
Perspectives on miRNAs Targeting DKK1 for Developing Hair Regeneration Therapy

 ,
,  ,
,
Abstract
:1. Introduction
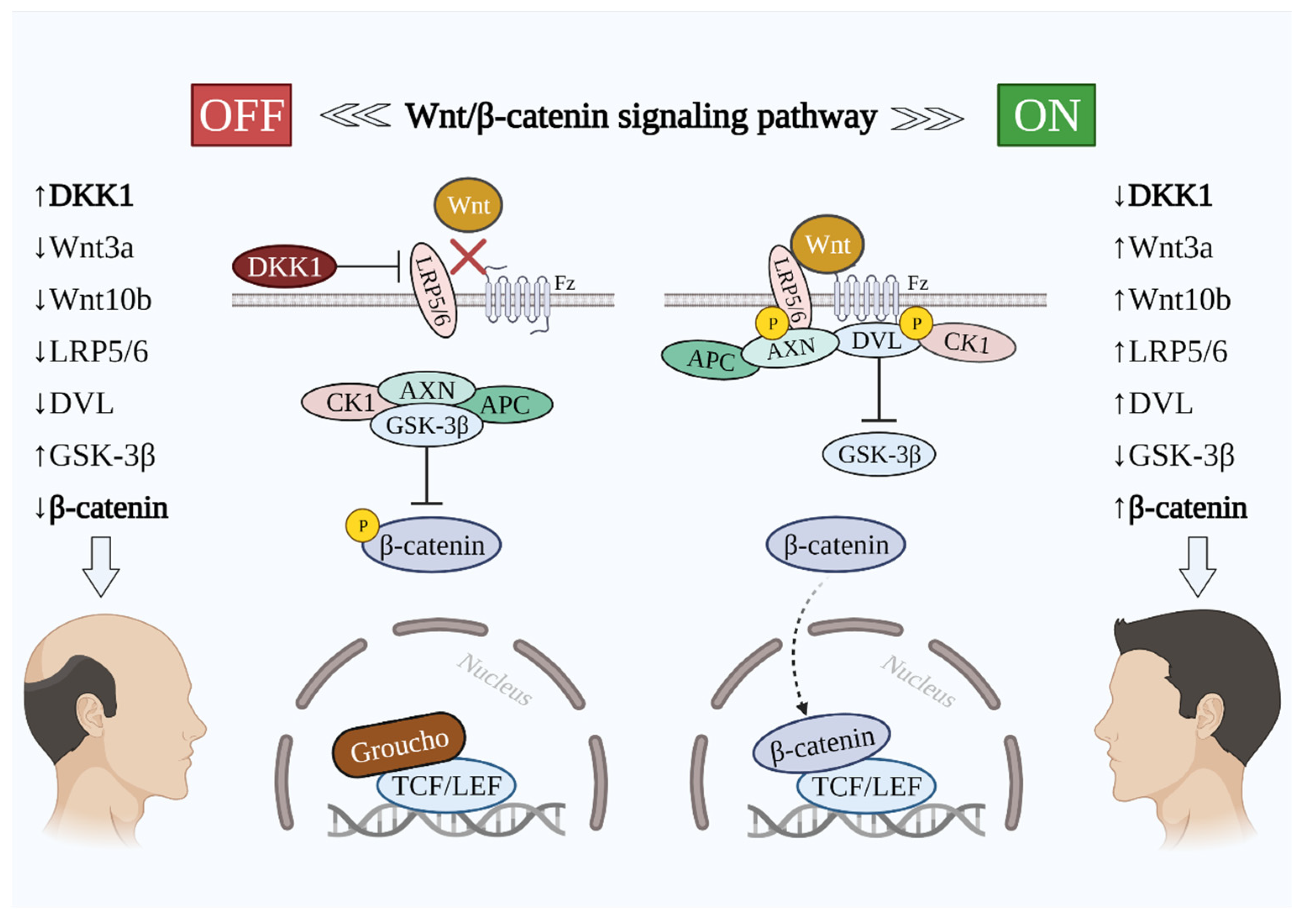
2. The Importance of Wnt/β-Catenin Signaling in Hair Growth
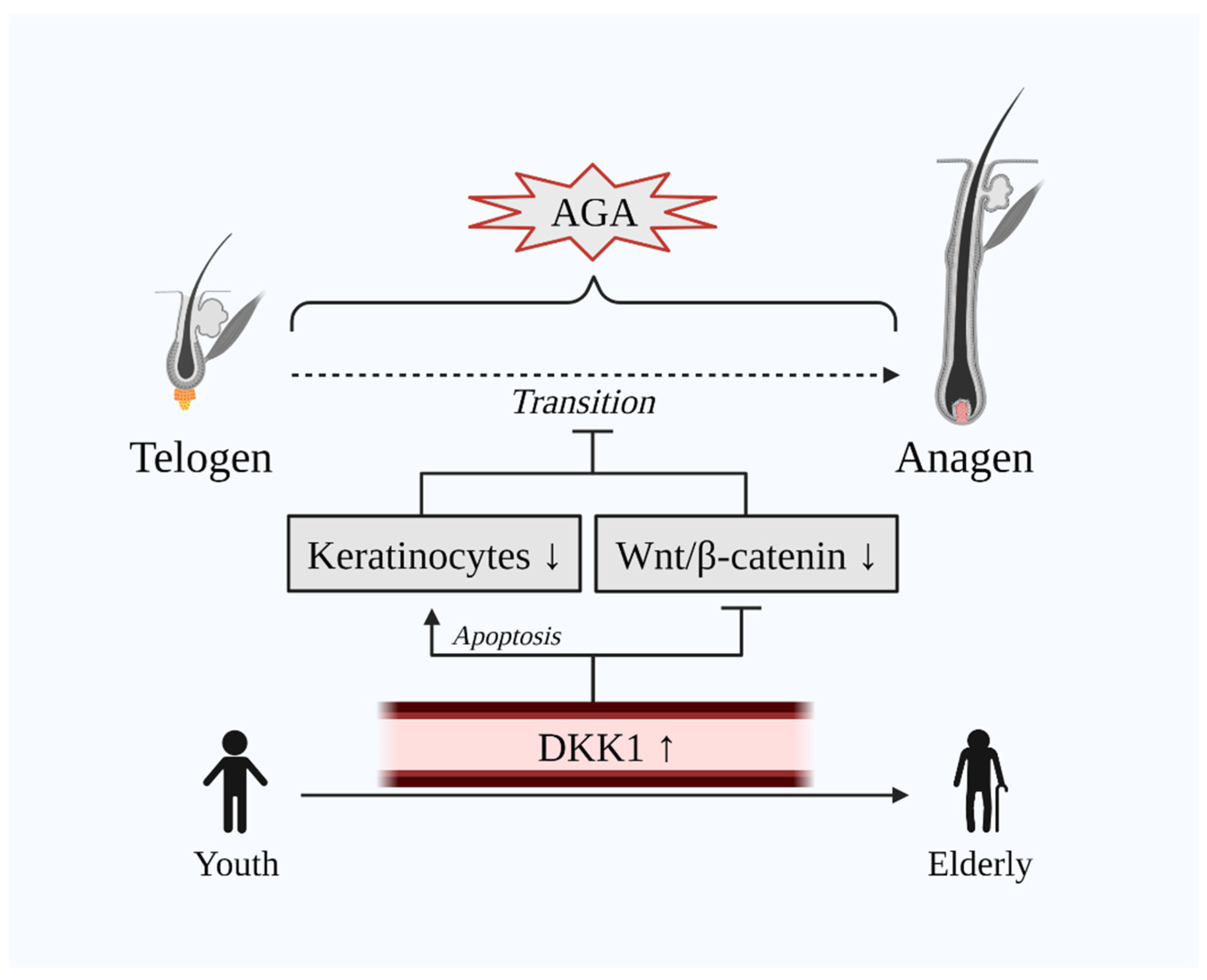
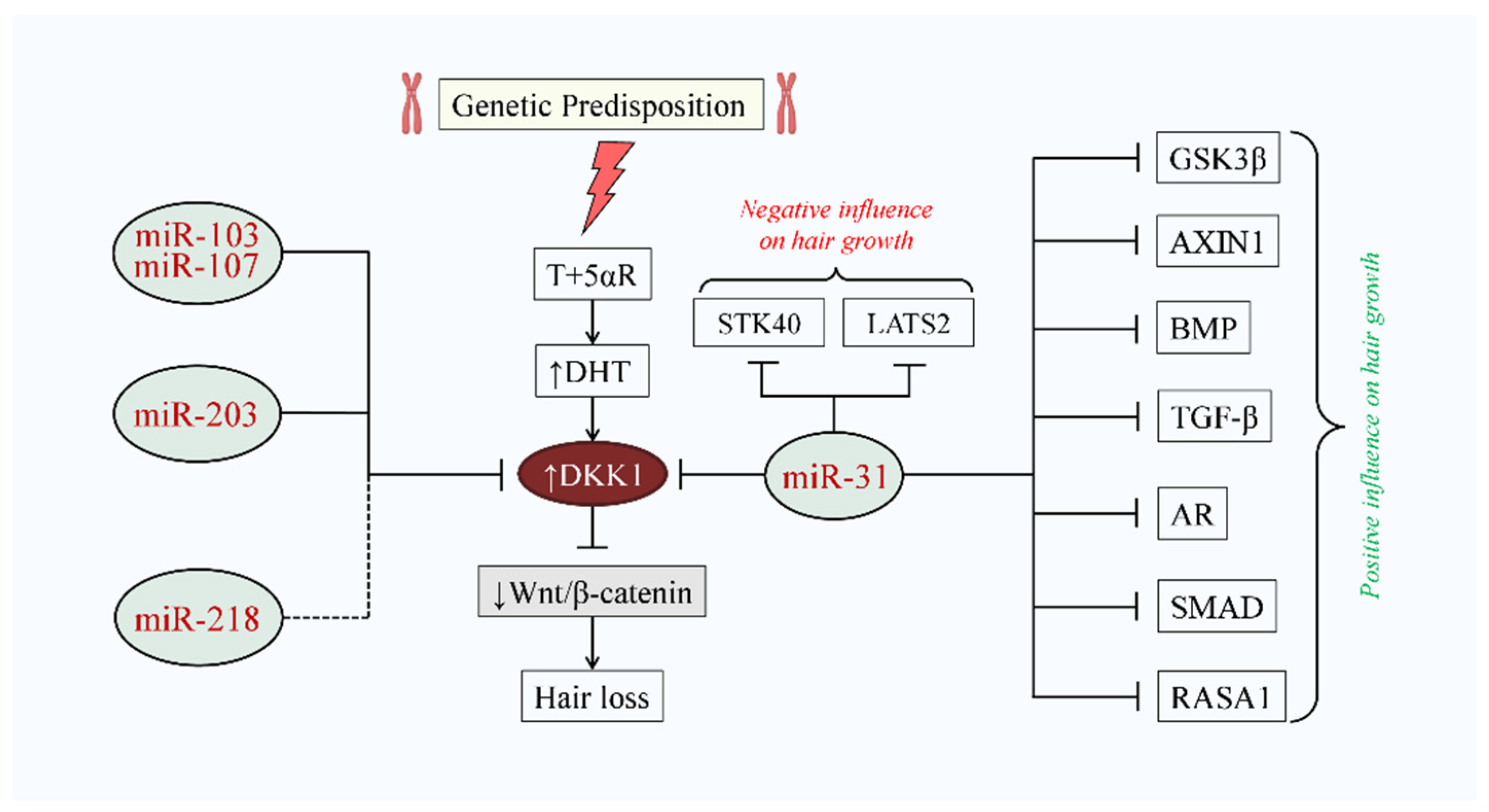
3. DKK1 Implication in AGA
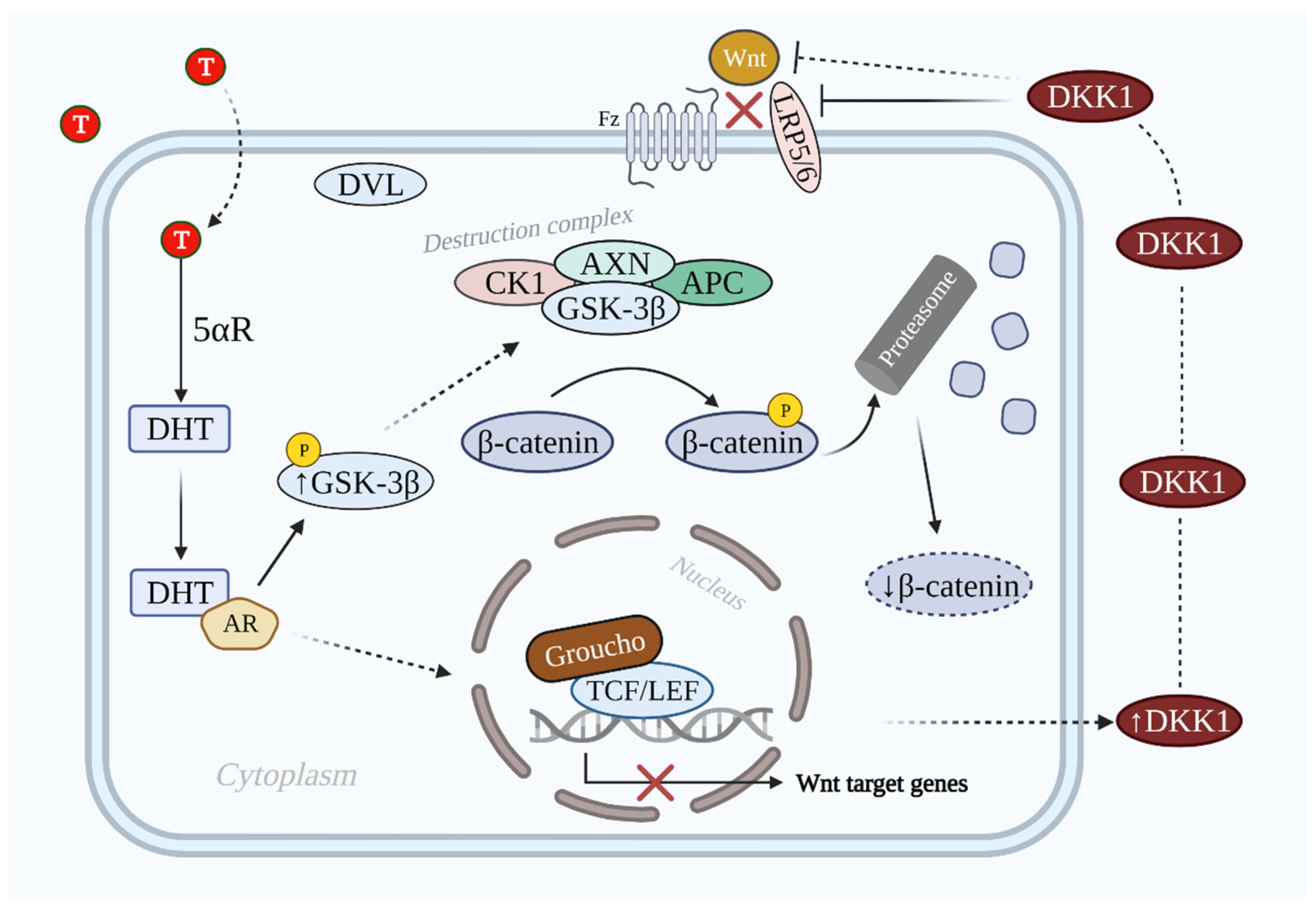
3.1. DHT-Induced DKK1-Mediated AGA
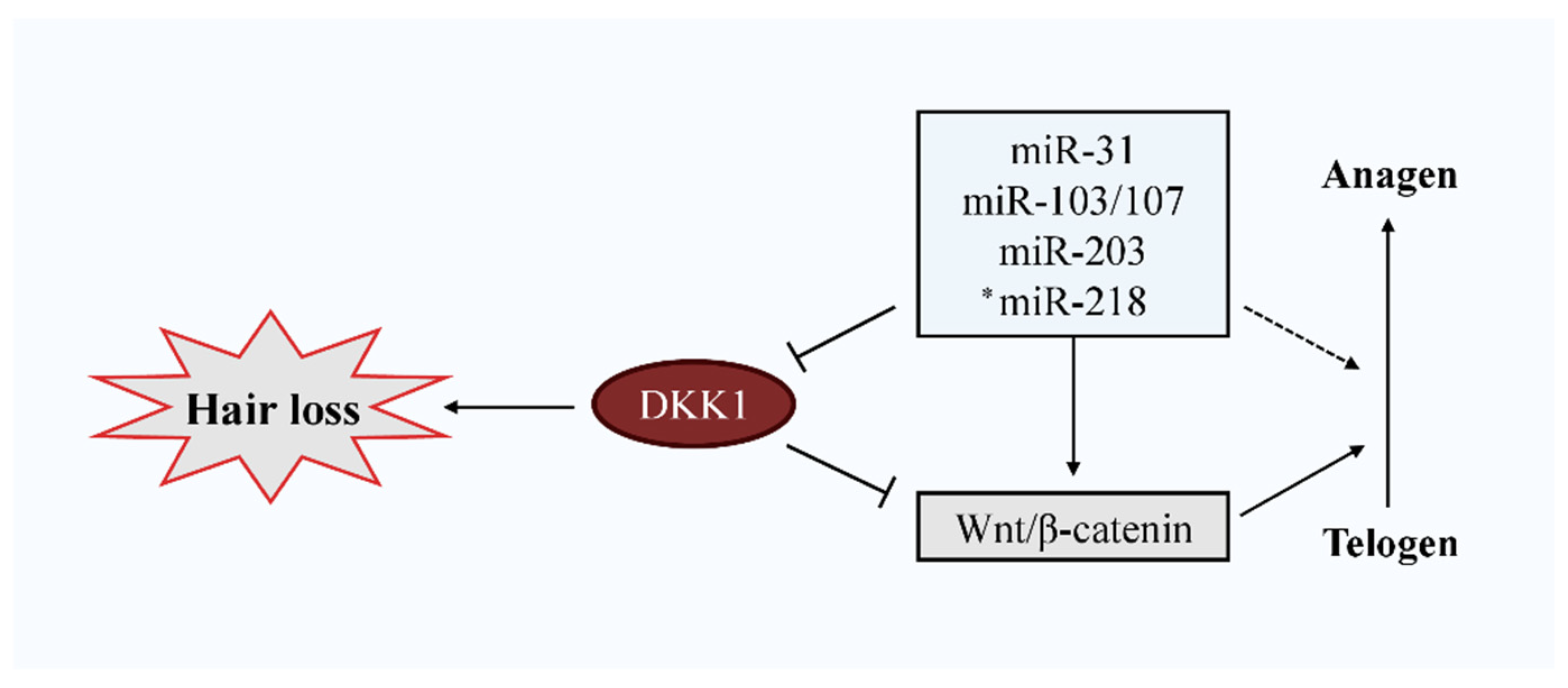
3.2. Negative Influence of DKK1 on Hair Growth
3.3. Molecular Mechanism of Wnt/β-Catenin Signaling Inhibition by DKK1
3.4. Compounds Capable to Inhibit DKK1 Expression and Promote Hair Growth
4. miRNAs Targeting DKK1 Expression
5. Available and Recently Studied Therapeutics for AGA
6. miRNA Involvement in Hair Growth Regulation
6.1. miR-29
6.2. miR-31
6.3. miR-103/107
6.4. miR-152
6.5. miR-203
6.6. miR-218
7. Main Challenge: Delivery Systems
8. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, C.L.; Huang, W.Y.; Wang, E.H.C.; Tai, K.Y.; Lin, S.J. Functional complexity of hair follicle stem cell niche and therapeutic targeting of niche dysfunction for hair regeneration. J. Biomed. Sci. 2020, 27, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, B.Y. Targeting wnt/β-catenin pathway for developing therapies for hair loss. Int. J. Mol. Sci. 2020, 21, 4915. [Google Scholar] [CrossRef] [PubMed]
- Grymowicz, M.; Rudnicka, E.; Podfigurna, A.; Napierala, P.; Smolarczyk, R.; Smolarczyk, K.; Meczekalski, B. Hormonal E ff ects on Hair Follicles. Int. J. Mol. Sci. 2020, 21, 5342. [Google Scholar] [CrossRef] [PubMed]
- Houschyar, K.S.; Borrelli, M.R.; Tapking, C.; Popp, D.; Puladi, B.; Ooms, M.; Chelliah, M.P.; Rein, S.; Pförringer, D.; Thor, D.; et al. Molecular mechanisms of hair growth and regeneration: Current understanding and novel paradigms. Dermatology 2020, 236, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Zhang, K.; Ye, J.X.; Lian, X.H.; Yang, T. Wnt10b promotes growth of hair follicles via a canonical Wnt signalling pathway. Clin. Exp. Dermatol. 2011, 36, 534–540. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhang, K.; Yang, K.; Ye, J.X.; Xing, Y.Z.; Guo, H.Y.; Deng, F.; Lian, X.H.; Yang, T. Adenovirus-mediated wnt10b overexpression induces hair follicle regeneration. J. Investig. Dermatol. 2013, 133, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Liu, Y.; Song, Z.; Hao, F.; Yang, X. Identification of Wnt/β-catenin signaling pathway in dermal papilla cells of human scalp hair follicles: TCF4 regulates the proliferation and secretory activity of dermal papilla cell. J. Dermatol. 2014, 41, 84–91. [Google Scholar] [CrossRef]
- Lin, W.; Xiang, L.; Shi, H.; Zhang, J.; Jiang, L.; Cai, P.; Lin, Z.; Lin, B.; Huang, Y.; Zhang, H.; et al. Fibroblast Growth Factors Stimulate Hair Growth through β-Catenin and Shh Expression in C57BL/6 Mice. Biomed Res. Int. 2015, 2015, 730139. [Google Scholar] [CrossRef] [Green Version]
- Gentile, P.; Garcovich, S. Advances in Regenerative Stem Cell Therapy in Androgenic Alopecia and Hair Loss: Wnt Pathway, Growth-Factor, and Mesenchymal Stem Cell Signaling Impact Analysis on Cell Growth and Hair Follicle Development. Cells 2019, 8, 466. [Google Scholar] [CrossRef] [Green Version]
- Kwack, M.H.; Sung, Y.K.; Chung, E.J.; Im, S.U.; Ahn, J.S.; Kim, M.K.; Kim, J.C. Dihydrotestosterone-inducible dickkopf 1 from balding dermal papilla cells causes apoptosis in follicular keratinocytes. J. Investig. Dermatol. 2008, 128, 262–269. [Google Scholar] [CrossRef] [Green Version]
- Fawzi, M.M.T.; Mahmoud, S.B.; Shaker, O.G.; Saleh, M.A. Assessment of tissue levels of dickkopf-1 in androgenetic alopecia and alopecia areata. J. Cosmet. Dermatol. 2016, 15, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dickkopf 1 promotes regression of hair follicles. J. Investig. Dermatol. 2012, 132, 1554–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Premanand, A.; Rajkumari, B.R. Androgen modulation of Wnt/β-catenin signaling in androgenetic alopecia. Arch. Dermatol. Res. 2018, 310, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Sferrazza, G.; Corti, M.; Brusotti, G.; Pierimarchi, P.; Temporini, C.; Serafino, A.; Calleri, E. Nature-derived compounds modulating Wnt/β-catenin pathway: A preventive and therapeutic opportunity in neoplastic diseases. Acta Pharm. Sin. B 2020, 10, 1814–1834. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X. Targeting the Wnt/β-catenin signaling pathway in cancer. J. Hematol. Oncol. 2020, 13, 165. [Google Scholar] [CrossRef]
- Bao, J.; Yang, Y.; Xia, M.; Sun, W.; Chen, L. Wnt signaling: An attractive target for periodontitis treatment. Biomed. Pharmacother. 2021, 133, 110935. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef]
- Huang, C.; Ye, Z.; Wan, J.; Liang, J.; Liu, M.; Xu, X.; Li, L. Secreted Frizzled-Related Protein 2 Is Associated with Disease Progression and Poor Prognosis in Breast Cancer. Dis. Markers 2019, 2019, 6149381. [Google Scholar] [CrossRef]
- Zhu, G.; Song, J.; Chen, W.; Yuan, D.; Wang, W.; Chen, X.; Liu, H.; Su, H.; Zhu, J. Expression and Role of Dickkopf-1 (Dkk1) in Tumors: From the Cells to the Patients. Cancer Manag. Res. 2021, 13, 659–675. [Google Scholar] [CrossRef]
- Choi, Y.S.; Zhang, Y.; Xu, M.; Yang, Y.; Ito, M.; Peng, T.; Cui, Z.; Nagy, A.; Hadjantonakis, A.K.; Lang, R.A.; et al. Distinct functions for Wnt/β-Catenin in hair follicle stem cell proliferation and survival and interfollicular epidermal homeostasis. Cell Stem Cell 2013, 13, 720–733. [Google Scholar] [CrossRef] [Green Version]
- Flores-hernández, E.; Velázquez, D.M.; Castañeda-patlán, M.C.; Fuentes-garcía, G.; Fonseca-camarillo, G.; Yamamoto-furusho, J.K.; Romero-Avila, M.T.; García-Sáinz, J.A.; Robles-Flores, M. Canonical and non-canonical Wnt signaling are simultaneously activated by Wnts in colon cancer cells. Cell. Signal. 2020, 72, 109636. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.F.; Kaur, P.; Bunnag, N.; Suresh, J.; Sung, I.C.H.; Tan, Q.H.; Gruber, J.; Tolwinski, N.S. WNT Signaling in Disease. Cells 2019, 8, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, P.; Yang, J.; Liu, L.; Yang, X.; Yao, Z.; Ma, C.; Zhu, H.; Su, J.; Zhao, Q.; Feng, K.; et al. ERK1/2 inhibition reduces vascular calcification by activating miR-126-3p-DKK1/LRP6 pathway. Theranostics 2021, 11, 1129–1146. [Google Scholar] [CrossRef] [PubMed]
- Tosti, A.; Zaiac, M.N.; Canazza, A.; Sanchis-gomar, F.; Pareja-galeano, H.; Alis, R.; Lucia, A.; Emanuele, E. Topical application of the Wnt/β-catenin activator methyl vanillate increases hair count and hair mass index in women with androgenetic alopecia. J. Cosmet. Dermatol. 2016, 15, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, J.; Jing, J.; Wu, X.; Lv, Z. Serum levels of androgen-associated hormones are correlated with curative effect in androgenic alopecia in young men. Med. Sci. Monit. 2018, 24, 7770–7777. [Google Scholar] [CrossRef]
- Chen, X.; Liu, B.; Li, Y.; Han, L.; Tang, X.; Deng, W.; Lai, W. Dihydrotestosterone regulates hair growth through the Wnt/β-catenin Pathway in C57BL/6 Mice and in vitro organ culture. Front. Pharmacol. 2020, 10, 1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, D.; Huang, J.; Li, K.; Chen, Y.; He, Y.; Sun, Y.; Guo, Y.; Du, L.; Qu, Q.; Miao, Y.; et al. Dihydrotestosterone-induced hair regrowth inhibition by activating androgen receptor in C57BL6 mice simulates androgenetic alopecia. Biomed. Pharmacother. 2021, 137, 111247. [Google Scholar] [CrossRef]
- Kishimoto, J.; Burgeson, R.E.; Morgan, B.A. Wnt signaling maintains the hair-inducing activity of the dermal papilla. Genes Dev. 2000, 14, 1181–1185. [Google Scholar] [CrossRef]
- Shin, H.R.; Kwack, M.H.; Shin, S.H.; Oh, J.W.; Kang, B.M.; Kim, A.A.; Kim, J.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Identification of transcriptional targets of Wnt/β-catenin signaling in dermal papilla cells of human scalp hair follicles: EP2 is a novel transcriptional target of Wnt3a. J. Dermatol. Sci. 2010, 58, 91–96. [Google Scholar] [CrossRef]
- Xing, F.; Yi, W.J.; Miao, F.; Su, M.Y.; Lei, T.C. Baicalin increases hair follicle development by increasing canonical Wnt/β-catenin signaling and activating dermal papillar cells in mice. Int. J. Mol. Med. 2018, 41, 2079–2085. [Google Scholar] [CrossRef] [Green Version]
- Lei, M.; Guo, H.; Qiu, W.; Lai, X.; Yang, T.; Randall, B.; Chuong, C.; Lian, X.; Yang, L. Modulating hair follicle size with Wnt10b-DKK1 pair during hair regeneration. Exp. Dermatol. 2015, 23, 407–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemayel, R.; Chenette, E.J. β-catenin signalling in dermal papilla cells leads to a hairy situation. FEBS J. 2016, 283, 2820–2822. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, W.; Sinha, S.; Biernaskie, J. Immune modulation of hair follicle regeneration. npj Regen. Med. 2020, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schlake, T.; Sick, S. Canonical WNT Signalling Controls Hair Follicle Spacing. Cell Adh. Migr. 2007, 1, 149–151. [Google Scholar] [CrossRef] [Green Version]
- Enshell-Seijffers, D.; Lindon, C.; Kashiwagi, M.; Morgan, B.A. β-catenin Activity in the Dermal Papilla Regulates Morphogenesis and Regeneration of Hair. Dev. Cell 2010, 18, 633–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rishikaysh, P.; Dev, K.; Diaz, D.; Shaikh Qureshi, W.M.; Filip, S.; Mokry, J. Signaling involved in hair follicle morphogenesis and development. Int. J. Mol. Sci. 2014, 15, 1647–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimelman, D.; Xu, W. β-catenin destruction complex: Insights and questions from a structural perspective. Oncogene 2006, 25, 7482–7491. [Google Scholar] [CrossRef] [Green Version]
- Daniels, D.L.; Weis, W.I. β-catenin directly displaces Groucho/TLE repressors from Tcf/Lef in Wnt-mediated transcription activation. Nat. Struct. Mol. Biol. 2005, 12, 364–371. [Google Scholar] [CrossRef]
