Stick around: Cell–Cell Adhesion Molecules during Neocortical Development


Abstract
:1. Introduction
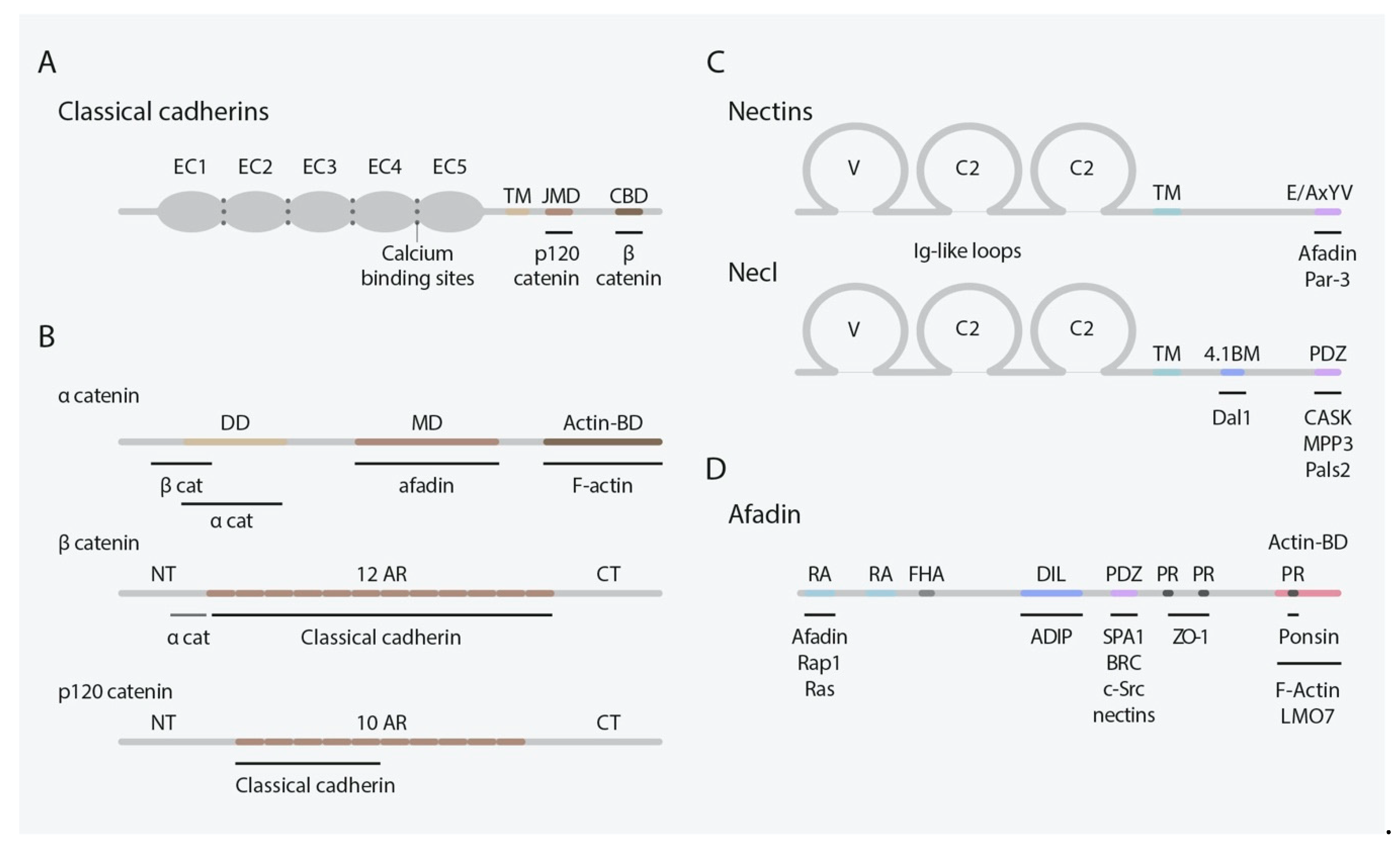
2. Classical Cadherins
3. Nectins and Nectin-Like Molecules
4. Cadherin and Nectin Roles during Mammalian Neocorticogenesis
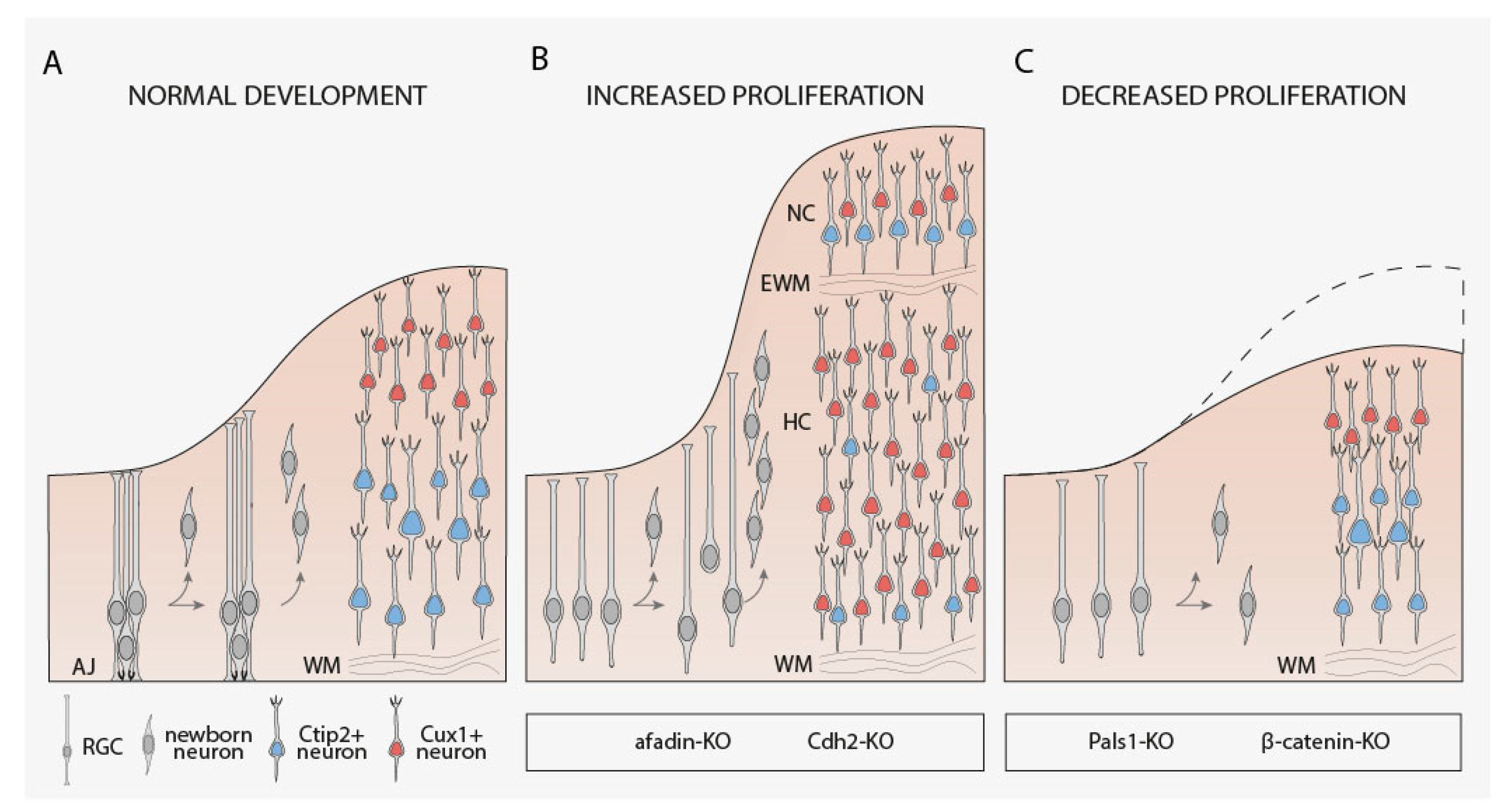
4.1. RGCs Maintenance, Proliferation, and Fate Determination
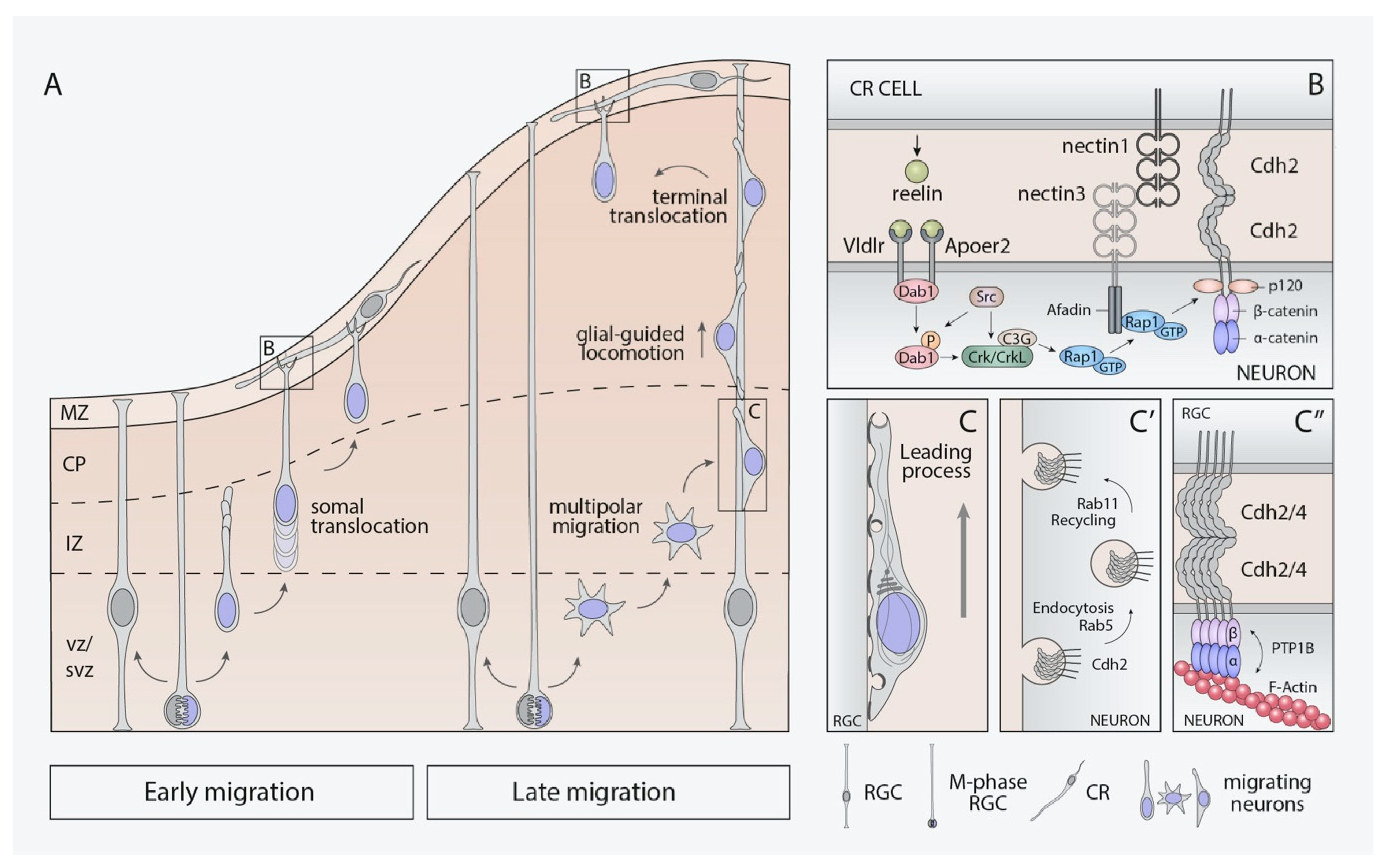
4.2. Neuronal Migration
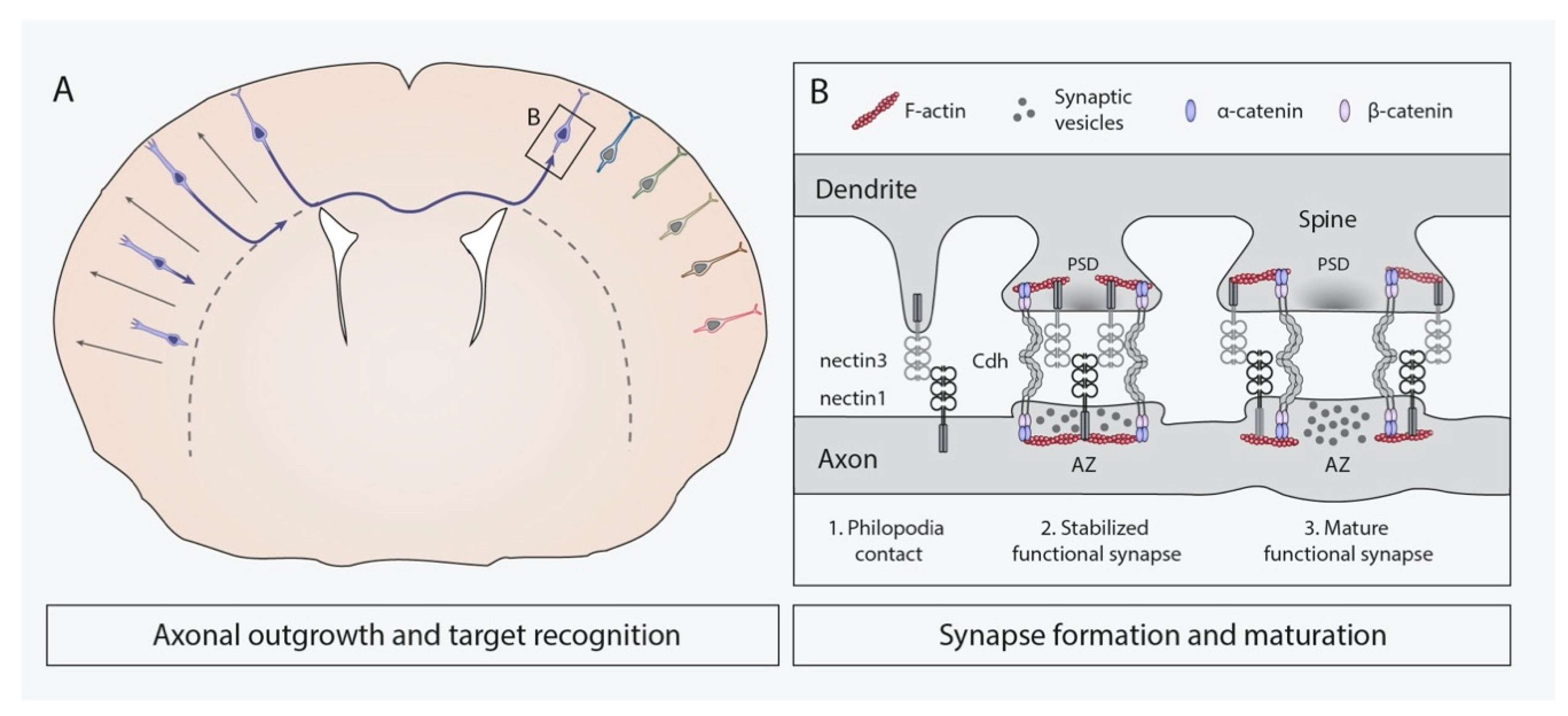
4.3. Axonal Outgrowth and Target Recognition
4.4. Synapse Formation and Remodeling
5. Relevance of Classical Cadherins, Nectins, and Necls in Human Neurodevelopmental Disorders
6. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shapiro, L.; Love, J.; Colman, D.R. Adhesion molecules in the nervous system: Structural insights into function and diversity. Neuroscience 2007, 30, 451–474. [Google Scholar] [CrossRef] [PubMed]
- Rutishauser, U.; Thiery, J.P.; Brackenbury, R.; Sela, B.A.; Edelman, G.M. Mechanisms of adhesion among cells from neural tissues of the chick embryo. Proc. Natl. Acad. Sci. USA 1976, 73, 577–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco, S.J.; Müller, U. Shaping our minds: Stem and progenitor cell diversity in the mammalian neocortex. Neuron 2013, 77, 19–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodato, S.; Arlotta, P. Generating neuronal diversity in the mammalian cerebral cortex. Annu. Rev. Cell Dev. Biol. 2014, 31, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, S.; Takeichi, M. Cadherins in brain morphogenesis and wiring. Physiol. Rev. 2012, 92, 597–634. [Google Scholar] [CrossRef]
- Mandai, K.; Rikitake, Y.; Mori, M.; Takai, Y. Chapter six nectins and nectin-like molecules in development and disease. Curr. Top. Dev. Biol. 2015, 112, 197–231. [Google Scholar] [CrossRef]
- Friedman, L.G.; Benson, D.L.; Huntley, G.W. Chapter thirteen cadherin-based transsynaptic networks in establishing and modifying neural connectivity. Curr. Top. Dev. Biol. 2015, 112, 415–465. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Garay, I. Molecular mechanisms of cadherin function during cortical migration. Front. Cell Dev. Biol. 2020, 8, 588152. [Google Scholar] [CrossRef]
- Hippenmeyer, S. Cellular and molecular control of neuronal migration. Adv. Exp. Med. Biol. 2013, 800, 1–24. [Google Scholar] [CrossRef]
- Walmod, P.S. Introduction. Cell adhesion molecules: Implications in neurological diseases. Adv. Neurobiol. 2014, 8, xvii–xxi. [Google Scholar]
- Sakurai, T. The role of cell adhesion molecules in brain wiring and neuropsychiatric disorders. Mol. Cell Neurosci. 2017, 81, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Dulabon, L.; Olson, E.C.; Taglienti, M.G.; Eisenhuth, S.; McGrath, B.; Walsh, C.A.; Kreidberg, J.A.; Anton, E.S. Reelin binds alpha3beta1 integrin and inhibits neuronal migration. Neuron 2000, 27, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Geiger, B.; Bershadsky, A.; Pankov, R.; Yamada, K.M. Transmembrane crosstalk between the extracellular matrix and the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2001, 2, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Solecki, D.J. Sticky situations: Recent advances in control of cell adhesion during neuronal migration. Curr. Opin. Neurobiol. 2012, 22, 791–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerrisk, M.E.; Cingolani, L.A.; Koleske, A.J. Chapter 5 ECM receptors in neuronal structure, synaptic plasticity, and behavior. Prog. Brain Res. 2014, 214, 101–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Togashi, H.; Sakisaka, T.; Takai, Y. Cell adhesion molecules in the central nervous system. Cell Adhes. Migr. 2009, 3, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sytnyk, V.; Leshchyns’ka, I.; Schachner, M. Neural cell adhesion molecules of the immunoglobulin superfamily regulate synapse formation, maintenance, and function. Trends Neurosci. 2017, 40, 295–308. [Google Scholar] [CrossRef]
- Hatta, K.; Takeichi, M. Expression of N-cadherin adhesion molecules associated with early morphogenetic events in chick development. Nature 1986, 320, 447–449. [Google Scholar] [CrossRef]
- Martinez-Garay, I.; Gil-Sanz, C.; Franco, S.J.; Espinosa, A.; Molnár, Z.; Mueller, U. Cadherin 2/4 signaling via PTP1B and catenins is crucial for nucleokinesis during radial neuronal migration in the neocortex. Development 2016, 143, 2121–2134. [Google Scholar] [CrossRef] [Green Version]
- Schwabe, T.; Gontang, A.C.; Clandinin, T.R. More than just glue: The diverse roles of cell adhesion molecules in the drosophila nervous system. Cell Adhes. Migr. 2009, 3, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Shan, W.-S.; Tanaka, H.; Phillips, G.R.; Arndt, K.; Yoshida, M.; Colman, D.R.; Shapiro, L. Functional cis-heterodimers of N- and R-cadherins. J. Cell Biol. 2000, 148, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Gil-Sanz, C.; Franco, S.J.; Martinez-Garay, I.; Espinosa, A.; Harkins-Perry, S.; Müller, U. Cajal–retzius cells instruct neuronal migration by coincidence signaling between secreted and contact-dependent guidance cues. Neuron 2013, 79, 461–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.E.; de Wit, J.; Ghosh, A. Molecular mechanisms of synaptic specificity in developing neural circuits. Neuron 2010, 68, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulson, A.F.; Prasad, M.S.; Thuringer, A.H.; Manzerra, P. Regulation of cadherin expression in nervous system development. Cell Adhes. Migr. 2013, 8, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Missaire, M.; Hindges, R. The role of cell adhesion molecules in visual circuit formation: From neurite outgrowth to maps and synaptic specificity. Dev. Neurobiol. 2015, 75, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Takeichi, M. Cadherins: A molecular family important in selective cell-cell adhesion. Annu. Rev. Biochem. 1990, 59, 237–252. [Google Scholar] [CrossRef]
- Halbleib, J.M.; Nelson, W.J. Cadherins in development: Cell adhesion, sorting, and tissue morphogenesis. Gene Dev. 2006, 20, 3199–3214. [Google Scholar] [CrossRef] [Green Version]
- Gul, I.S.; Hulpiau, P.; Saeys, Y.; van Roy, F. Evolution and diversity of cadherins and catenins. Exp. Cell Res. 2017, 358, 3–9. [Google Scholar] [CrossRef]
- Brasch, J.; Harrison, O.J.; Honig, B.; Shapiro, L. Thinking outside the cell: How cadherins drive adhesion. Trends Cell Biol. 2012, 22, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Tepass, U.; Truong, K.; Godt, D.; Ikura, M.; Peifer, M. Cadherins in embryonic and neural morphogenesis. Nat. Rev. Mol. Cell Biol. 2000, 1, 91–100. [Google Scholar] [CrossRef]
