Barley Varieties Stoneham and Sydney Exhibit Mild Antibiosis and Antixenosis Resistance to the Wheat Curl Mite, Aceria tosichella (Keifer)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Material, A. tosichella Biotypes and Environmental Conditions
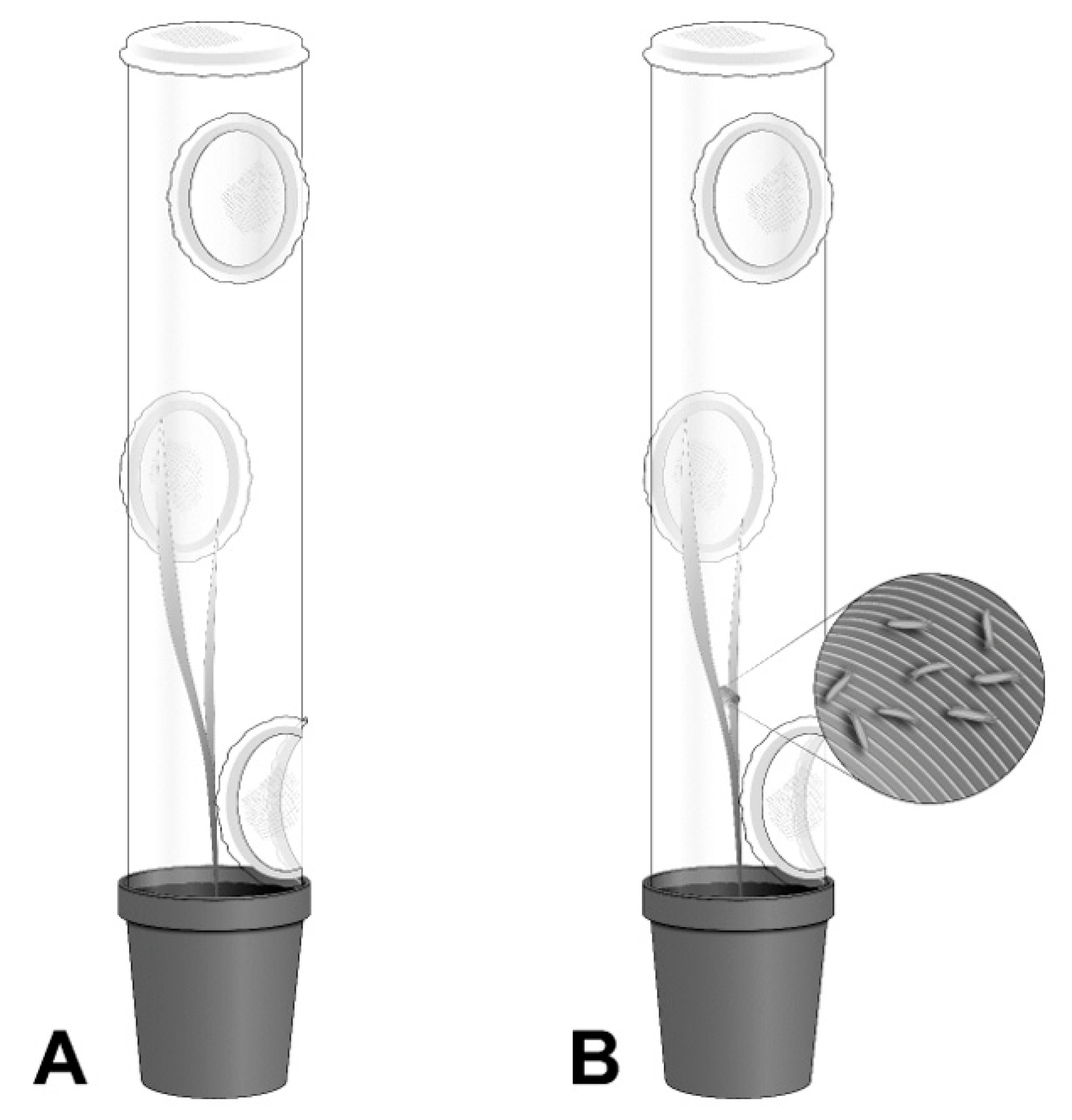
2.2. Non-Choice Experiments
2.3. Choice Experiments
2.4. Statistical Analyses
2.4.1. Non-Choice Experiments
2.4.2. Choice Experiments
3. Results
3.1. Non-Choice Experiments
3.2. Choice Experiments
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slykhuis, J.T. Mite transmission of plant viruses. In Advances in Virus Research; Smith, K.M., Lauffer, M.A., Eds.; Academic Press: Cambridge, MA, USA, 1965; Volume 11, pp. 97–137. [Google Scholar]
- Skoracka, A.; Rector, B.G.; Hein, G.L. The interface between wheat and the wheat curl mite, Aceria tosichella, the primary vector of globally important viral diseases. Front. Plant Sci. 2018, 9, 1098. [Google Scholar] [CrossRef] [PubMed]
- Navia, D.; de Mendonça, R.S.; Skoracka, A.; Szydło, W.; Knihinicki, D.; Hein, G.L.; da Silva Pereira, P.R.V.; Truol, G.; Lau, D. Wheat curl mite, Aceria tosichella, and transmitted viruses: An expanding pest complex affecting cereal crops. Exp. Appl. Acarol. 2013, 59, 95–143. [Google Scholar] [CrossRef] [PubMed]
- Lehnhoff, E.; Miller, Z.; Menalled, F.; Ito, D.; Burrows, M. Wheat and barley susceptibility and tolerance to multiple isolates of Wheat Streak Mosaic Virus. Plant Dis. 2015, 99, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Ito, D.; Miller, Z.; Menalled, F.; Moffet, M.; Burrows, M. Relative susceptibility among alternative host species prevalent in the Great Plains to Wheat Streak Mosaic Virus. Plant Dis. 2012, 96, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Harvey, T.L.; Martin, T.J.; Seifers, D.L. Wheat yield reduction due to wheat curl mite (Acari: Eriophyidae) infestations. J. Agric. Urban Entomol. 2002, 19, 9–13. [Google Scholar]
- Gillespie, R.L.; Roberts, D.E.; Bentley, E.M. Population dynamics and dispersal of wheat curl mites (Acari: Eryophyidae) in North Central Washington. J. Kans. Entomol. Soc. 1997, 70, 361–364. [Google Scholar]
- Carew, M.; Schiffer, M.; Umina, P.; Weeks, A.; Hoffmann, A. Molecular markers indicate that the wheatcurl mite, Aceria tosichella Keifer, may be a species complex in Australia. Bull. Entomol. Res. 2009, 99, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Hein, G.L.; French, R.; Siriwetwiwat, B.; Amrine, J.W. Genetic characterization of North American populations of the wheat curl mite and dry bulb mite. J. Econ. Entomol. 2012, 105, 1801–1808. [Google Scholar] [CrossRef] [PubMed]