- Chen, H.; Lang, Y.; Han-nan, L.; Liu, Y.; Liao, C.; Nana, A.W.; Yen, Y.; Chen, R. miR-103/107 prolong Wnt/β-catenin signaling and colorectal cancer stemness by targeting Axin2. Sci. Rep. 2019, 9, 9687. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, E.A.; Elgarhy, L.H.; Hasby, E.A.; Mohammad, L. Dickkopf-1 Expression in Androgenetic Alopecia and Alopecia Areata in Male Patients. Am. J. Dermatopathol. 2019, 41, 122–127. [Google Scholar] [CrossRef]
- Kwack, M.H.; Ahn, J.S.; Jang, J.H.; Kim, J.C.; Sung, Y.K.; Kim, M.K. SFRP2 augments Wnt/β-catenin signalling in cultured dermal papilla cells. Exp. Dermatol. 2016, 25, 813–815. [Google Scholar] [CrossRef]
- Paul, S.; Licona-Vázquez, I.; Serrano-Cano, F.I.; Frías-Reid, N.; Pacheco-Dorantes, C.; Pathak, S.; Chakraborty, S.; Srivastava, A. Current insight into the functions of microRNAs in common human hair loss disorders: A mini review. Hum. Cell 2021, 34, 1040–1050. [Google Scholar] [CrossRef]
- Niehrs, C. Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene 2006, 25, 7469–7481. [Google Scholar] [CrossRef] [Green Version]
- Seib, D.R.M.; Corsini, N.S.; Ellwanger, K.; Plaas, C.; Mateos, A.; Pitzer, C.; Niehrs, C.; Celikel, T.; Martin-Villalba, A. Loss of dickkopf-1 restores neurogenesis in old age and counteracts cognitive decline. Cell Stem Cell 2013, 12, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dovjak, P.; Dorfer, S.; Föger-Samwald, U.; Kudlacek, S.; Marculescu, R.; Pietschmann, P. Serum levels of sclerostin and dickkopf-1: Effects of age, gender and fracture status. Gerontology 2014, 60, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.H.; Sood, T.; Zito, P.M. Androgenetic Alopecia; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Yang, C.C.; Hsieh, F.N.; Lin, L.Y.; Hsu, C.K.; Sheu, H.M.; Chen, W. Higher body mass index is associated with greater severity of alopecia in men with male-pattern androgenetic alopecia in Taiwan: A cross-sectional study. J. Am. Acad. Dermatol. 2014, 70, 297–302.e1. [Google Scholar] [CrossRef]
- Morinaga, H.; Mohri, Y.; Grachtchouk, M.; Asakawa, K.; Matsumura, H.; Oshima, M.; Takayama, N.; Kato, T.; Nishimori, Y.; Sorimachi, Y.; et al. Obesity accelerates hair thinning by stem cell-centric converging mechanisms. Nature 2021, 595, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Zmuda, J.M.; Cvejkus, R.K.; Kershaw, E.E.; Kuipers, A.L.; Oczypok, E.A.; Wheeler, V.; Bunker, C.H.; Miljkovic, I. Wnt pathway inhibitor DKK1: A potential novel biomarker for adiposity. J. Endocr. Soc. 2019, 3, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colditz, J.; Picke, A.; Hofbauer, L.C.; Rauner, M. Contributions of Dickkopf-1 to Obesity-Induced Bone. J. Bone Miner. Res. 2020, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Lim, C.; Lee, J.Y.; Im, K.R.; Yoon, K.S.; Song, J.M. Visible-to-near IR quantum dot-based hypermulticolor high-content screening of herbal medicines for the efficacy monitoring of hair growth promotion and hair loss inhibition. J. Biomol. Screen. 2013, 18, 462–473. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Wang, H.; Jing, J.; Yu, L.; Wu, X.; Lu, Z. Morroniside regulates hair growth and cycle transition via activation of the Wnt/β -catenin signaling pathway. Sci. Rep. 2018, 8, 13785. [Google Scholar] [CrossRef] [PubMed]
- Cselenyi, C.S.; Lee, E. Context-dependent activation or Inhibition of Wnt-beta-catenin signaling by kremen. Sci. Signal. 2008, 1, pe10. [Google Scholar] [CrossRef] [PubMed]
- Rothbächer, U.; Lemaire, P. Crème de la Kremen of Wnt signalling inhibition. Nat. Cell Biol. 2002, 4, E172–E173. [Google Scholar] [CrossRef]
- Luo, J.; Chen, M.; Liu, Y.; Xie, H.; Yuan, J.; Zhou, Y.; Ding, J.; Deng, Z.; Li, J. Nature-derived lignan compound VB-1 exerts hair growth-promoting effects by augmenting Wnt/β-catenin signaling in human dermal papilla cells. PeerJ 2018, 6, e4737. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, S.U.N.A.; Hong, Y.D.; Park, B.C.; Na, Y. Panax ginseng extract antagonizes the effect of DKK-1-induced catagen-like changes of hair follicles. Int. J. Mol. Med. 2017, 40, 1194–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.E.; Choi, H.C.; Nam, G.; Choi, B.Y. Costunolide promotes the proliferation of human hair follicle dermal papilla cells and induces hair growth in C57BL/6 mice. J. Cosmet. Dermatol. 2019, 18, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Shi, Q.; Nan, W.; Wang, Y.; Wang, S.; Yang, F.; Li, G. Ginkgolide B and bilobalide promote the growth and increase β-catenin expression in hair follicle dermal papilla cells of American minks. BioFactors 2019, 45, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.S.; Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Attenuation of Dickkopf 1-Induced Hair Growth Inhibition in Cultured Human Hair Follicles by Tianeptine. Ann. Dermatol. 2017, 29, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Feinbaum, R.; Ambros, V.; Lee, R. The C. elegans Heterochronic Gene lin-4 Encodes Small RNAs with Antisense Complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar]
- Pasquinelli, A.E.; Reinhart, B.J.; Slack, F.; Martindale, M.Q.; Kuroda, M.I.; Maller, B.; Hayward, D.C.; Ball, E.E.; Degnan, B.; Mu, P.; et al. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 2000, 408, 86–89. [Google Scholar] [CrossRef]
- Gazerani, P. Current Evidence on Potential Uses of MicroRNA Biomarkers for Migraine: From Diagnosis to Treatment. Mol. Diagn. Ther. 2019, 23, 681–694. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.P.; Castresana, J.S.; Shahi, M.H. Glioblastoma and MiRNAs. Cancers 2021, 13, 1581. [Google Scholar] [CrossRef] [PubMed]
- Bouchie, A. First microRNA mimic enters clinic. Nat. Biotechnol. 2013, 31, 577. [Google Scholar] [CrossRef]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.S. Therapeutic advances of miRNAs: A preclinical and clinical update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef]
- Zhang, S.; Cheng, Z.; Wang, Y.; Han, T. The Risks of miRNA Therapeutics: In a Drug Target Perspective. Drug Des. Dev. Ther. 2021, 15, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. 2018 FDA drug approvals. Nat. Rev. Drug Discov. 2019, 18, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. 2019 FDA drug approvals. Nat. Rev. Drug Discov. 2020, 19, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.M.; Bahal, R.; Rasmussen, T.P.; Manautou, J.E.; Zhong, X.B. The growth of siRNA-based therapeutics: Updated clinical studies. Biochem. Pharmacol. 2021, 189, 114432. [Google Scholar] [CrossRef]
- Ozcan, G.; Ozpolat, B.; Coleman, R.L.; Sood, A.K.; Lopez-Berestein, G. Preclinical and clinical development of siRNA-based therapeutics. Adv. Drug Deliv. Rev. 2015, 87, 108–119. [Google Scholar] [CrossRef] [Green Version]
- Hoy, S.M. Patisiran: First Global Approval. Drugs 2018, 78, 1625–1631. [Google Scholar] [CrossRef]
- Ledford, H. Gene-silencing drug approved Outrage over changes to EPA chemical assessments. Nature 2018, 560, 291–292. [Google Scholar] [CrossRef]
- Ning, M.S.; Andl, T. Control by a hair’s breadth: The role of microRNAs in the skin. Cell. Mol. Life Sci. 2013, 70, 1149–1169. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xia, S.; Wang, T.; Wang, S.; Yuan, D.; Li, F.; Wang, X. Chi-miR-30b-5p inhibits dermal papilla cells proliferation by targeting CaMKIIδ gene in cashmere goat. BMC Genom. 2020, 21, 430. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.I.; Alam, M.; Emelianov, V.U.; Poterlowicz, K.; Patel, A.; Sharov, A.A.; Mardaryev, A.N.; Botchkareva, N. V MicroRNA-214 controls skin and hair follicle development by modulating the activity of the Wnt pathway. J. Cell Biol. 2014, 207, 549–567. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Chen, Y.; Yang, N.; Chen, Q.; Wu, X. miR-218-5p regulates skin and hair follicle development through Wnt/β-catenin signaling pathway by targeting SFRP2. J. Cell. Physiol. 2019, 234, 1–13. [Google Scholar] [CrossRef]
- Hu, S.; Li, Z.; Lutz, H.; Huang, K.; Su, T.; Cores, J.; Dinh, P.U.C.; Cheng, K. Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling. Sci. Adv. 2020, 6, eaba1685. [Google Scholar] [CrossRef] [PubMed]
- Andl, T.; Reddy, S.T.; Gaddapara, T.; Millar, S.E. WNT signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653. [Google Scholar] [CrossRef]
- Li, S.; Yin, Y.; Yao, L.; Lin, Z.; Sun, S.; Zhang, J.; Li, X. TNF-α treatment increases DKK1 protein levels in primary osteoblasts via upregulation of DKK1 mRNA levels and downregulation of miR-335-5p. Mol. Med. Rep. 2020, 22, 1017–1025. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tu, Q.; Bonewald, L.F.; He, X.; Stein, G.; Lian, J.; Chen, J. Effects of miR-335-5p in modulating osteogenic differentiation by specifically downregulating Wnt antagonist DKK1. J. Bone Miner. Res. 2011, 26, 1953–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, L.D.; Reygagne, P.; Benech, P.; Scalvino, S.; So, S.L.K.; Hamidou, Z.; Bianovici, S.; Pouch, J.; Ducos, B.; Bonnet, M.; et al. Study of gene expression alteration in male androgenetic alopecia: Evidence of predominant molecular signalling pathways. Br. J. Dermatol. 2017, 177, 1322–1336. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, J.; Chen, Z.; Wang, K.; Shi, L. MicroRNA-1-3p inhibits proliferation and migration of oral squamous cell carcinoma cells by targeting DKK1. Biochem. Cell Biol. 2018, 96, 355–364. [Google Scholar] [CrossRef]
