Manure Fertilization Gives High-Quality Earthworm Coprolites with Positive Effects on Plant Growth and N Metabolism





Abstract
:1. Introduction
2. Results
2.1. Chemical Composition and Apparent Molecular Weight of HS
2.2. Spectroscopic Characterization of HS
2.3. Hormone-like Activity of HS
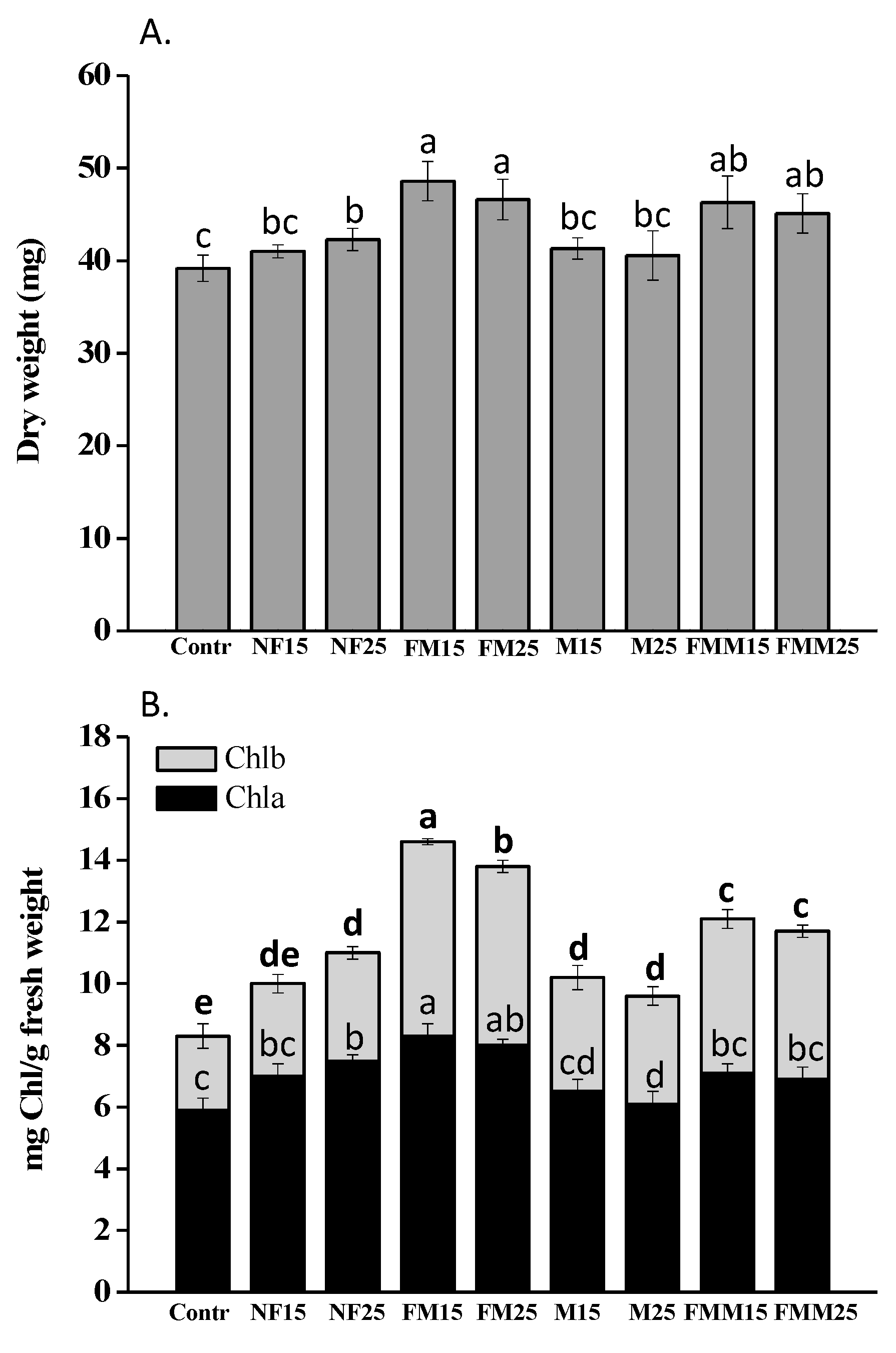
2.4. Effects of HS on Oat Plant Growth, Chlorophyll and Nutrient Content
2.5. Effects of HS on the Activity of N Assimilation Enzymes
3. Discussion
4. Materials and Methods
4.1. Experimental Setup and HS Extraction
4.2. Chemical Analyses and Apparent Molecular Weight Distribution of HS
4.3. ATR/FT-IR Analysis of HS
4.4. Determination of the Hormone-like Activities of HS
4.5. Plant Growth Conditions
4.6. Quantification of Chlorophyll Content and Mineral Nutrients
4.7. Enzyme Extraction and Assay Conditions
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Baker, G.H.; Brown, G.; Butt, K.; Curry, J.P.; Scullion, J. Introduced earth-worms in agricultural and reclaimed land: Their ecology and influences on soil properties, plant production and other soil biota. Biol. Invasions 2006, 8, 1301–1316. [Google Scholar] [CrossRef]
- Atiyeh, R.M.; Lee, S.; Edwards, C.A.; Arancon, N.Q.; Metzger, J.D. The in-fluence of humic acids derived from earthworm-processed organic wastes on plant growth. Bioresour. Technol. 2002, 84, 7–14. [Google Scholar] [CrossRef]
- Choosai, C.; Jouquet, P.; Hanboonsong, Y.; Hartmann, C. Effects of earthworms on soil properties and rice production in the rainfed paddy fields of Northeast Thailand. Appl. Soil Ecol. 2010, 45, 298–303. [Google Scholar] [CrossRef]
- Van Groenigen, J.W.; Lubbers, I.M.; Vos, H.M.J.; Brown, G.G.; De Deyn, G.B.; van Groenigen, K.J. Earthworms increase plant production: A meta-analysis. Sci. Rep. 2014, 4, 6365. [Google Scholar] [CrossRef] [PubMed]
- Kahsnitz, H.G. Investigations on the influence of earthworms on soil and plant. Bot. Arch. 1992, 1, 315–331. [Google Scholar]
- Gilot, C. Effects of a tropical geophagous earthworm, M. anomala (Megascolecidae), on soil characteristics and production of a yam crop in Cote d’Ivoire. Soil Biol. Biochem. 1997, 29, 353–359. [Google Scholar] [CrossRef]
- Muscolo, A.; Bovalo, F.; Gionfriddo, F.; Nardi, S. Earthworm humic matter produces auxin-like effects on Daucus carota cell growth and nitrate metabolism. Soil Biol. Biochem. 1999, 31, 1303–1311. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Nardi, S. Humic substance: Relationship between structure and activity. Deeper information suggests univocal findings. J. Geochem. Explor. 2013, 129, 57–63. [Google Scholar] [CrossRef]
- Vaccaro, S.; Muscolo, A.; Pizzeghello, D.; Spaccini, R.; Piccolo, A.; Nardi, S. Effect of a compost and its water-soluble fractions on key enzymes of nitrogen metabolism in maize seedlings. J. Agric. Food Chem. 2009, 57, 11267–11276. [Google Scholar] [CrossRef]
