Integrative Effects of Rice-Straw Biochar and Silicon on Oil and Seed Quality, Yield and Physiological Traits of Helianthus annuus L. Grown under Water Deficit Stress

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Field Practices
2.2. Measurements
2.2.1. Physiological and Yield Traits
2.2.2. Mycorrhizal Spores and Plant Analysis at Harvest
2.2.3. Quality Traits
2.3. Statistical Analysis
3. Results
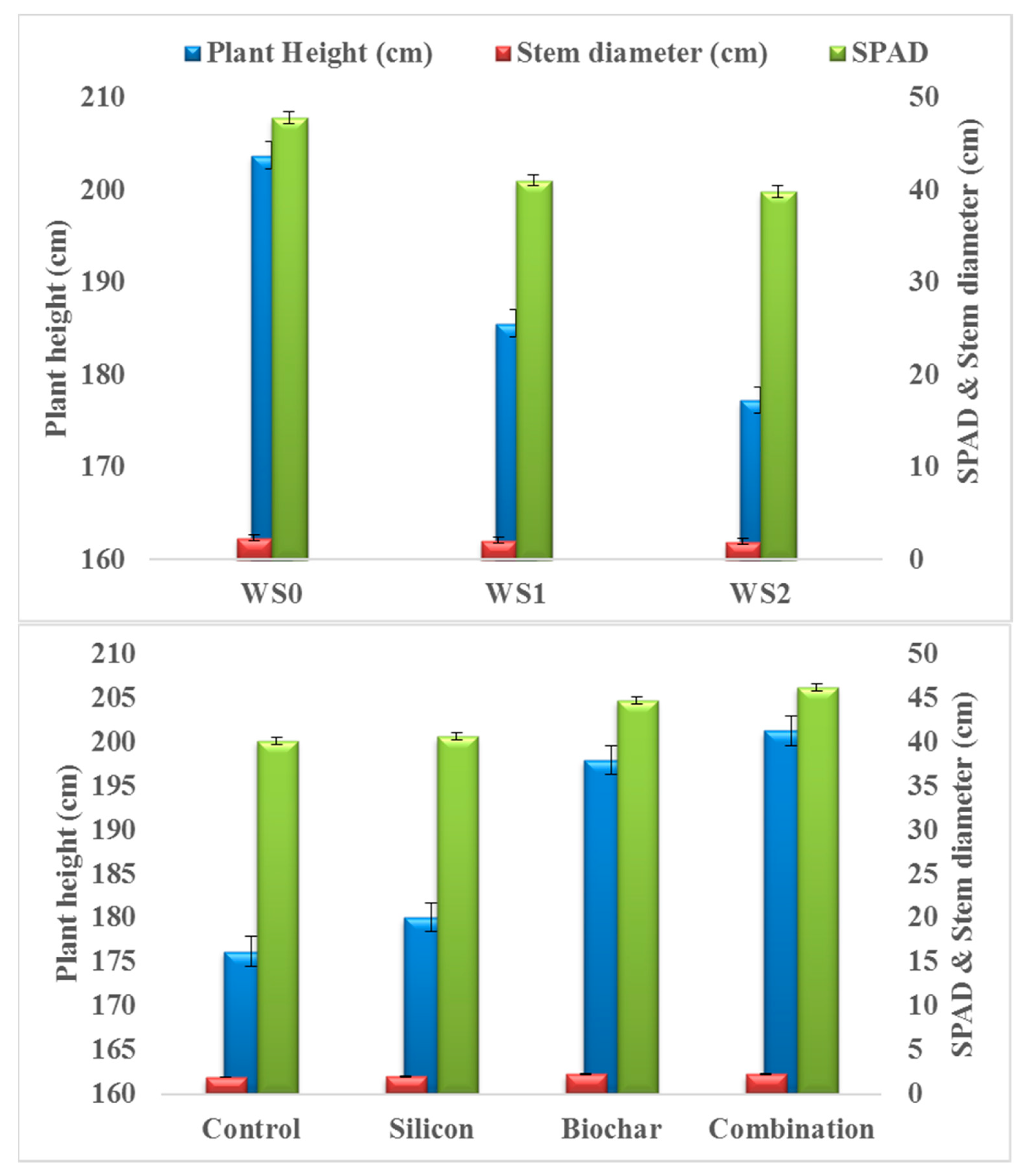
3.1. Physiological and Yield Traits
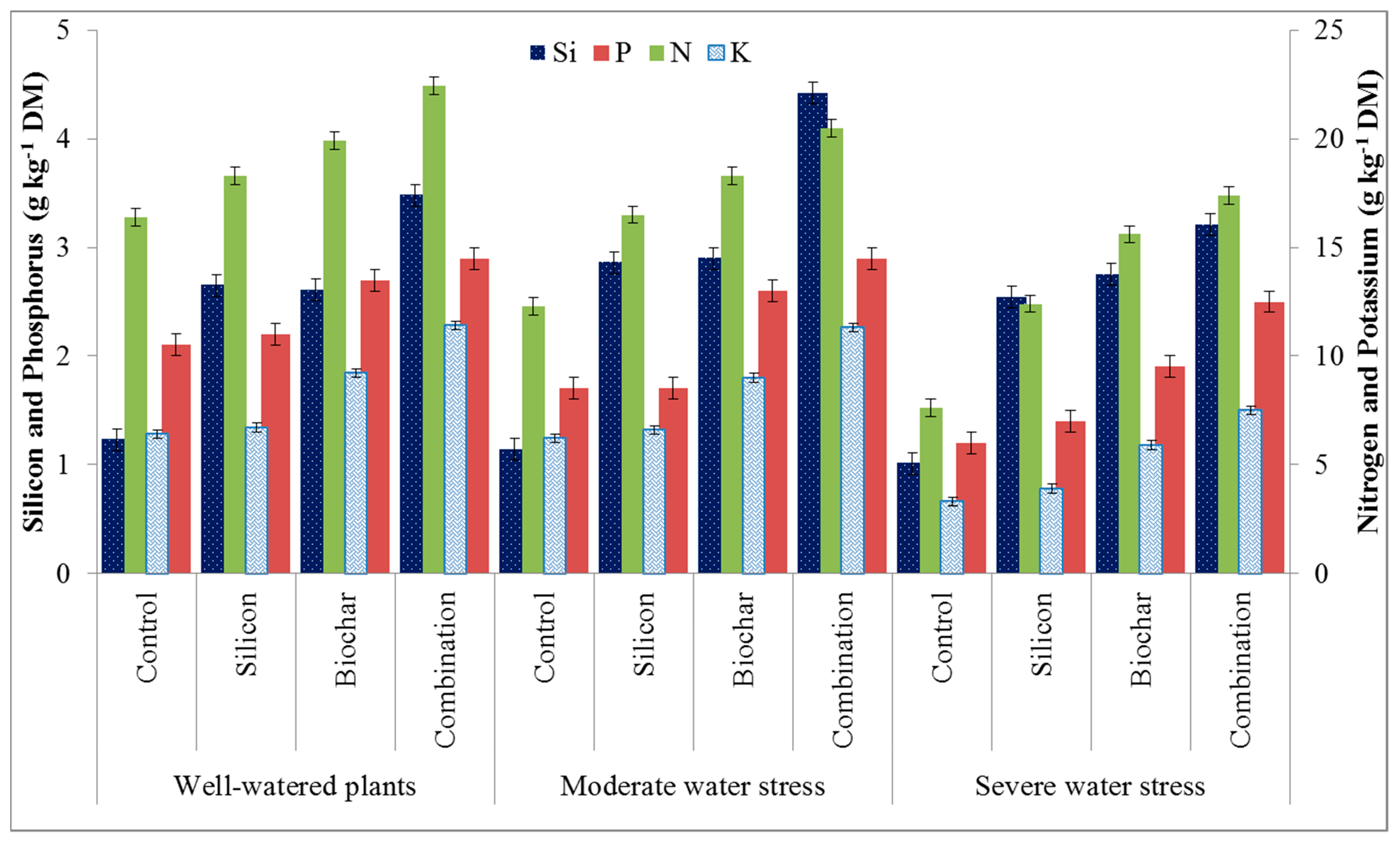
3.2. Mycorrhizal Spores and Plant Elemental Analysis
3.3. Oil and Seed Quality Traits
4. Discussion
4.1. Physiological and Yield Traits
4.2. Mycorrhizal Spores and Plant Elemental Analysis
4.3. Oil and Seed Quality Traits
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, X.; Wang, J.; Wang, S.; Xing, G. Successive straw biochar application as a strategy to sequester carbon and improve fertility: A pot experiment with two rice/wheat rotations in paddy soil. Plant Soil 2014, 378, 279–294. [Google Scholar] [CrossRef]
- Lehmann, J.; Joseph, S. Biochar for Environmental Management: Science, Technology and Implementation, 2nd ed.; Routledge, Taylor & Francis Publisher: Oxfordshire, UK, 2015. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Kheir, A.S. Maize productivity, heavy metals uptake and their availability in contaminated clay and sandy alkaline soils as affected by inorganic and organic amendments. Chemosphere 2018, 204, 514–522. [Google Scholar] [CrossRef]
- Shi, J.; Fan, X.; Tsang, D.C.W.; Wang, F.; Shen, Z.; Hou, D.; Alessi, D.S. Removal of lead by rice husk biochars produced at different temperatures and implications for their environmental utilizations. Chemosphere 2019, 235, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Rafique, M.; Ortas, I.; Rizwan, M.; Chaudhary, J.H.; Gurmani, A.R.; Munis, M.F.H. Residual effects of biochar and phosphorus on growth and nutrient accumulation by maize (Zea mays L.) amended with microbes in texturally different soils. Chemosphere 2019, 238, 124710. [Google Scholar] [CrossRef] [PubMed]
- Githinji, L. Effect of biochar application rate on soil physical and hydraulic properties of a sandy loam. Arch. Agron. Soil Sci. 2014, 60, 457–470. [Google Scholar] [CrossRef]
- Stewart, C.E.; Zheng, J.; Botte, J.; Cotrufo, M.F. Co-generated fast pyrolysis biochar mitigates green-house gas emissions and increases carbon sequestration in temperate soils. Glob. Chang. Biol. Bioenergy 2013, 5, 153–164. [Google Scholar] [CrossRef]
- Xiao, X.; Chen, B.; Zhu, L. Transformation, morphology and dissolution of silicon and carbon in rice straw derived biochars under different pyrolytic temperatures. Environ. Sci. Technol. 2014, 48, 3411–3419. [Google Scholar] [CrossRef]
