
Dynamic Role of Phospholipases A2 in Health and Diseases in the Central Nervous System

 , , ,
, , ,  and
and 
Abstract
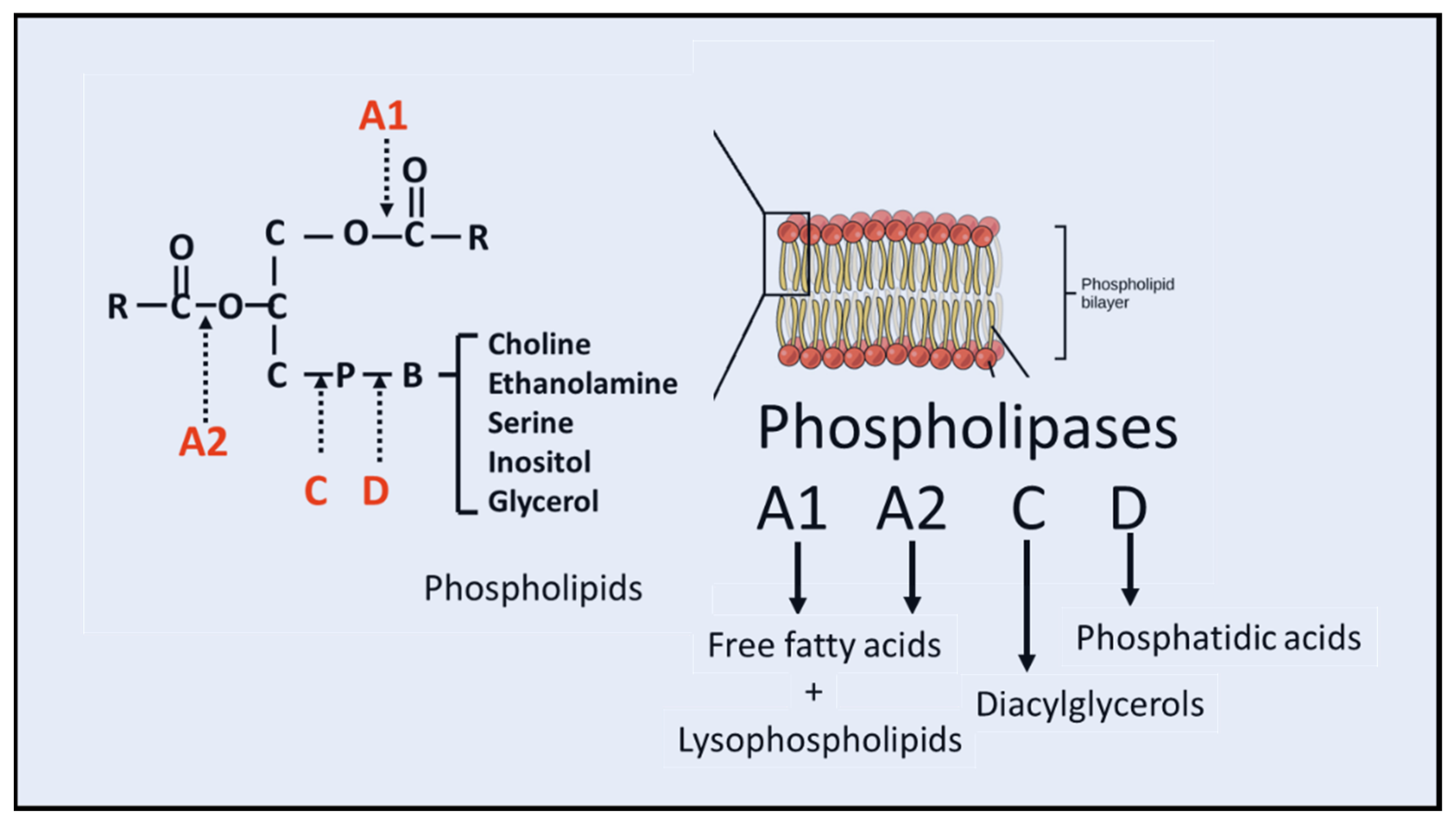
:1. Introduction
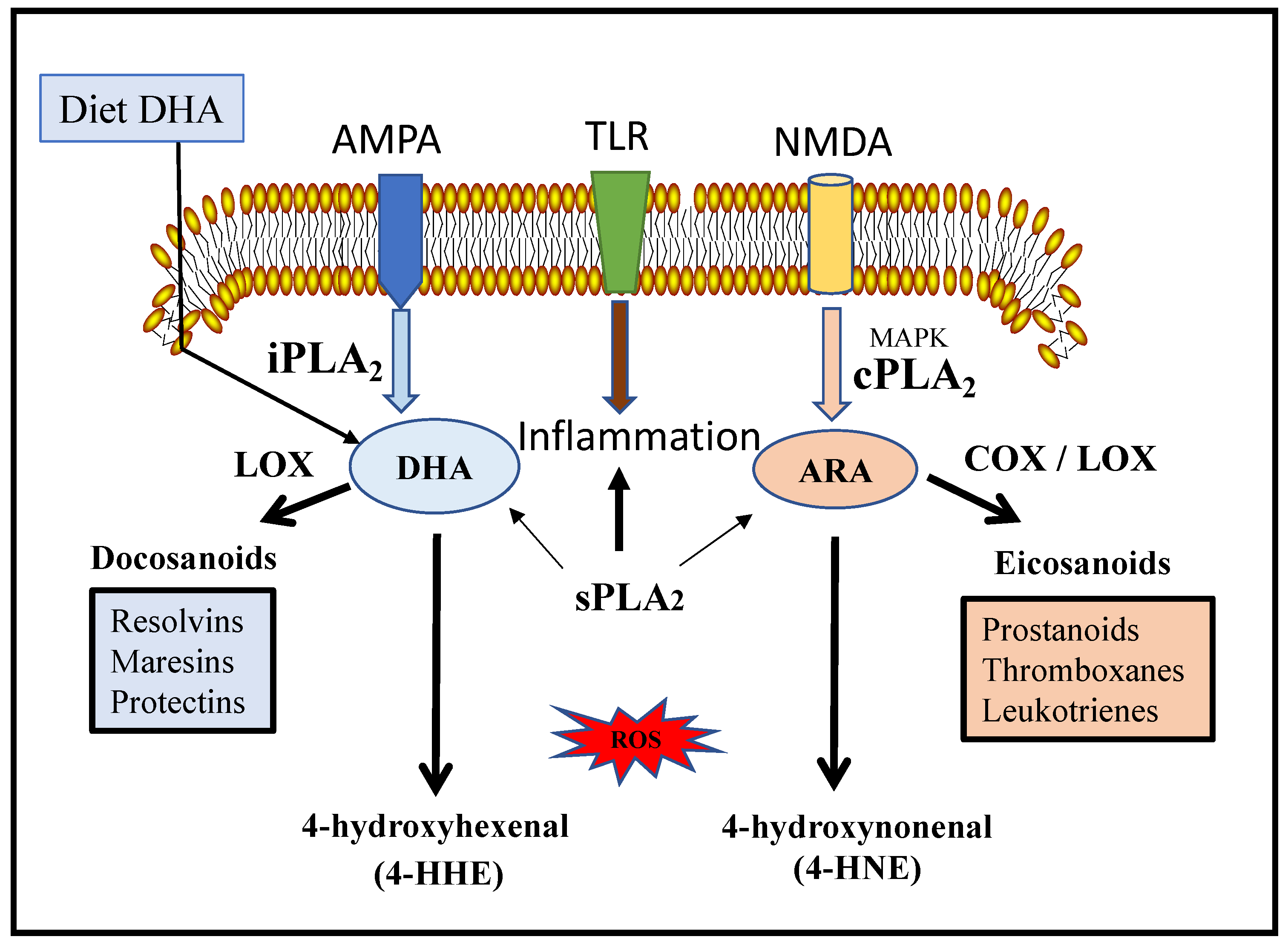
2. Structure and Function of cPLA2
3. cPLA2 in Cerebral Ischemia, Spinal Cord and Traumatic Brain Injury
3.1. Cerebral Ischemia
3.2. Traumatic Brain Injury (TBI)
3.3. Spinal Cord Injury (SCI)
4. cPLA2 in Other Neurodegenerative Diseases
Alzheimer’s Disease (AD)
5. Structure and Function of iPLA2
5.1. GVIA PLA2 (iPLA2β)
5.2. iPLA2γ
6. iPLA2 and Neurological Diseases
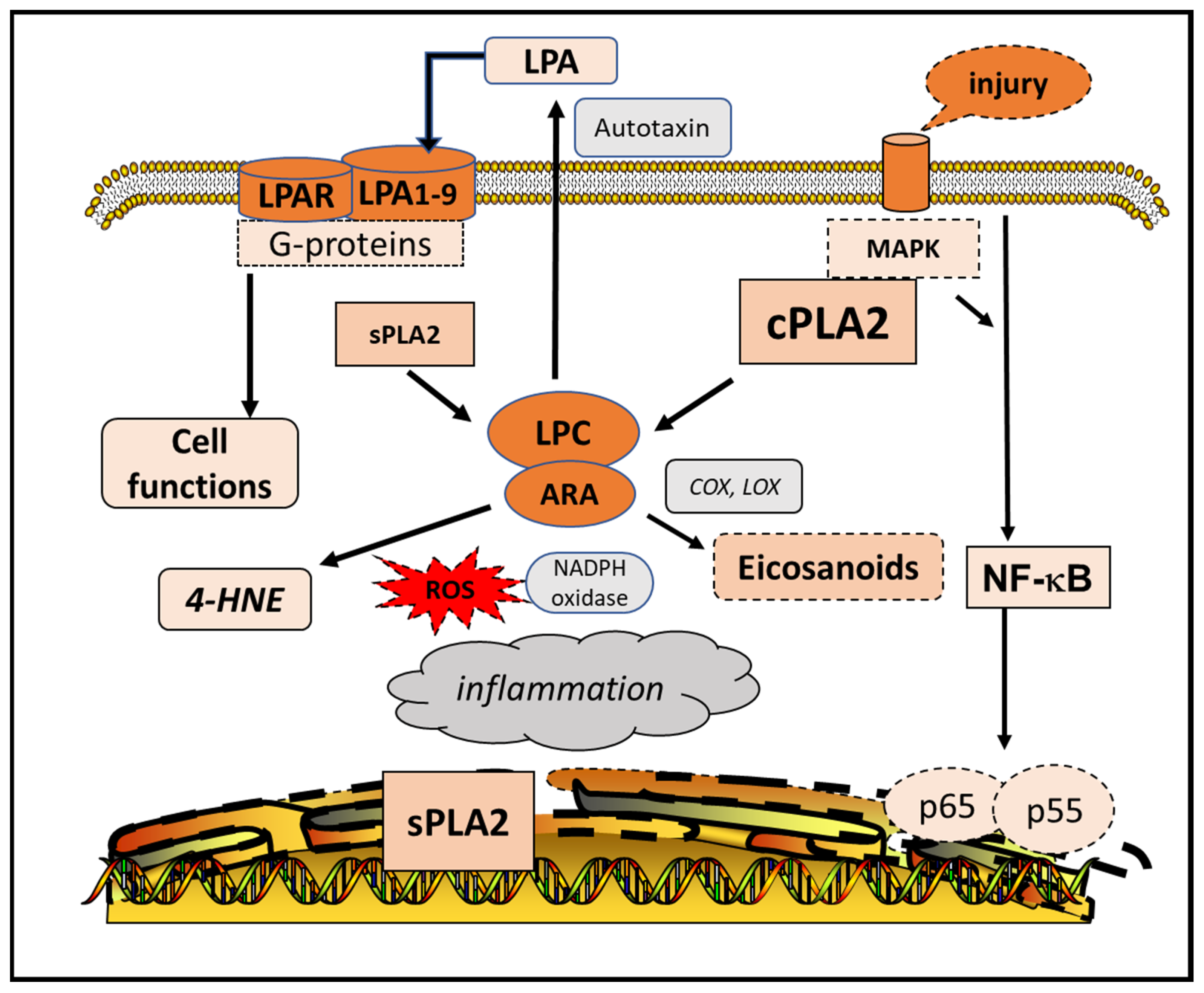
7. Structure and Function of sPLA2
8. sPLA2 in Extracellular Vesicles
9. sPLA2-IIA in Neurodegenerative Diseases
9.1. Alzheimer’s Diseases
9.2. Cerebral Ischemia
9.3. Spinal Cord Injury
10. PLA2 and Lysophospholipids
11. PLA2-Mediated Production of Oxylipins and Lipid Peroxidation Products
12. Peroxidation of ARA and DHA
13. Phospholipase Products through Lipidomics and Advanced Mass Spectrometry Analysis
13.1. Phospholipid Species
13.2. Oxylipins
14. Summary and Future Directions
Funding
Conflicts of Interest
References
- Kita, Y.; Shindou, H.; Shimizu, T. Cytosolic phospholipase A2 and lysophospholipid acyltransferases. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2018, 1864, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2006, 1761, 1246–1259. [Google Scholar] [CrossRef]
- Burke, J.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, W.-Y.; Farooqui, T.; Kokotos, G.; Farooqui, A.A. Synthetic and Natural Inhibitors of Phospholipases A2: Their Importance for Understanding and Treatment of Neurological Disorders. ACS Chem. Neurosci. 2015, 6, 814–831. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Chuang, D.Y.; Zong, Y.; Jiang, J.; Lee, J.C.M.; Gu, Z.; Simonyi, A. Role of Cytosolic Phospholipase A2 in Oxidative and Inflammatory Signaling Pathways in Different Cell Types in the Central Nervous System. Mol. Neurobiol. 2014, 50, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.Y.; Shelat, P.B.; Jensen, M.B.; He, Y.; Sun, A.Y.; Simonyi, A. Phospholipases A2 and Inflammatory Responses in the Central Nervous System. NeuroMol. Med. 2009, 12, 133–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, E.A.; Cao, J.; Hsu, Y.-H.; Magrioti, V.; Kokotos, G. Phospholipase A2 Enzymes: Physical Structure, Biological Function, Disease Implication, Chemical Inhibition, and Therapeutic Intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [Green Version]
- Niknami, M.; Patel, M.; Witting, P.K.; Dong, Q. Molecules in focus: Cytosolic phospholipase A2-α. Int. J. Biochem. Cell Biol. 2009, 41, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, C.; Jones, J.W.; Hegdekar, N.; Thayer, J.A.; Kumar, A.; Faden, A.I.; Kane, M.A.; Lipinski, M.M. PLA2G4A/cPLA2-mediated lysosomal membrane damage leads to inhibition of autophagy and neurodegeneration after brain trauma. Autophagy 2019, 16, 466–485. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, H.; Moriyama, Y.; Makiyama, T.; Emori, S.; Yamashita, H.; Yamazaki, R.; Murayama, T. Lactosylceramide Interacts with and Activates Cytosolic Phospholipase A2α. J. Biol. Chem. 2013, 288, 23264–23272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.-C.; Gutiérrez-Vázquez, C.; Rothhammer, V.; Mayo, L.; Wheeler, M.A.; Tjon, E.C.; Zandee, S.; Blain, M.; de Lima, K.A.; Takenaka, M.C.; et al. Metabolic Control of Astrocyte Pathogenic Activity via cPLA2-MAVS. Cell 2019, 179, 1483–1498.e22. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Balram, A.; Li, W. Convergence: Lactosylceramide-Centric Signaling Pathways Induce Inflammation, Oxidative Stress, and Other Phenotypic Outcomes. Int. J. Mol. Sci. 2021, 22, 1816. [Google Scholar] [CrossRef] [PubMed]
- Shelat, P.B.; Chalimoniuk, M.; Wang, J.-H.; Strosznajder, J.B.; Lee, J.C.; Sun, A.Y.; Simonyi, A.; Sun, G.Y. Amyloid beta peptide and NMDA induce ROS from NADPH oxidase and AA release from cytosolic phospholipase A2in cortical neurons. J. Neurochem. 2008, 106, 45–55. [Google Scholar] [CrossRef]
- Malada-Edelstein, Y.F.; Hadad, N.; Levy, R. Regulatory role of cytosolic phospholipase A2 alpha in the induction of CD40 in microglia. J. Neuroinflamm. 2017, 14, 33. [Google Scholar] [CrossRef] [Green Version]
- Chuang, D.Y.; Cui, J.; Simonyi, A.; Engel, V.A.; Chen, S.; Fritsche, K.L.; Thomas, A.L.; Applequist, W.L.; Folk, W.R.; Lubahn, D.B.; et al. Dietary Sutherlandia and Elderberry Mitigate Cerebral Ischemia-Induced Neuronal Damage and Attenuate p47phox and Phospho-ERK1/2 Expression in Microglial Cells. ASN Neuro 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Li, R.; Liu, P.N.; Geng, X.; Mooney, B.P.; Chen, C.; Cheng, J.; Fritsche, K.L.; Beversdorf, D.Q.; Lee, J.C.; et al. Quantitative Proteomics Reveals Docosahexaenoic Acid-Mediated Neuroprotective Effects in Lipopolysaccharide-Stimulated Microglial Cells. J. Proteome Res. 2020, 19, 2236–2246. [Google Scholar] [CrossRef] [PubMed]