- Tanabe, K.; Takeichi, M.; Nakagawa, S. Identification of a nonchordate-type classic cadherin in vertebrates: Chicken Hz-cadherin is expressed in horizontal cells of the neural retina and contains a nonchordate-specific domain complex. Dev. Dynam. 2004, 229, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.C.; Takeichi, M. Cadherins in neuronal morphogenesis and function. Dev. Growth Differ. 2008, 50, S119–S130. [Google Scholar] [CrossRef] [PubMed]
- McCrea, P.D.; Maher, M.T.; Gottardi, C.J. Chapter five nuclear signaling from cadherin adhesion complexes. Curr. Top. Dev. Biol. 2015, 112, 129–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, W.; Takeichi, M. Adherens junction: Molecular architecture and regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a002899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Kojima, S.; Borisy, G.G.; Green, K.J. P120 catenin associates with kinesin and facilitates the transport of cadherin–catenin complexes to intercellular junctions. J. Cell Biol. 2003, 163, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.B.; Carnahan, R.H. Regulation of cadherin stability and turnover by P120ctn: Implications in disease and cancer. Semin. Cell Dev. Biol. 2004, 15, 657–663. [Google Scholar] [CrossRef]
- McCrea, P.D.; Gu, D. The catenin family at a glance. J. Cell Sci. 2010, 123, 637–642. [Google Scholar] [CrossRef] [Green Version]
- Heuberger, J.; Birchmeier, W. Interplay of cadherin-mediated cell adhesion and canonical wnt signaling. Cold Spring Harb. Perspect. Biol. 2010, 2, a002915. [Google Scholar] [CrossRef]
- Aken, E.V.; Papeleu, P.; Potter, P.D.; Laey, J.J.D.; Mareel, M. Cadherin expression in the eye. Bull. Soc. Belg. D Ophtalmol. 2000, 278, 55–59. [Google Scholar]
- Bekirov, I.H.; Needleman, L.A.; Zhang, W.; Benson, D.L. Identification and localization of multiple classic cadherins in developing rat limbic system. Neuroscience 2002, 115, 213–227. [Google Scholar] [CrossRef]
- Price, S.R.; Garcia, N.V.D.M.; Ranscht, B.; Jessell, T.M. Regulation of motor neuron pool sorting by differential expression of type II cadherins. Cell 2002, 109, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Takai, Y.; Nakanishi, H. Nectin and afadin: Novel organizers of intercellular junctions. J. Cell Sci. 2003, 116, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takai, Y.; Ikeda, W.; Ogita, H.; Rikitake, Y. The immunoglobulin-like cell adhesion molecule nectin and its associated protein afadin. Annu. Rev. Cell Dev. Biol. 2008, 24, 309–342. [Google Scholar] [CrossRef] [PubMed]
- Mandai, K.; Nakanishi, H.; Satoh, A.; Obaishi, H.; Wada, M.; Nishioka, H.; Itoh, M.; Mizoguchi, A.; Aoki, T.; Fujimoto, T.; et al. Afadin: A novel actin filament-binding protein in with one PDZ domain localized at cadherin-based cell-to-cell adherens junction. J. Cell Biol. 1997, 139, 1061. [Google Scholar] [CrossRef]
- Takahashi, K.; Nakanishi, H.; Miyahara, M.; Mandai, K.; Satoh, K.; Satoh, A.; Nishioka, H.; Aoki, J.; Nomoto, A.; Mizoguchi, A.; et al. Nectin/PRR: An immunoglobulin-like cell adhesion molecule recruited to cadherin-based adherens junctions through interaction with afadin, a PDZ domain-containing protein. J. Cell Biol. 1999, 145, 539–549. [Google Scholar] [CrossRef]
- Satoh-Horikawa, K.; Nakanishi, H.; Takahashi, K.; Miyahara, M.; Nishimura, M.; Tachibana, K.; Mizoguchi, A.; Takai, Y. Nectin-3, a new member of immunoglobulin-like cell adhesion molecules that shows homophilic and heterophilic cell-cell adhesion activities. J. Biol. Chem. 2000, 275, 10291–10299. [Google Scholar] [CrossRef] [Green Version]
- Reymond, N.; Fabre, S.; Lecocq, E.; Adelaïde, J.; Dubreuil, P.; Lopez, M. Nectin4/PRR4, a new afadin-associated member of the nectin family that trans-interacts with nectin1/PRR1 through V domain interaction. J. Biol. Chem. 2001, 276, 43205–43215. [Google Scholar] [CrossRef] [Green Version]
- Saito, S.; Matsushima, M.; Shirahama, S.; Minaguchi, T.; Kanamori, Y.; Minami, M.; Nakamura, Y. Complete genomic structure, DNA polymorphisms, and alternative splicing of the Human AF -6 gene. DNA Res. 1998, 5, 115–120. [Google Scholar] [CrossRef]
- Ohno, S. Intercellular junctions and cellular polarity: The PAR–APKC complex, a conserved core cassette playing fundamental roles in cell polarity. Curr. Opin. Cell Biol. 2001, 13, 641–648. [Google Scholar] [CrossRef]
- Takekuni, K.; Ikeda, W.; Fujito, T.; Morimoto, K.; Takeuchi, M.; Monden, M.; Takai, Y. Direct binding of cell polarity protein PAR-3 to cell-cell adhesion molecule nectin at neuroepithelial cells of developing mouse. J. Biol. Chem. 2003, 278, 5497–5500. [Google Scholar] [CrossRef] [Green Version]
- Narita, H.; Yamamoto, Y.; Suzuki, M.; Miyazaki, N.; Yoshida, A.; Kawai, K.; Iwasaki, K.; Nakagawa, A.; Takai, Y.; Sakisaka, T. Crystal structure of the cis-dimer of nectin-1 implications for the architecture of cell-cell junctions. J. Biol. Chem. 2011, 286, 12659–12669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, W.; Kakunaga, S.; Itoh, S.; Shingai, T.; Takekuni, K.; Satoh, K.; Inoue, Y.; Hamaguchi, A.; Morimoto, K.; Takeuchi, M.; et al. Tage4/Nectin-like molecule-5 heterophilically trans-interacts with cell adhesion molecule nectin-3 and enhances cell migration. J. Biol. Chem. 2003, 278, 28167–28172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizutani, K.; Takai, Y. Nectin spot: A novel type of nectin-mediated cell adhesion apparatus. Biochem. J. 2016, 473, 2691–2715. [Google Scholar] [CrossRef] [PubMed]
- Takai, Y.; Irie, K.; Shimizu, K.; Sakisaka, T.; Ikeda, W. Nectins and nectin-like molecules: Roles in cell adhesion, migration, and polarization. Cancer Sci. 2003, 94, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Galibert, L.; Diemer, G.S.; Liu, Z.; Johnson, R.S.; Smith, J.L.; Walzer, T.; Comeau, M.R.; Rauch, C.T.; Wolfson, M.F.; Sorensen, R.A.; et al. Nectin-like protein 2 defines a subset of T-cell zone dendritic cells and is a ligand for class-I-restricted T-cell-associated molecule. J. Biol. Chem. 2005, 280, 21955–21964. [Google Scholar] [CrossRef] [Green Version]
- Kakunaga, S.; Ikeda, W.; Itoh, S.; Deguchi-Tawarada, M.; Ohtsuka, T.; Mizoguchi, A.; Takai, Y. Nectin-like molecule-1/TSLL1/SynCAM3: A neural tissue-specific immunoglobulin-like cell-cell adhesion molecule localizing at non-junctional contact sites of presynaptic nerve terminals, axons and glia cell processes. J. Cell Sci. 2005, 118, 1267–1277. [Google Scholar] [CrossRef] [Green Version]