- Calderín García, A.; Huertas Tavares, O.; Martínez Balmori, D.; dos Santos Almeida, V.; Pasqualoto Canellas, L.; García-Mina, J.M.; Berbara1, L.L. Structure-function relationship of vermicompost humic fractions for use in agriculture. J. Soil Sediment 2018, 18, 1365–1375. [Google Scholar] [CrossRef]
- Darwin, C. The formation of vegetable mold through the action of worms, with observations of their habits. In The Essential Darwin; Ridley, M., Ed.; Allen and Unwin: London, UK, 1881; pp. 237–256. [Google Scholar]
- Evans, A.C. The importance of earthworms. Farming 1948, 2, 59–60. [Google Scholar]
- Muscolo, A.; Cutrupi, S.; Nardi, S. IAA detection in humic matter. Soil Biol. Biochem. 1998, 30, 1199–1201. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Okorokova-Facanha, A.L.; Façanha, A.R. Humic acids isolated from earthworm compost enhance root elongation, lateral root emergence, and plasma membrane H-ATPase activity in maize roots. Plant Physiol. 2002, 130, 1951–1957. [Google Scholar] [CrossRef] [PubMed]
- Nardi, S.; Morari, F.; Berti, A.; Tosoni, M.; Giardini, L. Soil organic matter properties after 40 years of different use of organic and mineral fertilisers. Eur. J. Agron. 2004, 21, 357–367. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Schiavon, M.; Ertani, A. Plant biostimulants: Physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 2016, 73, 18–23. [Google Scholar] [CrossRef]
- Piccolo, A. The supramolecular structure of humic substances. Soil Sci. 2001, 166, 810–832. [Google Scholar] [CrossRef]
- Sutton, R.; Sposito, G. Molecular structure in soil humic substances: The new view. Environ. Sci. Technol. 2005, 39, 9009–9015. [Google Scholar] [CrossRef]
- Swift, R.S. Macromolecular properties of soil humic substances: Fact, fiction and opinion. Soil Sci. 1999, 164, 790–802. [Google Scholar] [CrossRef]
- Clapp, C.E.; Hayes, M.H.B. Sizes and shapes of humic substances. Soil Sci. 1999, 164, 777–789. [Google Scholar] [CrossRef]
- Wershaw, R.L. Molecular aggregation of humic substances. Soil Sci. 1999, 164, 803–813. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Gerke, J. Concepts and misconceptions of humic substances as the stable part of soil organic matter: A review. Agronomy 2018, 8, 76. [Google Scholar] [CrossRef]
- Shah, Z.H.; Rehman, H.M.; Akhtar, T.; Alsamadany, H.; Hamooh, B.T.; Mujtaba, T.; Daur, I.; Al Zahrani, Y.; Alzahrani, H.A.S.; Ali, S.; et al. Humic Substances: Determining Potential Molecular Regulatory Processes in Plants. Front. Plant Sci. 2018, 9, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiavon, M.; Pizzeghello, D.; Muscolo, A.; Nardi, S. High molecular size humic substances enhance phenylpropanoid metabolism in maize (Zea mays L.). J. Chem. Ecol. 2010, 36, 662–669. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 2014, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Zandonadi, D.B.; Santos, M.P.; Caixeta, L.S.; Marinho, E.B.; Peres, L.E.P.; Façanha, A.R. Plant proton pumps as markers of biostimulant action. Sci. Hort. 2016, 73, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Nardi, S.; Ertani, A.; Francioso, O. Soil–root cross--talking: The role of humic substances. J. Plant Nutr. Soil Sci. 2017, 180, 5–13. [Google Scholar] [CrossRef]