- Skoracka, A.; Kuczyński, L.; Szydło, W.; Rector, B. The wheat curl mite Aceria tosichella (Acari: Eriophyoidea) is a complex of cryptic lineages with divergent host ranges: Evidence from molecular and plant bioassay data. Biol. J. Linn. Soc. 2013, 109, 165–180. [Google Scholar] [CrossRef]
- Malik, R.; Smith, C.M.; Brown-Guedira, G.L.; Harvey, T.L.; Gill, B.S. Assessment of Aegilops tauschii for resistance to biotypes of wheat curl mite (Acari: Eriophyidae). J. Econ. Entomol. 2003, 96, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Skoracka, A.; Lewandowski, M.; Rector, B.G.; Szydło, W.; Kuczyński, L. Spatial and host-related variation in prevalence and population density of wheat curl mite (Aceria tosichella) cryptic genotypes in agricultural landscapes. PLoS ONE 2017, 12, e0169874. [Google Scholar] [CrossRef] [PubMed]
- Brey, C.W.; Johnson, G.D.; Blodgett, S.L. Survey of montana grasses for wheat curl mite (Acari: Eriophyidae), the vector of Wheat Streak Mosaic Virus. J. Agric. Entomol. 1998, 15, 173–181. [Google Scholar]
- Ranabhat, N.B.; Seipel, T.; Lehnhoff, E.A.; Miller, Z.J.; Owen, K.E.; Menalled, F.D.; Burrows, M.E. Temperature and alternative hosts influence Aceria tosichella infestation and Wheat Streak Mosaic Virus infection. Plant Dis. 2018, 102, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Tatineni, S.; Hein, G.L. Genetics and mechanisms underlying transmission of Wheat streak mosaic virus by the wheat curl mite. Curr. Opin. Virol. 2018, 33, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Chuang, W.-P.; Rojas, L.M.A.; Khalaf, L.K.; Zhang, G.; Fritz, A.K.; Whitfield, A.E.; Smith, C.M. Wheat genotypes with combined resistance to wheat curl mite, Wheat Streak Mosaic Virus, Wheat Mosaic Virus, and Triticum Mosaic Virus. J. Econ. Entomol. 2017, 110, 711–718. [Google Scholar] [PubMed]
- Khalaf, L.; Chuang, W.-P.; Aguirre-Rojas, L.; Klein, P.; Smith, C.M. Changes in virulence of North American A. tosichella populations to mite resistance genes in wheat. Arthropod. Plant Interact. 2019, 13, 807–818. [Google Scholar] [CrossRef]
- Carver, B.F.; Smith, C.M.; Chuang, W.-P.; Hunger, R.M.; Edwards, J.T.; Yan, L.; Brown-Guedira, G.; Gill, B.S.; Bai, G.; Bowden, R.L. Registration of OK05312, a high-yielding hard winter wheat donor of Cmc4 for wheat curl mite resistance. J. Plant Regist. 2016, 10, 75–79. [Google Scholar] [CrossRef]
- Martin, T.; Harvey, T.; Bender, C.G.; Seifers, S.L.; Hatchett, J.H. Wheat curl mite resistant germplasm. Crop. Sci. 1983, 23, 809. [Google Scholar] [CrossRef]