- Biederman, L.A.; Harpole, W.S. Biochar and its effects on plant productivity and nutrient cycling: A meta-analysis. Glob. Chang. Biol. Bioenergy 2013, 5, 202–214. [Google Scholar] [CrossRef]
- Rizwan, M.; Rehman, M.; Ali, S.; Abbas, T.; Maqbool, A.; Bashir, A. Biochar is a potential source of silicon fertilizer: An overview. In Biochar from Biomass and Waste; Ok, Y.S., Tsang, D.C.W., Bolan, N., Novak, J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 225–235. [Google Scholar] [CrossRef]
- Li, Z.; Song, Z.; Singh, B.; Wang, H. The impact of crop residue biochars on silicon and nutrient cycles in croplands. Sci. Total Environ. 2019, 659, 673–680. [Google Scholar] [CrossRef]
- Hashem, A.; Kumar, A.; Al-Dbass, A.M.; Alqarawi, A.A.; Al-Arjani, A.F.; Singh, G.; Farooq, M.; Abd-Allah, E.F. Arbuscular mycorrhizal fungi and biochar improves drought tolerance in chickpea. Saudi J. Biol. Sci. 2019, in press. [Google Scholar] [CrossRef]
- Gavili, E.; Moosavi, A.A.; Kamgar Haghighi, A.A.K. Does biochar mitigate the adverse effects of drought on the agronomic traits and yield components of soybean? Ind. Crops Prod. 2019, 128, 445–454. [Google Scholar] [CrossRef]
- Mehari, Z.H.; Elad, Y.; Rav-David, D.; Graber, E.R.; Harel, Y.M. Induced systemic resistance in tomato (Solanum lycopersicum) against Botrytis cinerea by biochar amendment involves jasmonic acid signaling. Plant Soil 2015, 395, 31–44. [Google Scholar] [CrossRef]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s role in abiotic and biotic plant stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.; Brown, P.; Cakmak, I.; Feng, M.J.; Rengel, Z.; Zhao, F. Beneficial elements. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 249–269. [Google Scholar] [CrossRef]
- Gunes, A.; Kadioglu, Y.K.; Pilbeam, D.J.; Inal, A.; Coban, S.; Aksu, A. Influence of silicon on sunflower cultivars under drought stress, II: Essential and nonessential element uptake determined by polarized energy dispersive X-ray fluorescence. Commun. Soil Sci. Plant Anal. 2008, 39, 1904–1927. [Google Scholar] [CrossRef]
- Tubana, B.T.; Heckman, J.R. Silicon in soils and plants. In Silicon and Plant Disease; Rodrigues, F.A., Datnoff, L.E., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 7–51. [Google Scholar]
- Tubana, B.S.; Babu, T.; Datnof, L.E. A Review of silicon in soils and plants and its role in us agriculture: History and future perspectives. Soil Sci. 2016, 181, 393–411. [Google Scholar] [CrossRef]
- Norton, L.D.; Hall, G.F.; Smeck, N.E.; Bigham, J.M. Fraginap bonding in a late-Wisconsian loss-derived soil in East-Central Ohio. Soil Sci. Soc. Am. J. 1984, 48, 1360. [Google Scholar] [CrossRef]