- Mirfazeli, E.S.; Arefian, E.; Nadri, S.; Rezazadeh Valojerdi, R.; Kehtari, M.; Zeynali, B. DKK1 expression is suppressed by miR-9 during induced dopaminergic differentiation of human trabecular meshwork mesenchymal stem cells. Neurosci. Lett. 2019, 707, 134250. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, P.; Gu, J. miR-29a modulates tumor necrosis factor-α-induced osteogenic inhibition by targeting Wnt antagonists. Dev. Growth Differ. 2015, 57, 264–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, C.; Li, F.; Li, X.; Tian, Y.; Zhang, Y.; Sheng, X.; Song, Y.; Meng, Q.; Yuan, S.; Luan, L.; et al. MiR-31 promotes mammary stem cell expansion and breast tumorigenesis by suppressing Wnt signaling antagonists. Nat. Commun. 2017, 8, 1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Liu, H.; Li, C.; Liang, H. miR-33a-5p inhibits the progression of esophageal cancer through the DKK1-mediated Wnt/β-catenin pathway. Aging 2021, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Liu, T.; Yu, C.; Yang, X.; Shao, Y.; Shi, J.; Ye, X.; Zheng, X.; Yan, J.; Xu, D.; et al. LncRNA TUG1 alleviates cardiac hypertrophy by targeting miR-34a/DKK1/Wnt-β-catenin signalling. J. Cell. Mol. Med. 2020, 24, 3678–3691. [Google Scholar] [CrossRef]
- Xiang, J.; Fu, H.Q.; Xu, Z.; Fan, W.; Liu, F.; Chen, B. lncRNA SNHG1 attenuates osteogenic differentiation via the miR-101/DKK1 axis in bone marrow mesenchymal stem cells. Mol. Med. Rep. 2020, 22, 3715–3722. [Google Scholar] [CrossRef]
- Liu, J.U.N.; Wu, M.; Feng, G.; Li, R.U.I.; Wang, Y.; Jiao, J. Downregulation of LINC00707 promotes osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by regulating DKK1 via targeting miR-103a-3p. Int. J. Mol. Med. 2020, 46, 1029–1038. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, J.; Shao, Z.; Pu, F.; Wang, B.; Wu, Q.; Zhang, Y.; Zeng, X.; Guo, X.; Yang, S.; et al. In vitro effect of microRNA-107 targeting Dkk-1 by regulation of Wnt/β-catenin signaling pathway in osteosarcoma. Medicine 2017, 96, 27. [Google Scholar] [CrossRef]
- Liao, Y.; Jia, X.; Ren, Y.; Deji, Z.; Gesang, Y.; Ning, N.; Feng, H.; Yu, H.; Wei, A. Suppressive role of microRNA-130b-3p in ferroptosis in melanoma cells correlates with DKK1 inhibition and Nrf2-HO-1 pathway activation. Hum. Cell 2021. [Google Scholar] [CrossRef]
- Di, G.; Kong, L.; Zhao, Q.; Ding, T. MicroRNA-146a knockdown suppresses the progression of ankylosing spondylitis by targeting dickkopf 1. Biomed. Pharmacother. 2018, 97, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, B.; George, S.K.; Liu, B. Downregulation of MicroRNA-152 contributes to high expression of DKK1 in multiple myeloma. RNA Biol. 2015, 12, 1314–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Lin, Y.; Kang, X.; Liu, Z.; Zhang, W.; Xu, F. microRNA-186 in extracellular vesicles from bone marrow mesenchymal stem cells alleviates idiopathic pulmonary fibrosis via interaction with SOX4 and DKK1. Stem Cell Res. Ther. 2021, 12, 96. [Google Scholar] [CrossRef]
- Cheng, R.; Lu, C.; Zhang, G.; Zhang, G.; Zhao, G. Overexpression of miR-203 increases the sensitivity of NSCLC A549/H460 cell lines to cisplatin by targeting Dickkopf-1. Oncol. Rep. 2017, 37, 2129–2136. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Wang, Y.; Sun, Q.; Du, X. MiR-203 is involved in osteoporosis by regulating DKK1 and inhibiting osteogenic differentiation of MSCs. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5098–5105. [Google Scholar] [CrossRef]
- Jia, Y.; Chen, L.; Guo, S.; Li, Y. Baicalin induced colon cancer cells apoptosis through miR-217/DKK1-mediated inhibition of Wnt signaling pathway. Mol. Biol. Rep. 2019, 46, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Yu, M.; Xie, X.; Huang, G.; Peng, Y.; Ren, D. miR-217 targeting DKK1 promotes cancer stem cell properties via activation of the Wnt signaling pathway in hepatocellular carcinoma. Oncol. Rep. 2017, 38, 2351–2359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.; Jin, Y.; Zheng, J.; Liu, K.; Zhao, J.; Zhang, S.; Wu, F.; Sun, Z. MiR-217 promotes cell proliferation and osteogenic differentiation of BMSCs by targeting DKK1 in steroid-associated osteonecrosis. Biomed. Pharmacother. 2019, 109, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hu, H.; Zhang, X.; Liu, G.; Ran, B.; Zhang, P.; Liao, M.; Wu, Y. MiR-291a-3p regulates the BMSCs differentiation via targeting DKK1 in dexamethasone-induced osteoporosis. Kaohsiung J. Med. Sci. 2019, 36, 35–42. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, Y.; Yang, Z.; Zhu, Y.; Xie, Y.; Zhou, F.; Cai, L. Elevation of miR-302b prevents multiple myeloma cell growth and bone destruction by blocking DKK1 secretion. Cancer Cell Int. 2021, 21, 187. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, F.; Wang, F.; Wu, N. Long noncoding RNA MAGI1-IT1 regulates cardiac hypertrophy by modulating miR-302e/DKK1/Wnt/beta-catenin signaling pathway. J. Cell. Physiol. 2019, 235, 245–253. [Google Scholar] [CrossRef]
- Luan, Y.; Xie, B.; Wei, W. REST-repressed lncRNA NPPA-AS1 regulates cervical cancer progression by modulating miR-302e/DKK1/Wnt/β-catenin signaling pathway. J. Cell. Biochem. 2021, 122, 16–28. [Google Scholar] [CrossRef]
- Li, J.; Feng, Z.; Chen, L.; Wang, X.; Deng, H. MicroRNA-335-5p inhibits osteoblast apoptosis induced by high glucose. Mol. Med. Rep. 2016, 13, 4108–4112. [Google Scholar] [CrossRef]
- Lin, S.C.; Wu, H.L.; Yeh, L.Y.; Yang, C.C.; Kao, S.Y.; Chang, K.W. Activation of the mir-371/372/373 mirna cluster enhances oncogenicity and drug resistance in oral carcinoma cells. Int. J. Mol. Sci. 2020, 21, 9442. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Zhang, H.; Wang, C.; Liang, J.; Chen, G.; Li, W.; Tang, H.; Hou, J. MIR-373-3p Targets DKK1 to Promote EMT-Induced Metastasis via the Wnt/β-Catenin Pathway in Tongue Squamous Cell Carcinoma. Biomed. Res. Int. 2017, 2017, 6010926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; He, Y.; Rui, J.; Xu, M.-Q. miR-410 acts as an oncogene in colorectal cancer cells by targeting dickkopf-related protein 1 via the Wnt/β-catenin signaling pathway. Oncol. Lett. 2019, 17, 807–814. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Lin, J.; Wang, G.; Lu, J. MicroRNA-433-3p promotes osteoblast differentiation through targeting DKK1 expression. PLoS ONE 2017, 12, e0179860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Hu, X.-Y.; Cao, C.; Zhao, Y.-W.; He, S.-H. MiR-488 promotes fracture healing by targeting DKK1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8965–8972. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Li, N.; Peng, C.; Deng, Y.; Wang, J.; Deng, M.; Lu, M.; Yin, J.; Zheng, G.; Liu, H.; et al. miR-493 mediated DKK1 down-regulation confers proliferation, invasion and chemo-resistance in gastric cancer cells. Oncotarget 2016, 7, 7044–7054. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, C.; Chen, M.; Li, Z.; Tian, S.; Jiang, J. miR-522 contributes to cell proliferation of hepatocellular carcinoma by targeting DKK1 and SFRP2. Tumor Biol. 2016, 37, 11321–11329. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Y.; Li, Q.; Ma, D.; Nie, A.; Shen, X. Biochemical and Biophysical Research Communications LncRNA Linc-PINT inhibits miR-523-3p to hamper retinoblastoma progression by upregulating Dickkopf-1 (DKK1). Biochem. Biophys. Res. Commun. 2020, 530, 47–53. [Google Scholar] [CrossRef]
- Liu, L.; Zuo, Y.; Xu, Y.; Zhang, Z.; Li, Y.; Pang, J. MiR-613 inhibits proliferation and invasion and induces apoptosis of rheumatoid arthritis synovial fibroblasts by direct downregulation of DKK1. Cell. Mol. Biol. Lett. 2019, 24, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Liao, H.; Zhang, Y.; Cao, Z. MicroRNA-3064-3p regulates the differentiation of cementoblasts through targeting DKK1. J. Periodontal Res. 2018, 53, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Hua, Q.; Zhou, Y. CircRNA has_circ_0006427 suppresses the progression of lung adenocarcinoma by regulating miR-6783-3p/DKK1 axis and inactivating Wnt/β-catenin signaling pathway. Biochem. Biophys. Res. Commun. 2018, 508, 37–45. [Google Scholar] [CrossRef]
- Yao, Y.; Zhou, Y.; Hua, Q. circRNA hsa_circ_0018414 inhibits the progression of LUAD by sponging miR-6807-3p and upregulating DKK1. Mol. Ther. Nucleic Acid 2021, 23, 783–796. [Google Scholar] [CrossRef]
- Min, K.; Lee, S.K. EBV miR-BART10-3p Promotes Cell Proliferation and Migration by Targeting DKK1. Int. J. Biol. Sci. 2019, 15, 657–667. [Google Scholar] [CrossRef]
- Dong, M.; Gong, L.; Chen, J.; Zhang, X.; Zhang, Y.; Hui, D. EBV-miR-BART10-3p and EBV-miR-BART22 promote metastasis of EBV-associated gastric carcinoma by activating the canonical Wnt signaling pathway. Cell. Oncol. 2020, 43, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Kumaresan, M.; Mysore, V. Controversies in Hair Transplantation. Muthuvel Kumaresan Venkatram Mysore 2018, 11, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Suchonwanit, P.; Thammarucha, S.; Leerunyakul, K. Minoxidil and its use in hair disorders: A review. Drug Des. Dev. Ther. 2019, 13, 2777–2786. [Google Scholar] [CrossRef] [Green Version]
- Panchaprateep, R.; Lueangarun, S. Efficacy and safety of oral minoxidil 5 mg once daily in the treatment of male patients with androgenetic alopecia: An open-label and global photographic assessment. Dermatol. Ther. 2020, 10, 1345–1357. [Google Scholar] [CrossRef]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef] [Green Version]
- Kwack, M.H.; Kang, B.M.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Minoxidil activates β-catenin pathway in human dermal papilla cells: A possible explanation for its anagen prolongation effect. J. Dermatol. Sci. 2011, 62, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Choi, N.; Shin, S.; Song, S.U.; Sung, J. Minoxidil Promotes Hair Growth through Stimulation of Growth Factor Release from Adipose-Derived Stem Cells. Int. J. Mol. Sci. 2018, 19, 691. [Google Scholar] [CrossRef] [Green Version]
- Andy, G.; John, M.; Mirna, S.; Rachita, D.; Michael, K.; Maja, K.; Aseem, S.; Zeljana, B. Controversies in the treatment of androgenetic alopecia: The history of finasteride. Dermatol. Ther. 2019, 32, e12647. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Na, J.; Bak, D.H.O.; Lee, B.C.; Lee, E.; Choi, M.I.J.I.; Ryu, C.H.O.; Lee, S.; Mun, S.K.; Park, B.C.; et al. Development of finasteride polymer microspheres for systemic application in androgenic alopecia. Int. J. Mol. Med. 2019, 43, 2409–2419. [Google Scholar] [CrossRef] [PubMed]
