Spatial and Temporal Distribution of Ecballium elaterium in Almond Orchards


Abstract
:1. Introduction
2. Material and Methods
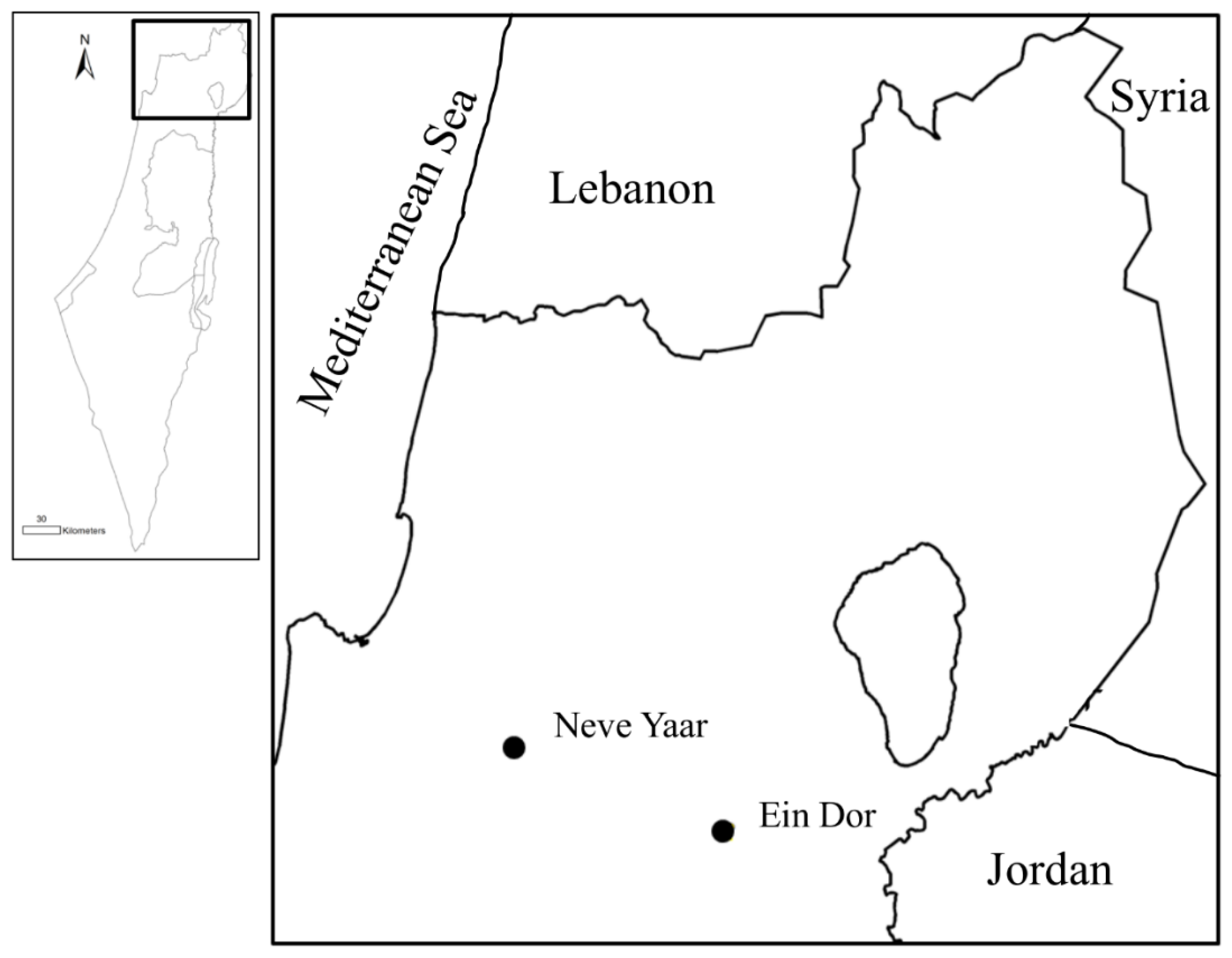
2.1. Study Sites and Data Collection
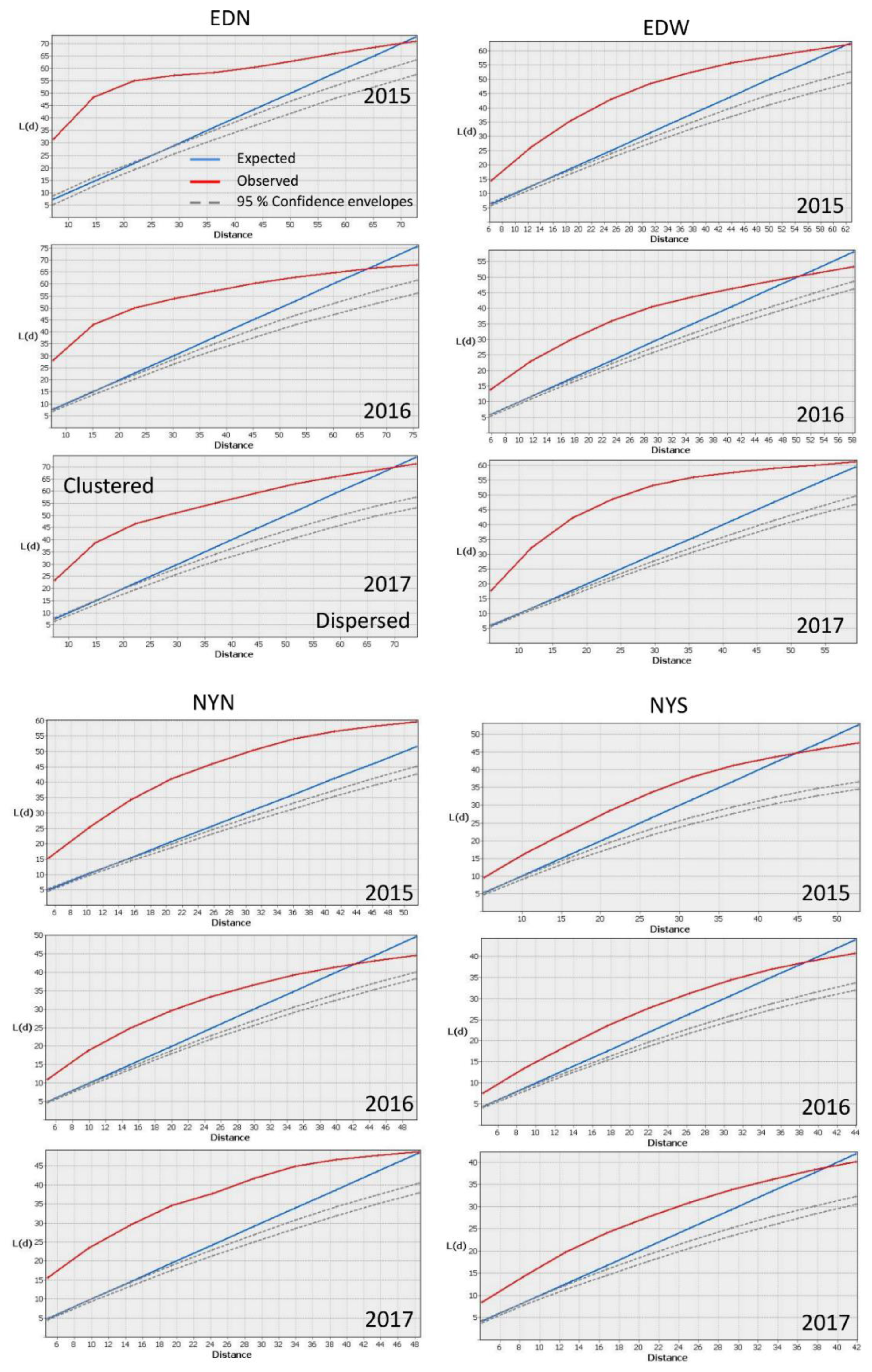
2.2. Spatially Analyzing Patterns
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Steward, B.L.; Tian, L.F. Real-time machine vision weed-sensing. ASAE Pap. 1998. Paper number 983033. [Google Scholar]
- Lopez-Granados, F. Weed detection for site-specific weed management: Mapping and real-time approaches. Weed Res. 2011, 51, 1–11. [Google Scholar] [CrossRef]
- Dieleman, J.A.; Mortensen, D.A.; Cooper, E.M.; Sims, J.T.; Cunningham, S.D.; Huang, J.W.; Berti, W.R. Characterizing the spatial pattern of Abutilon theophrasti seedling patches. Weed Res. 1999, 39, 455–467. [Google Scholar] [CrossRef]
- Heijting, S.; Van Der Werf, W.; Stein, A.; Kropff, M.J. Are weed patches stable in location? Application of an explicitly two-dimensional methodology. Weed Res. 2007, 47, 381–395. [Google Scholar] [CrossRef]
- Andújar, D.; Barroso, J.; Fernández-Quintanilla, C.; Dorado, J. Spatial and temporal dynamics of Sorghum halepense patches in maize crops. Weed Res. 2012, 52, 411–420. [Google Scholar] [CrossRef]
- Calha, I.M.; Sousa, E.; González-Andújar, J.L. Infestation maps and spatial stability of main weed species in maize culture. Planta Daninha 2014, 32, 275–282. [Google Scholar] [CrossRef]
- San Martín, C.; Andújar, D.; Fernández-Quintanilla, C.; Dorado, J. Spatial distribution patterns of weed communities in corn fields of central Spain. Weed Sci. 2015, 63, 936–945. [Google Scholar] [CrossRef]
- Metcalfe, H.; Milne, A.E.; Webster, R.; Lark, R.M.; Murdoch, A.J.; Kanelo, L.; Storkey, J. Defining the habitat niche of Alopecurus myosuroides at the field scale. Weed Res. 2018, 58, 165–176. [Google Scholar] [CrossRef]
- Walter, A.M.; Christensen, S.; Simmelsgaard, S.E. Spatial correlation between weed species densities and soil properties. Weed Res. 2002, 42, 26–38. [Google Scholar] [CrossRef]