- Riendeau, D.; Guay, J.; Weech, P.; Laliberté, F.; Yergey, J.; Li, C.; Desmarais, S.; Perrier, H.; Liu, S.; Nicoll-Griffith, D. Arachidonyl trifluoromethyl ketone, a potent inhibitor of 85-kDa phospholipase A2, blocks production of arachidonate and 12-hydroxyeicosatetraenoic acid by calcium ionophore-challenged platelets. J. Biol. Chem. 1994, 269, 15619–15624. [Google Scholar] [CrossRef]
- Farooqui, A.A.; Ong, W.Y.; Horrocks, L.A. Inhibitors of brain phospholipase A2 activity: Their neuropharmacological effects and thera-peutic importance for the treatment of neurologic disorders. Pharmacol. Rev. 2006, 58, 591–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szaingurten-Solodkin, I.; Hadad, N.; Levy, R. Regulatory role of cytosolic phospholipase A2alpha in NADPH oxidase activity and in in-ducible nitric oxide synthase induction by aggregated Abeta1-42 in microglia. Glia 2009, 57, 1727–1740. [Google Scholar] [CrossRef] [PubMed]
- Anwar, K.; Voloshyna, I.; Littlefield, M.J.; Carsons, S.E.; Wirkowski, P.A.; Jaber, N.L.; Sohn, A.; Eapen, S.; Reiss, A.B. COX-2 Inhibition and Inhibition of Cytosolic Phospholipase A2 Increase CD36 Expression and Foam Cell Formation in THP-1 Cells. Lipids 2010, 46, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.M.; Dwyer-Nield, L.D.; Hurteau, G.J.; Keith, R.L.; O’Leary, E.; You, M.; Bonventre, J.V.; Nemenoff, R.A.; Malkinson, A.M. Decreased lung tumorigenesis in mice genetically deficient in cytosolic phospholipase A2. Carcinogenesis 2004, 25, 1517–1524. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, K.; Miyazaki, A.; Nabe, T.; Fushimi, H.; Iriyama, N.; Kanai, S.; Sato, T.; Uozumi, N.; Shimizu, T.; Akiba, S. Group IVA phospholipase A 2 participates in the progression of hepatic fibrosis. FASEB J. 2012, 26, 4111–4121. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Wei, X.; Du, P.; Zhao, H.; Liu, A.; Chen, Y. beta-Arrestin-2-ERK1/2 cPLA2alpha axis mediates TLR4 signaling to influence eicosanoid induction in ischemic brain. FASEB J. 2019, 33, 6584–6595. [Google Scholar] [CrossRef]
- Street, I.P.; Lin, H.K.; Laliberte, F.; Ghomashchi, F.; Wang, Z.; Perrier, H.; Tremblay, N.M.; Huang, Z.; Weech, P.K.; Gelb, M.H. Slow- and tight-binding inhibitors of the 85-kDa human phospholipase A2. Biochemistry 1993, 32, 5935–5940. [Google Scholar] [CrossRef] [PubMed]
- Schanstra, J.P.; Luong, T.T.; Makridakis, M.; Van Linthout, S.; Lygirou, V.; Latosinska, A.; Alesutan, I.; Boehme, B.; Schelski, N.; Von Lewinski, D.; et al. Systems biology identifies cytosolic PLA2 as a target in vascular calcification treatment. JCI Insight 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Sharma, S.; Kaushik, G.; Avti, P.K.; Pandey, S.; Sarma, P.; Medhi, B.; Khanduja, K.L. Therapeutic potential of arachidonyl trifluromethyl ketone, a cytosolic phospholipaseA2 IVA specific inhibitor, in cigarette smoke condensate-induced pathological conditions in alveolar type I & II epithelial cells. Toxicol. In Vitro 2018, 54, 215–223. [Google Scholar] [CrossRef]
- Rodriguez de Turco, E.B.; Belayev, L.; Liu, Y.; Busto, R.; Parkins, N.; Bazan, N.G.; Ginsberg, M.D. Systemic fatty acid responses to transient focal cerebral ischemia: Influence of neuroprotectant therapy with human albumin. J. Neurochem. 2002, 83, 515–524. [Google Scholar] [CrossRef]
- Sun, G.Y.; MacQuarrie, R.A. Deacylation-Reacylation of Arachidonoyl Groups in Cerebral Phospholipids. Ann. N. Y. Acad. Sci. 1989, 559, 37–55. [Google Scholar] [CrossRef]
- Golovko, S.A.; Golovko, M. Plasma Unesterified Fatty-Acid Profile Is Dramatically and Acutely Changed under Ischemic Stroke in the Mouse Model. Lipids 2018, 53, 641–645. [Google Scholar] [CrossRef]
- Shimizu, H.; Ito, A.; Sakurada, K.; Nakamura, J.; Tanaka, K.; Komatsu, M.; Takeda, M.; Saito, K.; Endo, Y.; Kozaki, T.; et al. AK106-001616, a Potent and Selective Inhibitor of Cytosolic Phospholipase A2: In Vivo Efficacy for Inflammation, Neuropathic Pain, and Pulmonary Fibrosis. J. Pharmacol. Exp. Ther. 2019, 369, 511–522. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.-K.; Zhang, Y.P.; Titsworth, W.L.; Jiang, X.; Han, S.; Lu, P.-H.; Shields, C.B.; Xu, X.-M. A novel role of phospholipase A2 in mediating spinal cord secondary injury. Ann. Neurol. 2006, 59, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Deng, L.; Zhang, Y.P.; Lu, Q.; Wang, X.; Hu, J.; Oakes, E.; Bonventre, J.V.; Shields, C.B.; Xu, X. Cytosolic phospholipase A2 protein as a novel therapeutic target for spinal cord injury. Ann. Neurol. 2014, 75, 644–658. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Mejia, R.O.; Mucke, L. Phospholipase A2 and arachidonic acid in Alzheimer’s disease. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2010, 1801, 784–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.-M.; Simonyi, A.; Sun, A.Y.; Sun, G.Y. Phospholipases A2 and neural membrane dynamics: Implications for Alzheimer’s disease. J. Neurochem. 2011, 116, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; He, Y.; Chuang, D.Y.; Lee, J.C.; Gu, Z.; Simonyi, A.; Sun, A.Y. Integrating Cytosolic Phospholipase A2 with Oxidative/Nitrosative Signaling Pathways in Neurons: A Novel Therapeutic Strategy for AD. Mol. Neurobiol. 2012, 46, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, E.L.; Forlenza, O.V.; Gattaz, W.F. Phospholipase A2 activation as a therapeutic approach for cognitive enhancement in ear-ly-stage Alzheimer disease. Psychopharmacology 2009, 202, 37–51. [Google Scholar] [CrossRef]