- Maurel, P.; Einheber, S.; Galinska, J.; Thaker, P.; Lam, I.; Rubin, M.B.; Scherer, S.S.; Murakami, Y.; Gutmann, D.H.; Salzer, J.L. Nectin-like proteins mediate axon–schwann cell interactions along the internode and are essential for myelination. J. Cell Biol. 2007, 178, 861–874. [Google Scholar] [CrossRef]
- Niederkofler, V.; Baeriswyl, T.; Ott, R.; Stoeckli, E.T. Nectin-like molecules/SynCAMs are required for post-crossing commissural axon guidance. Development 2010, 137, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, I.; Adamsky, K.; Eshed, Y.; Milo, R.; Sabanay, H.; Sarig-Nadir, O.; Horresh, I.; Scherer, S.S.; Rasband, M.N.; Peles, E. A central role for Necl4 (SynCAM4) in Schwann cell–axon interaction and myelination. Nat. Neurosci. 2007, 10, 861–869. [Google Scholar] [CrossRef]
- Pellissier, F.; Gerber, A.; Bauer, C.; Ballivet, M.; Ossipow, V. The adhesion molecule Necl-3/SynCAM-2 localizes to myelinated axons, binds to oligodendrocytes and promotes cell adhesion. BMC Neurosci. 2007, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Honda, T.; Sakisaka, T.; Yamada, T.; Kumazawa, N.; Hoshino, T.; Kajita, M.; Kayahara, T.; Ishizaki, H.; Tanaka-Okamoto, M.; Mizoguchi, A.; et al. Involvement of nectins in the formation of puncta adherentia junctions and the mossy fiber trajectory in the mouse hippocampus. Mol. Cell Neurosci. 2006, 31, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, W.; Nakanishi, H.; Miyoshi, J.; Mandai, K.; Ishizaki, H.; Tanaka, M.; Togawa, A.; Takahashi, K.; Nishioka, H.; Yoshida, H.; et al. Afadin: A key molecule essential for structural organization of cell-cell junctions of polarized epithelia during embryogenesis. J. Cell Biol. 1999, 146, 1117–1132. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Fujita, N.; Yamada, A.; Ooshio, T.; Okamoto, R.; Irie, K.; Takai, Y. Regulation of the assembly and adhesion activity of E-cadherin by nectin and afadin for the formation of adherens junctions in madin-darby canine kidney cells. J. Biol. Chem. 2006, 281, 5288–5299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götz, M.; Huttner, W.B. The cell biology of neurogenesis. Nat. Rev. Mol. Cell Biol. 2005, 6, 777–788. [Google Scholar] [CrossRef]
- Taverna, E.; Götz, M.; Huttner, W.B. The cell biology of neurogenesis: Toward an understanding of the development and evolution of the neocortex. Annu. Rev. Cell Dev. Biol. 2014, 30, 1–38. [Google Scholar] [CrossRef]
- Gelman, D.M.; Marín, O. Generation of interneuron diversity in the mouse cerebral cortex. Eur. J. Neurosci. 2010, 31, 2136–2141. [Google Scholar] [CrossRef]
- Greig, L.C.; Woodworth, M.B.; Galazo, M.J.; Padmanabhan, H.; Macklis, J.D. Molecular logic of neocortical projection neuron specification, development and diversity. Nat. Rev. Neurosci. 2013, 14, 755–769. [Google Scholar] [CrossRef] [Green Version]
- Sultan, K.T.; Brown, K.N.; Shi, S.-H. Production and organization of neocortical interneurons. Front. Cell Neurosci. 2013, 7, 221. [Google Scholar] [CrossRef] [Green Version]
- Miyata, T.; Kawaguchi, A.; Saito, K.; Kawano, M.; Muto, T.; Ogawa, M. Asymmetric production of surface-dividing and non-surface-dividing cortical progenitor cells. Development 2004, 131, 3133–3145. [Google Scholar] [CrossRef] [Green Version]
- Noctor, S.C.; Martínez-Cerdeño, V.; Ivic, L.; Kriegstein, A.R. Cortical neurons arise in symmetric and asymmetric division zones and migrate through specific phases. Nat. Neurosci. 2004, 7, 136–144. [Google Scholar] [CrossRef]
- Gal, J.S.; Morozov, Y.M.; Ayoub, A.E.; Chatterjee, M.; Rakic, P.; Haydar, T.F. Molecular and morphological heterogeneity of neural precursors in the mouse neocortical proliferative zones. J. Neurosci. 2006, 26, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Cerdeño, V.; Cunningham, C.L.; Camacho, J.; Antczak, J.L.; Prakash, A.N.; Cziep, M.E.; Walker, A.I.; Noctor, S.C. Comparative analysis of the subventricular zone in rat, ferret and macaque: Evidence for an outer subventricular zone in rodents. PLoS ONE 2012, 7, e30178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fietz, S.A.; Kelava, I.; Vogt, J.; Wilsch-Bräuninger, M.; Stenzel, D.; Fish, J.L.; Corbeil, D.; Riehn, A.; Distler, W.; Nitsch, R.; et al. OSVZ progenitors of human and ferret neocortex are epithelial-like and expand by integrin signaling. Nat. Neurosci. 2010, 13, 690–699. [Google Scholar] [CrossRef] [PubMed]
- Hansen, D.V.; Lui, J.H.; Parker, P.R.L.; Kriegstein, A.R. Neurogenic radial glia in the outer subventricular zone of human neocortex. Nature 2010, 464, 554–561. [Google Scholar] [CrossRef]
- Reillo, I.; de Romero, C.J.; García-Cabezas, M.Á.; Borrell, V. A role for intermediate radial glia in the tangential expansion of the mammalian cerebral cortex. Cereb. Cortex 2011, 21, 1674–1694. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tsai, J.-W.; LaMonica, B.; Kriegstein, A.R. A new subtype of progenitor cell in the mouse embryonic neocortex. Nat. Neurosci. 2011, 14, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Vaid, S.; Camp, J.G.; Hersemann, L.; Oegema, C.E.; Heninger, A.-K.; Winkler, S.; Brandl, H.; Sarov, M.; Treutlein, B.; Huttner, W.B.; et al. A novel population of Hopx-dependent basal radial glial cells in the developing mouse neocortex. Development 2018, 145, dev.169276. [Google Scholar] [CrossRef] [Green Version]
- Harris, T.J.C.; Peifer, M. APKC controls microtubule organization to balance adherens junction symmetry and planar polarity during development. Dev. Cell 2007, 12, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Kadowaki, M.; Nakamura, S.; Machon, O.; Krauss, S.; Radice, G.L.; Takeichi, M. N-cadherin mediates cortical organization in the mouse brain. Dev. Biol. 2007, 304, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Gil-Sanz, C.; Landeira, B.; Ramos, C.; Costa, M.R.; Müller, U. Proliferative defects and formation of a double cortex in mice lacking Mltt4 and Cdh2 in the dorsal telencephalon. J. Neurosci. 2014, 34, 10475–10487. [Google Scholar] [CrossRef] [Green Version]
- Cappello, S.; Böhringer, C.R.J.; Bergami, M.; Conzelmann, K.-K.; Ghanem, A.; Tomassy, G.S.; Arlotta, P.; Mainardi, M.; Allegra, M.; Caleo, M.; et al. A radial glia-specific role of RhoA in double cortex formation. Neuron 2012, 73, 911–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machon, O.; van den Bout, C.J.; Backman, M.; Kemler, R.; Krauss, S. Role of beta-catenin in the developing cortical and hippocampal neuroepithelium. Neuroscience 2003, 122, 129–143. [Google Scholar] [CrossRef]