- Ballaré, C.L.; Scopel, A.L.; Ghersa, C.M.; Sánchez, R.A. The population ecology of Datura ferox in soybean crops. A simulation approach incorporating seed dispersal. Agric. Ecosyst. Environ. 1987, 19, 177–188. [Google Scholar] [CrossRef]
- Blank, L.; Carmel, Y. Woody vegetation patch types affect herbaceous species richness and composition in a Mediterranean ecosystem. Community Ecol. 2012, 13, 72–81. [Google Scholar] [CrossRef]
- Cardina, J.; Johnson, G.A.; Sparrow, D.H. The nature and consequence of weed spatial distribution. Weed Sci. 1997, 45, 364–373. [Google Scholar] [CrossRef]
- Thill, D.C.; Mallory-Smith, C.A. The nature and consequence of weed spread in cropping systems. Weed Sci. 1997, 45, 337–342. [Google Scholar] [CrossRef]
- Blanco-Moreno, J.M.; Chamorro, L.; Masalles, R.M.; Recasens, J.; Sans, F.X. Spatial distribution of Lolium rigidum seedlings following seed dispersal by combine harvesters. Weed Res. 2004, 44, 375–387. [Google Scholar] [CrossRef]
- Garibay, S.V.; Richner, W.; Stamp, P.; Nakamoto, T.; Yamagishi, J.; Abivardi, C.; Edwards, P.J. Extent and implications of weed spatial variability in arable crop fields. Plant Prod. Sci. 2001, 4, 259–269. [Google Scholar] [CrossRef]
- Gerhards, R.; Christensen, S. Real-time weed detection, decision making and patch spraying in maize, sugarbeet, winter wheat and winter barley. Weed Res. 2003, 43, 385–392. [Google Scholar] [CrossRef]
- Gerhards, R.; Wyse-Pester, D.Y.; Johnson, G.A. Characterizing spatial stability of weed populations using interpolated maps. Weed Sci. 1997, 45, 108–119. [Google Scholar] [CrossRef]
- Wilson, B.J.; Brain, P. Long-term stability of distribution of Alopecurus myosuroides Huds. within cereal fields. Weed Res. 1991, 31, 367–373. [Google Scholar] [CrossRef]
- Wyse-Pester, D.Y.; Mortensen, D.A.; Gotway, C.A. Statistical methods to quantify spatial stability of weed population. Proc. North Cent. Weed Control Conf. 1995, 50, 512. [Google Scholar]
- Johnson, G.A.; Mortensen, D.A.; Gotway, C.A. Spatial and temporal analysis of weed seedling populations using geostatistics. Weed Sci. 1996, 44, 704–710. [Google Scholar] [CrossRef]