- Sanchez-Mejia, R.O.; Newman, J.; Toh, S.; Yu, G.-Q.; Zhou, Y.; Halabisky, B.; Cissé, M.; Scearce-Levie, K.; Cheng, I.H.; Gan, L.; et al. Phospholipase A2 reduction ameliorates cognitive deficits in a mouse model of Alzheimer’s disease. Nat. Neurosci. 2008, 11, 1311–1318. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Bao, X.; Shan, L.; Ling, Y.; Ding, Y.; Wang, J.; Cao, Y.; Wang, Q.; Cui, W.; Xu, S. Calcium-Sensing Receptor Mediates beta-Amyloid-Induced Synaptic Formation Impairment and Cognitive Deficits via Reg-ulation of Cytosolic Phospholipase A2/Prostaglandin E2 Metabolic Pathway. Front. Aging Neurosci. 2020, 12, 144. [Google Scholar] [CrossRef]
- Kaya, I.; Jennische, E.; Lange, S.; Tarik Baykal, A.; Malmberg, P.; Fletcher, J.S. Brain region-specific amyloid plaque-associated myelin lipid loss, APOE deposition and disruption of the myelin sheath in familial Alzheimer’s disease mice. J. Neurochem. 2020, 154, 84–98. [Google Scholar] [CrossRef] [Green Version]
- Mehla, J.; Lacoursiere, S.G.; Lapointe, V.; McNaughton, B.L.; Sutherland, R.J.; McDonald, R.J.; Mohajerani, M.H. Age-dependent behavioral and biochemical characterization of single APP knock-in mouse (APPNL-G-F/NL-G-F) model of Alzheimer’s disease. Neurobiol. Aging 2018, 75, 25–37. [Google Scholar] [CrossRef]
- Sakakibara, Y.; Sekiya, M.; Saito, T.; Saido, T.C.; Iijima, K.M. Amyloid-beta plaque formation and reactive gliosis are required for induction of cognitive deficits in App knock-in mouse models of Alzheimer’s disease. BMC Neurosci. 2019, 20, 13. [Google Scholar] [CrossRef]
- Emre, C.; Do, K.V.; Jun, B.; Hjorth, E.; Alcalde, S.G.; Kautzmann, M.-A.I.; Gordon, W.C.; Nilsson, P.; Bazan, N.G.; Schultzberg, M. Age-related changes in brain phospholipids and bioactive lipids in the APP knock-in mouse model of Alzheimer’s disease. Acta Neuropathol. Commun. 2021, 9, 1–26. [Google Scholar] [CrossRef]
- Granger, M.W.; Liu, H.; Fowler, C.; Blanchard, A.P.; Taylor, M.W.; Sherman, S.P.M.; Xu, H.; Le, W.; Bennett, S.A.L. Distinct disruptions in Land’s cycle remodeling of glycerophosphocholines in murine cortex mark symptomatic onset and progression in two Alzheimer’s disease mouse models. J. Neurochem. 2018, 149, 499–517. [Google Scholar] [CrossRef]
- Pérez-González, M.; Mendioroz, M.; Badesso, S.; Sucunza, D.; Roldan, M.; Espelosín, M.; Ursua, S.; Luján, R.; Cuadrado-Tejedor, M.; Garcia-Osta, A. PLA2G4E, a candidate gene for resilience in Alzheimer’s disease and a new target for dementia treatment. Prog. Neurobiol. 2020, 191, 101818. [Google Scholar] [CrossRef] [PubMed]
- Dahlgren, K.N.; Manelli, A.M.; Stine, W.B., Jr.; Baker, L.K.; Krafft, G.A.; LaDu, M.J. Oligomeric and fibrillar species of amyloid-beta peptides differentially affect neuronal viability. J. Biol. Chem. 2002, 277, 32046–33253. [Google Scholar] [CrossRef] [Green Version]
- Hicks, J.B.; Lai, Y.; Sheng, W.; Yang, X.; Zhu, D.; Sun, G.Y.; Lee, J.C.-M. Amyloid-β peptide induces temporal membrane biphasic changes in astrocytes through cytosolic phospholipase A2. Biochim. Biophys. Acta (BBA)—Biomembr. 2008, 1778, 2512–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, D.; Lai, Y.; Shelat, P.B.; Hu, C.; Sun, G.Y.; Lee, J.C. Phospholipases A2 mediate amyloid-beta peptide-induced mitochondrial dysfunction. J. Neurosci. 2006, 26, 11111–11119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagy-Bross, C.; Kasianov, K.; Solomonov, Y.; Braiman, A.; Friedman, A.; Hadad, N.; Lévy, R. The role of cytosolic phospholipase A2α in amyloid precursor protein induction by amyloid beta1-42: Implication for neurodegeneration. J. Neurochem. 2015, 132, 559–571. [Google Scholar] [CrossRef]
- Sagy-Bross, C.; Hadad, N.; Levy, R. Cytosolic phospholipase A2alpha upregulation mediates apoptotic neuronal death induced by ag-gregated amyloid-beta peptide1-42. Neurochem. Int. 2013, 63, 541–550. [Google Scholar] [CrossRef]
- Desbene, C.; Malaplate-Armand, C.; Youssef, I.; Garcia, P.; Stenger, C.; Sauvee, M.; Fischer, N.; Rimet, D.; Koziel, V.; Escanye, M.C.; et al. Critical role of cPLA2 in Abeta oligomer-induced neurodegeneration and memory deficit. Neurobiol. Aging 2012, 33, 1123.e17–1123.e29. [Google Scholar] [CrossRef]
- Teng, T.; Dong, L.; Ridgley, D.M.; Ghura, S.; Tobin, M.K.; Sun, G.Y.; LaDu, M.J.; Lee, J.C. Cytosolic Phospholipase A2 Facilitates Oligomeric Amyloid-beta Peptide Association with Microglia via Regulation of Mem-brane-Cytoskeleton Connectivity. Mol. Neurobiol. 2019, 56, 3222–3234. [Google Scholar] [CrossRef]
