Somatic Embryogenesis and Plant Regeneration from Cotyledon and Hypocotyl Explants of Fagopyrum esculentum Moench lpls Mutant

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Callus Induction
2.2. Callus Maintenance and Embryogenic Callus Induction
2.3. Embryogenic Callus Maintenance, Somatic Embryos Germination and Plant Regeneration
2.4. Histological Analysis
2.5. Observations and Statistical Analyses
3. Results
3.1. Callus Induction
3.2. Callus Maintenance and Embryogenic Callus Induction
3.3. Embryogenic Callus Maintenance, Somatic Embryos Germination and Plant Regeneration
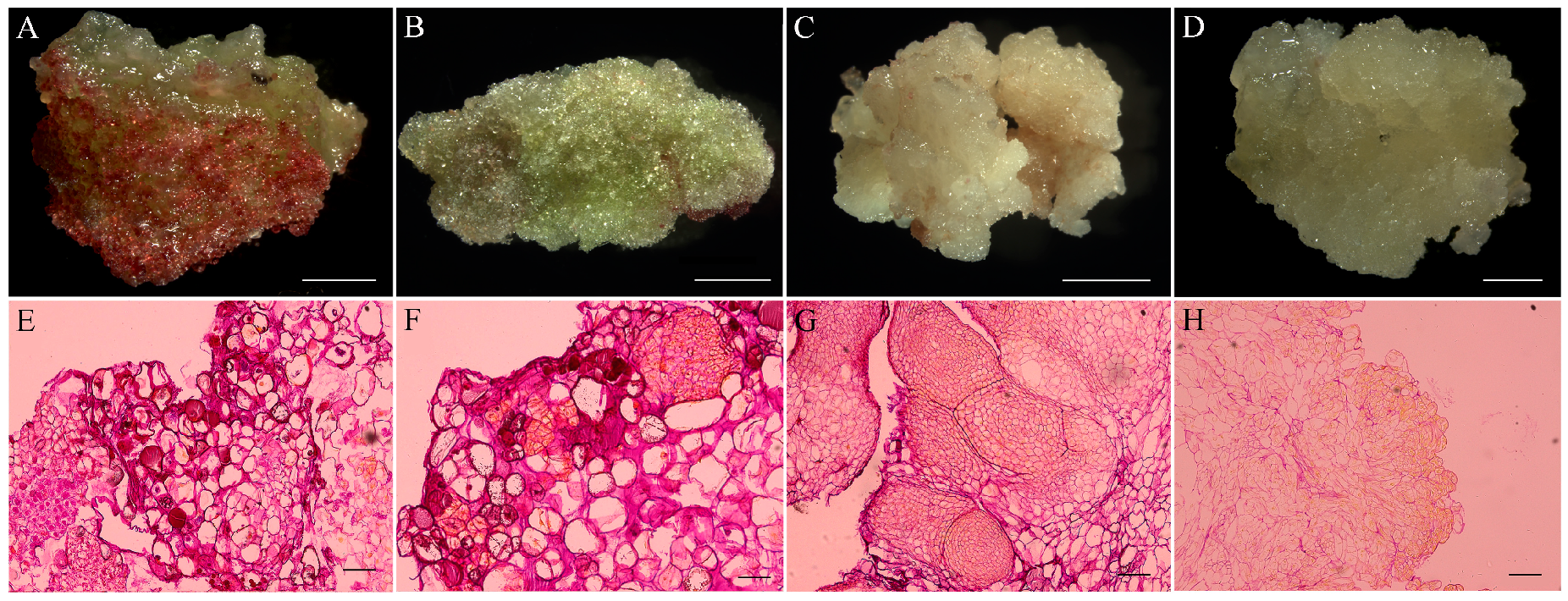
3.4. Somatic Embryogenesis of Fagopyrum esculentum and Histological Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2,4-D | 2,4-Dichlorophenoxyacetic acid |
| 6-BA | 6-Benzylaminopurine |
| BAP | 6-Benzylaminopurine |
| ABA | abscisic acid |
| FAA | Formalin acetic acid alcohol |
| IAA | Indole-3-acetic acid |
| KT | Kinetin |
| MS | Murashige and Skoog |
| NAA | α-Naphthalene acetic acid |
| TDZ | N-Phenyl-N′-1,2,3-thiadiazol-5-ylurea |
References
- Mizuno, N.; Yasui, Y. Gene flow signature in the S-allele region of cultivated buckwheat. BMC Plant Biol. 2019, 19, 125. [Google Scholar] [CrossRef] [PubMed]
- Quinet, M.; Cawoy, V.; Lefèvre, I.; Van Miegroet, F.; Jacquemart, A.L.; Kinet, J.M. Inflorescence structure and control of flowering time and duration by light in buckwheat (Fagopyrum esculentum Moench). J. Exp. Bot. 2004, 55, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Cawoy, V.; Kinet, J.M.; Jacquemart, A.L. Morphology of nectaries and biology of nectar production in the distylous species Fagopyrum esculentum. Ann. Bot. 2008, 102, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Yabe, S.; Hara, T.; Ueno, M.; Enoki, H.; Kimura, T.; Nishimura, S.; Yasui, Y.; Ohsawa, R.; Iwata, H. Rapid genotyping with DNA micro-arrays for high-density linkage mapping and QTL mapping in common buckwheat (Fagopyrum esculentum Moench.). Breed. Sci. 2014, 64, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Oomah, B.D.; Mazza, G. Flavonoids and antioxidative activities in buckwheat. J. Agric. Food chem. 1996, 44, 1746–1750. [Google Scholar] [CrossRef]
- Giménez-Bastida, A.; Zieliński, H. Buckwheat as a Functional Food and Its Effects on Health. J. Agric. Food Chem. 2015, 63, 7896–7913. [Google Scholar] [CrossRef] [PubMed]
- Kreft, M. Buckwheat phenolic metabolites in health and disease. Nutr. Res. Rev. 2016, 29, 30–39. [Google Scholar] [CrossRef]
- Li, L.Y.; Fang, Z.W.; Li, X.P.; Liu, Z.X. Isolation and Characterization of the C-class MADS-box Gene from the Distylous Pseudo-cereal Fagopyrum esculentum. J. Plant Biol. 2017, 60, 189–198. [Google Scholar] [CrossRef]