- Nunes, R.O.; Domiciano, G.A.; Alves, W.S.; Melo, A.C.A.; Nogueira, F.C.S.; Canellas, L.P.; Olivares, F.L.; Zingali, R.B.; Soares, M.R. Evaluation of the effects of humic acids on maize root architecture by label-free proteomics analysis. Sci Rep. 2019, 9, 12019. [Google Scholar] [CrossRef]
- Muscolo, A.; Sidari, M.; Attinà, E.; Francioso, O.; Tugnoli, V.; Nardi, S. Biological activity of humic substances is related to their chemical structure. Soil Sci. Soc. Am. J. 2007, 71, 75–85. [Google Scholar] [CrossRef]
- Canellas, L.P.; Spaccini, R.; Piccolo, A.; Dobbss, L.B.; Okorokova-Façanha, A.L.; Santos, G.A.; Olivares, F.L.; Façanha, A.R. Relationships between chemical characteristics and root growth promotion of humic acids isolated from Brazilian Oxisols. Soil Sci. 2009, 174, 611–620. [Google Scholar] [CrossRef]
- Chan, P.L.S.; Griffiths, D.A. The vermicomposting of pre-treated pig manure. Biol. Wastes 2001, 24, 57–69. [Google Scholar] [CrossRef]
- Xiang, C.; Zhang, P.; Pan, G. Changes in diversity, protein content, and amino acid composition of earthworms from a paddy soil under different long-term fertilizations in the Tai Lake Region. Acta Ecologica Sinica 2006, 26, 1667–1673. [Google Scholar] [CrossRef]
- Eriksen-Hamel, N.S.; Whalen, J.K. Growth rates of Aporrectodea caliginosa (Oligochaetae: Lumbricidae) as influenced by soil temperature and moisture in disturbed and undisturbed soil columns. Pedobiologia 2006, 50, 207–215. [Google Scholar] [CrossRef]
- Eriksen-Hamel, N.S.; Speratti, A.B.; Whalen, J.K.; Légère, A.; Madramootoo, C.A. Earthworm populations and growth rates related to long-term crop residue and tillage management. Soil Tillage Res. 2008, 104, 311–316. [Google Scholar] [CrossRef]
- Fonte, S.J.; Winsome, T.; Six, J. Earthworm populations in relation to soil organic matter dynamics and management in California tomato cropping systems. Appl. Soil Ecol. 2009, 41, 206–214. [Google Scholar] [CrossRef]
- Simonsen, J.; Posner, J.; Rosemeyer, M.; Baldock, J. Endogeic and anecic earthworm abundance in six Midwestern cropping systems. Appl. Soil Ecol. 2010, 44, 147–155. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Cesco, S.; Varanini, Z.; Pinton, R. Humic substances contribute to plant iron nutrition acting as chelators and biostimulants. Front. Plant Sci. 2019, 10, 675. [Google Scholar] [CrossRef]
- Ferrari, E.; Francioso, O.; Nardi, S.; Saladini, M.; Dal Ferro, N.; Morari, F. DRIFT and HR MAS NMR characterization of humic substances from a soil treated with different organic and mineral fertilizers. J. Mol. Struct. 2011, 998, 216–224. [Google Scholar] [CrossRef]
- Tinti, A.; Tugnoli, V.; Bonora, S.; Francioso, O. Recent applications of vibrational mid-Infrared (IR) spectroscopy for studying soil components: A review. J. Centr. Eur. Agric. 2015, 16, 1–22. [Google Scholar] [CrossRef]