- Reichenberg, J.S.; Kelsey, P.A.; Therrien, C.A.; Gavino, C.; Reichenberg, J.S. Adverse effects and safety of 5-alpha reductase inhibitors (finasteride, dutasteride): A systematic review. J. Clin. Aesthet. Dermatol. 2016, 9, 56–62. [Google Scholar]
- Arif, T.; Dorjay, K.; Adil, M.; Sami, M. Dutasteride in Androgenetic Alopecia: An Update. Curr. Clin. Pharmacol. 2017, 12, 31–35. [Google Scholar] [CrossRef] [PubMed]
- York, K.; Meah, N.; Bhoyrul, B.; Sinclair, R.; York, K.; Meah, N.; Bhoyrul, B.; Sinclair, R.; York, K.; Meah, N.; et al. Expert Opinion on Pharmacotherapy Treatment review for male pattern hair-loss Treatment review for male pattern hair-loss. Expert Opin. Pharmacother. 2020, 21, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Motofei, I.G.; Rowland, D.L.; Tampa, M.; Sarbu, M.-I.; Mitran, M.-I.; Mitran, C.-I.; Stoian, A.P.; Diaconu, C.C.; Paunica, S.; Georgescu, S.R. Finasteride and androgenic alopecia; from therapeutic options to medical implications. J. Dermatolog. Treat. 2019, 31, 415–421. [Google Scholar] [CrossRef]
- Marihart, S.; Harik, M.; Djavan, B. Dutasteride: A Review of Current Data on a Novel Dual Inhibitor of. Rev. Urol. 2005, 7, 203–210. [Google Scholar]
- Zhou, Z.; Song, S.; Gao, Z.; Wu, J.; Ma, J.; Cui, Y. The efficacy and safety of dutasteride compared with finasteride in treating men with androgenetic alopecia: A systematic review and meta-analysis. Clin. Interv. Aging 2019, 14, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.S.; Ghatak, S.; El Masry, M.S.; Gnyawali, S.C.; Roy, S.; Amer, M.; Everts, H.; Sen, C.K.; Khanna, S. Epidermal E-Cadherin Dependent β-Catenin Pathway Is Phytochemical Inducible and Accelerates Anagen Hair Cycling. Mol. Ther. 2017, 25, 2502–2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamblin, M.R. NIH Public Access. Laser Surg. Med. 2014, 46, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Liao, P.P.; Liu, L.H.; Fang, X.; Li, W.; Guan, S.M. Baicalin and geniposide inhibit the development of atherosclerosis by increasing Wnt1 and inhibiting dickkopf-related protein-1 expression. J. Geriatr. Cardiol. 2016, 13, 846–854. [Google Scholar] [CrossRef]
- Bejaoui, M.; Villareal, M.O.; Isoda, H. β-catenin-mediated hair growth induction effect of 3,4,5-tri-O-caffeoylquinic acid. Aging 2019, 11, 4216–4237. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.I.; Choi, Y.K.; Koh, Y.S.; Hyun, J.W.; Kang, J.H.; Lee, K.S.; Lee, C.M.; Yoo, E.S.; Kang, H.K. Vanillic acid stimulates anagen signaling via the PI3K/Akt/β-catenin pathway in dermal papilla cells. Biomol. Ther. 2020, 28, 354–360. [Google Scholar] [CrossRef]
- Lim, K.M.; An, S.; Lee, O.K.; Lee, M.J.; Lee, J.P.; Lee, K.S.; Lee, G.T.; Lee, K.K.; Bae, S. Analysis of changes in microRNA expression profiles in response to the troxerutin-mediated antioxidant effect in human dermal papilla cells. Mol. Med. Rep. 2015, 12, 2650–2660. [Google Scholar] [CrossRef] [Green Version]
- Ohn, J.; Been, K.W.; Kim, J.Y.; Kim, E.J.; Park, T.; Yoon, H.; Ji, J.S.; Okada-iwabu, M.; Iwabu, M.; Yamauchi, T.; et al. Discovery of a transdermally deliverable pentapeptide for activating AdipoR1 to promote hair growth. EMBO Mol. Med. 2021, 13, e13790. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.L.; Gangadaran, P.; Bak, S.S.; Oh, J.M.; Kalimuthu, S.; Lee, H.W.; Baek, S.H.; Zhu, L.; Sung, Y.K.; Jeong, S.Y.; et al. Extracellular vesicles derived from MSCs activates dermal papilla cell in vitro and promotes hair follicle conversion from telogen to anagen in mice. Sci. Rep. 2017, 7, 15560. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.R.; Logarinho, E. Tissue engineering strategies for human hair follicle regeneration: How far from a hairy goal? Concise review. Stem Cells Transl. Med. 2020, 9, 342–350. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Lin, H.; Wang, L.; Guo, K.; Jing, R.; Li, X. Suppression of FGF5 and FGF18 Expression by Cholesterol-Modified siRNAs Promotes Hair Growth in Mice. Front. Pharmacol. 2021, 12, 666860. [Google Scholar] [CrossRef] [PubMed]
- Kwack, M.H.; Seo, C.H.; Gangadaran, P.; Ahn, B.C.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Exosomes derived from human dermal papilla cells promote hair growth in cultured human hair follicles and augment the hair-inductive capacity of cultured dermal papilla spheres. Exp. Dermatol. 2019, 28, 854–857. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, C.; Harikumar, K.B. The origin and functions of exosomes in cancer. Front. Oncol. 2018, 8, 66. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowska, A.; Braniewska, A.; Kozar-kaminska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. 2017, 26, 865–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Brien, J.O.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, B.P.; Shih, I.; Jones-rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of Mammalian MicroRNA Targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Andl, T.; Murchison, E.P.; Liu, F.; Zhang, Y.; Yunta-gonzalez, M.; Tobias, J.W.; Andl, C.D.; Seykora, J.T.; Hannon, G.J.; Millar, S.E. Report The miRNA-Processing Enzyme Dicer Is Essential for the Morphogenesis and Maintenance of Hair Follicles. Curr. Biol. 2006, 16, 1041–1049. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Hu, T.; Han, L.; Liu, B.; Tang, X.; Chen, H.; Chen, X.; Wan, M. miRNA microarray profiling in patients with androgenic alopecia and the effects of miR-133b on hair growth. Exp. Mol. Pathol. 2021, 118, 104589. [Google Scholar] [CrossRef]
- Goodarzi, H.R.; Abbasi, A.; Saffari, M.; Fazelzadeh Haghighi, M.; Tabei, M.B.; Noori Daloii, M.R. Differential expression analysis of balding and nonbalding dermal papilla microRNAs in male pattern baldness with a microRNA amplification profiling method. Br. J. Dermatol. 2012, 166, 1010–1016. [Google Scholar] [CrossRef]
- Bernard, B.A. Advances in Understanding Hair Growth. F1000Research 2016, 5, F1000. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, N.; Fukui, S.; Takatani, A.; Shimizu, T.; Umeda, M.; Nishino, A.; Igawa, T.; Koga, T.; Kawashiri, S.; Ichinose, K.; et al. Osteogenic differentiation of fibroblast-like synovial cells in rheumatoid arthritis is induced by microRNA-218 through a ROBO/Slit pathway. Arthritis Res. Ther. 2018, 20, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.C.; Pirro, A.E.; Amling, M.; Delling, G.; Baron, R.; Bronson, R.; Demay, M.B. Targeted ablation of the vitamin D receptor: An animal model of vitamin D-dependent rickets type II with alopecia. Proc. Natl. Acad. Sci. USA 1997, 94, 9831–9835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisse, T.S.; Saini, V.; Zhao, H.; Luderer, H.F.; Gori, F.; Demay, M.B. The Vitamin D Receptor Is Required for Activation of cWnt and Hedgehog Signaling in Keratinocytes. Mol. Endocrinol. 2014, 28, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Yi, R.; O’Carroll, D.; Pasolli, H.A.; Zhang, Z.; Dietrich, F.S.; Tarakhovsky, A.; Fuchs, E. Morphogenesis in skin is governed by discrete sets of differentially expressed microRNAs. Nat. Genet. 2006, 38, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Li, J.; Jiang, Q.; Wu, S.; Jiang, H.; Zhang, Q. Differential expression of miR-let7a in hair follicle cycle of Liaoning cashmere goats and identification of its targets. Funct. Integr. Genom. 2018, 18, 701–707. [Google Scholar] [CrossRef]
- Liu, N.; Niu, S.; Cao, X.-R.; Cheng, J.-Q.; Gao, S.-Y.; Yu, X.-J.; Wang, H.-D.; Dong, C.-S.; He, X.-Y. Let-7b regulates alpaca hair growth by downregulating ectodysplasin A. Mol. Med. Rep. 2018, 17, 4688–4694. [Google Scholar] [CrossRef] [PubMed]
- Zhai, B.; Zhang, L.; Wang, C.; Zhao, Z.; Zhang, M.; Li, X. Identification of microRNA-21 target genes associated with hair follicle development in sheep. PeerJ 2019, 7, e7167. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.; Li, M.; Zheng, Y.; Yin, Y.; Jin, F.; Li, X.; Dong, J.; Liu, X.; Zhang, K.; Li, D.; et al. EZH2-mediated inhibition of microRNA-22 promotes differentiation of hair follicle stem cells by elevating STK40 expression. Aging 2020, 12, 12726–12739. [Google Scholar] [CrossRef]
- Yuan, S.; Li, F.; Meng, Q.; Zhao, Y.; Chen, L.; Zhang, H.; Xue, L.; Zhang, X.; Lengner, C.; Yu, Z. Post-transcriptional Regulation of Keratinocyte Progenitor Cell Expansion, Differentiation and Hair Follicle Regression by miR-22. PLoS Genet. 2015, 11, e1005253. [Google Scholar] [CrossRef]
- Amelio, I.; Lena, A.M.; Bonanno, E.; Melino, G.; Candi, E. miR-24 affects hair follicle morphogenesis targeting Tcf-3. Cell Death Dis. 2013, 4, e922. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Zhang, X.; Peng, Y.; Zhang, L.; Yu, Y.; Hua, P.; Zhu, P.; Yan, X.; Li, Y. miR-24 controls the regenerative competence of hair follicle progenitors by targeting Plk3. Cell Rep. 2021, 35, 109225. [Google Scholar] [CrossRef] [PubMed]
- Kapinas, K.; Kessler, C.; Ricks, T.; Gronowicz, G.; Delany, A.M. miR-29 modulates Wnt signaling in human osteoblasts through a positive feedback loop*. J. Biol. Chem. Chem. 2010, 285, 25221–25231. [Google Scholar] [CrossRef] [Green Version]
- Kapinas, K.; Kessler, C.B.; Delany, A.M. miR-29 Suppression of Osteonectin in Osteoblasts: Regulation During Differentiation and by Canonical Wnt Signaling. J. Cell. Biochem. 2009, 108, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.; Chang, P.; Ho, C.; Huang, Y.; Shih, Y. Protective effects of miR-29a on diabetic glomerular dysfunction by modulation of DKK1/Wnt/β-catenin signaling. Sci. Rep. 2016, 6, 30575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, N.; Lin, E.; Zhang, H.; Liu, Y.; Cao, G.; Fu, C. LncRNA H19 Overexpression Activates Wnt Signaling to Maintain the Hair Follicle Regeneration Potential of Dermal Papilla Cells. Front. Genet. 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Liu, C.; Li, L.; Lan, M.; Yu, Y.; Gu, L.; Su, Y.; Zhang, K.; Zhang, Y.; Wang, T.; et al. miR-29a/b1 Inhibits Hair Follicle Stem Cell Lineage Progression by Spatiotemporally Suppressing WNT and BMP Signaling. Cell Rep. 2019, 29, 2489–2504. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Wang, J.; Ma, J.; Zhang, L.; Chu, C.; Hu, H.; Wang, Y.; Li, Y. miR-31-5p promotes proliferation and inhibits apoptosis of goat hair follicle stem cells by targeting RASA1/MAP3K1 pathway. Exp. Cell Res. 2021, 398, 112441. [Google Scholar] [CrossRef]