- Cawoy, V.; Lutts, S.; Ledent, J.F.; Kinet, J.M. Resource availability regulates reproductive meristem activity, development of reproductive structures and seed set in buckwheat (Fagopyrum esculentum). Physiol. Plant. 2007, 131, 341–353. [Google Scholar] [CrossRef]
- Kadokura, S.; Sugimoto, K.; Tarr, P.; Suzuki, T.; Matsunaga, S. Characterization of somatic embryogenesis initiated from the Arabidopsis shoot apex. Dev. Biol. 2018, 442, 13–27. [Google Scholar] [CrossRef]
- Saeed, W.; Naseem, S.; Gohar, D.; Ali, Z. Efficient and reproducible somatic embryogenesis and micropropagation in tomato via novel structures—Rhizoid Tubers. PLoS ONE 2019, 14, e0215929. [Google Scholar] [CrossRef] [PubMed]
- Ming, N.G.J.; Binte Mostafiz, S.; Johon, N.S.; Abdullah Zulkifli, N.S.; Wagiran, A. Combination of Plant Growth Regulators, Maltose, and Partial Desiccation Treatment Enhance Somatic Embryogenesis in Selected Malaysian Rice Cultivar. Plants (Basel) 2019, 8, 144. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; La Rota, M.; Hoerster, G.; Hastings, C.; Wang, N.; Chamberlin, M.; Wu, E.; Jones, T.; Gordon-Kamm, W. Rapid genotype “independent” Zea mays L. (maize) transformation via direct somatic embryogenesis. In Vitro Cell Dev. Biol. Plant 2018, 54, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Nešković, M.; Vujičić, R.; Budimir, S. Somatic embryogenesis and bud formation from immature embryos of buckwheat (Fagopyrum esculentum Moench). Plant Cell Rep. 1987, 6, 423–426. [Google Scholar] [CrossRef]
- Kwon, S.J.; Han, M.H.; Huh, Y.S.; Roy, S.K.; Lee, C.W.; Woo, S.H. Plantlet regeneration via somatic embryogenesis from hypocotyls of common buckwheat (Fagopyrum esculentum Moench). Korean J. Crop Sci. 2013, 58, 331–335. [Google Scholar] [CrossRef]
- Saraswat, R.; Kumar, M. Plant regeneration in buckwheat (Fagopyrum esculentum Moench) via somatic embryogenesis and induction of meristemoids in abnormal embryos. Plant Tissue Cult. & Biotech. 2019, 29, 33–47. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Zeng, S.J.; Cheng, S.J.; Zhang, J.L.; Zhao, F.P. Embryo Culture and Propagation of Dendrobium in Vitro. Acta Hortic. Sin. 1998, 25, 75–80. [Google Scholar]
- Jiang, J.L. Study on the Effects of Several Additives on the PLB Proliferation and Tube Seedlings Growth of Oncidium ’Gower Ramsey’. Subtrop. Plant Sci. 2012, 41, 8–14. [Google Scholar] [CrossRef]
- Liu, Z.X.; Xiong, H.Y.; Li, L.Y.; Fei, Y.J. Functional Conservation of an AGAMOUS Orthologous Gene Controlling Reproductive Organ Development in the Gymnosperm species Taxus chinensis var. Mairei. J. Plant Biol. 2018, 61, 50–59. [Google Scholar] [CrossRef]