- Carletti, P.; Roldan, M.L.; Francioso, O.; Nardi, S.; Sanchez-Cortes, S. Structural characterization of humic-like substances with conventional and surface-enhanced spectroscopic techniques. J. Mol. Struct. 2010, 982, 169–175. [Google Scholar] [CrossRef]
- Brunow, G. Lignin, Humic Substances and Coal; Hofrichter, M., Steinbüchel, A., Eds.; Wiley-VCH: Weinheim, Germany, 2001; p. 89. [Google Scholar]
- Osono, T.; Takeda, H. Effects of organic chemical quality and mineral nitrogen addition on decomposition of beech leaf litter by Xylaria sp. Eur. J. Soil Biol. 2001, 37, 17–23. [Google Scholar] [CrossRef]
- Thevenot, M.; Dignac, M.F.; Rumpel, C. Fate of lignins in soils: A review. Soil Biol. Biochem. 2010, 42, 1200–1211. [Google Scholar] [CrossRef]
- Audus, L.J. Plant Growth Substances Chemistry and Physiology; L. Hill: London, UK, 1972; Volume 1, pp. 24–65. [Google Scholar]
- Lemtiri, A.; Colinet, G.; Alabi, T.; Cluzeau, D.; Zirbes, L.; Haubruge, E.; Francis, F. Impacts of earthworms on soil components and dynamics. Biotechnol. Agron. Soc. Environ. 2014, 18, 121–133. [Google Scholar]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Edwards, C.A.; Lofty, J.R. The effects of direct drilling and minimal cultivation on earthworm populations. J. Appl. Ecol. 1982, 19, 723–734. [Google Scholar] [CrossRef]
- Nardi, S.; Concheri, G.; Dell’Agnola, G. Biological activity of humus. In Humic Substances in Terrestrial Ecosystems; Piccolo, A., Ed.; Elsevier: Amsterdam, The Netherlands, 1996; pp. 361–406. [Google Scholar]
- Russell, L.; Stokes, A.R.; Macdonald, H.; Muscolo, A.; Nardi, S. Stomatal responses to humic substances and auxin are sensitive to inhibitors of phospholipase A2. Plant Soil 2006, 283, 175–185. [Google Scholar] [CrossRef]
- Pizzeghello, D.; Zanella, A.; Carletti, P.; Nardi, S. Chemical and biochemical characterization of dissolved organic matter from Silver fir and beech forest soils. Chemosphere 2006, 65, 190–200. [Google Scholar] [CrossRef]
- Nardi, S.; Muscolo, A.; Vaccaro, S.; Baiano, S.; Spaccini, R.; Piccolo, A. Relationship between molecular characteristics of soil humic fractions and glycolytic pathway and krebs cycle in maize seedlings. Soil Biol. Biochem. 2007, 39, 3138–3146. [Google Scholar] [CrossRef]
- Ertani, A.; Nardi, S.; Francioso, O.; Pizzeghello, D.; Tinti, A.; Schiavon, M. Metabolite targeted analysis and physiological traits of Zea mays L. In response to application of a leonardite-humate and lignosulfonate-based products for their evaluation as potential biostimulants. Agronomy 2019, 9, 445. [Google Scholar] [CrossRef]
- García, A.C.; de Souza, L.G.; Pereira, M.G.; Castro, R.N.; García-Mina, J.M.; Zonta, E.; Lisboa, F.J.; Berbara, R.L. Structure-property-function relationship in humic substances to explain the biological activity in plants. Sci. Rep. 2016, 6, 20798. [Google Scholar] [CrossRef]