- Mardaryev, A.N.; Ahmed, M.I.; Vlahov, N.V.; Fessing, M.Y.; Gill, J.H.; Sharov, A.A.; Botchkareva, N.V. Micro-RNA-31 controls hair cycle-associated changes in gene expression programs of the skin and hair follicle. FASEB J. 2010, 24, 3869–3881. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Zhong, L.; Li, X.; Liu, W.; Zhao, Y.; Li, J. Down-regulation of microRNA-31-5p inhibits proliferation and invasion of osteosarcoma cells through Wnt/β-catenin signaling pathway by enhancing AXIN1. Exp. Mol. Pathol. 2019, 108, 32–41. [Google Scholar] [CrossRef]
- Luan, L.; Shi, J.; Yu, Z.; Andl, T. The major miR-31 target genes STK40 and LATS2 and their implications in the regulation of keratinocyte growth and hair differentiation. Exp. Dermatol. 2017, 26, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kobeissi, A.; Dong, Y.; Kaplan, N.; Yang, W.; He, C. MicroRNAs-103/107 Regulate Autophagy in the Epidermis. J. Investig. Dermatol. 2018, 138, 1481–1490. [Google Scholar] [CrossRef] [Green Version]
- Mokabber, H.; Najafzadeh, N.; Vardi, M.M. miR-124 promotes neural differentiation in mouse bulge stem cells by repressing Ptbp1 and Sox9. J. Cell. Physiol. 2019, 234, 8941–8950. [Google Scholar] [CrossRef] [PubMed]
- Peng, T.; Peng, J.J.; Miao, G.Y.; Tan, Z.Q.; Liu, B.; Zhou, E. miR-125/CDK2 axis in cochlear progenitor cell proliferation. Mol. Med. Rep. 2021, 23, 102. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Hai, E.; Di, Z.; Ma, R.; Shang, F.; Wang, M.; Liang, L.; Rong, Y.; Pan, J.; Su, R.; et al. Chi-miR-130b-3p regulates Inner Mongolia cashmere goat skin hair follicles in fetuses by targeting Wnt family member 10A. G3 Genes Genomes Genet. 2021, 11, jkaa023. [Google Scholar] [CrossRef]
- Ding, Y.; Xue, X.; Liu, Z.; Ye, Y.; Xiao, P.; Pu, Y. Expression Profiling and Functional Characterization of miR-26a and miR-130a in Regulating Zhongwei Goat Hair Development via the TGF-β/SMAD Pathway. Int. J. Mol. Sci. 2020, 21, 5076. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, J.; Liu, Z.; Chen, R.; Fu, D.; Yang, L.; Wang, J.; Du, L.; Wen, L.; Miao, Y.; et al. miR-140-5p in Small Extracellular Vesicles from Human Papilla Cells Stimulates Hair Growth by Promoting Proliferation of Outer Root Sheath and Hair Matrix Cells. Front. Cell Dev. Biol. 2020, 8, 593638. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Gao, W.; Jin, C.; Wang, L.; Wang, Y.; Chen, W.; Zou, S.; Huang, S.; Li, Z.; Wang, J.; et al. Preliminary study on microR-148a and microR-10a in dermal papilla cells of Hu sheep. BMC Genet. 2019, 20, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Li, Q.; Su, B.; Yu, M. MicroRNA-148b promotes proliferation of hair follicle cells by targeting NFAT5. Front. Agric. Sci. Eng. 2016, 3, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Qu, J.; Li, Y.; Feng, Y.; Ma, J.; Zhang, L.; Chu, C.; Hu, H.; Wang, Y.; Ji, D. miR-149-5p Regulates Goat Hair Follicle Stem Cell Proliferation and Apoptosis by Targeting the CMTM3/AR Axis During Superior-Quality Brush Hair Formation. Front. Genet. 2020, 11, 529757. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudian-Sani, M.R.; Jami, M.S.; Mahdavinezhad, A.; Amini, R.; Farnoosh, G.; Saidijam, M. The effect of the microRNA-183 family on hair cell-specific markers of human bone marrow-derived mesenchymal stem cells. Audiol. Neurotol. 2018, 23, 208–215. [Google Scholar] [CrossRef]
- Zhou, W.E.I.; Du, J.; Jiang, D.I.; Wang, X.; Chen, K.; Tang, H. microRNA-183 is involved in the differentiation and regeneration of Notch signaling-prohibited hair cells from mouse cochlea. Mol. Med. Rep. 2018, 18, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.W.; Han, J.H.; Wu, L.; Choi, J.Y. microRNA-183 is Essential for Hair Cell Regeneration after Neomycin Injury in Zebrafish. Yonsei Med. J. 2018, 59, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Yang, F.; Zhao, M.; Ma, L.; Li, H.; Xie, Y.; Nai, R. The intragenic mRNA-microRNA regulatory network during telogen-anagen hair follicle transition in the cashmere goat. Sci. Rep. 2018, 8, 14227. [Google Scholar] [CrossRef] [Green Version]
- Zhu, N.; Huang, K.; Liu, Y.; Zhang, H.; Lin, E.; Zeng, Y.; Li, H.; Xu, Y.; Cai, B.; Yuan, Y.; et al. miR-195-5p Regulates Hair Follicle Inductivity of Dermal Papilla Cells by Suppressing Wnt/β-Catenin Activation. Biomed. Res. Int. 2018, 2018, 4924356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoefert, J.E.; Bjerke, G.A.; Wang, D.; Yi, R. The microRNA-200 family coordinately regulates cell adhesion and proliferation in hair morphogenesis. J. Cell Biol. 2018, 217, 2185–2204. [Google Scholar] [CrossRef] [Green Version]
- Warshauer, E.; Samuelov, L.; Sarig, O.; Vodo, D.; Bindereif, A.; Kanaan, M.; Gat, U.; Fuchs-telem, D.; Shomron, N.; Farberov, L.; et al. RBM28, a protein deficient in ANE syndrome, regulates hair follicle growth via miR-203 and p63. Exp. Dermatol. 2015, 24, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Li, J.; Li, J.; Wu, S.; Jiang, H. Expression of miRNA-203 and its target gene in hair follicle cycle development of Cashmere goat. Cell Cycle 2021, 20, 204–210. [Google Scholar] [CrossRef]
- Luo, Z.; Dou, J.; Xie, F.; Lu, J.; Han, Q. miR-203a-3p promotes loureirin A-induced hair follicle stem cells differentiation by targeting Smad1. Anat. Rec. 2020, 304, 531–540. [Google Scholar] [CrossRef]
- Liu, G.; Li, S.; Liu, H.; Zhu, Y.; Bai, L.; Sun, H.; Gao, S.; Jiang, W.; Li, F. The functions of ocu-miR-205 in regulating hair follicle development in Rex rabbits. BMC Dev. Biol. 2020, 20, 8. [Google Scholar] [CrossRef] [PubMed]
- Du, K.T.; Deng, J.Q.; He, X.G.; Liu, Z.p.; Peng, C.; Zhang, M.S. MiR-214 Regulates the Human Hair Follicle Stem Cell Proliferation and Differentiation by Targeting EZH2 and Wnt/β-Catenin Signaling Way In Vitro. Tissue Eng. Regen. Med. 2018, 15, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Li, M.; Li, X.; Zheng, Y.; Zhang, K.; Liu, X.; Cai, B.; Yin, G. DNMT1-mediated methylation inhibits microRNA-214-3p and promotes hair follicle stem cell differentiate into adipogenic lineages. Stem Cell Res. Ther. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Mohammadi, P.; Ali, M.; Kass, K.; Shari, A.; Moradi, S.; Khosravani, P.; Aghdami, R.; Taheri, P.; Hosseini, G.; Baharvand, H.; et al. Defining microRNA signatures of hair follicular stem and progenitor cells in healthy and androgenic alopecia patients. J. Dermatol. Sci. 2021, 101, 49–57. [Google Scholar] [CrossRef]
- Aksenenko, M.; Palkina, N.; Komina, A.; Ruksha, T. MiR-92a-1-5p and miR-328-3p Are Up-Regulated in Skin of Female Pattern Hair Loss Patients. Ann. Dermatol. 2019, 31, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.J.; Lin, G.Y.; Zhu, J.Y.; Yin, G.Q.; Huang, D.; Yan, Y.Y. LncRNA-PCAT1 maintains characteristics of dermal papilla cells and promotes hair follicle regeneration by regulating miR-329/Wnt10b axis. Exp. Cell Res. 2020, 394, 112031. [Google Scholar] [CrossRef]
- Hai, E.; Han, W.; Wu, Z.; Ma, R.; Shang, F.; Wang, M.; Liang, L.; Rong, Y.; Pan, J.; Wang, Z.; et al. Chi-miR-370-3p regulates hair follicle morphogenesis of Inner Mongolian cashmere goats. G3 Genes Genomes Genet. 2021, 11, jkab091. [Google Scholar] [CrossRef]
- Lin, B.; Zhu, J.; Ye, J.; Lu, S.; Liao, M.; Meng, X. LncRNA-XIST promotes dermal papilla induced hair follicle regeneration by targeting miR-424 to activate hedgehog signaling. Cell. Signal. 2020, 72, 109623. [Google Scholar] [CrossRef]
- Kriegel, A.J.; Liu, Y.; Fang, Y.; Ding, X.; Liang, M.; Kriegel, A.J.; Liu, Y.; Fang, Y.; Ding, X.; Liang, M. The miR-29 family: Genomics, cell biology, and relevance to renal and cardiovascular injury. Physiol. Genom. 2012, 44, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Navasardyan, I.; Bonavida, B. Chapter 8—YY1 Is Involved in the Pathogenesis and Malignant Properties of Human Triple-Negative Breast Cancer (TNBC). In YY1 in the Control of the Pathogenesis and Drug Resistance of Cancer: A Critical Therapeutic Target; Bonavida, B., Ed.; Academic Press: Cambridge, MA, USA, 2021; ISBN 9780128219096. [Google Scholar]
- Lin, P.-C.; Chiu, Y.-L.; Banerjee, S.; Park, K.; Mosquera, J.M.; Giannopoulou, E.; Alves, P.; Tewari, A.K.; Gerstein, M.B.; Beltran, H.; et al. Epigenetic repression of miR-31 disrupts androgen receptor homeostasis and contributes to prostate cancer progression. Cancer Res. 2014, 73, 1232–1244. [Google Scholar] [CrossRef] [Green Version]
- Hibberts, N.A.; Howell, A.E.; Randall, V.A. Balding hair follicle dermal papilla cells contain higher levels of androgen receptors than those from non-balding scalp. J. Endocrinol. 1998, 156, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.A.; Stebbing, M.; Harrap, S.B. Polymorphism of the Androgen Receptor Gene is Associated with Male Pattern Baldness. J. Investig. Dermatol. 2001, 116, 452–455. [Google Scholar] [CrossRef]
- Kim, B.K.; Yoon, S.K. Hairless Up-Regulates Tgf-β2 Expression via Down-Regulation of miR-31 in the Skin of “Hairpoor” (HrHp) Mice. J. Cell. Physiol. 2015, 230, 2075–2085. [Google Scholar] [CrossRef]
- Yu, T.; Ma, P.; Wu, D.; Shu, Y.; Gao, W. Functions and mechanisms of microRNA-31 in human cancers. Biomed. Pharmacother. 2018, 108, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Ma, X.; Lv, C.; Sheng, X.; Li, X.; Zhao, R.; Song, Y.; Andl, T.; Plikus, M.V.; Sun, J.; et al. Stress responsive miR-31 is a major modulator of mouse intestinal stem cells during regeneration and tumorigenesis. Elife 2017, 6, e29538. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, X.; Liu, F.; Zhu, P.; Zhang, L.; Peng, Y.; Yan, X.; Li, Y.; Hua, P.; Liu, C.; et al. A stress-induced miR-31–CLOCK–ERK pathway is a key driver and therapeutic target for skin aging. Nat. Aging 2021, 1, 795–809. [Google Scholar] [CrossRef]