- Deshmukh, R.; Belanger, R.R. Molecular evolution of aquaporins and Si influx in plants. The functional role of Si in plant Biology. Funct. Ecol. 2016, 30, 1277–1285. [Google Scholar] [CrossRef]
- Kaur, H.; Greger, M. A Review on si uptake and transport system. Plants 2019, 8, 81. [Google Scholar] [CrossRef]
- Ma, J.F.; Takahashi, E. Si-accumulating plants in the plant kingdom. In Soil, Fertilizer and Plant Si Research in Japan; Elsevier Science: Amsterdam, The Netherlands, 2002; pp. 63–71. [Google Scholar]
- Hodson, M.; White, P.; Mead, A.; Broadley, M. Phylogenetic variation in the Si composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef]
- Wang, M.; Gao, L.; Dong, S.; Sun, Y.; Shen, Q.; Guo, S. Role of silicon on plant-pathogen interactions. Front. Plant Sci. 2017, 8, 701. [Google Scholar] [CrossRef]
- Rodrigues, F.A.; Resende, R.S.; Dallagnol, L.J.; Datnoff, L.E. Silicon Potentiates Host Defense Mechanisms against Infection by Plant Pathogens; Springer International Publishing: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Van Bockhaven, J.; De Vleesschauwer, D.; Höfte, M. Towards establishing broad-spectrum disease resistance in plants: Silicon leads the way. J. Exp. Bot. 2013, 64, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Vivancos, J.; Labbe, C.; Menzies, J.G.; Belanger, R.R. Silicon-mediated resistance of Arabidopsis against powdery mildew involves mechanisms other than the salicylic acid (SA)-dependent defence pathway. Mol. Plant Pathol. 2015, 16, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shi, Y.; Gong, H.; Zhao, H.; Li, H.; Hu, Y.; Wang, Y. Beneficial effects of silicon on photosynthesis of tomato seedlings under water stress. J. Integ. Agric. 2018, 17, 2151–2159. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Chen, K.; Zhao, Z.; Chen, G.; Zhou, W. Effects of silicon on defense of wheat against oxidative stress under drought at different developmental stages. Biol. Plant. 2008, 52, 592–596. [Google Scholar] [CrossRef]
- Liu, P.; Yin, L.; Deng, X.; Wang, S.; Tanaka, K.; Zhang, S. Aquaporin-mediated increase in root hydraulic conductance is involved in silicon-induced improved root water uptake under osmotic stress in Sorghum bicolor L. J. Exp. Bot. 2014, 65, 4747–4756. [Google Scholar] [CrossRef] [PubMed]
- Korndörfer, G.H.; Lepsch, I. Effect of silicon on plant growth and crop yield. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndorfer, G.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2001; pp. 133–147. [Google Scholar] [CrossRef]
- Gong, H.J.; Chen, K.M. The regulatory role of silicon on water relations, photosynthetic gas exchange and carboxylation activities of wheat leaves in field drought conditions. Acta Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Trenholm, L.E.; Datnoff, L.E.; Nagara, R.T. Influence of silicon on drought and shade tolerance of St. Augustinegrass. HortTechnology 2004, 14, 487–490. [Google Scholar] [CrossRef]
