Development of Allergenicity and Toxicity Assessment Methods for Evaluating Transgenic Sugarcane Overexpressing Sucrose–Phosphate Synthase

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Animals and Housing
2.3. Allergenicity Assessment
2.3.1. Bioinformatics Analysis
2.3.2. Heat Stability Treatment
2.3.3. In Vitro Simulated Gastric Fluids (SGF) and Simulated Intestine Fluid (SIF) Assays
2.3.4. In Vivo Allergenicity Assay in Rats and Measurement of the Protein-Specific IgE Antibody
2.4. Acute Oral Toxicity Assessment
2.5. Statistical Analysis
3. Results and Discussion
3.1. Bioinformatics Assessment of the Protein SPS Expressed in Transgenic Sugarcane
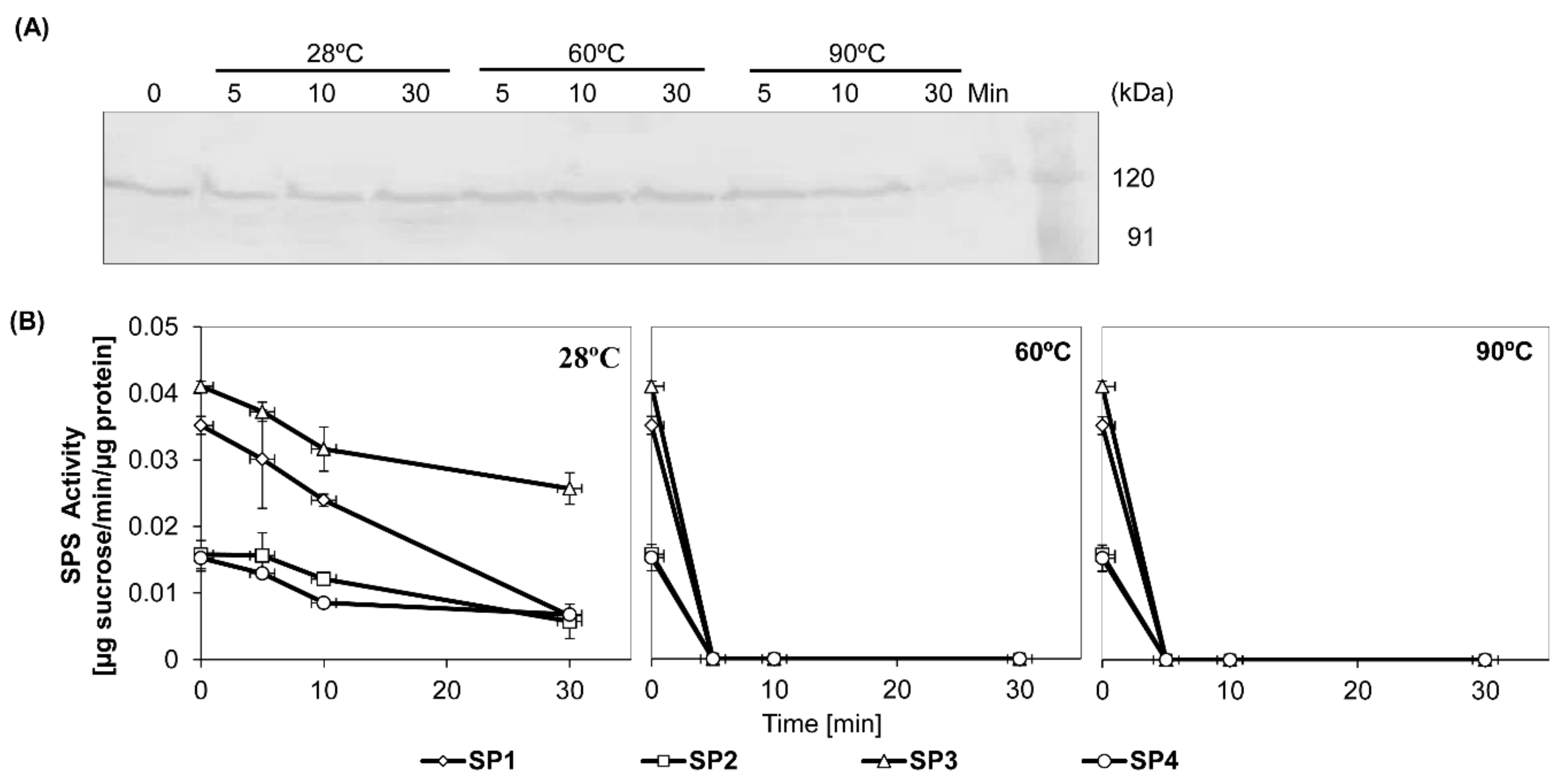
3.2. Thermal Stability Assessment
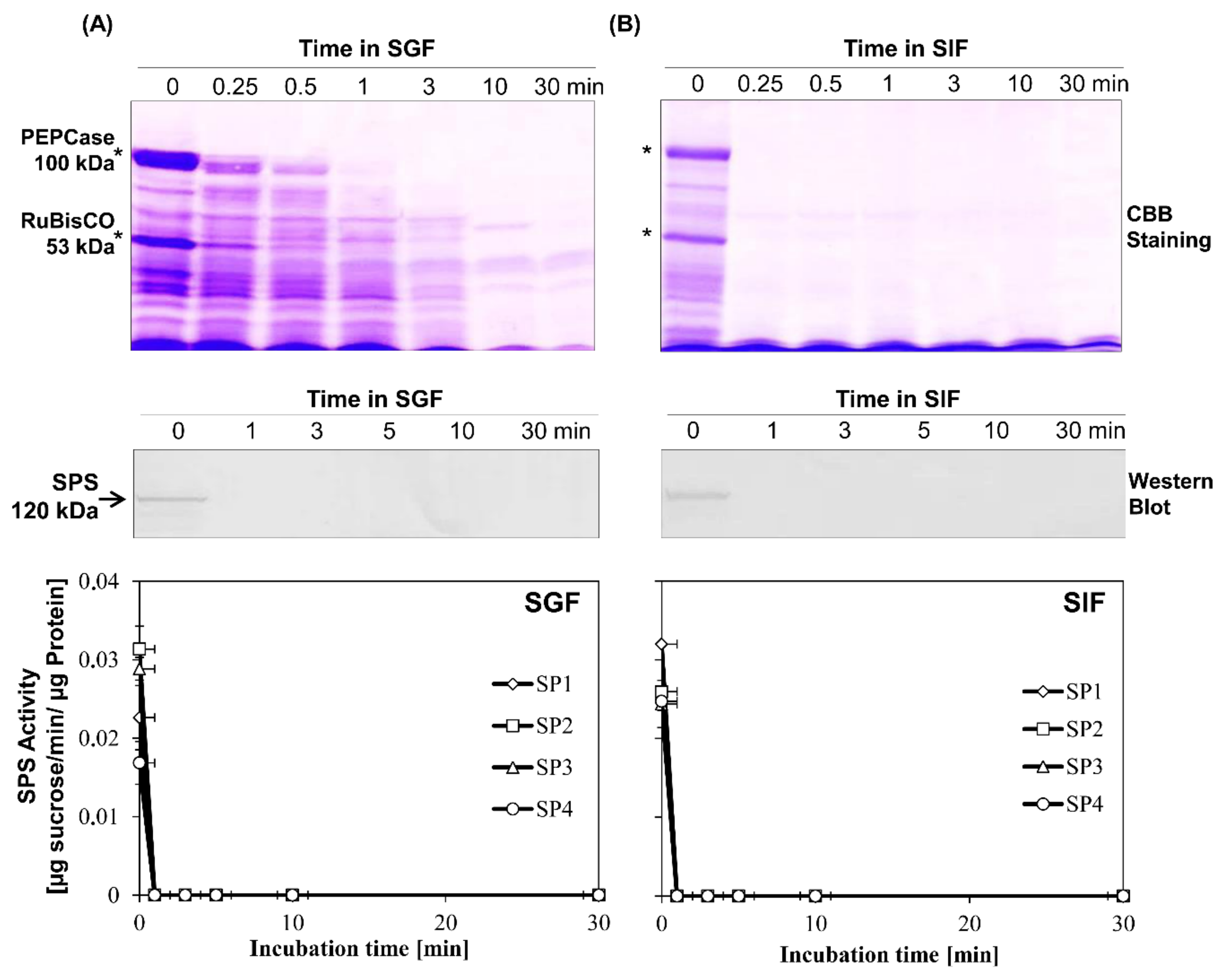
3.3. In Vitro Simulated Gastric Fluids (SGF) and Simulated Intestine Fluid (SIF) Assessment
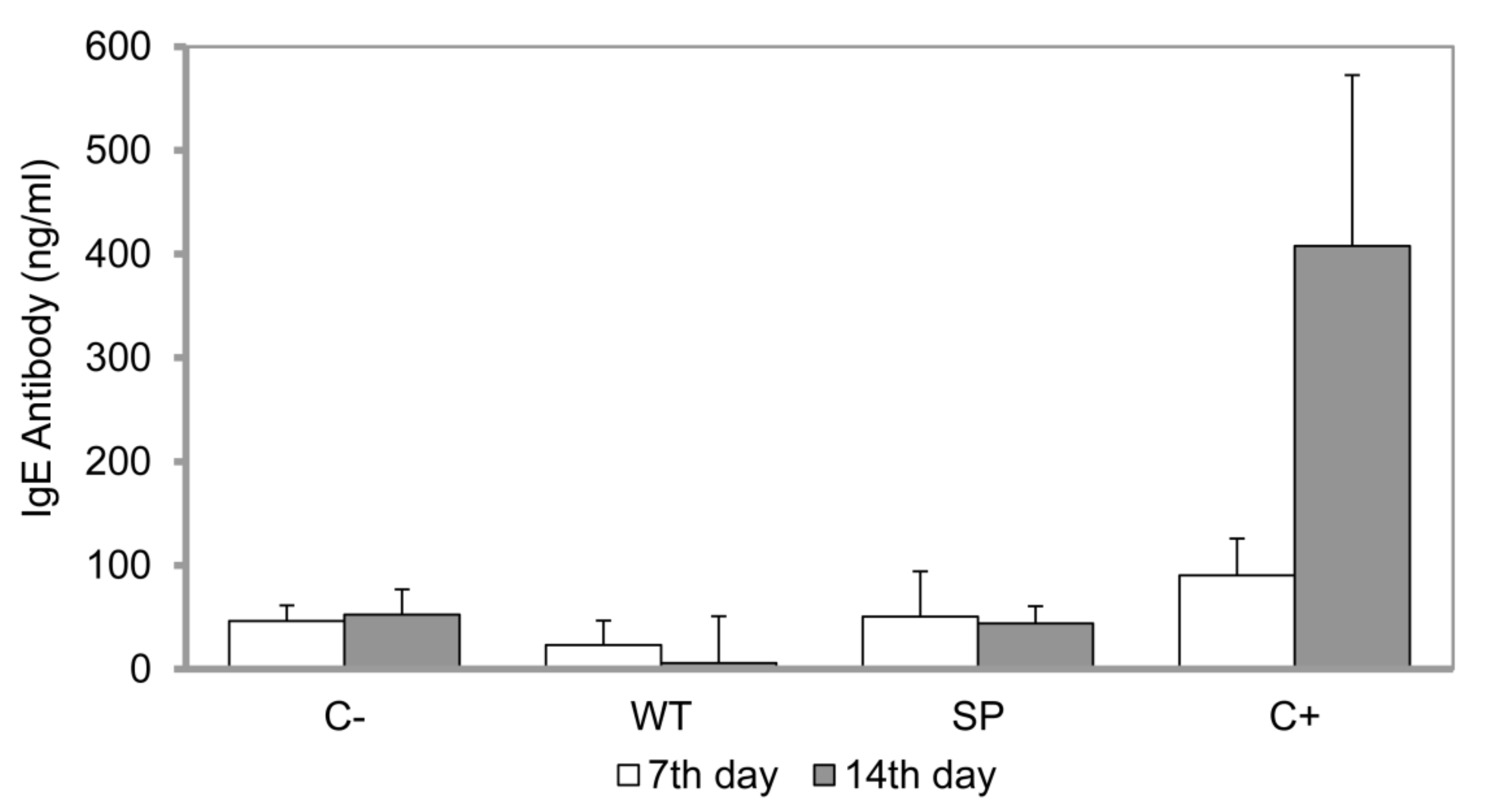
3.4. In Vivo Allergenicity Assay in Rats and Measurement of the Protein-Specific IgE Antibody
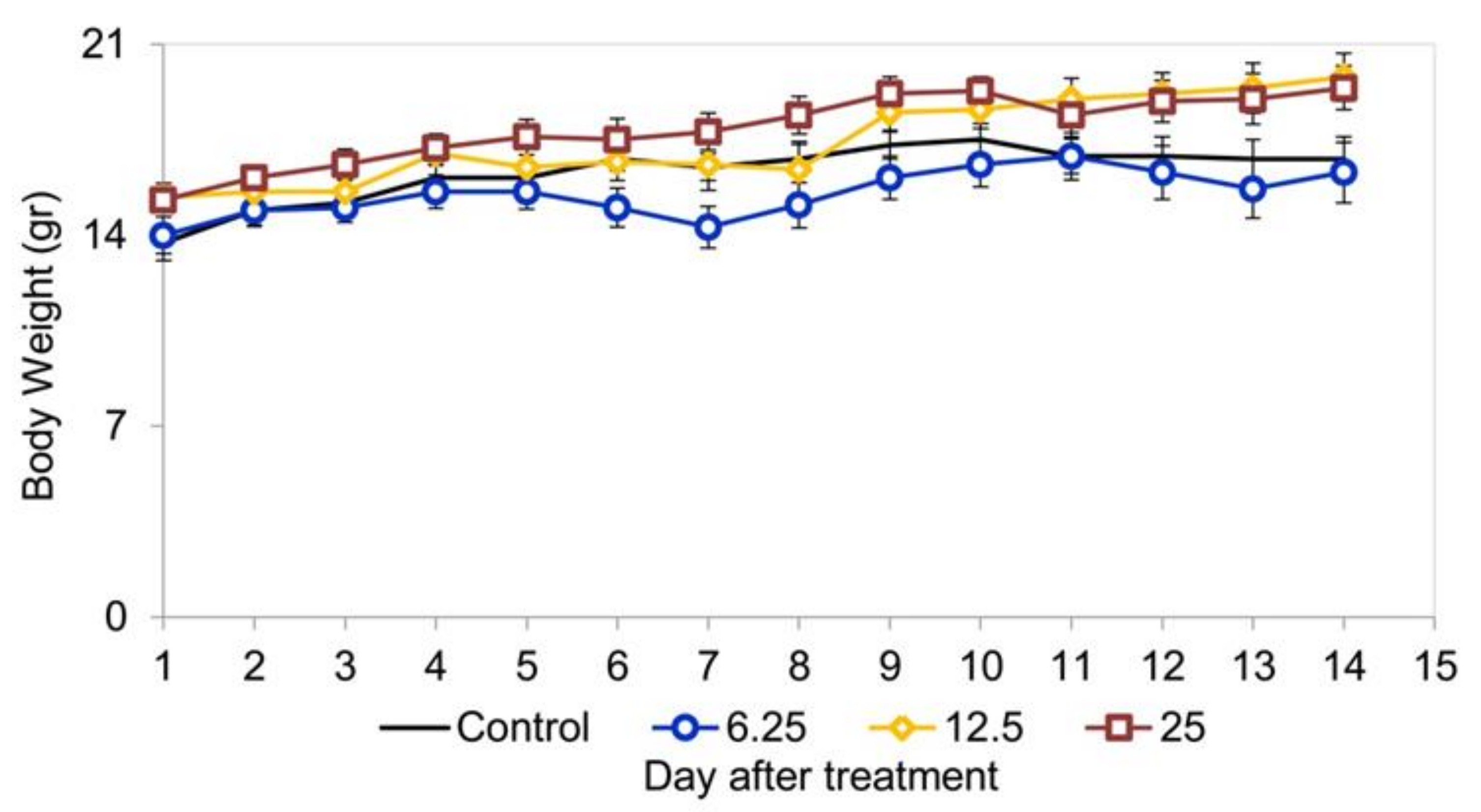
3.5. Acute Oral Toxicity Assessment
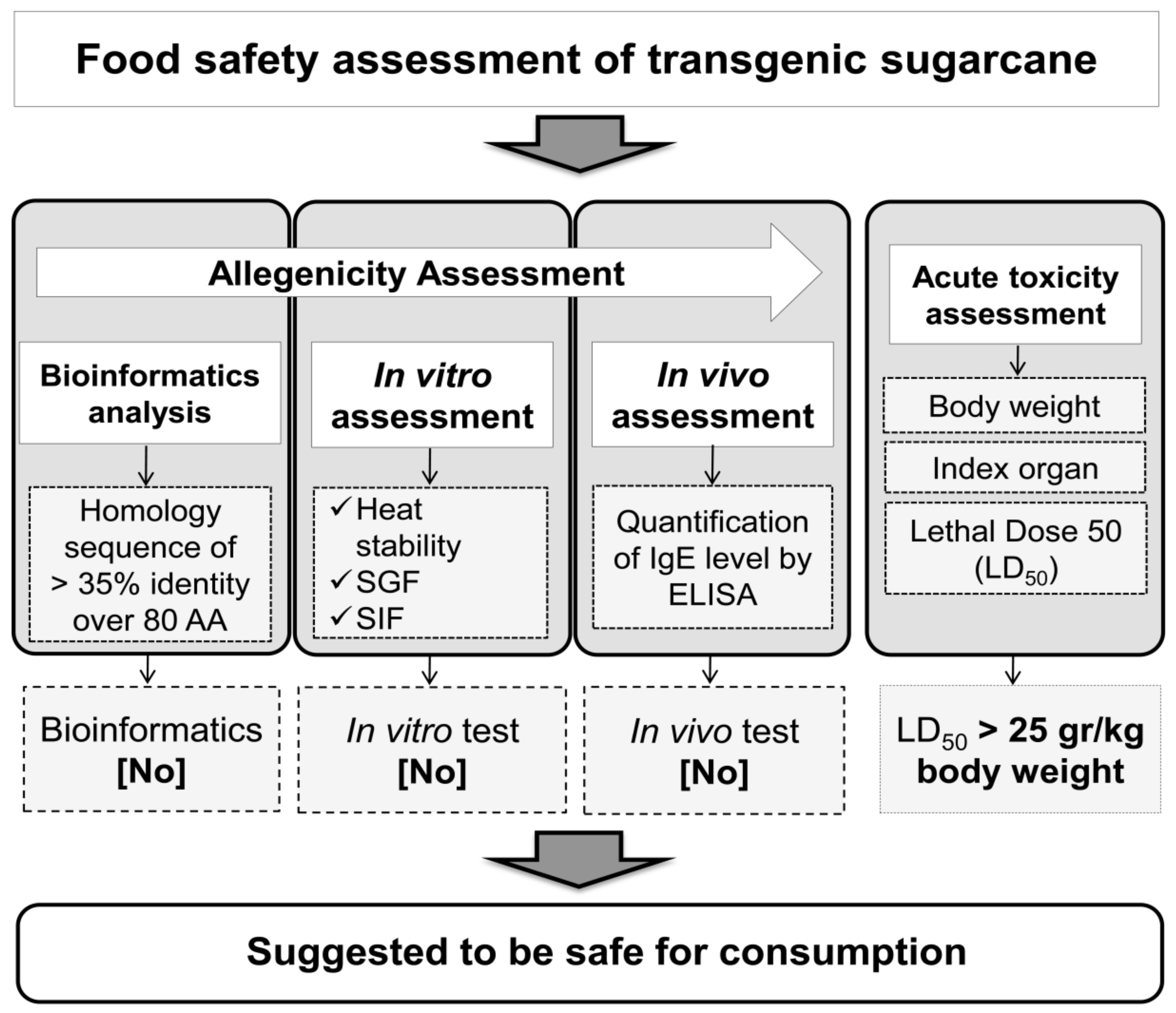
3.6. A Model for Food Safety Assessment of Transgenic Sugarcane
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- FAO/WHO. Evaluation of Allergenicity of Genetically Modified Foods; Report of a Joint FAO/WHO Expert Consultation of Allergenicity of Foods Derived from Biotechnology: Rome, Italy, 2001. [Google Scholar]
- Delaney, B. Safety assessment of foods from genetically modified crops in countries with developing economies. Food Chem. Toxicol. 2015, 86, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Farias, D.F.; Viana, M.P.; Oliveira, G.R.; Santos, V.O.; Pinto, C.E.M.; Viana, D.A.; Vasconcelos, I.M.; de-Sa, M.F.G.; Carvalho, A.F.U. Food safety assessment of Cry8Ka5 mutant protein using Cry1Ac as a control Bt protein. Food Chem. Toxicol. 2015, 81, 81–91. [Google Scholar] [CrossRef]
