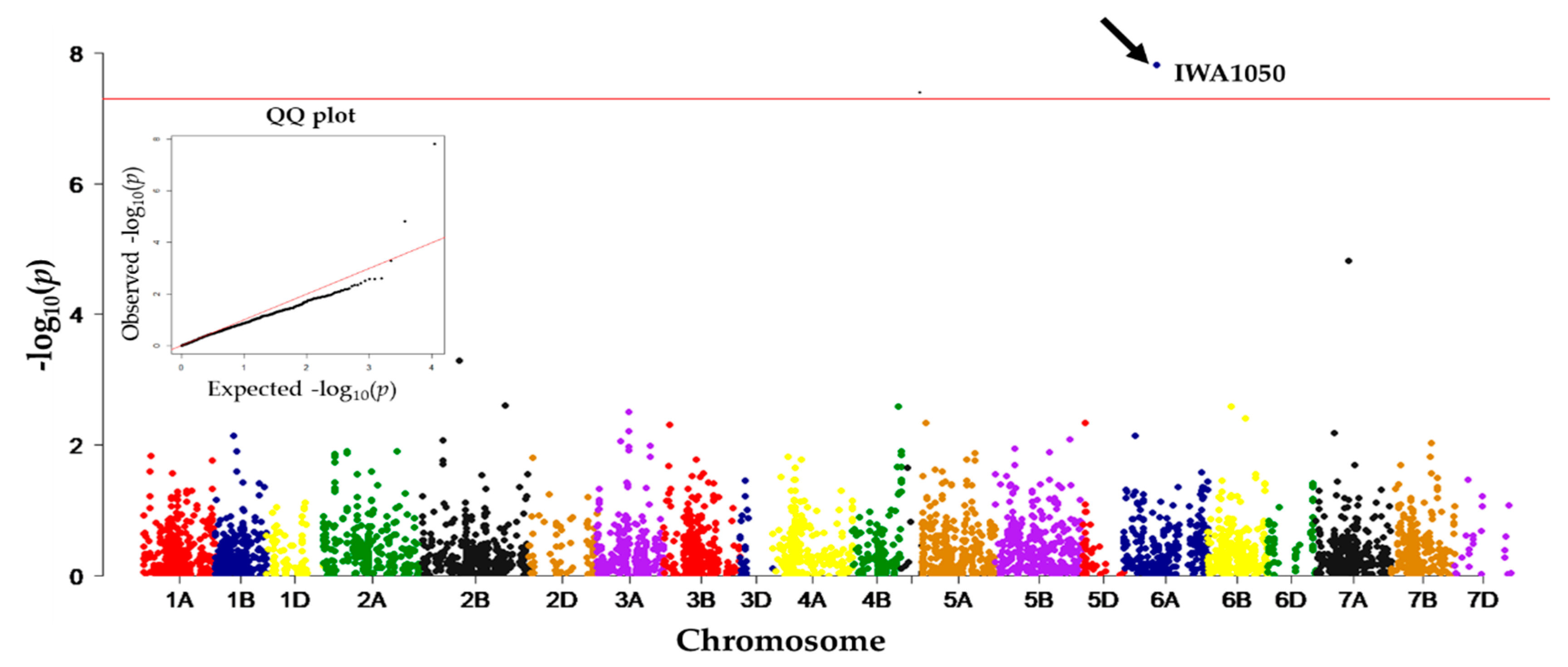
| IWA1050 | 6A | 85.53 | 0.41 | 192 | 1.52 × 10−8 | Electrolyte leakage | chlorophyll a/b binding protein |
| Source of Variation | df a | Sum of Squares b | % Explained Variation |
|---|---|---|---|
| Latitude | 1 | 21.6 *** | 52.5 |
| Test temperature | 2 | 17.0 *** | 41.5 |
| Replications | 1 | 2.3 | 5.6 |
| Subsamples | 2 | 0.2 | 0.4 |
| Model | 6 | 41.1 | |
| Error | 2949 | 258.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sthapit Kandel, J.; Huang, M.; Zhang, Z.; Skinner, D.Z.; See, D.R. Genetic Diversity of Clinal Freezing Tolerance Variation in Winter Wheat Landraces. Agronomy 2018, 8, 95. https://doi.org/10.3390/agronomy8060095
Sthapit Kandel J, Huang M, Zhang Z, Skinner DZ, See DR. Genetic Diversity of Clinal Freezing Tolerance Variation in Winter Wheat Landraces. Agronomy. 2018; 8(6):95. https://doi.org/10.3390/agronomy8060095
Chicago/Turabian StyleSthapit Kandel, Jinita, Meng Huang, Zhiwu Zhang, Daniel Z. Skinner, and Deven R. See. 2018. "Genetic Diversity of Clinal Freezing Tolerance Variation in Winter Wheat Landraces" Agronomy 8, no. 6: 95. https://doi.org/10.3390/agronomy8060095
APA StyleSthapit Kandel, J., Huang, M., Zhang, Z., Skinner, D. Z., & See, D. R. (2018). Genetic Diversity of Clinal Freezing Tolerance Variation in Winter Wheat Landraces. Agronomy, 8(6), 95. https://doi.org/10.3390/agronomy8060095