- Myers, M.W.; Curran, W.S.; Vangessel, M.J.; Majek, B.A.; Scott, B.A.; Mortensen, D.A.; Calvin, D.D.; Karsten, H.D.; Roth, G.W. The effect of weed density and application timing on weed control and corn grain yield. Weed Technol. 2005, 19, 102–107. [Google Scholar] [CrossRef]
- Lati, R.N.; Filin, S.; Eizenberg, H. Effect of tuber density and trifloxysulfuron application timing on purple nutsedge (Cyperus rotundus) control. Weed Sci. 2012, 60, 494–500. [Google Scholar] [CrossRef]
- Cousens, R. A simple model relating yield loss to weed density. Ann. Appl. Biol. 1985, 107, 239–252. [Google Scholar] [CrossRef]
- Longchamps, L.; Panneton, B.; Reich, R.; Simard, M.-J.; Leroux, G.D. Spatial pattern of weeds based on multispecies infestation maps created by imagery. Weed Sci. 2016, 64, 474–485. [Google Scholar] [CrossRef]
- Colbach, N.; Forcella, F.; Johnson, G.A. Spatial and temporal stability of weed populations over five years. Weed Sci. 2000, 48, 366–377. [Google Scholar] [CrossRef]
- Mueller-Warrant, G.W.; Whittaker, G.W.; Young, W.C. GIS analysis of spatial clustering and temporal change in weeds of grass seed crops. Weed Sci. 2008, 56, 647–669. [Google Scholar] [CrossRef]
- Mitchel, A. The ESRI Guide to GIS Analysis, Volume 2: Spatial Measurements and Statistics; ESRI Press: Redlands, CA, USA, 2005. [Google Scholar]
- Dafni, A.; Benítez, G.C.; Blanché, C.; Rammón-Laca, L.; Petanidou, T.; Aytaç, B.; Horvat, M.; Lucchese, F.; Geva-Kleinberger, A. The etymological, ecological, historical and ethnobotanical roots of the vernacular names of Ecballium elaterium (L.) Rich. (Squirting cucumber). J. Ethnobiol. Tradit. Med. Photon 2013, 119, 515–537. [Google Scholar]
- Lewes, D. Observations on the internal pressure of the ripening fruit of Ecballium elaterium. Kew Bull. 1951, 6, 443–444. [Google Scholar] [CrossRef]
- Beckie, H.J.; Reboud, X. Selecting for weed resistance: Herbicide rotation and mixture. Weed Technol. 2009, 23, 363–370. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef]