- Zhao, K.; Ren, F.; Han, F.; Liu, Q.; Wu, G.; Xu, Y.; Zhang, J.; Wu, X.; Wang, J.; Li, P.; et al. Edible safety assessment of genetically modified rice T1C-1 for Sprague Dawley rats through horizontal gene transfer, allergenicity and intestinal microbiota. PLoS ONE 2016, 11, e0163352. [Google Scholar] [CrossRef] [PubMed]
- Codex Alimentarius Commission. Joint FAO/WHO Food Standard Program, Food and Agriculture Organization of the United Nations; Food Derived from Biotechnology: Rome, Italy, 2003. [Google Scholar]
- Parrott, W.; Chassy, B.; Ligon, J.; Meyer, L.; Petrick, J.; Zhou, J.; Herman, R.; Delaney, B.; Levine, M. Application of food and feed safety assessment principles to evaluate transgenic approaches to gene modulation in crops. Food Chem. Toxicol. 2010, 48, 1773–1790. [Google Scholar] [CrossRef] [PubMed]
- Dunn, S.E.; Vicini, J.L.; Glenn, K.C.; Fleischer, D.M.; Greenhawt, M.J. The allergenicity of genetically modified foods from genetically engineered crops: A narrative and systematic review. Ann. Allergy Asthma Immunol. 2017, 119, 214–222. [Google Scholar] [CrossRef]
- Lee, R.; Reiner, D.; Dekan, G.; Moore, A.E.; Epstein, M.M.; Higgins, T.J.V. Genetically modified α-amylase inhibitor peas are not specifically allergenic in mice. PLoS ONE 2013, 8, e52972. [Google Scholar] [CrossRef]
- Sugiharto, B. Biotechnology of drought-tolerant sugarcane. In Sugarcane Technology and Research; Olieveira, A.D., Ed.; InTech Open: London, UK, 2018; ISBN 978-1-78923-151-9. [Google Scholar]
- Waltz, E. Beating the heat; Despite the complexity of drought tolerance, researcher is making progress in the search for crops that can produce seed with limited water. Nat. Biotechnol. 2014, 32, 610–613. [Google Scholar] [CrossRef]