- Hafez, E.H.; Seleiman, M.F. Response of barley quality traits, yield and antioxidant enzymes to water-stress and chemical inducers. Inter. J. Plant Prod. 2017, 11, 477–490. [Google Scholar] [CrossRef]
- Bray, E.A.; Bailey-Serres, J.; Weretilnyk, E. Responses to abiotic stresses. In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 1158–1203. [Google Scholar]
- Petcu, E.; Arsintescu, A.; Stanciu, D. The effect of drought stress on fatty acid composition in some Romanian sunflower hybrids. Rom. Agric. Res. 2001, 15, 39–42. [Google Scholar]
- Eslami, M. The effect of drought stress on oil percent and yield and the type of sunflower (Helianthus annuus L.) fatty acids. Agri. Sci. Develop. 2015, 4, 1–3. [Google Scholar]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveere, M.; Farooq, M.; Nawazd, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Louise, H.; Thomas, J.; Kendall, C.; DeJonge, H.Z.; Sean, M.G. Water productivity under strategic growth stage-based deficit irrigation in maize. Agric. Water Manag. 2019, 212, 433–440. [Google Scholar] [CrossRef]
- Moradi-Ghahderijani, M.; Jafarian, S.; Keshavarz, H. Alleviation of water stress effects and improved oil yield in sunflower by application of soil and foliar amendments. Rhizosphere 2017, 4, 54–61. [Google Scholar] [CrossRef]
- Seleiman, M. Use of plant nutrients in improving abiotic stress tolerance in wheat. In Wheat Production in Changing Environments; Hasanuzzaman, M., Nahar, K., Hossain, M.A., Eds.; Springer: Singapore, 2019; pp. 481–495. [Google Scholar] [CrossRef]
- García-López, J.; Lorite, I.J.; García-Ruiz, R.; Domínguez, J. Evaluation of three simulation approaches for assessing yield of rainfed sunflower in a Mediterranean environment for climate change impact modelling. Clim. Chang. 2014, 124, 147–162. [Google Scholar] [CrossRef]
- Flagella, Z.; Rotunno, T.E.; Tatantino, E.; Di Caterina, R.; De Caro, A. Changes in seed yield and oil fatty acid composition of high oleic sunflower (Helianthus annuus L.) hybrids in relation to the sowing date and the water regime. Eur. J. Agron. 2002, 17, 221–230. [Google Scholar] [CrossRef]
- Anastasi, U.; Santonocetoa, C.; Giuffrèa, A.M.; Sortinob, O.; Grestab, F.; Abbateb, V. Yield performance and grain lipid composition of standard and oleic sunflower as affected by water supply. Field Crops Res. 2010, 119, 145–153. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization of the United Nations Statistics Division. 2019. Available online: http://faostat.fao.org/site/567/DesktopDefault.aspx (accessed on 20 August 2019).