- Lien, W.-H.; Klezovitch, O.; Fernandez, T.E.; Delrow, J.; Vasioukhin, V. AE-catenin controls cerebral cortical size by regulating the hedgehog signaling pathway. Science 2006, 311, 1609–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Maruo, T.; Majima, T.; Ishizaki, H.; Tanaka-Okamoto, M.; Miyoshi, J.; Mandai, K.; Takai, Y. Genetic deletion of afadin causes hydrocephalus by destruction of adherens junctions in radial glial and ependymal cells in the midbrain. PLoS ONE 2013, 8, e80356. [Google Scholar] [CrossRef]
- Bian, W.-J.; Miao, W.-Y.; He, S.-J.; Qiu, Z.; Yu, X. Coordinated spine pruning and maturation mediated by inter-spine competition for cadherin/catenin complexes. Cell 2015, 162, 808–822. [Google Scholar] [CrossRef] [Green Version]
- Li, M.-Y.; Miao, W.-Y.; Wu, Q.-Z.; He, S.-J.; Yan, G.; Yang, Y.; Liu, J.-J.; Taketo, M.M.; Yu, X. A critical role of presynaptic cadherin/catenin/P140Cap complexes in stabilizing spines and functional synapses in the neocortex. Neuron 2017, 94, 1155–1172. [Google Scholar] [CrossRef] [Green Version]
- Ribic, A.; Crair, M.C.; Biederer, T. Synapse-selective control of cortical maturation and plasticity by parvalbumin-autonomous action of SynCAM 1. Cell Rep. 2019, 26, 381–393. [Google Scholar] [CrossRef] [Green Version]
- van den Bout, C.J.; Machon, O.; Røsok, Ø.; Backman, M.; Krauss, S. The mouse enhancer element D6 directs cre recombinase activity in the neocortex and the hippocampus. Mech. Develop. 2002, 110, 179–182. [Google Scholar] [CrossRef]
- Kim, S.; Lehtinen, M.K.; Sessa, A.; Zappaterra, M.W.; Cho, S.-H.; Gonzalez, D.; Boggan, B.; Austin, C.A.; Wijnholds, J.; Gambello, M.J.; et al. The apical complex couples cell fate and cell survival to cerebral cortical development. Neuron 2010, 66, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Yabut, O.R.; Pleasure, S.J. Sonic hedgehog signaling rises to the surface: Emerging roles in neocortical development. Adv. Neurol. 2018, 3, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Hou, S.; Han, Y.-G. Hedgehog signaling promotes basal progenitor expansion and the growth and folding of the neocortex. Nat. Neurosci. 2016, 19, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, N.; Tanaka, S.; Horiike, T.; Shinmyo, Y.; Kawasaki, H. A discrete subtype of neural progenitor crucial for cortical folding in the gyrencephalic mammalian brain. Elife 2020, 9, e54873. [Google Scholar] [CrossRef] [PubMed]
- Lui, J.H.; Hansen, D.V.; Kriegstein, A.R. Development and evolution of the human neocortex. Cell 2011, 146, 18–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehay, C.; Kennedy, H.; Kosik, K.S. The outer subventricular zone and primate-specific cortical complexification. Neuron 2015, 85, 683–694. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Martínez, M.Á.; Romero, C.D.J.; Fernández, V.; Cárdenas, A.; Götz, M.; Borrell, V. A restricted period for formation of outer subventricular zone defined by Cdh1 and Trnp1 levels. Nat. Commun. 2016, 7, 11812. [Google Scholar] [CrossRef]
- Namba, T.; Huttner, W.B. Neural progenitor cells and their role in the development and evolutionary expansion of the neocortex. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e256. [Google Scholar] [CrossRef]
- Fang, W.-Q.; Chen, W.-W.; Jiang, L.; Liu, K.; Yung, W.-H.; Fu, A.K.Y.; Ip, N.Y. Overproduction of upper-layer neurons in the neocortex leads to autism-like features in mice. Cell Rep. 2014, 9, 1635–1643. [Google Scholar] [CrossRef] [Green Version]
- Guerrini, R.; Parrini, E. Neuronal migration disorders. Neurobiol. Dis. 2010, 38, 154–166. [Google Scholar] [CrossRef]
- Lodato, S.; Shetty, A.S.; Arlotta, P. Cerebral cortex assembly: Generating and reprogramming projection neuron diversity. Trends Neurosci. 2014, 38, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Marín, O.; Rubenstein, J.L.R. Cell migration in the forebrain. Annu. Rev. Neurosci. 2003, 26, 441–483. [Google Scholar] [CrossRef]
- Anderson, S.A.; Marín, O.; Horn, C.; Jennings, K.; Rubenstein, J.L. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development 2001, 128, 353–363. [Google Scholar] [PubMed]
- Batista-Brito, R.; Fishell, G. Chapter 3 the developmental integration of cortical interneurons into a functional network. Curr. Top. Dev. Biol. 2009, 87, 81–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griveau, A.; Borello, U.; Causeret, F.; Tissir, F.; Boggetto, N.; Karaz, S.; Pierani, A. A novel role for Dbx1-derived cajal–retzius cells in early regionalization of the cerebral cortical neuroepithelium. PLoS Biol. 2010, 8, e1000440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Assimacopoulos, S.; Jones, K.R.; Grove, E.A. Massive loss of cajal–retzius cells does not disrupt neocortical layer order. Development 2006, 133, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frotscher, M. Cajal—retzius cells, reelin, and the formation of layers. Curr. Opin. Neurobiol. 1998, 8, 570–575. [Google Scholar] [CrossRef]
- Barber, M.; Pierani, A. Tangential migration of glutamatergic neurons and cortical patterning during development: Lessons from cajal–retzius cells. Dev. Neurobiol. 2016, 76, 847–881. [Google Scholar] [CrossRef] [Green Version]
- Nadarajah, B.; Brunstrom, J.E.; Grutzendler, J.; Wong, R.O.L.; Pearlman, A.L. Two modes of radial migration in early development of the cerebral cortex. Nat. Neurosci. 2001, 4, 143–150. [Google Scholar] [CrossRef]
- Molyneaux, B.J.; Arlotta, P.; Menezes, J.R.L.; Macklis, J.D. Neuronal subtype specification in the cerebral cortex. Nat. Rev. Neurosci. 2007, 8, 427–437. [Google Scholar] [CrossRef]
- Tabata, H.; Nakajima, K. Multipolar migration: The third mode of radial neuronal migration in the developing cerebral cortex. J. Neurosci. 2003, 23, 9996–10001. [Google Scholar]
- Rakic, P. Mode of cell migration to the superficial layers of fetal monkey neocortex. J. Comp. Neurol. 1972, 145, 61–83. [Google Scholar] [CrossRef]
- D’Arcangelo, G.; Miao, G.G.; Chen, S.-C.; Scares, H.D.; Morgan, J.I.; Curran, T. A protein related to extracellular matrix proteins deleted in the mouse mutant reeler. Nature 1995, 374, 719–723. [Google Scholar] [CrossRef]