- Gao, S.; Yang, Y.; Wang, C.; Guo, J.; Zhou, D.; Wu, Q.; Su, Y.; Xu, L.; Que, Y. Transgenic sugarcane with a cry1Ac gene exhibited better phenotypic traits and enhanced resistance against sugarcane borer. PLoS ONE 2016, 11, e0153929. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.Z.; Yang, B.P.; Feng, X.Y.; Cao, Z.Y.; Feng, C.L.; Wang, J.G.; Xiong, G.R.; Shen, L.B.; Zeng, J.; Zhao, T.T.; et al. Development and characterization of transgenic sugarcane with insect resistance and herbicide tolerance. Front. Plant Sci. 2017, 8, 1–10. [Google Scholar] [CrossRef]
- Yao, W.; Ruan, M.; Qin, L.; Yang, C.; Chen, R.; Chen, B.; Zhang, M. Field performance of transgenic sugarcane lines resistant to sugarcane mosaic virus. Front. Plant Sci. 2017, 8, 104. [Google Scholar] [CrossRef]
- Apriasti, R.; Widyaningrum, S.; Hidayati, W.N.; Sawitri, W.D.; Darsono, N.; Hase, T.; Sugiharto, B. Full sequence of the coat protein gene is required for the induction of pathogen-derived resistance against sugarcane mosaic virus in transgenic sugarcane. Mol. Biol. Rep. 2018, 45, 2749–2758. [Google Scholar] [CrossRef] [PubMed]
- Astwood, J.D.; Leach, J.N.; Fuchs, R.L. Stability of food allergens to digestion in vitro. Nat. Biotechnol. 1996, 14, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Knippels, L.M.; Penninks, A.H.; Smit, J.J.; Houben, G.F. Immune-mediated effects upon oral challenge of ovalbumin-sensitized Brown Norway rats: Further characterization of a rat food allergy model. Toxicol. Appl. Pharmacol. 1999, 156, 161–169. [Google Scholar] [CrossRef]
- Domingo, J.L. Toxicity studies of genetically modified plants: A review of the published literature. Crit. Rev. Food Sci. Nutr. 2007, 47, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Fares, N.H.; El-Sayed, A.K. Fine structural changes in the ileum of mice fed on delta-endotoxin-treated potatoes and transgenic potatoes. Nat. Toxins 1998, 6, 219–233. [Google Scholar] [CrossRef]