- Thomas, J.B.; Conner, R.L.; Graf, R.J. Radiant hard red winter wheat. Can. J. Plant Sci. 2012, 92, 169–175. [Google Scholar] [CrossRef]
- Aguirre-Rojas, M.L.; Khalaf, K.L.; Garcés-Carrera, S.; Sinha, K.D.; Chuang, W.-P.; Smith, M.C. Resistance to wheat curl mite in arthropod-resistant rye-wheat translocation lines. Agronomy 2017, 7, 74. [Google Scholar] [CrossRef]
- Mittal, S.; Dahleen, L.S.; Mornhinweg, D. Locations of quantitative trait loci conferring Russian wheat aphid resistance in barley germplasm STARS-9301B. Crop. Sci. 2008, 48, 1452–1458. [Google Scholar] [CrossRef]
- Mornhinweg, D.H.; Porter, D.R.; Webster, J.A. Inheritance of Russian wheat aphid resistance in spring barley. Crop. Sci. 1995, 35, 1368–1371. [Google Scholar] [CrossRef]
- Mornhinweg, D.W.; Bregitzer, P.P.; Porter, D.R.; Peairs, F.B.; Baltensperger, D.D.; Hein, G.L.; Randolph, T.A.; Koch, M.; Walker, T. Registration of ‘Sidney’ spring feed barley resistant to Russian wheat aphid. J. Plant Regist. 2009, 3, 214–218. [Google Scholar] [CrossRef]
- Mittal, S.; Dahleen, L.S.; Mornhinweg, D. Barley germplasm STARS-9577B lacks a Russian wheat aphid resistance allele at a quantitative trait locus present in STARS-9301B. Crop. Sci. 2009, 49, 1999–2004. [Google Scholar] [CrossRef]
- Mornhinweg, D.W.; Porter, D.R.; Webster, J.A. Inheritance of Russian wheat aphid resistance in spring barley germplasm line STARS-9577B. Crop. Sci. 2002, 42, 1891–1893. [Google Scholar] [CrossRef]
- Painter, R.H. The mechanisms of resistance. In Insect Resistance in Crop Plants; The MacMillan Co.: New York, NY, USA, 1951; pp. 23–83. [Google Scholar]
- Smith, C.M.; Clement, S.L. Molecular bases of plant resistance to arthropods. Annu. Rev. Entomol. 2011, 57, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.R.; Saxena, R.C. Technique for locating planthopper (Homoptera: Delphacidae) and leafhopper (Homoptera: Cicadellidae) eggs in rice plants. J. Econ. Entomol. 1986, 79, 271–273. [Google Scholar] [CrossRef] [Green Version]
- Reese, J.C.; Schwenke, J.R.; Lamont, P.S.; Zehr, D. Importance and quantification of plant tolerance in crop pest management programs for aphids: Greenbug resistance in sorghum. J. Agric. Entomol. 1994, 11, 255–270. [Google Scholar]
- Peterson, R.K.D.; Varella, A.C.; Higley, L.G. Tolerance: The forgotten child of plant resistance. PeerJ 2017, 5, e3934. [Google Scholar] [CrossRef] [PubMed]
- Mornhinweg, D.W.; Bregitzer, P.P.; Porter, D.R.; Peairs, F.B.; Baltensperger, D.D.; Hein, G.L.; Randolph, T.A.; Koch, M.; Walker, T. Registration of ‘Stoneham’ spring feed barley resistant to Russian wheat aphid. J. Plant Regist. 2012, 6, 1–5. [Google Scholar] [CrossRef]