- CRDC Integrated weed management Hub. Section 3: Herbicide Resistance. Grains Research and Development Corporation, Australia Government; CRDC Integrated weed management Hub: Canberra, Australia, 2014. [Google Scholar]
- Sezen, U.U.; Chazdon, R.L.; Holsinger, K.E. Proximity is not a proxy for parentage in an animal-dispersed Neotropical canopy palm. Proc. R. Soc. B 2009, 276, 2037–2044. [Google Scholar] [CrossRef] [PubMed]
- Wehenkel, C.; Brazão-Protázio, J.M.; Carrillo-Parra, A.; Martínez-Guerrero, J.H.; Crecente-Campo, F. Spatial distribution patterns in the very rare and species-rich Picea chihuahuana tree community (Mexico). PLoS ONE 2015, 10, e0140442. [Google Scholar]
- Ripley, B.D. Spatial Statistics; John Wiley & Sons: Hoboken, NJ, USA, 2005; Volume 575. [Google Scholar]
- Lynch, H.J.; Moorcroft, P.R. A spatiotemporal Ripley’s K-function to analyze interactions between spruce budworm and fire in British Columbia, Canada. Can. J. For. Res. 2008, 38, 3112–3119. [Google Scholar] [CrossRef]
- Birger, N. Environmental Friendly Management of Ecballium elaterium in Almond Orchards Based on Understanding the Relationship Between Spatial Distribution, Weed Biology and Chemical Control. Master’s Thesis November 2018. [Google Scholar]
- Roham, R.; Pirdashti, H.; Yaghubi, M.; Nematzadeh, G. Spatial distribution of nutsedge (Cyperus spp. L.) seed bank in rice growth cycle using geostatistics. Crop Prot. 2014, 55, 133–141. [Google Scholar] [CrossRef]
- Andújar, D.; Ruiz, D.; Ribeiro, Á.; Fernández-Quintanilla, C.; Dorado, J. Spatial distribution patterns of johnsongrass (Sorghum halepense) in commercial corn fields in Spain. Weed Sci. 2011, 59, 82–89. [Google Scholar] [CrossRef]
- Mitskas, M.B.; Tsolis, C.E.; Eleftherohorinos, I.G.; Damalas, C.A. Interference between corn and johnsongrass (Sorghum halepense) from seed or rhizomes. Weed Sci. 2003, 51, 540–545. [Google Scholar] [CrossRef]
- Barroso, J.; Andújar, D.; San Martín, C.; Fernández-Quintanilla, C.; Dorado, J. Johnsongrass (Sorghum halepense) seed dispersal in corn crops under Mediterranean conditions. Weed Sci. 2012, 60, 34–41. [Google Scholar] [CrossRef]