- Ghobadi, M.; Sh, T.; Mohammad, E.G.; Gholam, R.M.; Saeid, J.H. Antioxidant capacity, photosynthetic characteristics and water relations of sunflower (Helianthus annuus L.) cultivars in response to drought stress. Ind. Crops Prod. 2013, 50, 29–38. [Google Scholar] [CrossRef]
- Meier, U. BBCH-Monograph: Growth Stages of Mono-and Dicotyledonous Plants, 2nd ed.; Federal Biological Research Centre for Agriculture and Forestry: Berlin, Germany, 2001. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Kleemola, J.; Stoddard, F.L.; Mäkelä, P.S.A. Improved sustainability of feedstock production with sludge and interacting mychorriza. Chemosphere 2013, 91, 1236–1242. [Google Scholar] [CrossRef]
- Allen, M.F.; Moore, T.S.; Christensen, J.R.M. Growth ofvesicular-arbuscular mycorrhizal and non-mycorrhizal Bouteloua gracilis in adefinedmedium. Mycologia 1979, 71, 666–669. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Stoddard, F.L.; Makela, P. Feedstock quality and growth of bioenergy crops fertilized with sewage sludge. Chemosphere 2012, 89, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Pomeranz, Y.; Clifton, E. Food Analysis: Theory and Practice, 3rd ed.; Kluwer Academic Publisher: San Diego, CA, USA, 1994. [Google Scholar] [CrossRef]
- Rotunno, T.; Sevi, A.; Caterina, R.D.; Muscio, A. Effects of graded levels of dietary rumen-protected fat on milk characteristics of Comisana ewes. Small Rumin. Res. 1998, 30, 137–145. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Kheir, A.M.S.; Al-Dhumri, S.; Alghamdi, A.G.; Omar, E.-S.H.; Aboelsoud, H.M.; Abdella, K.A.; Abou El Hassan, W.H. Exploring optimal tillage improved soil characteristics and productivity of wheat irrigated with different water qualities. Agronomy 2019, 9, 233. [Google Scholar] [CrossRef]
- Pfister, M.; Saha, S. Effects of biochar and fertilizer management on sunflower (Helianthus annuus L.) feedstock and soil properties. Arch. Agron. Soil Sci. 2017, 63, 651–662. [Google Scholar] [CrossRef]
- Baronti, S.; Vaccari, F.P.; Miglietta, F.; Calzolari, C.; Lugato, E.; Orlandini, S.; Pini, R.; Zulian, C.; Genesio, L. Impact of biochar application on plant water relations in Vitis vinifera (L.). Eur. J. Agron. 2014, 53, 38–44. [Google Scholar] [CrossRef]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, L.; Higgs, D. Effect of silicon on plant growth and mineral nutrition of maize grown under water-stress conditions. J. Plant Nutr. 2006, 29, 1469–1480. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Tezara, W.; Mitchell, V.J.; Driscoll, S.D.; Lawlor, D.W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 1999, 401, 914. [Google Scholar] [CrossRef]
- Ramzani, P.M.A.; Khan, W.U.D.; Iqbal, M.; Kausar, S.; Ali, S.; Rizwan, M.; Virk, Z.A. Effect of different amendments on rice (Oryza sativa L.) growth, yield, nutrient uptake and grain quality in Ni-contaminated soil. Environ. Sci. Pollut. Res. 2016, 23, 18585–18595. [Google Scholar] [CrossRef]
- Paneque, M.; De, R.J.; Franco, N.J.; Colmenero, F.J.; Knicker, H. Effect of biochar amendment on morphology, productivity and water relations of sunflower plants under non-irrigation conditions. Catena 2016, 147, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Agarie, S.; Uchida, H.; Agata, W.; Kubota, F.; Kaufman, P.B. Effects of silicon on transpiration and leaf conductance in rice plants (Oryza sativa L.). Plant Prod. Sci. 1998, 1, 89–95. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Gong, H.J. Beneficial effects of silicon on salt and drought tolerance in plants. Agron. Sustain. Dev. 2014, 34, 455–472. [Google Scholar] [CrossRef]