- Venkataiah, P.; Bhanuprakash, P.; Suman Kalyan, S.; Subhash, K. Somatic embryogenesis and plant regeneration of Capsicum baccatum L. J. Genet. Eng. Biotechnol. 2016, 14, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, P.E.V.; Benítez-García, I.; Peralta, A.L.L.; Paredes-López, O.; Del Villar-Martínez, A.A. Somatic embryogenesis from leaf explants of Tagetes erecta L. Plant Biotechnol. (Tokyo) 2017, 34, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Rathore, M.S.; Paliwal, N.; Anand, K.G.V.; Agarwal, P.K. Somatic embryogenesis and in vitro plantlet regeneration in Salicornia brachiata Roxb. Plant Cell Tissue Organ Cult. 2015, 120, 355–360. [Google Scholar] [CrossRef]
- Dey, T.; Bhattacharya, S.; Ghosh, P.D. Somatic embryogenesis from rhizome explants of Cymbopogon winterianus. Biol. Plant. 2010, 54, 325–328. [Google Scholar] [CrossRef]
- Gumerova, E.A.; Galeeva, E.I.; Chuyenkova, S.A.; Rumyantseva, N.I. Somatic Embryogenesis and Bud Formation on Cultured Fagopyrum esculentum Hypocotyls. Rus. J. Plant Physiol. 2003, 50, 640–645. [Google Scholar] [CrossRef]
- Woo, S.H.; Nair, A.; Adachi, T.; Campbell, C.G. Plant regeneration from cotyledon tissues of common buckwheat (Fagopyrum esculentum Moench). In Vitro Cell. Dev. Biol.-Plant. 2000, 36, 358–361. [Google Scholar] [CrossRef]
- Klčová, L.; Gubišová, M. Evaluation of Different Approaches to Buckwheat (Fagopyrum esculentum Moench) Micropropagation. Czech J. Genet. Plant Breed. 2008, 44, 66–72. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, Y.K.; Uddin, M.R.; Park, N. An efficient protocol for shoot organogenesis and plant regeneration of buckwheat (Fagopyrum esculentum Moench). Rom. Biotechnol. Lett. 2009, 14, 4524–4529. [Google Scholar]
- Zhang, Q.; Chen, J.; Henny, R.J. Direct somatic embryogenesis and plant regeneration from leaf, petiole, and stem explants of Golden Pothos. Plant Cell Rep. 2005, 23, 587–595. [Google Scholar] [CrossRef]
- Liu, C.; Xia, X.; Yin, W.; Huang, L.; Zhou, J. Shoot regeneration and somatic embryogenesis from needles of redwood (Sequoia sempervirens (D.Don.) Endl.). Plant Cell Rep. 2006, 25, 621–628. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fei, Y.; Wang, L.-X.; Fang, Z.-W.; Liu, Z.-X. Somatic Embryogenesis and Plant Regeneration from Cotyledon and Hypocotyl Explants of Fagopyrum esculentum Moench lpls Mutant. Agronomy 2019, 9, 768. https://doi.org/10.3390/agronomy9110768
Fei Y, Wang L-X, Fang Z-W, Liu Z-X. Somatic Embryogenesis and Plant Regeneration from Cotyledon and Hypocotyl Explants of Fagopyrum esculentum Moench lpls Mutant. Agronomy. 2019; 9(11):768. https://doi.org/10.3390/agronomy9110768
Chicago/Turabian StyleFei, Yue, Lan-Xiang Wang, Zheng-Wu Fang, and Zhi-Xiong Liu. 2019. "Somatic Embryogenesis and Plant Regeneration from Cotyledon and Hypocotyl Explants of Fagopyrum esculentum Moench lpls Mutant" Agronomy 9, no. 11: 768. https://doi.org/10.3390/agronomy9110768
APA StyleFei, Y., Wang, L. -X., Fang, Z. -W., & Liu, Z. -X. (2019). Somatic Embryogenesis and Plant Regeneration from Cotyledon and Hypocotyl Explants of Fagopyrum esculentum Moench lpls Mutant. Agronomy, 9(11), 768. https://doi.org/10.3390/agronomy9110768