- Alcántara, S.; Ruiz, M.; D’Arcangelo, G.; Ezan, F.; de Lecea, L.; Curran, T.; Sotelo, C.; Soriano, E. Regional and cellular patterns of reelin mRNA expression in the forebrain of the developing and adult mouse. J. Neurosci. 1998, 18, 7779–7799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, M.; Miyata, T.; Nakajima, K.; Yagyu, K.; Seike, M.; Ikenaka, K.; Yamamoto, H.; Mikoshiba, K. The reeler gene-associated antigen on cajal–retzius neurons is a crucial molecule for laminar organization of cortical neurons. Neuron 1995, 14, 899–912. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S. Two new mutants, ‘trembler’ and ‘reeler’, with neurological actions in the house mouse (mus musculus L.). J. Genet. 1951, 50, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Rice, D.S.; Curran, T. Role of the reelin signaling pathway in central nervous system development. Annu. Rev. Neurosci. 2001, 24, 1005–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luque, J.M.; Morante-Oria, J.; Fairén, A. Localization of ApoER2, VLDLR and Dab1 in radial glia: Groundwork for a new model of reelin action during cortical development. Dev. Brain Res. 2003, 140, 195–203. [Google Scholar] [CrossRef]
- Huang, Y.; Magdaleno, S.; Hopkins, R.; Slaughter, C.; Curran, T.; Keshvara, L. Tyrosine phosphorylated disabled 1 recruits crk family adapter proteins. Biochem. Bioph. Res. Commun. 2004, 318, 204–212. [Google Scholar] [CrossRef]
- Trommsdorff, M.; Gotthardt, M.; Hiesberger, T.; Shelton, J.; Stockinger, W.; Nimpf, J.; Hammer, R.E.; Richardson, J.A.; Herz, J. Reeler/disabled-like disruption of neuronal migration in knockout mice lacking the VLDL receptor and ApoE receptor 2. Cell 1999, 97, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, M.; Rice, D.S.; D’Arcangelo, G.; Yoneshima, H.; Nakajima, K.; Mikoshiba, K.; Howell, B.W.; Cooper, J.A.; Goldowitz, D.; Curran, T. Scrambler and yotari disrupt the disabled gene and produce a reeler -like phenotype in mice. Nature 1997, 389, 730–733. [Google Scholar] [CrossRef]
- Arnaud, L.; Ballif, B.A.; Förster, E.; Cooper, J.A. Fyn tyrosine kinase is a critical regulator of disabled-1 during brain development. Curr. Biol. 2003, 13, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Howell, B.W.; Lanier, L.M.; Frank, R.; Gertler, F.B.; Cooper, J.A. The disabled 1 phosphotyrosine-binding domain binds to the internalization signals of transmembrane glycoproteins and to phospholipids. Mol. Cell. Biol. 1999, 19, 5179–5188. [Google Scholar] [CrossRef] [Green Version]
- Bock, H.H.; Jossin, Y.; Liu, P.; Förster, E.; May, P.; Goffinet, A.M.; Herz, J. Phosphatidylinositol 3-Kinase interacts with the adaptor protein Dab1 in response to reelin signaling and is required for normal cortical lamination. J. Biol. Chem. 2003, 278, 38772–38779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballif, B.A.; Arnaud, L.; Arthur, W.T.; Guris, D.; Imamoto, A.; Cooper, J.A. Activation of a Dab1/CrkL/C3G/Rap1 pathway in reelin-stimulated neurons. Curr. Biol. 2004, 14, 606–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jossin, Y.; Cooper, J.A. Reelin, Rap1 and N-cadherin orient the migration of multipolar neurons in the developing neocortex. Nat. Neurosci. 2011, 14, 697–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, A.P.; Polleux, F. Establishment of axon-dendrite polarity in developing neurons. Annu. Rev. Neurosci. 2009, 32, 347–381. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Funahashi, Y.; Watanabe, T.; Takano, T.; Nakamuta, S.; Namba, T.; Kaibuchi, K. Radial glial cell–neuron interaction directs axon formation at the opposite side of the neuron from the contact site. J. Neurosci. 2015, 35, 14517–14532. [Google Scholar] [CrossRef] [Green Version]
- Franco, S.J.; Martinez-Garay, I.; Gil-Sanz, C.; Harkins-Perry, S.R.; Müller, U. Reelin regulates cadherin function via Dab1/Rap1 to control neuronal migration and lamination in the neocortex. Neuron 2011, 69, 482–497. [Google Scholar] [CrossRef] [Green Version]
- Kawauchi, T.; Sekine, K.; Shikanai, M.; Chihama, K.; Tomita, K.; Kubo, K.; Nakajima, K.; Nabeshima, Y.; Hoshino, M. Rab GTPases-dependent endocytic pathways regulate neuronal migration and maturation through N-cadherin trafficking. Neuron 2010, 67, 588–602. [Google Scholar] [CrossRef] [Green Version]
- Inoue, S.; Hayashi, K.; Fujita, K.; Tagawa, K.; Okazawa, H.; Kubo, K.; Nakajima, K. Drebrin-like (Dbnl) controls neuronal migration via regulating N-cadherin expression in the developing cerebral cortex. J. Neurosci. 2018, 39, 1618–1634. [Google Scholar] [CrossRef] [Green Version]
- Hara, Y.; Fukaya, M.; Hayashi, K.; Kawauchi, T.; Nakajima, K.; Sakagami, H. ADP ribosylation factor 6 regulates neuronal migration in the developing cerebral cortex through FIP3/arfophilin-1-dependent endosomal trafficking of N-cadherin. ENeuro 2016, 3. [Google Scholar] [CrossRef]
- Luccardini, C.; Hennekinne, L.; Viou, L.; Yanagida, M.; Murakami, F.; Kessaris, N.; Ma, X.; Adelstein, R.S.; Mège, R.-M.; Métin, C. N-cadherin sustains motility and polarity of future cortical interneurons during tangential migration. J. Neurosci. 2013, 33, 18149–18160. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.L.; Rakic, P.; Goldman-Rakic, P.S. Early phenotype expression of cortical neurons: Evidence that a subclass of migrating neurons have callosal axons. Proc. Natl. Acad. Sci. USA 1991, 88, 1354–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keynes, R.; Cook, G.M.W. Axon guidance molecules. Cell 1995, 83, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Stoeckli, E.T. Understanding axon guidance: Are we nearly there yet? Development 2018, 145, dev151415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jontes, J.D. The cadherin superfamily in neural circuit assembly. Cold Spring Harb. Perspect. Biol. 2017, 10, a029306. [Google Scholar] [CrossRef] [PubMed]
- Ranscht, B. Cadherins: Molecular codes for axon guidance and synapse formation. Int. J. Dev. Neurosci. 2000, 18, 643–651. [Google Scholar] [CrossRef]
- Okabe, N.; Shimizu, K.; Ozaki-Kuroda, K.; Nakanishi, H.; Morimoto, K.; Takeuchi, M.; Katsumaru, H.; Murakami, F.; Takai, Y. Contacts between the commissural axons and the floor plate cells are mediated by nectins. Dev. Biol. 2004, 273, 244–256. [Google Scholar] [CrossRef] [Green Version]
- Redies, C. Cadherins and the formation of neural circuitry in the vertebrate CNS. Cell Tissue Res. 1997, 290, 405–413. [Google Scholar] [CrossRef]
- Huntley, G.W.; Benson, D.L. Neural (N)-cadherin at developing thalamocortical synapses provides an adhesion mechanism for the formation of somatopically organized connections. J. Comp. Neurol. 1999, 407, 453–471. [Google Scholar] [CrossRef]
- Gil, O.D.; Needleman, L.; Huntley, G.W. Developmental patterns of cadherin expression and localization in relation to compartmentalized thalamocortical terminations in rat barrel cortex. J. Comp. Neurol. 2002, 453, 372–388. [Google Scholar] [CrossRef]