- Hubbell, S.P.; Foster, R.B.; O’Brien, S.T.; Harms, K.E.; Condit, R.; Wechsler, B.; Wright, S.J.; De Lao, S.L. Light-gap disturbances, recruitment limitation, and tree diversity in a neotropical forest. Science 1999, 283, 554–557. [Google Scholar] [CrossRef] [Green Version]
- Plotkin, J.B.; Chave, J.; Ashton, P.S. Cluster analysis of spatial patterns in Malaysian tree species. Am. Nat. 2002, 160, 629–644. [Google Scholar] [CrossRef]
- Gunatilleke, C.V.S.; Gunatilleke, I.; Esufali, S.; Harms, K.E.; Ashton, P.M.S.; Burslem, D.; Ashton, P.S. Species–habitat associations in a Sri Lankan dipterocarp forest. J. Trop. Ecol. 2006, 22, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Mendelsohn, O.; Dayan, T.; Aidlin-Harari, S.; Silberstein, M.; Orlov, V.; Blank, L. Mediterranean fruit fly subplot hot spots prediction by experts’ experience. J. Appl. Entomol. 2018, 142, 371–379. [Google Scholar] [CrossRef]
- Bar-Massada, A.; Belmaker, J. Non-stationarity in the co-occurrence patterns of species across environmental gradients. J. Ecol. 2017, 105, 391–399. [Google Scholar] [CrossRef]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N. Species associations in a heterogeneous Sri Lankan dipterocarp forest. Am. Nat. 2007, 170, E77–E95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmeyer, H.; Dunker, M.; Stafford, J.V. Variable weed densities and soil properties in a weed mapping concept for patchy weed control. In Proceedings of the Precision agriculture ’99, Part 1; Sheffield Academic Press: Odense Denmark, 1999; pp. 453–462. [Google Scholar]
- Andújar, D.; Rueda-Ayala, V.; Dorado, J.; Gerhards, R.; Fernández-Quintanilla, C. Multivariate Analysis of the Agricultural Management Presence of Sorghum halepense (L.) Pers. Relationships in Maize Crops. Gesunde Pflanz. 2014, 66, 17–22. [Google Scholar]
- Timmermann, C.; Gerhards, R.; Kühbauch, W. The economic impact of site-specific weed control. Precis. Agric. 2003, 4, 249–260. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Name | Abbreviations | Plantation Year | Survey Area (m2) |
|---|---|---|---|