- Mourelatos, Z.; Paushkin, S.; Sharma, A.; Charroux, B.; Abel, L.; Rappsilber, J.; Mann, M.; Dreyfuss, G. miRNPs: A novel class of ribonucleoproteins containing numerous microRNAs. Genes Dev. 2002, 16, 720–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Sun, S.; Xu, J.; Luo, Y.; Xin, Y.; Wang, Y. MicroRNA-152 inhibits cell proliferation of osteosarcoma by directly targeting Wnt/β-catenin signaling pathway in a DKK1-dependent manner. Oncol. Rep. 2018, 40, 767–774. [Google Scholar] [CrossRef]
- Owens, P.; Han, G.; Li, A.G.; Wang, X. The Role of Smads in Skin Development. J. Investig. Dermatol. 2008, 128, 783–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taipaleenmäki, H.; Farina, N.H.; Van Wijnen, A.J.; Stein, L.; Hesse, E.; Stein, G.S.; Lian, J.B. Antagonizing miR-218-5p attenuates Wnt signaling and reduces metastatic bone disease of triple negative breast cancer cells. Oncotarget 2016, 7, 79032–79046. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Han, Y.; Li, Y.; Li, X.; Sun, T.; Chen, G. miR-218-5p inhibits the stem cell properties and invasive ability of the A2B5+CD133- subgroup of human glioma stem cells. Oncol. Rep. 2016, 35, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; He, J.; Shao, M.; Cui, B.; Peng, F.; Li, J.; Ran, Y.; Jin, D. Biomedicine & pharmacotherapy downregulation of miR-218-5p promotes invasion of oral squamous cell carcinoma cells via activation of CD44-ROCK signaling. Biomed. Pharmacother. 2018, 106, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Peng, P.; Chen, T.; Wang, Q.; Zhang, Y.; Zheng, F.; Huang, S.; Tang, Y.; Yang, C.; Ding, W.; Ren, D.; et al. Decreased miR-218–5p Levels as a Serum Biomarker in Bone Metastasis of Prostate Cancer. Oncol. Res. Treat. 2019, 42, 165–185. [Google Scholar] [CrossRef]
- Xia, C.; Jiang, H.; Ye, F.; Zhuang, Z. The Multifunction Of miR-218-5p-Cx43 Axis in Breast Cancer. OncoTargets Ther. 2019, 12, 8319–8328. [Google Scholar] [CrossRef] [Green Version]
- Son, S.W.; Lee, H.Y.; Moeng, S.; Kuh, H.J.; Choi, S.Y.; Park, J.K. Participation of microRNAs in the treatment of cancer with phytochemicals. Molecules 2020, 25, 4701. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in oligonucleotide drug delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.A.M.; Richardson, K.L.; Kalinowski, F.C.; Epis, M.R.; Horsham, J.L.; Kabir, T.D.; De Pinho, M.H.; Beveridge, D.J.; Stuart, L.M.; Wintle, L.C.; et al. Evaluation of MicroRNA Delivery In Vivo. Methods Mol. Biol. 2018, 1699, 155–178. [Google Scholar] [CrossRef]
- Bajan, S.; Hutvagner, G. RNA-Based Therapeutics: From Antisense Oligonucleotides to miRNAs. Cells 2020, 9, 137. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.F.; Weirauch, U.; Thomas, M. MicroRNA Replacement Therapy for miR-145 and miR-33a Is Efficacious in a Model of Colon Carcinoma Is Efficacious in a Model of Colon Carcinoma. Cancer Res. 2011, 71, 5214–5224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, N. An overview of viral and nonviral delivery systems for microRNA. Int. J. Pharm. Investig. 2015, 5, 179–181. [Google Scholar] [CrossRef]
- Sun, X.; Guo, Q.; Wei, W.; Robertson, S.; Yuan, Y.; Luo, X. Current Progress on MicroRNA-Based Gene Delivery in the Treatment of Osteoporosis and Osteoporotic Fracture. Int. J. Endocrinol. 2019, 2019, 6782653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasparello, J.; Lomazzi, M.; Papi, C.; Aversa, E.D.; Sansone, F.; Casnati, A.; Donofrio, G.; Gambari, R.; Finotti, A. Efficient Delivery of MicroRNA and AntimiRNA Molecules Using an Argininocalix[4]arene Macrocycle. Mol. Ther. Nucleic Acid 2019, 18, 748–763. [Google Scholar] [CrossRef] [Green Version]
- Gasparello, J.; Manicardi, A.; Casnati, A.; Corradini, R.; Gambari, R.; Finotti, A.; Sansone, F. Efficient cell penetration and delivery of peptide nucleic acids by an argininocalix[4]arene. Sci. Rep. 2019, 9, 3036. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Lee, R.J. The role of helper lipids in lipid nanoparticles (LNPs) designed for oligonucleotide delivery. Adv. Drug Deliv. Rev. 2016, 99, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Lee, L.; Paoletti, C.; Campisi, M.; Osaki, T.; Adriani, G.; Kamm, R.D.; Mattu, C.; Chiono, V. MicroRNA delivery through nanoparticles. J. Control. Release 2019, 313, 80–95. [Google Scholar] [CrossRef]
- Gokita, K.; Inoue, J.; Ishihara, H.; Kojima, K.; Inazawa, J. Therapeutic Potential of LNP-Mediated Delivery of miR-634 for Cancer Therapy. Mol. Ther. Nucleic Acids 2020, 19, 330–338. [Google Scholar] [CrossRef]
- Shi, S.; Han, L.; Gong, T.; Zhang, Z.; Sun, X. Systemic Delivery of microRNA-34a for Cancer Stem Cell Therapy. Angew. Chem. Int. Ed. Engl. 2013, 52, 3901–3905. [Google Scholar] [CrossRef]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J.A. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int. J. Pharm. 2021, 601, 120586. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Feng, Y.; Zhang, R.; Zhang, W.; Shu, Y.; Zeng, Z.; Huang, S.; Zhang, L.; Huang, B.; Wu, D.; et al. A simplified system for the effective expression and delivery of functional mature microRNAs in mammalian cells. Cancer Gene Ther. 2020, 27, 424–437. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, I.; Chatterjee, A. Recent advances in miRNA delivery systems. Methods Protoc. 2021, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, X.; Zhang, W.; Yu, L.; Wang, Y.; Deng, Z.; Liu, M.; Mo, S.; Wang, R.; Zhao, J.; et al. Trends in the biological functions and medical applications of extracellular vesicles and analogues. Acta Pharm. Sin. B 2021, 11, 2114–2135. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Disorder/Biological Process | miR Level | DKK1 Relative Expression | miR Predicted Binding Site in 3′UTR of DKK1 | Study Type | Ref. |
|---|---|---|---|---|---|---|
| miR-1-3p | Oral squamous cell carcinoma | ↓ | ↑ |  | in vitro | [82] |
| miR-9-5p | Dopaminergic neuron differentiation | N/A | ↓ |  | in vitro | [83] |
| miR-29a | Bone metabolism disorder | ↓ | ↑ |  | in vitro | [84] |
| miR-31 | Breast cancer | ↓ | ↑ |  | in vitro | [85] |
| miR-33a-5p | Esophageal cancer | ↓ | ↑ |  | in vivo in vitro | [86] |
| miR-34a | Cardiac hypertrophy | ↑ | ↓ |  | in vivo | [87] |
| miR-101-3p | Osteogenic differentiation | ↑ | ↓ |  | in vitro | [88] |
| miR-103a-3p | Osteogenic differentiation | ↑ | ↓ |  | in vitro | [89] |
| miR-107 | Osteosarcoma | ↓ | ↑ |  | in vitro | [90] |
| miR-130b-3p | Melanoma | ↑ | ↓ |  | in vitro | [91] |
| miR-146a | Ankylosing spondylitis | ↑ | ↓ |  | ex vivo | [92] |
| miR-152 | Multiple myeloma | ↓ | ↑ |  | in vitro in vivo | [93] |
| *miR-186-5p | Idiopathic pulmonary fibrosis | ↓ | ↑ |  | in vitro, ex vivo, in vivo | [94] |
| miR-203 | Lung adenocarcinoma | ↓ | ↑ |  | ex vivo, in vitro | [95] |
| Osteoporosis | ↑ |  | in vitro | [96] | ||
| miR-217 | Colon cancer | ↑ | ↓ |  | in vitro | [97] |
| Hepatocellular carcinoma | ↑ | ↓ |  | in vitro/ex vivo in vivo | [98] | |
 | ||||||
| Osteonecrosis | ↓ | ↑ |  | in vitro, ex vivo | [99] | |
| miR-291a-3p | Osteoporosis | ↓ | ↑ |  | in vitro, in vivo | [100] |
| miR-302b-3p | Multiple myeloma | ↓ | ↑ |  | in vitro, in vivo | [101] |
| miR-302e | Cardiac hypertrophy | ↑ | N/A |  | in vitro | [102] |
| Cervical cancer | ↓ | ↑ |  | in vitro, in vivo | [103] | |
| miR-335-5p | Diabetic osteoporosis | ↓ | ↑ |  | in vitro | [104] |
| miR-371/372/373 | Stem cell tumorigenesis | ↑ | ↓ |  | in vitro | [105] |
| miR-373-3p | Tongue squamous cell carcinoma | ↑ | ↓ |  | in vitro | [106] |
| miR-410 | Colorectal cancer | ↑ | ↓ |  | in vitro | [107] |
| miR-433-3p | Osteogenic differentiation | ↑ | ↓ |  | in vitro | [108] |
| miR-488 | Fracture | ↓ | ↑ |  | in vitro | [109] |
| miR-493-3p | Gastric cancer | ↑ | ↓ |  | in vitro | [110] |
| miR-522 | Hepatocellular carcinoma | ↑ | ↓ |  | in vitro | [111] |
| miR-523-3p | Retinoblastoma | ↑ | ↓ |  | in vitro, ex vivo | [112] |
| miR-613 | Rheumatoid arthritis | ↓ | ↑ |  | in vitro | [113] |
| miR-3064-3p | Cementoblast differentiation | ↓ | ↑ |  | in vitro | [114] |
| miR-6783-3p | Lung adenocarcinoma | ↑ | ↓ |  | in vitro, in vivo | [115] |
| miR-6807-3p | Lung adenocarcinoma | ↑ | ↓ |  | in vitro, in vivo | [116] |
| miR-BART10-3p | Gastric carcinoma | ↑ | ↓ |  | in vitro | [117] |
| miR-BART22 | Gastric carcinoma | ↑ | ↓ |  | in vitro | [118] |
| # | Study Title | miRNA | Influence on Hair Growth | Study Model | Type of Administration | Biological Function | Target Protein/Gene | Finding | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Differential expression of miR-let7a in hair follicle cycle of Liaoning cashmere goats and identification of its targets | miR-let7a | Regulatory | Goat skin samples, cell culture | in vitro transfection | Regulating FGF, IGF, C-Myc signaling pathways | IGF-1R, C-Myc, FGF5 | miR-let7a regulates HF development via targeting C-myc, IGF-1R, and FGF5 | [157] |
| 2 | Let-7b regulates alpaca hair growth by downregulating ectodysplasin A | miR-let-7b | Regulatory | Alpaca, cell culture | in vitro transfection | Inhibiting EDA signaling | EDA | miR-let-7b regulates hair growth via targeting EDA | [158] |
| 3 | Identification of microRNA-21 target genes associated with hair follicle development in sheep | miR-21 | Regulatory | Sheep | in vitro transfection | Regulating CNKSR2-MAPK, KLF3, TNPO1-CCR2 signaling pathways | CNKSR2, KLF3, TNPO1 | miR-21 targets CNKSR2, KLF3, and TNPO1 that might play a regulatory role in HF development | [159] |