- Wesenberg, D.; Burrup, D.; Kolding, M.; Brown, B.; Burnett, C.R. Registration of “Eight-Twelve” barley. Crop. Sci. 1992, 32, 828. [Google Scholar] [CrossRef]
- Malik, R. Molecular Genetic Characterization of Wheat Curl Mite, Aceria Tosichella Keifer (Acari: Eriophyidae), and Wheat Genes Conferring Wheat Curl Mite Resistance. Ph.D. Thesis, Kansas State University, Manhattan, KS, USA, 2001. [Google Scholar]
- Carrera, S.G.; Davis, H.; Aguirre–Rojas, L.; Murugan, M.; Smith, C.M. Multiple categories of resistance to wheat curl mite (Acari: Eriophyidae) expressed in accessions of Aegilops tauschii. J. Econ. Entomol. 2012, 105, 2180–2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugan, M.; Cardona, P.S.; Duraimurugan, P.; Whitfield, A.E.; Schneweis, D.; Smith, C.M. Wheat curl mite resistance: Interactions of mite feeding with Wheat Streak Mosaic Virus infection. J. Econ. Entomol. 2011, 104, 1406–1414. [Google Scholar] [CrossRef] [PubMed]
- Kuczyński, L.; Rector, B.G.; Kiedrowicz, A.; Lewandowski, M.; Szydło, W.; Skoracka, A. Thermal niches of two invasive genotypes of the wheat curl mite Aceria tosichella: Congruence between physiological and geographical distribution data. PLoS ONE 2016, 11, e0154600. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, N.V. Estimate of deviation between empirical distribution functions in two independent samples. Bull. Mosc. Univ. 1939, 2, 3–16. [Google Scholar]
- Levene, H. Robust tests for the equality of variance. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling; Olkin, I., Ghurye, S.G., Hoeffding, W., Madow, W.G., Mann, H.B., Eds.; Stanford University Press: Palo Alto, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Brown, M.B.; Forsythe, A.B. Robust tests for equality of variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- SAS Institute. The GLIMMIX Procedure, SAS/STAT User’s Guide, version 9.2; SAS Institute, Inc.: Cary, NC, USA, 2009. [Google Scholar]
- Stroup, W.W. Generalized Linear Mixed Models: Modern Concepts, Methods and Applications; Chapman & Hall/CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D. SAS System for Mixed Models; SAS Institute, INC: Cary, NC, USA, 1996. [Google Scholar]
- Milliken, G.; Johnson, D. Analysis of Messy Data; Chapman & Hall/CRC Press: Boca Raton, FL, USA, 2009; Volume 1. [Google Scholar]
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2007. [Google Scholar]