- Benlloch-González, M.; Quintero, J.M.; García-Mateo, M.J.; Fournier, J.M.; Benlloch, M. Effect of water stress and subsequent re-watering on K+ and water flows in sunflower roots: A possible mechanism to tolerate water stress. Environ. Exper. Bot. 2015, 118, 78–84. [Google Scholar] [CrossRef]
- Jabari, H.; Akbari, G.A.; Daneshian, J.; Alahdadi, I.; Shahbazian, N. Effect of water deficit stress on agronomic characteristics of sunflower hybrids. Agric. Res. 2007, 9, 13–22. [Google Scholar]
- Wang, J.; Xiong, Z.; Kuzyakov, Y. Biochar stability in soil: Meta-analysis of decomposition and priming effects. Glob. Chang. Biol. Bioenergy 2016, 8, 512–523. [Google Scholar] [CrossRef]
- Ding, Y.; Liu, Y.-X.; Wu, W.; Shi, D.-Z.; Yang, M.; Zhong, Z. Evaluation of biochar effects on nitrogen retention and leaching in multi-layered soil columns. Water Air Soil Poll. 2010, 213, 47–55. [Google Scholar] [CrossRef]
- Rodriguez, D.J.; Philips, D.B.S.; Rodriguez-Garcia, R.; Angulo-Sanchez, J.L. Grain yield and fatty acid composition of sunflower seed for cultivars developed under dry land conditions. In Trends in New Crops and New Uses; Janick, J., Whipkey, A., Eds.; American Society for Horticultural Science Press: Alexandria, Egypt, 2002; pp. 139–142. [Google Scholar]
- Eneji, A.E.; Inanaga, S.; Muranaka, S.; Li, J.; Hattori, T.; An, P.; Tsuji, W. Growth and nutrient use in four grasses under drought stress as mediated by silicon fertilisers. J. Plant Nutr. 2008, 31, 355–365. [Google Scholar] [CrossRef]
- Mali, M.; Aery, N.C. Silicon effects on nodule growth, dry matter production and mineral nutrition of cowpea (Vigna unguiculata). J. Plant Nutr. Soil Sci. 2008, 171, 835–840. [Google Scholar] [CrossRef]
- Dorrell, D.G.; Vick, B.A. Properties and processing of oilseed sunflower. In Sunflower Technology and Production; Schneiter, A.A., Ed.; Agronomy Monograph 35, ASA, CSSA and SSSA: Madison, WI, USA, 1997; pp. 709–745. [Google Scholar]
- Baldini, M.; Giovanardi, R.; Tahmasebi-Enferadi, S.; Vannozzi, G.P. Effects of water regimes on fatty acid accumulation and final fatty acid composition in the oil of standard and high oleic sunflower hybrids. Ital. J. Agron. 2002, 6, 119–126. [Google Scholar]
- Göksoy, A.T.; Demir, A.O.; Turan, Z.M.; Dağüstü, N. Responses of sunflower (Helianthus annuus L.) to full and limited irrigation at different growth stages. Field Crops Res. 2004, 87, 167–178. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physical and chemical properties | ||||||||
| Properties | EC (dS m−1) | pH | OM (%) | N | P | K | Total Si (g kg−1) | |
| Seasons | Available (meq L−1) | |||||||
| 2016 | 0.80 | 7.2 | 1.75 | 28.01 | 9.73 | 297.4 | 0.31 | |
| 2017 | 0.84 | 7.0 | 1.82 | 27.34 | 9.44 | 291.6 | 0.36 | |
| Mechanical properties | ||||||||
| Properties | Sand (%) | Silt (%) | Clay (%) | Soil texture | ||||
| Seasons | ||||||||
| 2016 | 21.00 | 41.04 | 37.96 | Clay loam | ||||
| 2017 | 21.03 | 42.02 | 36.95 | Clay loam | ||||
| Traits | Stomatal Conductance gs (Mmol H2O m−2 S−1) | Proline (μ mol g−1 FW) | POX Activity (Mmol H2O2 FW min−1) | CAT Activity (Mmol H2O2 FW min−1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments | |||||||||