- Ertani, A.; Francioso, O.; Tugnoli, V.; Righi, V.; Nardi, S. Effect of commercial lignosulfonate-humates on Zea mays L. metabolism. J. Agric. Food Chem. 2011, 59, 11940–11948. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, G.; Schiavon, M.; Nardi, S.; Ertani, A.; Celano, G.; Colombo, C.M. Biostimulant potential of humic acids extracted from an amendment obtained via combination of olive mill wastewaters (OMW) and a pre-treated organic material derived from municipal solid waste (MSW). Front. Plant Sci. 2018, 9, 1028. [Google Scholar] [CrossRef] [PubMed]
- Schiavon, M.; Ertani, A.; Nardi, S. Effects of an alfalfa protein hydrolysate on the gene expression and activity of enzymes of the tricarboxylic acid (TCA) cycle and nitrogen metabolism in Zea mays L. J. Agric. Food Chem. 2008, 56, 11800–11808. [Google Scholar] [CrossRef] [PubMed]
- Baglieri, A.; Cadili, V.; Monterumici, C.M.; Gennaria, M.; Tabasso, S.; Montoneri, R.; Nardi, S.; Negre, M. Fertilization of bean plants with tomato plants hydrolysates. Effect on biomass production, chlorophyll content and N assimilation. Sci. Hort. 2014, 176, 194–199. [Google Scholar] [CrossRef]
- Giardini, L.; Giovanardi, R.; Borin, M. Confronto tra quattro rotazioni colturali eseguite per un dodicennio a diversi livelli di concimazione e di irrigazione: Aspetti agronomici ed economici. Rivista di Agronomia 1987, 21, 18–27. [Google Scholar]
- Giardini, L.; Borin, M.; Berti, A. Effetti del letame e del liquame bovino in avvicendamenti colturali a diverso livello di concimazione. Rivista di Agronomia 1999, 33, 118–129. [Google Scholar]
- Stevenson, F.J. Humus Chemistry: Genesis, Composition, Reactions, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1994; p. 496. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis, Part 3; SSSA Book Series No. 5; Madison, WI, USA, 1996; pp. 961–1010. Available online: http://www.scirp.org/(S(351jmbntvnsjt1aadkposzje))/reference/ReferencesPapers.aspx?ReferenceID=2017439 (accessed on 17 October 2019).
- Dell’Agnola, G.; Ferrari, G. Effect of humic acids on anion uptake by excised barley roots. Humus. Planta 1971, 5, 567–569. [Google Scholar]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolysates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Dall’Acqua, S.; Ertani, A.; Pilon-Smits, E.A.H.; Fabrega-Prats, M.; Schiavon, M. Selenium biofortification differentially affects sulfur metabolism and accumulation of phytochemicals in two rocket species (Eruca sativa mill. and Diplotaxis tenuifolia) grown in hydroponics. Plants 2019, 8, 68. [Google Scholar] [CrossRef]
- Welburn, A.R.; Lichtenthaler, H. Formulae and program to determine total carotenoids and chlorophylls A and B of leaf extracts in different solvents. In Advances in Photosynthesis Research. Advances in Agricultural Biotechnology; Sybesma, C., Ed.; Springer: Dordrecht, The Netherlands, 1984; Volume 2. [Google Scholar]