- Anderson, G.R.; Papa, D.; Peng, J.; Tahir, M.; Lapitan, N.L. V Genetic mapping of Dn7, a rye gene conferring resistance to the Russian wheat aphid in wheat. Theor. Appl. Genet. 2003, 107, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Brown-Guedira, G.L.; Smith, C.M.; Harvey, T.L.; Gill, B.S. Genetic mapping of wheat curl mite resistance genes Cmc3 and Cmc4 in common wheat. Crop. Sci. 2003, 43, 644–650. [Google Scholar] [CrossRef]
- Thomas, J.B.; Conner, R.L. Resistance to colonization by the wheat curl mite in Aegilops squarrosa and its inheritance after transfer to common wheat. Crop. Sci. 1986, 26, 527–530. [Google Scholar] [CrossRef]
- Marais, G.F.; Horn, M.; Du Torr, F. Intergeneric transfer (rye to wheat) of a gene (s) for Russian wheat aphid resistance. Plant Breed. 1994, 113, 265–271. [Google Scholar] [CrossRef]
- Porter, D.R.; Mornhinweg, D.W.; Webster, J.A. Insect resistance in barley germplasm. In Global Plant Genetic Resources for Insect-Resistant Crops; CRC Press: Boca Raton, FL, USA, 1999; pp. 51–62. [Google Scholar]


{kind=link}
| Mean (Lower, Upper CI) | Mean Percent Proportional Change ± SE | ||||
|---|---|---|---|---|---|
| Plant Variety | Arthropod R Gene(s) | # Mites ° | Tolerance Index ° | Dry Weight | Height |
| OK05312 wheat (R) | Cmc4 | 7.2 (0.9, 58.2) a | 5.9 (0.8, 25.9) | 18.6 ± 14.5 | 24.5 ± 8.7 |
| Eight-twelve barley | None | 40.5 (5.3, 308.2) ab | 0.5 (0.3, 6.1) | 6.9 ± 13.8 | 23.4 ± 8.4 |
| Stoneham barley | Rdn1, Rdn2* | 78.3 (10.3, 593.5) b | 0.9 (0.2, 4.5) | −3.6 ± 13.8 | 7.8 ± 8.4 |
| Sydney barley | Rdn1, Rdn2 | 83.2 (11.0, 630.8) b | 0.5 (0.3, 6.1) | 18.7 ± 13.8 | 7.6 ± 8.4 |
| Ike wheat (S) | None | 155.2 (20.5, 1175.0) b | 4.0 (0.3, 6.1) | 12.5 ± 13.8 | −0.7 ± 0.7 |
| Mean (Lower, Upper CI) | Mean Percent Proportional Change ± SE | ||||
|---|---|---|---|---|---|
| Plant Variety | Arthropod R Gene(s) | # Mites ° | Tolerance Index ° | Dry Weight | Height |
| OK05312 wheat (R) | Cmc4 | 33.4 (9.4, 118.2) a | 5.6 (1.0, 20.8) | 9.1 ± 11.8 | 24.2 ± 4.5 |
| Eight-twelve barley | None | 58.9 (16.7, 207.3) ab | 1.4 (0.4, 4.2) | 15.1 ± 11.8 | 4.6 ± 4.5 |
| Sydney barley | Rdn1, Rdn2 | 79.6 (21.6, 290.0) ab | 0.1 (0.3, 3.4) | 0.2 ± 11.8 | 4.6 ±4.5 |
| Stoneham barley | Rdn1, Rdn2* | 143.1 (37.5, 545.9) ab | 0.5 (0.3, 5.3) | 14.7 ± 11.8 | 23.1 ± 4.5 |
| Ike wheat (S) | None | 216.3 (61.8, 757.6) b | 0.1 (0.3, 3.3) | 19.9 ± 11.8 | 8.1 ± 4.5 |
| Fisher’s Exact Test P-Value for Each Variety Compared to: | ||||
|---|---|---|---|---|
| Plant Variety | Arthropod R Gene(s) | % Folded Plants | Ike (Susceptible Control) | OK05312 (Resistant Control) |