- Delaney, B.; Astwood, J.D.; Cunny, H.; Conn, R.E.; Herouet-Guicheney, C.; Macintosh, S.; Meyer, L.S.; Privalle, L.; Gao, Y.; Mattsson, J.; et al. Evaluation of protein safety in the context of agricultural biotechnology. Food Chem. Toxicol. 2008, 46, S71–S97. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.C. Chapter 2—Rodents model for toxicity testing and biomarkers. In Biomarkers in Toxicology; Gupta, R.C., Ed.; Academic Press: Cambridge, UK, 2014; pp. 8–68. ISBN 978-0-12-404630-6. [Google Scholar]
- OECD. Acute Oral Toxicity—Fixed Dose Procedure Section 4. In OECD Guideline for the Testing of Chemicals No 420; OECD Publishing: Paris, France, 2001; pp. 1–14. [Google Scholar]
- Song, Y.; Liang, C.; Wang, W.; Fang, J.; Sun, N.; Jia, X.; Li, N. Immunotoxicological evaluation of corn genetically modified with Bacillus thuringiensis Cry1Ah gene by a 30-day feeding study in BALB/c mice. PLoS ONE 2014, 9, e78566. [Google Scholar] [CrossRef]
- Sugiharto, B.; Sakakibara, H.; Sumadi; Sugiyama, T. Differential expression of two genes for sucrose-phosphate synthase in sugarcane: Molecular cloning of the cDNAs and comparative analysis of gene expression. Plant Cell Physiol. 1997, 38, 961–965. [Google Scholar] [CrossRef] [PubMed]
- Sawitri, W.D.; Narita, H.; Ishizaka-Ikeda, E.; Sugiharto, B.; Hase, T.; Nakagawa, A. Purification and characterization of recombinant sugarcane sucrose phosphate synthase expressed in E. coli and insect Sf9 cells: An importance of the N-terminal domain for an allosteric regulatory property. J. Biochem. 2016, 159, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.E.; Ebisawa, M.; Ferreira, F.; Sampson, H.A.; Van Ree, R.; Vieths, S.; Baumert, J.L.; Bohle, B.; Lalithambika, S.; Wise, J.; et al. AllergenOnline: A peer reviewed, curated allergen database to assess novel food proteins for potential cross-reactivity. Mol. Nutr. Food Res. 2016, 60, 1183–1198. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Roesler, K.R.; Rao, A.G. Rapid gastric fluid digestion and biochemical characterization of engineered proteins enriched in essential amino acids. J. Agric. Food Chem. 2001, 49, 3443–3451. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bellou, A.; Saint-Laudy, J.; Knippels, L.; Montemont, C.; Vauthier, E.; Gerard, P.; Pellegrom, H.; Koerkamp, E.K.; Lesesve, J.F.; Gueant, J.L.; et al. Brown Norway rat ovalbumin-specific immunoglobulin E antibodies increase the human basophil expression of CD63 marker. Scand. J. Immunol. 2002, 57, 271–278. [Google Scholar] [CrossRef]
- Verma, S.K.; Srikanth, R.; Das, SK.; Venkidachalam, G. An efficient and novel approach for clarification of sugarcane juice by micro- and ultrafiltration methods. Indian J. Chem. 1996, 3, 136–139. [Google Scholar]
- Shi, W.; Zhang, C.; Zhao, D.; Wang, L.; Li, P.; Li, H. Discovery of hepatotoxic equivalent combinatorial markers from Dioscorea bulbifera tuber by fingerprint-toxicity relationship modeling. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ladics, G.S.; Cressman, R.F.; Herouet-Guicheney, C.; Herman, R.A.; Privalle, L.; Song, P.; Ward, J.M.; McClain, S. Bioinformatics and the allergy assessment of agricultural biotechnology products: Industry practices and recommendations. Regul. Toxicol. Pharmacol. 2011, 60, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Young, G.J.; Zhang, S.; Mirsky, H.P.; Cressman, R.F.; Cong, B.; Ladics, G.S.; Zhong, C.X. Assessment of possible allergenicity of hypothetical ORFs in common food crops using current bioinformatic guidelines and its implications for the safety assessment of GM crops. Food Chem. Toxicol. 2012, 50, 3741–3751. [Google Scholar] [CrossRef] [Green Version]