- Geng, X.; Yang, B.; Li, R.; Teng, T.; Ladu, M.J.; Sun, G.Y.; Greenlief, C.M.; Lee, J.C. Effects of Docosahexaenoic Acid and Its Peroxidation Product on Amyloid-beta Peptide-Stimulated Microglia. Mol. Neurobiol. 2020, 57, 1085–1098. [Google Scholar] [CrossRef] [PubMed]
- Teng, T.; Ridgley, D.M.; Tsoy, A.; Sun, G.Y.; Askarova, S.; Lee, J.C. Azelnidipine Attenuates the Oxidative and NFκB Pathways in Amyloid-β-Stimulated Cerebral Endothelial Cells. ACS Chem. Neurosci. 2018, 10, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Mattson, M.P. Apolipoprotein E and oxidative stress in brain with relevance to Alzheimer’s disease. Neurobiol. Dis. 2020, 138, 104795. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, B.; Solomon, V.; Fonteh, A.; Rapoport, S.I.; Bennett, D.A.; Arvanitakis, Z.; Chui, H.C.; Miller, C.; Sullivan, P.M.; et al. Calcium-dependent cytosolic phospholipase A2 activation is implicated in neuroinflammation and oxidative stress associated with ApoE4. Mol. Neurodegener. 2021, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Turk, J.; Song, H.; Wohltmann, M.; Frankfater, C.; Lei, X.; Ramanadham, S. Metabolic Effects of Selective Deletion of Group VIA Phospholipase A2 from Macrophages or Pancreatic Islet Beta-Cells. Biomolecules 2020, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Turk, J.; White, T.D.; Nelson, A.J.; Lei, X.; Ramanadham, S. iPLA2beta and its role in male fertility, neurological disorders, metabolic disorders, and inflammation. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2019, 1864, 846–860. [Google Scholar] [CrossRef] [PubMed]
- Balboa, M.A.; Varela-Nieto, I.; Lucas, K.K.; Dennis, E.A. Expression and function of phospholipase A2 in brain. FEBS Lett. 2002, 531, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Mendes, C.T.; Gattaz, W.F.; Schaeffer, E.L.; Forlenza, O.V. Modulation of phospholipase A2 activity in primary cultures of rat cortical neurons. J. Neural Transm. 2005, 112, 1297–1308. [Google Scholar] [CrossRef]
- Kinghorn, K.J.; Castillo-Quan, J.I.; Bartolome, F.; Angelova, P.R.; Li, L.; Pope, S.; Cocheme, H.M.; Khan, S.; Asghari, S.; Bhatia, K.P.; et al. Loss ofPLA2G6leads to elevated mitochondrial lipid peroxidation and mitochondrial dysfunction. Brain 2015, 138, 1801–1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strokin, M.; Reiser, G. Mitochondria from a mouse model of the human infantile neuroaxonal dystrophy (INAD) with genetic defects in VIA iPLA2 have disturbed Ca2+ regulation with reduction in Ca2+ capacity. Neurochem. Int. 2016, 99, 187–193. [Google Scholar] [CrossRef] [PubMed]
- St-Gelais, F.; Menard, C.; Congar, P.; Trudeau, L.E.; Massicotte, G. Postsynaptic injection of calcium-independent phospholipase A2 inhibitors selectively increases AMPA receptor-mediated synaptic transmission. Hippocampus 2004, 14, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Blanchard, H.; Taha, A.Y.; Cheon, Y.; Kim, H.-W.; Turk, J.; Rapoport, S.I. iPLA2β Knockout Mouse, a Genetic Model for Progressive Human Motor Disorders, Develops Age-Related Neuropathology. Neurochem. Res. 2014, 39, 1522–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basselin, M.; Rosa, A.O.; Ramadan, E.; Cheon, Y.; Chang, L.; Chen, M.; Greenstein, D.; Wohltmann, M.; Turk, J.; Rapoport, S.I. Imaging decreased brain docosahexaenoic acid metabolism and signaling in iPLA(2)beta (VIA)-deficient mice. J. Lipid Res. 2010, 51, 3166–3173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strokin, M.; Sergeeva, M.; Reiser, G. Docosahexaenoic acid and arachidonic acid release in rat brain astrocytes is mediated by two separate isoforms of phospholipase A2and is differently regulated by cyclic AMP and Ca2+. Br. J. Pharmacol. 2003, 139, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- Cheon, Y.; Kim, H.W.; Igarashi, M.; Modi, H.R.; Chang, L.; Ma, K.; Greenstein, D.; Wohltmann, M.; Turk, J.; Rapoport, S.I.; et al. Disturbed brain phospholipid and docosahexaenoic acid metabolism in calcium-independent phospholipase A(2)-VIA (iPLA(2)beta)-knockout mice. Biochim. Biophys. Acta 2012, 1821, 1278–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, S.; Yoda, E.; Sasaki, Y.; Nakatani, Y.; Kuwata, H. Calcium-independent phospholipase A2gamma (iPLA2gamma) and its roles in cellular functions and diseases. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2019, 1864, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Jaburek, M.; Pruchova, P.; Holendova, B.; Galkin, A.; Jezek, P. Antioxidant Synergy of Mitochondrial Phospholipase PNPLA8/iPLA2gamma with Fatty Acid-Conducting SLC25 Gene Family Transporters. Antioxidants 2021, 10, 678. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Moon, S.H.; Jenkins, C.M.; Li, M.; Sims, H.F.; Guan, S.; Gross, R.W. The phospholipase iPLA2gamma is a major mediator releasing oxidized aliphatic chains from cardiolipin, integrating mito-chondrial bioenergetics and signaling. J. Biol. Chem. 2017, 292, 10672–10684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancuso, D.J.; Jenkins, C.M.; Gross, R.W. The genomic organization, complete mRNA sequence, cloning, and expression of a novel human intracellular membrane-associated calcium-independent phospholipase A(2). J. Biol. Chem. 2000, 275, 9937–9945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takata-Tanaka, H.; Takeya, R.; Sumimoto, H. A Novel Intracellular Membrane-Bound Calcium-Independent Phospholipase A2. Biochem. Biophys. Res. Commun. 2000, 272, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Allyson, J.; Bi, X.; Baudry, M.; Massicotte, G. Maintenance of Synaptic Stability Requires Calcium-Independent Phospholipase A2Activity. Neural Plast. 2012, 2012, 569149. [Google Scholar] [CrossRef] [Green Version]
- Ménard, C.; Chartier, E.; Patenaude, C.; Robinson, P.; Cyr, M.; Baudry, M.; Massicotte, G. Calcium-independent phospholipase A2 influences AMPA-mediated toxicity of hippocampal slices by regulating the GluR1 subunit in synaptic membranes. Hippocampus 2007, 17, 1109–1120. [Google Scholar] [CrossRef]
- Fusco, C.; Frattini, D.; Panteghini, C.; Pascarella, R.; Garavaglia, B. A Case of Infantile Neuroaxonal Dystrophy of Neonatal Onset. J. Child Neurol. 2014, 30, 368–370. [Google Scholar] [CrossRef] [PubMed]
- Mori, A.; Hatano, T.; Inoshita, T.; Shiba-Fukushima, K.; Koinuma, T.; Meng, H.; Kubo, S.-I.; Spratt, S.; Cui, C.; Yamashita, C.; et al. Parkinson’s disease-associated iPLA2-VIA/PLA2G6 regulates neuronal functions and α-synuclein stability through membrane remodeling. Proc. Natl. Acad. Sci. USA 2019, 116, 20689–20699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friend, S.F.; Nachnani, R.; Powell, S.B.; Risbrough, V.B. C-Reactive Protein: Marker of risk for post-traumatic stress disorder and its potential for a mechanistic role in trauma response and recovery. Eur. J. Neurosci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Hariprasad, G. Human Secretary Phospholipase A2 Mutations and Their Clinical Implications. J. Inflamm. Res. 2020, 13, 551–561. [Google Scholar] [CrossRef]
- Talib, L.L.; Diniz, B.; Zainaghi, I.A.; Forlenza, O.V.; Gattaz, W.F. A radioenzymatic assay to identify three groups of phospholipase A2 in platelets. Prostaglandins Leukot. Essent. Fat. Acids 2012, 86, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Duchez, A.-C.; Boudreau, L.H.; Naika, G.S.; Rousseau, M.; Cloutier, N.; Levesque, T.; Gelb, M.H.; Boilard, E. Respective contribution of cytosolic phospholipase A2α and secreted phospholipase A2 IIA to inflammation and eicosanoid production in arthritis. Prostaglandins Other Lipid Mediat. 2019, 143, 106340. [Google Scholar] [CrossRef] [PubMed]
- Dore, E.; Boilard, E. Roles of secreted phospholipase A2 group IIA in inflammation and host defense. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2018, 1864, 789–802. [Google Scholar] [CrossRef]
- Murakami, M.; Yamamoto, K.; Taketomi, Y. Phospholipase A2 in skin biology: New insights from gene-manipulated mice and lipidomics. Inflamm. Regen. 2018, 38, 31. [Google Scholar] [CrossRef]
- Murakami, M.; Miki, Y.; Sato, H.; Murase, R.; Taketomi, Y.; Yamamoto, K. Group IID, IIE, IIF and III secreted phospholipase A2s. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2019, 1864, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Zong, Y.; Mohammad, A.; Ajit, D.; Cui, J.; Han, D.; Hamilton, J.L.; Simonyi, A.; Sun, A.Y.; Gu, Z.; et al. Proinflammatory cytokines and lipopolysaccharide induce changes in cell morphology, and upregulation of ERK1/2, iNOS and sPLA2-IIA expression in astrocytes and microglia. J. Neuroinflamm. 2011, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Villanueva, E.; Little, J.P.; Lambeau, G.; Klegeris, A. Secreted phospholipase A2 group IIA is a neurotoxin released by stimulated human glial cells. Mol. Cell. Neurosci. 2012, 49, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.P.; Payette, P.; Mudgett, J.; Vadas, P.; Pruzanski, W.; Kwan, M.; Tang, C.; Rancourt, D.E.; Cromlish, W.A. A Natural Disruption of the Secretory Group II Phospholipase A2 Gene in Inbred Mouse Strains. J. Biol. Chem. 1995, 270, 22378–22385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grass, D.S.; Felkner, R.H.; Chiang, M.Y.; Wallace, R.E.; Nevalainen, T.J.; Bennett, C.F.; Swanson, M.E. Expression of human group II PLA2 in transgenic mice results in epidermal hyperplasia in the absence of inflammatory infiltrate. J. Clin. Investig. 1996, 97, 2233–2241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivandic, B.; Castellani, L.W.; Wang, X.P.; Qiao, J.H.; Mehrabian, M.; Navab, M.; Fogelman, A.M.; Grass, D.S.; Swanson, M.E.; de Beer, M.C.; et al. Role of group II secretory phospholipase A2 in atherosclerosis: 1. Increased atherogenesis and altered lipoproteins in transgenic mice expressing group IIa phospholipase A2. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1284–1290. [Google Scholar] [CrossRef] [Green Version]