| 4 | EZH2-mediated inhibition of microRNA-22 promotes differentiation of hair follicle stem cells by elevating STK40 expression | miR-22 | Negative | Mouse, cell culture | in vitro transfection | Inhibiting MEF2-ALP signaling | STK40 | miR-22 targets STK40 and inhibits the MEF2-ALP signaling pathway, and thus impedes the proliferation and differentiation of HFSC | [160] |
| 5 | Post-transcriptional regulation of keratinocyte progenitor cell expansion, differentiation and hair follicle regression by miR-22 | Negative | Mouse, cell culture | in vitro transfection | Activating apoptotic signaling pathways | DLX3, FOXN1, HOXC13 | miR-22 is a critical post-transcriptional regulator of the hair cycle and its activation results in hair loss | [161] | |
| 6 | miR-24 affects hair follicle morphogenesis targeting Tcf-3 | miR-24 | Negative | Mouse, cell culture | in vitro transfection | Regulating Wnt/β-catenin signaling | TCF-3 | miR-24 is expressed in the HF and it directly targets TCF-3, a regulator of the hair keratinocyte stemness | [162] |
| 7 | miR-24 controls the regenerative competence of hair follicle progenitors by targeting Plk3 | Negative | Mouse, cell culture | in vitro transfection | Regulating PLK3-CCNE1 signaling | PLK3 | miR-24 by targeting PLK3 limits the intrinsic growth competence of HF progenitor | [163] | |
| 8 | miR-29 modulates Wnt signaling in human osteoblasts through a positive feedback loop | miR-29a | N/A | Cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | DKK1, SFRP2, KRM2 | miR-29a potentiates Wnt signaling via downregulation of the key antagonists of Wnt signaling, DKK1, KRM2, and SFRP2, which contributes to a gene expression program important for osteoblast differentiation | [164] |
| 9 | miR-29 suppression of osteonectin in osteoblasts: regulation during differentiation and by canonical Wnt signaling | N/A | Cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | DKK1 | miR-29a and Wnt signaling antagonist DKK1 expressions are negatively correlated | [165] | |
| 10 | Protective effects of miR-29a on diabetic glomerular dysfunction by modulation of DKK1/Wnt/β-catenin signaling | N/A | Mouse, cell culture | Tail vain injection, in vitro transfection | Activating Wnt/β-catenin signaling | DKK1 | miR-29a in diabetic mice attenuates the expression of DKK1 which inactivates Wnt/β-catenin signaling | [166] | |
| 11 | miR-29a modulates tumor necrosis factor-α-induced osteogenic inhibition by targeting Wnt antagonists | N/A | Cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | DKK1, GSK-3β | miR-29a plays an important role in regulating TNF-α-mediated osteogenic inhibition partly by targeting DKK1 and GSK-3β | [84] | |
| 12 | LncRNA H19 overexpression activates Wnt signaling to maintain the hair follicle regeneration potential of dermal papilla cells | Positive | Mouse, cell culture | Subcutaneous injection, in vitro transfection | Activating Wnt/β-catenin signaling | DKK1, SFRP2, KRM2 | lncRNA H19 induces miR-29a which activates Wnt signaling and induces HF regeneration in vitro and in vivo | [167] | |
| 13 | miR-29a/b1 inhibits hair follicle stem cell lineage progression by spatiotemporally suppressing Wnt and BMP signaling | miR-29a/b1 | Negative | Mouse, Cell culture | in vitro transfection | Inhibiting Wnt/β-catenin, BMP signaling pathways | LRP6, CTNNB1, BMPR1a, CCNA2 | miR-29a/b1 overexpression causes hair loss by targeting Wnt and BMP | [168] |
| 14 | Chi-miR-30b-5p inhibits dermal papilla cells proliferation by targeting CaMKIIδ gene in cashmere goat | miR-30b-5p | Negative | Goat, cell culture | in vitro transfection | N/A | CaMKIIδ | Chi-miR-30b-5p targets CaMKIIδ and inhibits the proliferation of DP cells | [74] |
| 15 | miR-31-5p promotes proliferation and inhibits apoptosis of goat hair follicle stem cells by targeting RASA1/MAP3K1 pathway | miR-31 | Positive | Cell culture | in vitro transfection | Activating MAPK signaling | RASA1 | miR-31-5p suppresses apoptosis/promotes goat HFSC proliferation via inhibiting RASA1 and upregulating MAP3K1 | [169] |
| 16 | Micro-RNA-31 controls hair cycle-associated changes in gene expression programs of the skin and hair follicle | Optimal balance | Mouse, cell culture | in vitro transfection | Balancing FGF, BMP, Wnt/β-catenin signaling | FGF10, SOST, BAMBI | miR-31 is involved in maintaining an optimal balance of gene expression in HFs via targeting a number of key molecules; thus, it has a positive influence on HF proper growth and development | [170] | |
| 17 | MiR-31 promotes mammary stem cell expansion and breast tumorigenesis by suppressing Wnt signaling antagonists | N/A | Mouse, cell line | in vitro transfection | Activating Wnt/β-catenin, Inhibiting TGF-β, PRLR/STAT5 signaling pathways | DKK1, AXIN1, GSK-3β | miR-31 promotes Wnt/β-catenin signaling by targeting DKK1, thus, miR-31 represents a key regulator of breast tumorigenesis | [85] | |
| 18 | Down-regulation of microRNA-31-5p inhibits proliferation and invasion of osteosarcoma cells through Wnt/β-catenin signaling pathway by enhancing AXIN1 | N/A | Tumor samples cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | AXIN1 | miR-31-5p targets AXIN1, and via inhibiting the transcription of AXIN1, it activates Wnt/β-catenin signaling in OS cells | [171] | |
| 19 | The major miR-31 target genes STK40 and LATS2 and their implications in the regulation of keratinocyte growth and hair differentiation | Negative | Mouse, cell culture | in vitro transfection | Promoting aberrant keratinocyte growth | STK40, LATS2 | miR-31 targets a large number of gene expressions, including STK40 and LATS2, that play a role in keratinocyte growth and HF biology | [172] | |
| 20 | Downregulation of LINC00707 promotes osteogenic differentiation of human bone marrow-derived mesenchymal stem cells by regulating DKK1 via targeting miR-103a-3p | miR-103a-3p | N/A | Cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | DKK1 | LINC00707 regulates the expression of DKK1 by targeting miR-103a-3p | [89] |
| 21 | MicroRNAs-103/107 regulate autophagy in the epidermis | miR-103/107 | Positive | Mouse, cell culture | Subcutaneous injection | Activating PKC signaling | PLD1, PLD2 | miR-103/107 may play an important role in maintaining the stemness of HFSCs and its levels may decrease with age | [173] |
| 22 | In vitro effect of microRNA-107 targeting Dkk-1 by regulation of Wnt/β-catenin signaling pathway in osteosarcoma | miR-107 | N/A | OS tissue, cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | DKK1 | miR-107 inhibits the occurrence and development of OS by downregulating DKK1 through the Wnt/β-catenin signaling pathway | [90] |
| 23 | miR-124 promotes neural differentiation in mouse bulge stem cells by repressing Ptbp1 and Sox9 | miR-124 | Positive | Mouse, cell culture | in vitro transfection | Promoting HFSCs neuronal differentiation | SOX9, PTBP1 | miR-124 promotes HFSCs neuronal differentiation via targeting SOX9 and PTBP1 | [174] |
| 24 | miR-125/CDK2 axis in cochlear progenitor cell proliferation | miR-125 | Negative | Rat, cell culture | in vitro transfection | Inhibiting CDK pathway | CDK2 | miR-125 inhibits the proliferation of CPC by downregulating CDK2 | [175] |
| 25 | Chi-miR-130b-3p regulates Inner Mongolia cashmere goat skin hair follicles in fetuses by targeting Wnt family member 10A | miR-130b-3p | Negative | Cell culture | in vitro transfection | Inhibiting Wnt/β-catenin signaling | Wnt10a | The study suggests that Wnt10a is a target gene of chi-miR-130b-3p. Thus, chi-miR-130b-3p may regulate epithelial cell and dermal fibroblast proliferation | [176] |
| 26 | Expression profiling and functional characterization of miR-26a and miR-130a in regulating Zhongwei goat hair development via the TGF-β /SMAD pathway | miR-130a miR-26a | Negative (miR-130a) Positive (miR-26) | Goat, cell culture | in vitro transfection | Regulating TGF-β/SMAD pathway | SMAD5, SMAD1 | miR-26a and miR-130a regulate the HF development and growth through implication in TGF- β/SMAD signaling pathway | [177] |
| 27 | miRNA microarray profiling in patients with androgenic alopecia and the effects of miR-133b on hair growth | miR-133b | Negative | AGA scalp, cell culture | in vitro transfection | Inhibiting Wnt/β-catenin signaling | ALP, β-catenin, versican | miR-133b is abnormally highly expressed in patients with AGA. miR-133b may inactivate the Wnt/β-catenin pathway, and thus regulate hair growth | [150] |
| 28 | miR-140-5p in small extracellular vesicles from human papilla cells stimulates hair growth by promoting proliferation of outer root sheath and hair matrix cells | miR-140-5p | Positive | Cell culture | in vitro transfection | Inhibiting BMP signaling | BMP2 | miR-140-5p plays a critical role in hair growth and cell proliferation and it represents a therapeutic target for alopecia | [178] |
| 29 | Preliminary study on microR-148a and microR-10a in dermal papilla cells of Hu sheep | miR-148amiR-10a | N/A | Cell culture | in vitro transfection | Inhibiting BMP signaling, regulating TGF-β/Smads signaling | BMP7 | miR-148a and miR-10a inhibits the proliferation of Hu sheep dermal papilla cells | [179] |
| 30 | MicroRNA-148b promotes proliferation of hair follicle cells by targeting NFAT5 | miR-148b | Positive | Cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | NFAT5, Wnt10b | miR-148b activates the Wnt/β-catenin signaling pathway by targeting NFAT5 and promotes the proliferation of human HF cells | [180] |
| 31 | miR-149-5p regulates goat hair follicle stem cell proliferation and apoptosis by targeting the CMTM3/AR axis during superior-quality brush hair formation | miR-149-5p | Positive | Goat, cell culture | in vitro transfection | Regulating AR transcriptional activity | CMTM3 | miR-149-5p suppresses HF stem cell apoptosis by inhibiting CMTM3 and regulates the high-quality hair formation in goats | [181] |