- Goodman, R.E.; Vieths, S.; Sampson, H.A.; Hill, D.; Ebisawa, M.; Taylor, S.L.; van Ree, R. Allergenicity assessment of genetically modified crops--what makes sense? Nat. Biotechnol. 2008, 26, 73–81. [Google Scholar] [CrossRef]
- Herman, R.A.; Song, P.; Sekhar, A.R. Value of eight-amino-acid matches in predicting the allergenicity status of protein: An empirical bioinformatics investigation. Clin. Mol. Allergy 2009, 7, 1–7. [Google Scholar] [CrossRef]
- Goodman, R.E.; Tetteh, A.O. Suggested improvements for the allergenicity assessment of genetically modified plants used in foods. Curr. Allergy Asthma Rep. 2011, 11, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Goodman, R.E.; Tetteh, A.O.; Lu, M.; Tripathi, L. Bioinformatics analysis to assess potential risks of allergenicity and toxicity of HRAP and PFLP proteins in genetically modified bananas resistant to Xanthomonas wilt disease. Food Chem. Toxicol. 2017, 109, 81–89. [Google Scholar] [CrossRef]
- Bushey, D.F.; Bannon, G.A.; Delaney, B.F.; Graser, G.; Hefford, M.; Jiang, X.; Lee, T.C.; Madduri, K.M.; Pariza, M.; Privalle, L.S.; et al. Characteristics and safety assessment of intractable proteins in genetically modified crops. Regul. Toxicol. Pharmacol. 2014, 69, 154–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Cao, S.; He, X.; Luo, Y.; Guo, X.; Yuan, Y.; Huang, K. Safety assessment of Cry1Ab/Ac fusion protein. Food Chem. Toxicol. 2009, 47, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Huber, S.C.; Huber, J.L. Role of sucrose-phosphate synthase in sucrose metabolism in leaves. Plant Physiol. 1992, 99, 1275–1278. [Google Scholar] [CrossRef] [PubMed]
- Herouet, C.; Esdaile, D.J.; Mallyon, B.A.; Debruyne, E.; Schulz, A.; Currier, T.; Hendrickx, K.; van der Klis, R.J.; Rouan, D. Safety evaluation of the phosphinothricin acetyltransferase proteins encoded by the pat and bar sequences that confer tolerance to glufosinate-ammonium herbicide in transgenic plants. Regul. Toxicol. Pharmacol. 