- Bonventre, J.V. Cytosolic phospholipase A2α reigns supreme in arthritis and bone resorption. Trends Immunol. 2004, 25, 116–119. [Google Scholar] [CrossRef]
- Subra, C.; Grand, D.; Laulagnier, K.; Stella, A.; Lambeau, G.; Paillasse, M.; De Medina, P.; Monsarrat, B.; Perret, B.; SilventePoirot, S.; et al. Exosomes account for vesicle-mediated transcellular transport of activatable phospholipases and prostaglandins. J. Lipid Res. 2010, 51, 2105–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinkuolie, A.O.; Lawler, P.R.; Chu, A.Y.; Caulfield, M.; Mu, J.; Ding, B.; Nyberg, F.; Glynn, R.J.; Ridker, P.M.; Hurt-Camejo, E.; et al. Group IIA Secretory Phospholipase A2, Vascular Inflammation, and Incident Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1182–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, S.; Kazepidou, E.; Antonelou, M.H.; Leondaritis, G.; Tsapinou, A.; Koulouras, V.P.; Avgeropoulos, A.; Nakos, G.; Lekka, M.E. Secretory Phospholipase A2-IIA Protein and mRNA Pools in Extracellular Vesicles of Bronchoalveolar Lavage Fluid from Patients with Early Acute Respiratory Distress Syndrome: A New Perception in the Dissemination of Inflammation? Pharmaceuticals 2020, 13, 415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xia, R.; Jia, J.; Wang, L.; Li, K.; Li, Y.; Zhang, J. Oleanolic acid protects against cognitive decline and neuroinflammation-mediated neurotoxicity by blocking secretory phospholipase A2 IIA-activated calcium signals. Mol. Immunol. 2018, 99, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Moses, G.S.; Jensen, M.D.; Lue, L.-F.; Walker, D.G.; Sun, A.Y.; Simonyi, A.; Sun, G.Y. Secretory PLA2-IIA: A new inflammatory factor for Alzheimer’s disease. J. Neuroinflamm. 2006, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Sheng, W.; Ridgley, D.M.; Haidekker, M.A.; Sun, G.Y.; Lee, J.C. Astrocytes regulate α-secretase-cleaved soluble amyloid precursor protein secretion in neuronal cells: Involvement of group IIA secretory phospholipase A2. Neuroscience 2015, 300, 508–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.-N.; Wang, Q.; Simonyi, A.; Chen, J.-J.; Cheung, W.-M.; He, Y.Y.; Xu, J.; Sun, A.Y.; Hsu, C.Y.; Sun, G.Y. Induction of secretory phospholipase A2 in reactive astrocytes in response to transient focal cerebral ischemia in the rat brain. J. Neurochem. 2004, 90, 637–645. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, A.Y.; Pardeike, J.; Müller, R.H.; Simonyi, A.; Sun, G.Y. Neuroprotective effects of a nanocrystal formulation of sPLA2 inhibitor PX-18 in cerebral ischemia/reperfusion in gerbils. Brain Res. 2009, 1285, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titsworth, W.L.; Cheng, X.; Ke, Y.; Deng, L.; Burckardt, K.A.; Pendleton, C.; Liu, N.K.; Shao, H.; Cao, Q.L.; Xu, X.M. Differential expression of sPLA2 following spinal cord injury and a functional role for sPLA2-IIA in mediating oli-godendrocyte death. Glia 2009, 57, 1521–1537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quindlen-Hotek, J.C.; Kartha, S.; Winkelstein, B.A. Immediate inhibition of spinal secretory phospholipase A2 prevents the pain and elevated spinal neuronal hyperexcitability and neuroimmune regulatory genes that develop with nerve root compression. NeuroReport 2020, 31, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Kartha, S.; Weisshaar, C.L.; Philips, B.H.; Winkelstein, B.A. Pre-treatment with Meloxicam Prevents the Spinal Inflammation and Oxidative Stress in DRG Neurons that Accompany Painful Cervical Radiculopathy. Neuroscience 2018, 388, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Kartha, S.; Yan, L.; Ita, M.E.; Amirshaghaghi, A.; Luo, L.; Wei, Y.; Tsourkas, A.; Winkelstein, B.A.; Cheng, Z. Phospholipase A2 Inhibitor-Loaded Phospholipid Micelles Abolish Neuropathic Pain. ACS Nano 2020, 14, 8103–8115. [Google Scholar] [CrossRef]
- Sugasini, D.; Thomas, R.; Yalagala, P.C.; Tai, L.M.; Subbaiah, P.V. Dietary docosahexaenoic acid (DHA) as lysophosphatidylcholine, but not as free acid, enriches brain DHA and improves memory in adult mice. Sci. Rep. 2017, 7, 11263. [Google Scholar] [CrossRef]
- Sugasini, D.; Yalagala, P.C.R.; Goggin, A.; Tai, L.M.; Subbaiah, P.V. Enrichment of brain docosahexaenoic acid (DHA) is highly dependent upon the molecular carrier of dietary DHA: Lyso-phosphatidylcholine is more efficient than either phosphatidylcholine or triacylglycerol. J. Nutr. Biochem. 2019, 74, 108231. [Google Scholar] [CrossRef]
- Chan, J.P.; Wong, B.H.; Chin, C.F.; Galam, D.L.A.; Foo, J.C.; Wong, L.C.; Ghosh, S.; Wenk, M.R.; Cazenave-Gassiot, A.; Silver, D.L. The lysolipid transporter Mfsd2a regulates lipogenesis in the developing brain. PLoS Biol. 2018, 16, e2006443. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Ma, D.; Shui, G.; Wong, P.; Cazenave-Gassiot, A.; Zhang, X.; Wenk, M.R.; Goh, E.; Silver, D. Mfsd2a is a transporter for the essential omega-3 fatty acid docosahexaenoic acid. Nature 2014, 509, 503–506. [Google Scholar] [CrossRef]