- Poskanzer, K.; Needleman, L.A.; Bozdagi, O.; Huntley, G.W. N-cadherin regulates ingrowth and laminar targeting of thalamocortical axons. J. Neurosci. 2003, 23, 2294–2305. [Google Scholar] [CrossRef] [Green Version]
- Wakimoto, M.; Sehara, K.; Ebisu, H.; Hoshiba, Y.; Tsunoda, S.; Ichikawa, Y.; Kawasaki, H. Classic cadherins mediate selective intracortical circuit formation in the mouse neocortex. Cereb. Cortex 2015, 25, 3535–3546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Accogli, A.; Calabretta, S.; St-Onge, J.; Boudrahem-Addour, N.; Dionne-Laporte, A.; Joset, P.; Azzarello-Burri, S.; Rauch, A.; Krier, J.; Fieg, E.; et al. De novo pathogenic variants in N-cadherin cause a syndromic neurodevelopmental disorder with corpus collosum, axon, cardiac, ocular, and genital defects. Am. J. Hum. Genet. 2019, 105, 854–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippova, M.; Joshi, M.B.; Kyriakakis, E.; Pfaff, D.; Erne, P.; Resink, T.J. A guide and guard: The many faces of T-cadherin. Cell Signal. 2009, 21, 1035–1044. [Google Scholar] [CrossRef] [PubMed]
- Arlotta, P.; Molyneaux, B.J.; Chen, J.; Inoue, J.; Kominami, R.; Macklis, J.D. Neuronal subtype-specific genes that control corticospinal motor neuron development in vivo. Neuron 2005, 45, 207–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayano, Y.; Zhao, H.; Kobayashi, H.; Takeuchi, K.; Norioka, S.; Yamamoto, N. The role of T-cadherin in axonal pathway formation in neocortical circuits. Development 2014, 141, 4784–4793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molyneaux, B.J.; Arlotta, P.; Fame, R.M.; MacDonald, J.L.; MacQuarrie, K.L.; Macklis, J.D. Novel subtype-specific genes identify distinct subpopulations of callosal projection neurons. J. Neurosci. 2009, 29, 12343–12354. [Google Scholar] [CrossRef] [PubMed]
- Krishna-K, K.; Hertel, N.; Redies, C. Cadherin expression in the somatosensory cortex: Evidence for a combinatorial molecular code at the single-cell level. Neuroscience 2011, 175, 37–48. [Google Scholar] [CrossRef]
- Nuernberger, M.; Weth, F.; Redies, C. Layer-specific expression of multiple cadherins in the developing visual cortex (V1) of the ferret. Cereb. Cortex 2009, 19, 388–401. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.T.; Lim, K.; Giuliano, R.E.; Federoff, H.J. Temporal and spatial localization of nectin1 and lafadin during synaptogenesis in hippocampal neurons. J. Comp. Neurol. 2008, 507, 1228–1244. [Google Scholar] [CrossRef]
- Mizoguchi, A.; Nakanishi, H.; Kimura, K.; Matsubara, K.; Ozaki-Kuroda, K.; Katata, T.; Honda, T.; Kiyohara, Y.; Heo, K.; Higashi, M.; et al. Nectin an adhesion molecule involved in formation of synapses. J. Cell Biol. 2002, 156, 555–565. [Google Scholar] [CrossRef]
- Südhof, T.C. Towards an understanding of synapse formation. Neuron 2018, 100, 276–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagata, M.; Sanes, J.R.; Weiner, J.A. Synaptic adhesion molecules. Curr. Opin. Cell Biol. 2003, 15, 621–632. [Google Scholar] [CrossRef]
- Dalva, M.B.; McClelland, A.C.; Kayser, M.S. Cell adhesion molecules: Signalling functions at the synapse. Nat. Rev. Neurosci. 2007, 8, 206–220. [Google Scholar] [CrossRef] [Green Version]
- Harris, K.M.; Weinberg, R.J. Ultrastructure of synapses in the mammalian brain. Cold Spring Harb. Perspect. Biol. 2012, 4, a005587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, M.A.; Umemori, H. Seeking longterm relationship: Axon and target communicate to organize synaptic differentiation. J. Neurochem. 2006, 97, 1215–1231. [Google Scholar] [CrossRef]
- McAllister, A.K. Dynamic aspects of CNS synapse formation. Neuroscience 2007, 30, 425–450. [Google Scholar] [CrossRef] [Green Version]
- Friedman, L.G.; Riemslagh, F.W.; Sullivan, J.M.; Mesias, R.; Williams, F.M.; Huntley, G.W.; Benson, D.L. Cadherin8 expression, synaptic localization, and molecular control of neuronal form in prefrontal corticostriatal circuits. J. Comp. Neurol. 2015, 523, 75–92. [Google Scholar] [CrossRef] [Green Version]
- Takai, Y.; Miyoshi, J.; Ikeda, W.; Ogita, H. Nectins and nectin-like molecules: Roles in contact inhibition of cell movement and proliferation. Nat. Rev. Mol. Cell Biol. 2008, 9, 603–615. [Google Scholar] [CrossRef]
- Rikitake, Y.; Mandai, K.; Takai, Y. The role of nectins in different types of cell–cell adhesion. J. Cell Sci. 2012, 125, 3713–3722. [Google Scholar] [CrossRef] [Green Version]
- Tomorsky, J.; Parker, P.R.L.; Doe, C.Q.; Niell, C.M. Precise levels of nectin-3 are required for proper synapse formation in postnatal visual cortex. Neural Dev. 2020, 15, 13. [Google Scholar] [CrossRef]
- Wang, X.-D.; Su, Y.-A.; Wagner, K.V.; Avrabos, C.; Scharf, S.H.; Hartmann, J.; Wolf, M.; Liebl, C.; Kühne, C.; Wurst, W.; et al. Nectin-3 links CRHR1 signaling to stress-induced memory deficits and spine loss. Nat. Neurosci. 2013, 16, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-X.; Li, J.-T.; Xie, X.-M.; Gu, Y.; Si, T.-M.; Schmidt, M.V.; Wang, X.-D. Nectin-3 modulates the structural plasticity of dentate granule cells and long-term memory. Transl. Psychiat. 2017, 7, e1228. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, D.; Mandai, K.; Maruo, T.; Supriyanto, I.; Togashi, H.; Inoue, T.; Mori, M.; Takai, Y. Afadin regulates puncta adherentia junction formation and presynaptic differentiation in hippocampal neurons. PLoS ONE 2014, 9, e89763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaudoin, G.M.J.; Schofield, C.M.; Nuwal, T.; Zang, K.; Ullian, E.M.; Huang, B.; Reichardt, L.F. Afadin, a Ras/Rap effector that controls cadherin function, promotes spine and excitatory synapse density in the hippocampus. J. Neurosci. 2012, 32, 99–110. [Google Scholar] [CrossRef]
- Mori, M.; Rikitake, Y.; Mandai, K.; Takai, Y. Roles of Nectins and Nectin-Like Molecules in the Nervous System. In Cell Adhesion Molecules; Springer: New York, NY, USA, 2013; pp. 91–116. [Google Scholar] [CrossRef]
- Pielarski, K.N.; van Stegen, B.; Andreyeva, A.; Nieweg, K.; Jüngling, K.; Redies, C.; Gottmann, K. Asymmetric N-cadherin expression results in synapse dysfunction, synapse elimination, and axon retraction in cultured mouse neurons. PLoS ONE 2013, 8, e54105. [Google Scholar] [CrossRef]
- Frei, J.A.; Niescier, R.F.; Bridi, M.S.; Durens, M.; Nestor, J.E.; Yuan, X.; Dykxhoorn, D.M.; Nestor, M.W.; Huang, S.; Blatt, G.J.; et al. Altered expression of cadherin-8 and cadherin-11 in neural circuit development: Implications for autism. BiorXiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Yuste, R.; Bonhoeffer, T. Genesis of dendritic spines: Insights from ultrastructural and imaging studies. Nat. Rev. Neurosci. 2004, 5, 24–34. [Google Scholar] [CrossRef]