| 32 | Downregulation of MicroRNA-152 contributes to high expression of DKK1 in multiple myeloma | miR-152 | N/A | Mouse, cell culture | Femur injection, in vitro transfection | Activating Wnt/β-catenin signaling | DKK1 | miR-152 blocked DKK1 transcriptional activity by binding to the 3′UTR of DKK1 mRNA. miR-152 is a master regulator in the pathobiology of MM via targeting DKK1 | [93] |
| 33 | The effect of the microRNA-183 family on hair cell-specific markers of human bone marrow-derived mesenchymal stem cells | miR-182 | Positive | Cell culture | in vitro transfection | N/A | N/A | miR-182 plays a key role in hair cell differentiation | [182] |
| 34 | microRNA-183 is involved in the differentiation and regeneration of Notch signaling-prohibited hair cells from mouse cochlea | miR-183 | Positive | Mouse, cell culture, organ culture | in vitro transfection | Regulating Notch signaling | N/A | Inhibition of the Notch signaling pathway induces miR-183 expression, which participates in hair cell differentiation and regeneration | [183] |
| 35 | microRNA-183 is essential for hair cell regeneration after neomycin injury in zebrafish | miR-183miR-182 miR-96 | Positive | Zebrafish | Micro injection | N/A | N/A | miR-183 cluster has a crucial role in the regeneration of hair cells in zebrafish larvae and it can be a novel target for hair cell regeneration | [184] |
| 36 | The intragenic mRNA-microRNA regulatory network during telogen–anagen hair follicle transition in the cashmere goat | miR-195 | Positive | Goat | N/A | Regulating Wnt/β-catenin signaling | SMAD2 FZD6 | miR-195 plays a crucial role in the regulation of HF initiation | [185] |
| 37 | miR-195-5p regulates hair follicle inductivity of dermal papilla cells by suppressing Wnt/β-catenin activation | miR-195-5p | Negative | Scalp specimen, cell culture | in vitro transfection | Inhibiting Wnt/β-catenin signaling | LRP6 | miR-195-5p inhibits Wnt/β-catenin activation by targeting LRP6 | [186] |
| 38 | The microRNA-200 family coordinately regulates cell adhesion and proliferation in hair morphogenesis | miR-200 | Positive | Mouse, cell culture | in vitro transfection | Regulating Hippo/Yap signaling | CCNG2, CFL2, SNAI2, SEC23A, YWHAB, FAT1, PTK2, CDC42, CRK, ROCK2, WASL, ACTN1 | miR-200 family has a critical function in mammalian skin development via regulation of cell adhesion and orientation in the hair germ | [187] |
| 39 | miR-203 is involved in osteoporosis by regulating DKK1 and inhibiting osteogenic differentiation of MSCs | miR-203 | N/A | Human, cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | DKK1 | miR-203 by regulating DKK1 expression promotes the differentiation of rat MSCs into osteoblast-like cells | [96] |
| 40 | Overexpression of miR-203 increases the sensitivity of NSCLC A549/H460 cell lines to cisplatin by targeting Dickkopf-1 | N/A | Human advanced lung adenocarcinoma tissue, cell culture, mouse | Subcutaneous injection, in vitro transfection | Activating Wnt/β-catenin signaling | DKK1 | miR-203 by targeting the 3’UTR of DKK1 increases cisplatin sensitivity in A549/H460 cell lines | [95] | |
| 41 | RBM28, a protein deficient in ANE syndrome, regulates hair follicle growth via miR-203 and p63 | Positive | Cell culture, organ culture | in vitro transfection | N/A | P63 | RBM28 directly regulates hair growth by ameliorating the expression or activity of miR-203 that inhibits transcription factor p63 | [188] | |
| 42 | Expression of miRNA-203 and its target gene in hair follicle cycle development of Cashmere goat | Regulatory | Goat, cell culture | in vitro transfection | N/A | DDOST, NAE1 | miR-203 significantly downregulates the expression of DDOST and NAE1 and regulates the HF development in Cashmere goats | [189] | |
| 43 | miR-203a-3p promotes loureirin A-induced hair follicle stem cells differentiation by targeting Smad1 | Positive | Rat, cell culture | in vitro transfection | Inhibiting BMP signaling | SMAD1 | miR-203a-3p inhibits SMAD1 and promotes HFSCs differentiation | [190] | |
| 44 | The functions of ocu-miR-205 in regulating hair follicle development in Rex rabbits | miR-205 | Negative | Rabbit | Intradermal injection | Regulating PI3K/Akt, Wnt/β-catenin, Notch, BMP signaling pathways | NOG | ocu-miR-205 promotes the apoptosis of DP cells via inhibiting the expression of genes involved in the PI3K/Akt, Wnt, and Notch signaling pathways, and activates the BMP signaling pathway | [191] |
| 45 | MiR-214 regulates the human hair follicle stem cell proliferation and differentiation by targeting EZH2 and Wnt/β-catenin signaling way in vitro | miR-214 | Negative | Scalp tissue, cell culture | in vitro transfection | Regulating Wnt/β-catenin signaling | EZH2 | Overexpression of miR-214 decreases the expression of EZH2, β-catenin, and TCF-4, while downregulation of miR-214 promotes the proliferation and differentiation of HFSCs | [192] |
| 46 | MicroRNA-214 controls skin and hair follicle development by modulating the activity of the Wnt pathway | Negative | Mice, cell culture | in vitro transfection | Inhibiting Wnt/β-catenin signaling | β-catenin | miR-214 regulates the Wnt signaling pathway and β-catenin expression in the developing and postnatal skin and HFs | [75] | |
| 47 | DNMT1-mediated methylation inhibits microRNA-214-3p and promotes hair follicle stem cell differentiate into adipogenic lineages | Negative | Scalp tissue, cell culture | in vitro transfection | N/A | DNMT1, MAPK1 | Expression of DNMT1, MAPK1, and miR-214-3p in HFSc are negatively correlated. DNMT1 promotes adipogenesis of HFSc by mediating miR-214-3p/MAPK1/p-ERK1/2 axis | [193] | |
| 48 | Dermal exosomes containing miR-218-5p promote hair regeneration by regulating β-catenin signaling | miR-218-5p | Positive | Mouse, cell culture | Subcutaneous injection | Activating Wnt/β-catenin signaling | SFRP2 | miR-218-5p ameliorates HF development by downregulating SFRP2 and promoting β-catenin. miR-218-5p-overexpressed exosomes lead to the onset of anagen | [77] |
| 49 | miR-218-5p regulates skin and hair follicle development through Wnt/β-catenin signaling pathway by targeting SFRP2 | Positive | Rabbit, cell culture | in vitro transfection | Activating Wnt/β-catenin signaling | SFRP2 | miR-218-5p enhances the Wnt signaling pathway by targeting SFRP2 and induces HF development | [76] | |
| 50 | Osteogenic differentiation of fibroblast-like synovial cells in rheumatoid arthritis is induced by microRNA-218 through a ROBO/Slit pathway | N/A | Synovial tissues | in vitro transfection | Activating Wnt/β-catenin signaling | ROBO1 | miR-218 promotes the osteogenic differentiation of rheumatoid arthritis fibroblast-like synovial cells by targeting ROBO1 and suppressing DKK1 | [153] | |
| 51 | Defining microRNA signatures of hair follicular stem and progenitor cells in healthy and androgenic alopecia patients | miR-324-3p | Positive | Scalp sample, cell culture | in vitro transfection | Regulating MAPK, TGF-β signaling pathways | REL A, HSP A2, MAPK1/3, TGF-β3 | miR-324-3p regulates pathways implicated in HF growth and development | [194] |
| 52 | MiR-92a-1-5p and miR-328-3p are upregulated in skin of female pattern hair loss patients | miR-328-3p miR-92a-1-5p | Negative | Human | N/A | Implicating in multiple signaling pathways that are involved in FPHL | N/A | miR-92a-1-5p and miR-328-3p are involved in many signaling pathways and have a negative effect on FPHL | [195] |
| 53 | LncRNA-PCAT1 maintains characteristics of dermal papilla cells and promotes hair follicle regeneration by regulating miR-329/Wnt10b axis | miR-329 | Negative | Scalp sample, mouse, cell culture | in vitro transfection | Inhibiting Wnt/β-catenin signaling | Wnt10b, ALP, BMP2, Versican, NCAM | PCAT1 promotes DP cells’ proliferation via activating Wnt/β-catenin signaling, while miR-329 negatively affects DP cells | [196] |
| 54 | Chi-miR-370-3p regulates hair follicle morphogenesis of Inner Mongolian cashmere goats | miR-370-3p | Positive | Goat, cell culture | in vitro transfection | Inhibiting TGF, FGF signaling | TGF-βR2, FGFR2 | Chi-miR-370-3p inhibits the proliferation of epithelial cells and dermal fibroblasts via targeting FGFR2 and TGF-βR2, as well as induces HF morphog1enesis | [197] |
| 55 | Differential expression analysis of balding and nonbalding dermal papilla microRNAs in male pattern baldness with a microRNA amplification profiling method | miR-410 miR-221 miR-125b miR-106a | Negative | Human, cell culture | N/A | N/A | N/A | miR-221, miR-125b, miR-106a, and miR-410 are significantly upregulated in balding papilla cells and they may participate in the pathogenesis of male pattern baldness | [151] |
| 56 | LncRNA-XIST promotes dermal papilla induced hair follicle regeneration by targeting miR-424 to activate hedgehog signaling | miR-424 | Negative | Mouse, cell culture | in vitro transfection | Inhibiting SHH signaling | ALP, Versican, NCAM, GLI1/2 | miR-424 is sponged by XIST, which promotes SHH signaling and facilitates HF regeneration | [198] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papukashvili, D.; Rcheulishvili, N.; Liu, C.; Xie, F.; Tyagi, D.; He, Y.; Wang, P.G. Perspectives on miRNAs Targeting DKK1 for Developing Hair Regeneration Therapy. Cells 2021, 10, 2957. https://doi.org/10.3390/cells10112957
Papukashvili D, Rcheulishvili N, Liu C, Xie F, Tyagi D, He Y, Wang PG. Perspectives on miRNAs Targeting DKK1 for Developing Hair Regeneration Therapy. Cells. 2021; 10(11):2957. https://doi.org/10.3390/cells10112957
Chicago/Turabian StylePapukashvili, Dimitri, Nino Rcheulishvili, Cong Liu, Fengfei Xie, Deependra Tyagi, Yunjiao He, and Peng George Wang. 2021. "Perspectives on miRNAs Targeting DKK1 for Developing Hair Regeneration Therapy" Cells 10, no. 11: 2957. https://doi.org/10.3390/cells10112957
APA StylePapukashvili, D., Rcheulishvili, N., Liu, C., Xie, F., Tyagi, D., He, Y., & Wang, P. G. (2021). Perspectives on miRNAs Targeting DKK1 for Developing Hair Regeneration Therapy. Cells, 10(11), 2957. https://doi.org/10.3390/cells10112957