- Cater, R.J.; Chua, G.L.; Erramilli, S.K.; Keener, J.E.; Choy, B.C.; Tokarz, P.; Chin, C.F.; Quek, D.Q.Y.; Kloss, B.; Pepe, J.G.; et al. Structural basis of omega-3 fatty acid transport across the blood–brain barrier. Nature 2021, 595, 315–319. [Google Scholar] [CrossRef]
- Scala, M.; Chua, G.L.; Chin, C.F.; Alsaif, H.S.; Borovikov, A.; Riazuddin, S.; Riazuddin, S.; Manzini, M.C.; Severino, M.; Kuk, A.; et al. Biallelic MFSD2A variants associated with congenital microcephaly, developmental delay, and recognizable neuroimaging features. Eur. J. Hum. Genet. 2020, 28, 1509–1519. [Google Scholar] [CrossRef]
- Harel, T.; Quek, D.Q.Y.; Wong, B.H.; Cazenave-Gassiot, A.; Wenk, M.R.; Fan, H.; Berger, I.; Shmueli, D.; Shaag, A.; Silver, D.L.; et al. Homozygous mutation in MFSD2A, encoding a lysolipid transporter for docosahexanoic acid, is associated with microcephaly and hypomyelination. Neurogenetics 2018, 19, 227–235. [Google Scholar] [CrossRef]
- Pérez-Chacón, G.; Astudillo, A.M.; Balgoma, D.; Balboa, M.A.; Balsinde, J. Control of free arachidonic acid levels by phospholipases A2 and lysophospholipid acyltransferases. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2009, 1791, 1103–1113. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.C.; Folco, G. Lysophospholipid acyltransferases and leukotriene biosynthesis: Intersection of the Lands cycle and the ara-chidonate PI cycle. J. Lipid Res. 2019, 60, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Chacón, G.; Astudillo, A.M.; Ruipérez, V.; Balboa, M.A.; Balsinde, J. Signaling Role for Lysophosphatidylcholine Acyltransferase 3 in Receptor-Regulated Arachidonic Acid Reacylation Reactions in Human Monocytes. J. Immunol. 2009, 184, 1071–1078. [Google Scholar] [CrossRef] [Green Version]
- Johansen, A.; Rosti, R.O.; Musaev, D.; Sticca, E.; Harripaul, R.; Zaki, M.; Caglayan, A.O.; Azam, M.; Sultan, T.; Froukh, T.; et al. Mutations in MBOAT7, Encoding Lysophosphatidylinositol Acyltransferase I, Lead to Intellectual Disability Accompa-nied by Epilepsy and Autistic Features. Am. J. Hum. Genet. 2016, 99, 912–916. [Google Scholar] [CrossRef] [Green Version]
- Sundaram, J.R.; Chan, E.S.; Poore, C.P.; Pareek, T.; Cheong, W.F.; Shui, G.; Tang, N.; Low, C.-M.; Wenk, M.R.; Kesavapany, S. Cdk5/p25-Induced Cytosolic PLA2-Mediated Lysophosphatidylcholine Production Regulates Neuroinflammation and Triggers Neurodegeneration. J. Neurosci. 2012, 32, 1020–1034. [Google Scholar] [CrossRef]
- Yatomi, Y.; Kurano, M.; Ikeda, H.; Igarashi, K.; Kano, K.; Aoki, J. Lysophospholipids in laboratory medicine. Proc. Jpn. Acad. Ser. B 2018, 94, 373–389. [Google Scholar] [CrossRef] [Green Version]
- Aoki, J.; Inoue, A.; Okudaira, S. Two pathways for lysophosphatidic acid production. Biochim. Biophys. Acta 2008, 1781, 513–518. [Google Scholar] [CrossRef]
- Nakasaki, T.; Tanaka, T.; Okudaira, S.; Hirosawa, M.; Umemoto, E.; Otani, K.; Jin, S.; Bai, Z.; Hayasaka, H.; Fukui, Y.; et al. Involvement of the Lysophosphatidic Acid-Generating Enzyme Autotaxin in Lymphocyte-Endothelial Cell Interactions. Am. J. Pathol. 2008, 173, 1566–1576. [Google Scholar] [CrossRef] [Green Version]
- Herr, D.R.; Chew, W.S.; Satish, R.L.; Ong, W.-Y. Pleotropic Roles of Autotaxin in the Nervous System Present Opportunities for the Development of Novel Therapeutics for Neurological Diseases. Mol. Neurobiol. 2019, 57, 372–392. [Google Scholar] [CrossRef]
- Ma, L.; Uchida, H.; Nagai, J.; Inoue, M.; Aoki, J.; Ueda, H. Evidence for De Novo Synthesis of Lysophosphatidic Acid in the Spinal Cord through Phospholipase A2 and Autotaxin in Nerve Injury-Induced Neuropathic Pain. J. Pharmacol. Exp. Ther. 2010, 333, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Guo, M.; Feng, Y.; Dong, Q.; Cui, M. Lysophospholipids and Their G-Coupled Protein Signaling in Alzheimer’s Disease: From Physiological Performance to Patho-logical Impairment. Front. Mol. Neurosci. 2020, 13, 58. [Google Scholar] [CrossRef] [Green Version]
- Blaho, V.A.; Chun, J. ‘Crystal’ Clear? Lysophospholipid Receptor Structure Insights and Controversies. Trends Pharmacol. Sci. 2018, 39, 953–966. [Google Scholar] [CrossRef]
- Ramesh, S.; Govindarajulu, M.; Suppiramaniam, V.; Moore, T.; Dhanasekaran, M. Autotaxin(−)Lysophosphatidic Acid Signaling in Alzheimer’s Disease. Int. J. Mol. Sci. 2018, 19, 1827. [Google Scholar] [CrossRef] [Green Version]
- Fransson, J.; Gomez-Conde, A.I.; Romero-Imbroda, J.; Fernandez, O.; Leyva, L.; de Fonseca, F.R.; Chun, J.; Louapre, C.; Van-Evercooren, A.B.; Zujovic, V.; et al. Activation of Macrophages by Lysophosphatidic Acid through the Lysophosphatidic Acid Receptor 1 as a Novel Mecha-nism in Multiple Sclerosis Pathogenesis. Mol. Neurobiol. 2021, 58, 470–482. [Google Scholar] [CrossRef]