- Bhatt, D.H.; Zhang, S.; Gan, W.-B. Dendritic spine dynamics. Physiology 2009, 71, 261–282. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.; Kim, E. The postsynaptic organization of synapses. Cold Spring Harb. Perspect. Biol. 2011, 3, a005678. [Google Scholar] [CrossRef] [Green Version]
- Tada, T.; Sheng, M. Molecular mechanisms of dendritic spine morphogenesis. Curr. Opin. Neurobiol. 2006, 16, 95–101. [Google Scholar] [CrossRef]
- Chen, C.-C.; Lu, J.; Zuo, Y. Spatiotemporal dynamics of dendritic spines in the living brain. Front. Neuroanat. 2014, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mysore, S.P.; Tai, C.-Y.; Schuman, E.M. N-cadherin, spine dynamics, and synaptic function. Front. Neurosci. 2008, 2, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Togashi, H.; Abe, K.; Mizoguchi, A.; Takaoka, K.; Chisaka, O.; Takeichi, M. Cadherin regulates dendritic spine morphogenesis. Neuron 2002, 35, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Seong, E.; Yuan, L.; Arikkath, J. Cadherins and catenins in dendrite and synapse morphogenesis. Cell Adhes. Migr. 2015, 9, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Penzes, P.; Cahill, M.E.; Jones, K.A.; VanLeeuwen, J.-E.; Woolfrey, K.M. Dendritic spine pathology in neuropsychiatric disorders. Nat. Neurosci. 2011, 14, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Zoghbi, H.Y.; Bear, M.F. Synaptic dysfunction in neurodevelopmental disorders associated with autism and intellectual disabilities. Cold Spring Harb. Perspect. Biol. 2012, 4, a009886. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-C.; Frei, J.A.; Kilander, M.B.C.; Shen, W.; Blatt, G.J. A subset of autism-associated genes regulate the structural stability of neurons. Front. Cell Neurosci. 2016, 10, 263. [Google Scholar] [CrossRef] [Green Version]
- Redies, C.; Hertel, N.; Hübner, C.A. Cadherins and neuropsychiatric disorders. Brain Res. 2012, 1470, 130–144. [Google Scholar] [CrossRef]
- Hawi, Z.; Tong, J.; Dark, C.; Yates, H.; Johnson, B.; Bellgrove, M.A. The role of cadherin genes in five major psychiatric disorders: A literature update. Am. J. Med Genet. Part. B Neuropsychiatr. Genet. 2018, 177, 168–180. [Google Scholar] [CrossRef] [Green Version]
- Casey, J.P.; Magalhaes, T.; Conroy, J.M.; Regan, R.; Shah, N.; Anney, R.; Shields, D.C.; Abrahams, B.S.; Almeida, J.; Bacchelli, E.; et al. A novel approach of homozygous haplotype sharing identifies candidate genes in autism spectrum disorder. Hum. Genet. 2012, 131, 565–579. [Google Scholar] [CrossRef] [Green Version]
- Zhiling, Y.; Fujita, E.; Tanabe, Y.; Yamagata, T.; Momoi, T.; Momoi, M.Y. Mutations in the gene encoding CADM1 are associated with autism spectrum disorder. Biochem. Bioph. Res. Commun. 2008, 377, 926–929. [Google Scholar] [CrossRef] [PubMed]
- Crepel, A.; Wolf, V.D.; Brison, N.; Ceulemans, B.; Walleghem, D.; Peuteman, G.; Lambrechts, D.; Steyaert, J.; Noens, I.; Devriendt, K.; et al. Association of CDH11 with non-syndromic ASD. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2014, 165, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, H.; Ma, D.; Bucan, M.; Glessner, J.T.; Abrahams, B.S.; Salyakina, D.; Imielinski, M.; Bradfield, J.P.; Sleiman, P.M.A.; et al. Common genetic variants on 5p14.1 associate with autism spectrum disorders. Nature 2009, 459, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Pagnamenta, A.T.; Khan, H.; Walker, S.; Gerrelli, D.; Wing, K.; Bonaglia, M.C.; Giorda, R.; Berney, T.; Mani, E.; Molteni, M.; et al. Rare familial 16q21 microdeletions under a linkage peak implicate cadherin 8 (CDH8) in susceptibility to autism and learning disability. J. Med. Genet. 2011, 48, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Fujita, E.; Dai, H.; Tanabe, Y.; Zhiling, Y.; Yamagata, T.; Miyakawa, T.; Tanokura, M.; Momoi, M.Y.; Momoi, T. Autism spectrum disorder is related to endoplasmic reticulum stress induced by mutations in the synaptic cell adhesion molecule, CADM1. Cell Death Dis. 2010, 1, e47. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Liu, L.; Chen, W.; Gao, Q.; Li, H.; Wang, Y.; Qian, Q. The implicated roles of cell adhesion molecule 1 (CADM1) gene and altered prefrontal neuronal activity in attention-deficit/hyperactivity disorder: A “gene–brain–behavior relationship”? Front. Genet. 2019, 10, 882. [Google Scholar] [CrossRef]
- Lasky-Su, J.; Neale, B.M.; Franke, B.; Anney, R.J.L.; Zhou, K.; Maller, J.B.; Vasquez, A.A.; Chen, W.; Asherson, P.; Buitelaar, J.; et al. Genome-wide association scan of quantitative traits for attention deficit hyperactivity disorder identifies novel associations and confirms candidate gene associations. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2008, 147B, 1345–1354. [Google Scholar] [CrossRef] [PubMed]
- Lesch, K.-P.; Timmesfeld, N.; Renner, T.J.; Halperin, R.; Röser, C.; Nguyen, T.T.; Craig, D.W.; Romanos, J.; Heine, M.; Meyer, J.; et al. Molecular genetics of adult ADHD: Converging evidence from genome-wide association and extended pedigree linkage studies. J. Neural Transm. 2008, 115, 1573–1585. [Google Scholar] [CrossRef]
- Sklar, P.; Smoller, J.W.; Fan, J.; Ferreira, M.A.R.; Perlis, R.H.; Chambert, K.; Nimgaonkar, V.L.; McQueen, M.B.; Faraone, S.V.; Kirby, A.; et al. Whole-genome association study of bipolar disorder. Mol. Psychiatr. 2008, 13, 558–569. [Google Scholar] [CrossRef]
- Otsuka, I.; Watanabe, Y.; Sora, I.; Hishimoto, A.; Boku, S.; Mouri, K.; Shiroiwa, K.; Okazaki, S.; Nunokawa, A.; Shirakawa, O.; et al. Association analysis of the cadherin13 gene with schizophrenia in the japanese population. Neuropsych. Dis. Treat. 2015, 11, 1381–1393. [Google Scholar] [CrossRef] [Green Version]