| OK05312 wheat (R) | Cmc4 | 14.3 | 0.0101 * | - |
| Eight-twelve barley | None | 37.5 | 0.1189 ns | 0.2872 ns |
| Stoneham barley | Rdn1, Rdn2* | 62.5 | 0.5692 ns | 0.1189 ns |
| Sydney barley | Rdn1, Rdn2 | 75.0 | 1.0 ns | 0.0406 * |
| Ike wheat (S) | None | 87.5 | - | 0.0101 * |
| Fisher’s Exact Test P-Value for Each Variety Compared to: | ||||
|---|---|---|---|---|
| Plant Variety | Arthropod R Gene(s) | % Folded Plants | Ike (Susceptible Control) | OK05312 (Resistant Control) |
| OK05312 wheat (R) | Cmc4 | 0 | 0.4667 ns | - |
| Eight-twelve barley | None | 0 | 0.4667 ns | 1.0 ns |
| Sydney barley | Rdn1, Rdn2 | 25 | 1.0 ns | 0.4667 ns |
| Ike wheat (S) | None | 25 | - | 0.4667 ns |
| Stoneham barley | Rdn1, Rdn2* | 62.5 | 0.3147 ns | 0.0256 * |
| Plant Variety | Arthropod R Gene(s) | Mean # Mites (Lower, Upper CI) ° |
|---|---|---|
| OK05312 wheat (R) | Cmc4 | 2.2 (1.0, 5.1) a |
| Sydney barley | Rdn1, Rdn2 | 4.8 (2.2, 10.3) ab |
| Eight-twelve barley | None | 9.2 (4.4, 19.5) b |
| Ike wheat (S) | None | 10.1 (4.8, 21.1) b |
| Stoneham barley | Rdn1, Rdn2* | 17.7 (8.6, 36.6) b |
| Plant Variety | Arthropod R Gene(s) | Mean # Mites (Lower, Upper CI) |
|---|---|---|
| OK05312 wheat (R) | Cmc4 | 91.9 (176.5, 368.3) a |
| Eight-twelve barley | None | 176.3 (80.4, 272.2) ab |
| Ike wheat (S) | None | 272.4 (176.5, 368.3) bc |
| Sydney barley | Rdn1, Rdn2 | 331.7 (235.8, 427.6) c |
| Stoneham barley | Rdn1, Rdn2* | 347.8 (251.9, 443.7) c |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguirre-Rojas, L.M.; Khalaf, L.K.; Smith, C.M. Barley Varieties Stoneham and Sydney Exhibit Mild Antibiosis and Antixenosis Resistance to the Wheat Curl Mite, Aceria tosichella (Keifer). Agronomy 2019, 9, 748. https://doi.org/10.3390/agronomy9110748
Aguirre-Rojas LM, Khalaf LK, Smith CM. Barley Varieties Stoneham and Sydney Exhibit Mild Antibiosis and Antixenosis Resistance to the Wheat Curl Mite, Aceria tosichella (Keifer). Agronomy. 2019; 9(11):748. https://doi.org/10.3390/agronomy9110748
Chicago/Turabian StyleAguirre-Rojas, Lina Maria, Luaay Kahtan Khalaf, and Charles Michael Smith. 2019. "Barley Varieties Stoneham and Sydney Exhibit Mild Antibiosis and Antixenosis Resistance to the Wheat Curl Mite, Aceria tosichella (Keifer)" Agronomy 9, no. 11: 748. https://doi.org/10.3390/agronomy9110748
APA StyleAguirre-Rojas, L. M., Khalaf, L. K., & Smith, C. M. (2019). Barley Varieties Stoneham and Sydney Exhibit Mild Antibiosis and Antixenosis Resistance to the Wheat Curl Mite, Aceria tosichella (Keifer). Agronomy, 9(11), 748. https://doi.org/10.3390/agronomy9110748