- Lewis, O.A.M.; Watson, E.F.; Hewitt, E.J. Determination of nitrate reductase activity in barley leaves and roots. Ann. Bot. 1982, 49, 31–37. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | NF | FM | M | FMM | ||||
|---|---|---|---|---|---|---|---|---|
| pH | 8.20 | ±1.10a | 8.25 | ±1.02a | 8.31 | ±2.12a | 8.18 | ±1.88a |
| OC % | 0.75 | ±0.03c | 1.59 | ±0.06a | 1.33 | ±0.05b | 1.51 | ±0.10a |
| OM % | 1.29 | ±0.06c | 2.73 | ±0.04a | 2.28 | ±0.06b | 2.60 | ±0.09a |
| N % | 0.07 | ±0.01b | 0.16 | ±0.02a | 0.15 | ±0.04a | 0.18 | ±0.02a |
| C/N | 10.15 | ±1.11a | 9.94 | ±1.04b | 8.86 | ±1.15c | 9.43 | ±1.28c |
| HC % | 0.15 | ±0.02d | 0.53 | ±0.02a | 0.23 | ±0.03c | 0.33 | ±0.02b |
| Treatment | N | P | K | Ca | Fe | Mn | Zn | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % (g/100g d.wt.) | mg/g d.wt. | |||||||||||||
| Contr | 4.71 | ±0.10b | 6.60 | ±0.25b | 20.50 | ±1.03b | 1.10 | ±0.13c | 0.43 | ±0.04e | 0.061 | ±0.003d | 0.19 | ±0.02b |
| NF 15 | 5.01 | ±0.10a | 7.12 | ±0.33b | 21.80 | ±1.12b | 1.26 | ±0.10c | 0.48 | ±0.05e | 0.077 | ±0.004b | 0.22 | ±0.01ab |
| NF 25 | 5.00 | ±0.10a | 8.14 | ±0.33a | 21.39 | ±1.35b | 1.25 | ±0.12c | 0.49 | ±0.03e | 0.079 | ±0.004b | 0,19 | ±0.02b |
| FM 15 | 5.33 | ±0.31a | 8.12 | ±0.23a | 26.12 | ±0.85a | 1.82 | ±0.12a | 0.88 | ±0.01a | 0.102 | ±0.001a | 0.23 | ±0.01a |
| FM 25 | 5.12 | ±0.15a | 7.90 | ±0.21a | 25.33 | ±0.56a | 1.72 | ±0.12ab | 0.81 | ±0.01b | 0.080 | ±0.001b | 0.23 | ±0.03ab |
| M 15 | 4.63 | ±0.20b | 6.95 | ±0.14b | 22.95 | ±0.13b | 1.50 | ±0.15b | 0.66 | ±0.03cd | 0.071 | ±0.005bc | 0.20 | ±0.01b |
| M 25 | 4.50 | ±0.18b | 6.71 | ±0.10b | 22.61 | ±0.14b | 1.40 | ±0.13b | 0.63 | ±0.02d | 0.063 | ±0.002d | 0.19 | ±0.01b |
| FMM 15 | 5.02 | ±0.15a | 7.50 | ±0.35ab | 24.82 | ±1.03a | 1.66 | ±0.18ab | 0.74 | ±0.05bc | 0.080 | ±0.003b | 0.22 | ±0.03ab |
| FMM 25 | 4.90 | ±0.13a | 7.31 | ±0.18b | 24.24 | ±1.10ab | 1.53 | ±0.14ab | 0.70 | ±0.02c | 0.071 | ±0.003c | 0.21 | ±0.02ab |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiavon, M.; Ertani, A.; Francioso, O.; Nardi, S. Manure Fertilization Gives High-Quality Earthworm Coprolites with Positive Effects on Plant Growth and N Metabolism. Agronomy 2019, 9, 659. https://doi.org/10.3390/agronomy9100659
Schiavon M, Ertani A, Francioso O, Nardi S. Manure Fertilization Gives High-Quality Earthworm Coprolites with Positive Effects on Plant Growth and N Metabolism. Agronomy. 2019; 9(10):659. https://doi.org/10.3390/agronomy9100659
Chicago/Turabian StyleSchiavon, Michela, Andrea Ertani, Ornella Francioso, and Serenella Nardi. 2019. "Manure Fertilization Gives High-Quality Earthworm Coprolites with Positive Effects on Plant Growth and N Metabolism" Agronomy 9, no. 10: 659. https://doi.org/10.3390/agronomy9100659
APA StyleSchiavon, M., Ertani, A., Francioso, O., & Nardi, S. (2019). Manure Fertilization Gives High-Quality Earthworm Coprolites with Positive Effects on Plant Growth and N Metabolism. Agronomy, 9(10), 659. https://doi.org/10.3390/agronomy9100659