| Water Stress | Amendments | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 |
| WS0 | Control | 48.87 | 49.56 | 8.90 | 9.20 | 1.18 | 1.38 | 150.33 | 153.7 |
| Silicon | 50.78 | 51.48 | 9.54 | 9.84 | 1.04 | 1.24 | 125.33 | 127.0 | |
| Biochar | 53.73 | 54.42 | 8.70 | 9.00 | 0.75 | 0.95 | 72.67 | 76.6 | |
| Combination | 55.06 | 55.76 | 8.14 | 8.44 | 0.60 | 0.80 | 44.33 | 48.3 | |
| WS1 | Control | 38.76 | 40.10 | 10.59 | 10.56 | 2.21 | 2.75 | 229.20 | 192.0 |
| Silicon | 41.91 | 43.19 | 9.24 | 9.53 | 1.92 | 2.36 | 176.80 | 149.3 | |
| Biochar | 45.44 | 46.78 | 9.22 | 9.50 | 1.52 | 1.85 | 126.00 | 105.0 | |
| Combination | 47.19 | 48.57 | 9.02 | 9.30 | 1.34 | 1.55 | 97.60 | 85.3 | |
| WS2 | Control | 37.53 | 37.72 | 13.67 | 13.77 | 3.23 | 3.33 | 252.00 | 214.0 |
| Silicon | 38.60 | 38.79 | 12.69 | 12.79 | 3.06 | 3.16 | 191.60 | 162.6 | |
| Biochar | 40.96 | 41.16 | 11.82 | 11.92 | 2.55 | 2.65 | 137.20 | 119.3 | |
| Combination | 42.07 | 42.27 | 11.30 | 11.40 | 2.21 | 2.31 | 121.20 | 106.0 | |
| S.E.M | 0.56 | 0.59 | 0.042 | 0.113 | 0.049 | 0.051 | 3.78 | 3.4 | |
| Significant | |||||||||
| Water stress (WS) | * | ** | ** | ** | ** | ** | ** | * | |
| Amendments (A) | ** | ** | ** | ** | ** | ** | ** | ** | |
| WS × A | ** | ** | ** | ** | ** | ** | ** | ** | |
| Traits | Leaf Area Plant−1 (cm2) | Relative Water Content (%) | |||
|---|---|---|---|---|---|
| Treatments | |||||
| Water stress | Amendments | S1 | S2 | S1 | S2 |
| WS0 | Control | 5768.3 | 5868.3 | 80.43 | 83.93 |
| Biochar | 6765.3 | 6865.3 | 89.50 | 93.00 | |
| Silicon | 5889.3 | 5989.3 | 84.20 | 87.70 | |
| Combination | 6873.6 | 6973.7 | 92.10 | 95.60 | |
| WS1 | Control | 5104.1 | 5224.5 | 61.88 | 65.60 |
| Biochar | 5988.5 | 5506.9 | 76.51 | 80.54 | |
| Silicon | 5211.8 | 5333.4 | 69.29 | 73.17 | |
| Combination | 6102.7 | 5957.2 | 79.29 | 83.39 | |
| WS2 | Control | 3441.0 | 3471.0 | 45.03 | 46.53 |
| Biochar | 4710.0 | 4740.1 | 68.47 | 69.97 | |
| Silicon | 4099.2 | 4129.2 | 61.47 | 62.97 | |
| Combination | 5142.7 | 5172.8 | 71.43 | 72.93 | |
| S.E.M | 15.6 | 15.4 | 0.28 | 0.281 | |
| Significant | |||||
| Water stress (WS) | ** | ** | ** | ** | |
| Amendments (A) | ** | ** | ** | ** | |
| WS × A | ** | ** | ** | ** | |
| Traits | Head Diameter (cm) | Number of Seeds Per Head | 100-Seed Weight (g) | Seed Yield (kg ha−1) | |||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments | |||||||||
| Water Stress | Amendments | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 |
| WS0 | Control | 18.2 | 19.1 | 1033 | 1053 | 6.27 | 6.57 | 2914.0 | 2933.9 |
| Silicon | 18.6 | 19.5 | 1231 | 1251 | 6.43 | 6.73 | 3155.0 | 3174.9 | |
| Biochar | 22.0 | 22.9 | 1440 | 1460 | 7.13 | 7.43 | 3619.3 | 3639.3 | |
| Combination | 22.6 | 23.5 | 1555 | 1575 | 7.23 | 7.53 | 3973.0 | 3993.0 | |
| WS1 | Control | 16.7 | 17.3 | 942 | 956 | 5.57 | 5.58 | 2574.4 | 2316.1 |
| Silicon | 16.7 | 18.3 | 979 | 991 | 5.83 | 5.83 | 2661.0 | 2988.3 | |
| Biochar | 19.5 | 19.6 | 1120 | 1075 | 6.59 | 6.56 | 2964.3 | 2977.0 | |
| Combination | 19.8 | 19.8 | 1161 | 1158 | 6.81 | 6.78 | 3144.4 | 3268.6 | |
| WS2 | Control | 12.0 | 12.3 | 702 | 707 | 4.10 | 4.20 | 1294.6 | 1304.5 |
| Silicon | 14.0 | 14.3 | 810 | 815 | 4.53 | 4.63 | 1509.0 | 1518.9 | |
| Biochar | 17.6 | 17.9 | 932 | 937 | 4.87 | 4.97 | 1634.8 | 1644.8 | |
| Combination | 18.3 | 18.6 | 959 | 964 | 5.47 | 5.57 | 1953.5 | 1963.4 | |