- Bhalla, K.; Luo, Y.; Buchan, T.; Beachem, M.A.; Guzauskas, G.F.; Ladd, S.; Bratcher, S.J.; Schroer, R.J.; Balsamo, J.; DuPont, B.R.; et al. Alterations in CDH15 and KIRREL3 in patients with mild to severe intellectual disability. Am. J. Hum. Genet. 2008, 83, 703–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willemsen, M.H.; Fernandez, B.A.; Bacino, C.A.; Gerkes, E.; de Brouwer, A.P.; Pfundt, R.; Sikkema-Raddatz, B.; Scherer, S.W.; Marshall, C.R.; Potocki, L.; et al. Identification of ANKRD11 and ZNF778 as candidate genes for autism and variable cognitive impairment in the novel 16q24.3 microdeletion syndrome. Eur. J. Hum. Genet. 2010, 18, 429–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moya, P.R.; Dodman, N.H.; Timpano, K.R.; Rubenstein, L.M.; Rana, Z.; Fried, R.L.; Reichardt, L.F.; Heiman, G.A.; Tischfield, J.A.; King, R.A.; et al. Rare missense neuronal cadherin gene (CDH2) variants in specific obsessive-compulsive disorder and tourette disorder phenotypes. Eur. J. Hum. Genet. 2013, 21, 850–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiszniewski, W.; Gawlinski, P.; Gambin, T.; Bekiesinska-Figatowska, M.; Obersztyn, E.; Antczak-Marach, D.; Akdemir, Z.H.C.; Harel, T.; Karaca, E.; Jurek, M.; et al. Comprehensive genomic analysis of patients with disorders of cerebral cortical development. Eur. J. Hum. Genet. 2018, 26, 1121–1131. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Super Family | Family | Member | Common Interactions | Related Neurodevelopmental Process |
|---|---|---|---|---|
| Cadherins | Type I Classical Cadherins | Cdh1 | Cdh1 | Proliferation/differentiation, synaptogenesis |
| Cdh2 | Cdh2, Cdh4 | Proliferation/differentiation, migration, axon guidance, synaptogenesis | ||
| Cdh4 | Cdh4 | Migration, axon guidance | ||
| Type II Classical Cadherins | Cdh6 | Cdh6 | Axon guidance | |
| Cdh8 | Cdh8 | Proliferation/differentiation, Synaptogenesis | ||
| Cdh11 | Cdh11 | Synaptogenesis | ||
| Cdh13 | Cdh13 | Axon guidance | ||
| IgCAMs | nectin | nectin1 | nectin1, nectin3, nectin4, Necl1, tactile | Migration, axon guidance, synaptogenesis |
| nectin3 | nectin1, nectin2, nectin3, necl1, Necl2, Necl5, TIGIT | Migration, axon guidance, synaptogenesis | ||
| Necl | Necl1 | nectin1, nectin3, Necl1, Necl2, Necl3, Necl4 | - | |
| Necl2 | nectin3, Necl1, Necl2, Necl3, Necl2 | Axon guidance, Synaptogenesis | ||
| Necl3 | Necl1, Necl2 | Axon guidance | ||
| Necl4 | Necl1, Necl4 | - |
| Disrupted Adhesive Molecule | Nature of the Genetic Perturbation | Neocorticogenic Processes Affected and Subsequent Phenotypic Alterations | References |
|---|---|---|---|
| Cdh2 | D6-Cre-dependent recombination in Cdh2 F/F mice | Disruption of AJs integrity, laminar and RGCs organization, progenitor proliferation and cell cycle exit | [79] |
| Emx1-Cre-dependent recombination in Cdh2 F/F mice | Disruption of AJs integrity, cell fate choice and axonal pathfinding; enhanced progenitor proliferation; lamination defects and double cortex formation | [80] | |
| CaMKII-driven expression of the Cdh2-intracellular domain | Impaired spine pruning and maturation of neocortical excitatory PNs | [85] | |
| afadin | Emx1-Cre-dependent recombination in Mltt4 F/F mice | Disruption of AJs integrity, cell fate choice and axonal pathfinding; enhanced progenitor proliferation; lamination defects and double cortex formation | [80] |
| Nestin-Cre-dependent recombination in Mltt4 F/F mice | Disruption of AJs integrity; mislocalization of ependymal cells; hydrocephalus and neonatal death | [84] | |
| αE-catenin | Nestin-Cre dependent recombination in αE-catenin F/F mice | Disruption of AJs integrity; cell cycle shortening, increased proliferation and decreased apoptosis of progenitor cells; cortical dysplasia and hyperplasia | [83] |
| αN-catenin | Spontaneous mutation leading to αN-catenin gene deletion | Impaired spine pruning and maturation of neocortical excitatory PNs | [85] |
| β-catenin | D6-Cre-dependent recombination in β-catenin F/F mice | Disruption of AJs integrity, interkinetic nuclear migration, RGCs organization, radial migration of early-born neurons and cell fate choice; decreased progenitor proliferation | [82] |
| Nex-Cre-dependent recombination in β-catenin F(ex3)/F(ex3) mice generating a stable and active form of β-catenin | Enhanced synapse and spine stability and density in layer II/III excitatory PNs’ apical and basal dendrites | [86] | |
| Scnn1a-Tg3-Cre-dependent recombination in β-catenin F(ex3)/F(ex3) mice, generating a stabilized and active form of β-catenin only in layer IV excitatory PNs | Enhanced synapse and spine stability and density in layer II/III excitatory PNs’ basal dendrites | [86] | |
| CaMKII-Cre-ERT2-dependent recombination in β-catenin F/F mice | Impaired spine pruning and maturation of neocortical excitatory PNs | [85] | |
| Necl2 | Knockin mice with the lacZ-neo-cassette replacing part of Necl2 gene sequence, ablating Necl2 expression | Decreased density of thalamic synaptic inputs in PV+ interneurons from layer IV of the primary visual neocortex | [87] |
| C-CAMs | Genetic Marker * | Neurodevelopmental Disorder | Neocortex-Related Phenotype | References | |
|---|---|---|---|---|---|
| Cdh SF | CDH2 | De novo mutations (D353N, D597N, D597Y, D601T, C613W, D627G, Y676C, L855V, L856F) | ID | Yes | [143] |
| rs17445840, rs2289664, De novo mutations (N706S, V289I) | TD, OCD | No | [195] | ||
| CDH4 | De novo mutations (E451K, A852T, R659P) | MCD | Yes | [196] | |
| CDH7 | rs1444067, rs2850700, rs2658046, rs12970791, rs2850699, rs4455070 | BD | No | [191] | |
| CDH8 | De novo mutation (16q21 microdeletion) | ASD, LD | Yes | [186] | |
| CDH9/CDH10 | rs4307059, rs7704909, rs12518194, rs4327572, rs1896731, rs10038113 | ASD | No | [185] | |
| CDH11 | rs10500464 rs429065 | ASD, ADHD | No | [184] | |
| CDH13 | rs6565113, rs11646411 | ADHD | No | [189,190] | |
| rs8057927 | SCZ | No | [192] | ||
| CDH15 | De novo mutations (V8L, R60C, R92W, A122V) | ID | No | [193] | |
| De novo mutation (16q24.3 microdeletion) | ID | Yes | [194] | ||
| Necls | NECL2 | rs10891819 | ADHD | Yes | [188] |
| De novo mutations (H246N, Y251S) | ASD | No | [187] | ||
| De novo mutations (H246N, Y251S) | ASD | No | [183] | ||
| NECL3 | De novo mutation (3p12.1 microdeletion) | ASD | No | [182] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Agustín-Durán, D.; Mateos-White, I.; Fabra-Beser, J.; Gil-Sanz, C. Stick around: Cell–Cell Adhesion Molecules during Neocortical Development. Cells 2021, 10, 118. https://doi.org/10.3390/cells10010118
de Agustín-Durán D, Mateos-White I, Fabra-Beser J, Gil-Sanz C. Stick around: Cell–Cell Adhesion Molecules during Neocortical Development. Cells. 2021; 10(1):118. https://doi.org/10.3390/cells10010118
Chicago/Turabian Stylede Agustín-Durán, David, Isabel Mateos-White, Jaime Fabra-Beser, and Cristina Gil-Sanz. 2021. "Stick around: Cell–Cell Adhesion Molecules during Neocortical Development" Cells 10, no. 1: 118. https://doi.org/10.3390/cells10010118
APA Stylede Agustín-Durán, D., Mateos-White, I., Fabra-Beser, J., & Gil-Sanz, C. (2021). Stick around: Cell–Cell Adhesion Molecules during Neocortical Development. Cells, 10(1), 118. https://doi.org/10.3390/cells10010118