2005, 41, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Wehrmann, A.; Van Vliet, A.; Opsomer, C.; Botterman, J.; Schulz, A. The similarities of bar and pat gene products make them equally applicable for plant engineers. Nat. Biotechnol. 1996, 14, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Mathur, C.; Kathuria, P.C.; Dahiya, P.; Singh, A.B. Lack of detectable allergenicity in genetically modified maize containing “Cry” proteins as compared to native maize based on in silico & in vitro analysis. PLoS ONE 2015, 10, e0117340. [Google Scholar] [CrossRef]
- Menezes, S.P.; Santos, J.L.; Cardoso, T.H.S.; Pirovani, C.P.; Micheli, F.; Noronha, F.S.M.; Alves, A.C.; Faria, A.M.C.; Gesteira, A.S. Evaluation of the allergenicity potential of TcPR-10 protein from Theobromo cacao. PLoS ONE 2012, 7, e37969. [Google Scholar] [CrossRef]
- Fritsche, R. Animal models in food allergy: Assessment of allergenicity and preventive activity of infant formulas. Toxicol. Lett. 2003, 140–141, 303–309. [Google Scholar] [CrossRef]
- Adkinson, N.F.; Bochner, B.S.; Busse, W.W.; Holgate, S.T.; Lemanske, R.F.; Simons, F.E.R. Middleton’s Allergy Principles and Practice, 7th ed.; Elsevier: Philadelphia, PA, USA, 2009; p. 440. ISBN 978-0-323-04884-2. [Google Scholar]
- OECD. Novel Food and Feed Safety: Safety Assessment of Foods and Feeds Derived from Transgenic Crops; OECD Publishing: Paris, France, 2015; pp. 174–179. ISBN 978-92-64-18033-8. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Allergen | Species | Description | Sequence in FARRP * | Sequence Length | Matching Region of Contiguous Amino Acids |
|---|---|---|---|---|---|
| Chitinase | Zea mays L. | Food allergen | GI:260401081 | 279 | 62-GGGGGGGG-69 |
| Endochitinase A | Zea mays L. | Food allergen | GI:116329 | 281 | 61-GGGGGGGG-68 |
| Death of Animals | |||||