| S.E.M | 0.22 | 0.23 | 25 | 26 | 0.075 | 0.074 | 39.9 | 39.7 | |
| Significant | |||||||||
| Water stress (WS) | ** | ** | ** | ** | ** | * | ** | ** | |
| Amendments (A) | ** | ** | ** | ** | ** | * | ** | ** | |
| WS × A | * | * | ** | ** | ** | * | ** | ** | |
| Traits | Oil (%) | Oil Yield (kg ha−1) | Protein (%) | Oleic Acid (%) | Linoleic Acid (%) | Oleic/Linoleic Ratio | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | |||||||||||||
| Water Stress | Amendments | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 | S1 | S2 |
| WS0 | Control | 41.9 | 42.0 | 1221 | 1232 | 19.3 | 20.0 | 85.0 | 84.5 | 5.8 | 6.2 | 14.7 | 13.6 |
| Silicon | 43.4 | 43.6 | 1369 | 1384 | 20.0 | 20.8 | 86.4 | 86.9 | 4.7 | 5.0 | 18.4 | 17.4 | |
| Biochar | 45.3 | 45.5 | 1639 | 1655 | 19.5 | 20.0 | 87.4 | 87.9 | 4.2 | 4.5 | 20.8 | 19.5 | |
| Combination | 45.4 | 45.5 | 1803 | 1816 | 20.3 | 21.3 | 88.4 | 88.7 | 3.7 | 4.1 | 23.9 | 21.6 | |
| WS1 | Control | 40.9 | 41.4 | 1052 | 958 | 20.5 | 21.1 | 72.8 | 73.2 | 17.6 | 17.9 | 4.2 | 4.1 |
| Silicon | 42.4 | 42.4 | 1128 | 1267 | 21.2 | 21.7 | 74.6 | 75.6 | 16.5 | 16.8 | 4.5 | 4.6 | |
| Biochar | 45.1 | 45.2 | 1336 | 1345 | 20.7 | 21.6 | 82.7 | 83.0 | 7.3 | 7.5 | 11.3 | 11.1 | |
| Combination | 45.1 | 45.0 | 1418 | 1470 | 21.7 | 22.3 | 84.6 | 85.0 | 7.1 | 7.2 | 11.9 | 11.8 | |
| WS2 | Control | 33.6 | 33.7 | 435 | 439 | 22.5 | 22.7 | 60.5 | 60.7 | 31.5 | 31.2 | 1.8 | 1.9 |
| Silicon | 35.8 | 35.8 | 540 | 543 | 22.9 | 23.2 | 62.7 | 62.3 | 29.1 | 29.4 | 2.1 | 2.2 | |
| Biochar | 38.1 | 38.3 | 622 | 630 | 23.2 | 23.4 | 66.8 | 67.4 | 25.5 | 25.7 | 2.6 | 2.6 | |
| Combination | 39.1 | 39.1 | 763 | 767 | 23.6 | 23.9 | 67.9 | 68.1 | 23.9 | 24.3 | 2.9 | 2.8 | |
| S.E.M | 0.3 | 0.4 | 16.4 | 17.8 | 1.01 | 0.89 | 0.3 | 0.4 | 0.9 | 1.1 | 0.05 | 0.04 | |
| Significant | |||||||||||||
| Water stress (WS) | ** | ** | ** | ** | * | * | ** | ** | ** | ** | ** | ** | |
| Amendments (A) | * | * | ** | ** | * | * | ** | ** | ** | ** | ** | ** | |
| WS × A | * | * | ** | ** | * | * | ** | ** | ** | ** | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seleiman, M.F.; Refay, Y.; Al-Suhaibani, N.; Al-Ashkar, I.; El-Hendawy, S.; Hafez, E.M. Integrative Effects of Rice-Straw Biochar and Silicon on Oil and Seed Quality, Yield and Physiological Traits of Helianthus annuus L. Grown under Water Deficit Stress. Agronomy 2019, 9, 637. https://doi.org/10.3390/agronomy9100637
Seleiman MF, Refay Y, Al-Suhaibani N, Al-Ashkar I, El-Hendawy S, Hafez EM. Integrative Effects of Rice-Straw Biochar and Silicon on Oil and Seed Quality, Yield and Physiological Traits of Helianthus annuus L. Grown under Water Deficit Stress. Agronomy. 2019; 9(10):637. https://doi.org/10.3390/agronomy9100637
Chicago/Turabian StyleSeleiman, Mahmoud F., Yahya Refay, Nasser Al-Suhaibani, Ibrahim Al-Ashkar, Salah El-Hendawy, and Emad M. Hafez. 2019. "Integrative Effects of Rice-Straw Biochar and Silicon on Oil and Seed Quality, Yield and Physiological Traits of Helianthus annuus L. Grown under Water Deficit Stress" Agronomy 9, no. 10: 637. https://doi.org/10.3390/agronomy9100637
APA StyleSeleiman, M. F., Refay, Y., Al-Suhaibani, N., Al-Ashkar, I., El-Hendawy, S., & Hafez, E. M. (2019). Integrative Effects of Rice-Straw Biochar and Silicon on Oil and Seed Quality, Yield and Physiological Traits of Helianthus annuus L. Grown under Water Deficit Stress. Agronomy, 9(10), 637. https://doi.org/10.3390/agronomy9100637