| Ein Dor West | EDW | 2006 | 24,800 |
| Ein Dor North | EDN | 2007 | 32,800 |
| Newe Yaar North | NYN | 2007 | 18,900 |
| Newe Yaar South | NYS | 2007 | 6600 |
| 2015 | 2016 | 2017 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Oct-Dec (2014) | Jan–Mar | Apr–Jun | Oct–Dec (2015) | Jan–Mar | Apr–Jun | Oct–Dec (2016) | Jan–Mar | Apr–Jun | |
| Newe Yaar | |||||||||
| Average Temperature (°C) | 18.2 ± 5.2 | 13.4 ± 5 | 21.5 ± 5.9 | 18.4 ± 6.2 | 14.3 ± 5.2 | 23.0 ± 5.9 | 17.8 ± 6.5 | 12.4 ± 4.7 | 22.0 ± 5.7 |
| Precipitation (mm) | 186.8 | 379.9 | 78.1 | 141.2 | 334.6 | 17 | 312.1 | 179.6 | 23.9 |
| Ein Dor | |||||||||
| Average Temperature (°C) | 18.1 ± 5.4 | 13.3 ± 5.2 | 21.9 ± 6.3 | 18.5 ± 6.3 | 14.2 ± 5.2 | 23.0 ± 5.9 | 18.0 ± 6.9 | 12.4 ± 4.8 | 22.9 ± 6.1 |
| Precipitation (mm) | 178.6 | 297.7 | 68.5 | 122 | 308.8 | 13.3 | 242.1 | 158.8 | 20.5 |
| Plot Name | Number of Plants | Average Distance between Plants (M) | ANN Ratio | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | |
| EDW | 297 | 505 | 526 | 2.89 | 1.09 | 1.37 | 0.63 | 0.33 | 0.41 |
| EDN | 155 | 294 | 191 | 2.42 | 1.36 | 1.87 | 0.29 | 0.27 | 0.33 |
| NYN | 443 | 768 | 467 | 1.47 | 0.72 | 1.04 | 0.4 | 0.34 | 0.37 |
| NYS | 280 | 515 | 452 | 1.62 | 0.87 | 1.22 | 0.56 | 0.42 | 0.49 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blank, L.; Birger, N.; Eizenberg, H. Spatial and Temporal Distribution of Ecballium elaterium in Almond Orchards. Agronomy 2019, 9, 751. https://doi.org/10.3390/agronomy9110751
Blank L, Birger N, Eizenberg H. Spatial and Temporal Distribution of Ecballium elaterium in Almond Orchards. Agronomy. 2019; 9(11):751. https://doi.org/10.3390/agronomy9110751
Chicago/Turabian StyleBlank, Lior, Nitzan Birger, and Hanan Eizenberg. 2019. "Spatial and Temporal Distribution of Ecballium elaterium in Almond Orchards" Agronomy 9, no. 11: 751. https://doi.org/10.3390/agronomy9110751
APA StyleBlank, L., Birger, N., & Eizenberg, H. (2019). Spatial and Temporal Distribution of Ecballium elaterium in Almond Orchards. Agronomy, 9(11), 751. https://doi.org/10.3390/agronomy9110751