|---|---|---|---|---|---|
| Group Treatment | Sample Treatment | Dosage (gr/kg) | After 72 h | After 14 Days | Percentage (%) * |
| I | Control | 0 | 0 | 0 | 0 |
| II | Sugarcane Juice | 6.25 | 0 | 0 | 0 |
| II | Sugarcane Juice | 12.5 | 0 | 0 | 0 |
| IV | Sugarcane Juice | 25 | 0 | 0 | 0 |
| Dosage of Sugarcane Juice | ||||
|---|---|---|---|---|
| Organs | 0 gr/kg | 6.25 gr/kg | 12.5 gr/kg | 25 gr/kg |
| Heart | 0.37 ± 0.03 | 0.44 ± 0.03 | 0.41 ± 0.03 | 0.44 ± 0.04 |
| Liver | 5.74 ± 0.68 | 6.25 ± 0.19 | 7.06 ± 0.07 | 6.80 ± 0.10 |
| Spleen | 0.77 ± 0.17 | 0.79 ± 0.06 | 0.85 ± 0.02 | 0.84 ± 0.11 |
| Kidneys | 1.31 ± 0.10 | 1.56 ± 0.03 | 1.29 ± 0.07 | 1.43 ± 0.12 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neliana, I.R.; Sawitri, W.D.; Ermawati, N.; Handoyo, T.; Sugiharto, B. Development of Allergenicity and Toxicity Assessment Methods for Evaluating Transgenic Sugarcane Overexpressing Sucrose–Phosphate Synthase. Agronomy 2019, 9, 23. https://doi.org/10.3390/agronomy9010023
Neliana IR, Sawitri WD, Ermawati N, Handoyo T, Sugiharto B. Development of Allergenicity and Toxicity Assessment Methods for Evaluating Transgenic Sugarcane Overexpressing Sucrose–Phosphate Synthase. Agronomy. 2019; 9(1):23. https://doi.org/10.3390/agronomy9010023
Chicago/Turabian StyleNeliana, Intan Ria, Widhi Dyah Sawitri, Netty Ermawati, Tri Handoyo, and Bambang Sugiharto. 2019. "Development of Allergenicity and Toxicity Assessment Methods for Evaluating Transgenic Sugarcane Overexpressing Sucrose–Phosphate Synthase" Agronomy 9, no. 1: 23. https://doi.org/10.3390/agronomy9010023
APA StyleNeliana, I. R., Sawitri, W. D., Ermawati, N., Handoyo, T., & Sugiharto, B. (2019). Development of Allergenicity and Toxicity Assessment Methods for Evaluating Transgenic Sugarcane Overexpressing Sucrose–Phosphate Synthase. Agronomy, 9(1), 23. https://doi.org/10.3390/agronomy9010023