- Yang, B.; Fritsche, K.L.; Beversdorf, D.Q.; Gu, Z.; Lee, J.C.; Folk, W.R.; Greenlief, C.M.; Sun, G.Y. Yin-Yang Mechanisms Regulating Lipid Peroxidation of Docosahexaenoic Acid and Arachidonic Acid in the Central Nervous System. Front. Neurol. 2019, 10, 642. [Google Scholar] [CrossRef]
- Schebb, N.H.; Ostermann, A.I.; Yang, J.; Hammock, B.D.; Hahn, A.; Schuchardt, J.P. Comparison of the effects of long-chain omega-3 fatty acid supplementation on plasma levels of free and esterified oxylipins. Prostaglandins Other Lipid Mediat. 2014, 113–115, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Buczynski, M.W.; Dumlao, D.S.; Dennis, E.A. Thematic Review Series: Proteomics. An integrated omics analysis of eicosanoid biology. J. Lipid Res. 2009, 50, 1015–1038. [Google Scholar] [CrossRef] [Green Version]
- Palacios-Pelaez, R.; Lukiw, W.J.; Bazan, N.G. Omega-3 Essential Fatty Acids Modulate Initiation and Progression of Neurodegenerative Disease. Mol. Neurobiol. 2010, 41, 367–374. [Google Scholar] [CrossRef]
- Ferdouse, A.; Leng, S.; Winter, T.; Aukema, H.M. Dietary n-6 and n-3 PUFA alter the free oxylipin profile differently in male and female rat hearts. Br. J. Nutr. 2019, 122, 252–261. [Google Scholar] [CrossRef]
- Ferdouse, A.; Leng, S.; Winter, T.; Aukema, H.M. The Brain Oxylipin Profile Is Resistant to Modulation by Dietary n-6 and n-3 Polyunsaturated Fatty Acids in Male and Female Rats. Lipids 2019, 54, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Tourdot, B.E.; Ahmed, I.; Holinstat, M. The emerging role of oxylipins in thrombosis and diabetes. Front. Pharmacol. 2014, 4, 176. [Google Scholar] [CrossRef]
- Grapov, D.; Adams, S.H.; Pedersen, T.L.; Garvey, W.T.; Newman, J.W. Type 2 Diabetes Associated Changes in the Plasma Non-Esterified Fatty Acids, Oxylipins and Endocannabinoids. PLoS ONE 2012, 7, e48852. [Google Scholar] [CrossRef] [Green Version]
- Zivkovic, A.M.; Yang, J.; Georgi, K.; Hegedus, C.; Nording, M.; O’Sullivan, A.; German, J.B.; Hogg, R.J.; Weiss, R.H.; Bay, C.; et al. Serum oxylipin profiles in IgA nephropathy patients reflect kidney functional alterations. Metabolomics 2012, 8, 1102–1113. [Google Scholar] [CrossRef] [Green Version]
- Jira, W.; Spiteller, G.; Richter, A. Increased levels of lipid oxidation products in low density lipoproteins of patients suffering from rheumatoid arthritis. Chem. Phys. Lipids 1997, 87, 81–89. [Google Scholar] [CrossRef]
- Duffield, J.S.; Hong, S.; Vaidya, V.S.; Lu, Y.; Fredman, G.; Serhan, C.N.; Bonventre, J.V. Resolvin D Series and Protectin D1 Mitigate Acute Kidney Injury. J. Immunol. 2006, 177, 5902–5911. [Google Scholar] [CrossRef] [Green Version]
- Aoki, H.; Hisada, T.; Ishizuka, T.; Utsugi, M.; Kawata, T.; Shimizu, Y.; Okajima, F.; Dobashi, K.; Mori, M. Resolvin E1 dampens airway inflammation and hyperresponsiveness in a murine model of asthma. Biochem. Biophys. Res. Commun. 2008, 367, 509–515. [Google Scholar] [CrossRef]
- O’Connell, T.D.; Mason, R.P.; Budoff, M.J.; Navar, A.M.; Shearer, G.C. Mechanistic insights into cardiovascular protection for omega-3 fatty acids and their bioactive lipid metabolites. Eur. Heart J. Suppl. 2020, 22, J3–J20. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hossain, S.; Al Mamun, A.; Matsuzaki, K.; Arai, H. Docosahexaenoic acid: One molecule diverse functions. Crit. Rev. Biotechnol. 2016, 37, 579–597. [Google Scholar] [CrossRef]
- Yang, X.; Sheng, W.; Sun, G.Y.; Lee, J.C.-M. Effects of fatty acid unsaturation numbers on membrane fluidity and α-secretase-dependent amyloid precursor protein processing. Neurochem. Int. 2011, 58, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Schuchardt, J.P.; Schmidt, S.; Kressel, G.; Willenberg, I.; Hammock, B.D.; Hahn, A.; Schebb, N.H. Modulation of blood oxylipin levels by long-chain omega-3 fatty acid supplementation in hyper- and normolipidemic men. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Neubronner, J.; Schuchardt, J.P.; Kressel, G.; Merkel, M.; Von Schacky, C.; Hahn, A. Enhanced increase of omega-3 index in response to long-term n-3 fatty acid supplementation from triacylglycerides versus ethyl esters. Eur. J. Clin. Nutr. 2010, 65, 247–254. [Google Scholar] [CrossRef]
- Yang, B.; Li, R.; Woo, T.; Browning, J.D., Jr.; Song, H.; Gu, Z.; Cui, J.; Lee, J.C.; Fritsche, K.L.; Beversdorf, D.Q.; et al. Maternal Dietary Docosahexaenoic Acid Alters Lipid Peroxidation Products and (n-3)/(n-6) Fatty Acid Balance in Offspring Mice. Metabolites 2019, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.Y.; Appenteng, M.K.; Li, R.; Woo, T.; Yang, B.; Qin, C.; Pan, M.; Cieślik, M.; Cui, J.; Fritsche, K.L.; et al. Docosahexaenoic Acid (DHA) Supplementation Alters Phospholipid Species and Lipid Peroxidation Products in Adult Mouse Brain, Heart, and Plasma. NeuroMol. Med. 2020, 23, 118–129. [Google Scholar] [CrossRef]
- Lamaziere, A.; Richard, D.; Barbe, U.; Kefi, K.; Bausero, P.; Wolf, C.; Visioli, F. Differential distribution of DHA-phospholipids in rat brain after feeding: A lipidomic approach. Prostaglandins Leukot. Essent. Fat. Acids 2011, 84, 7–11. [Google Scholar] [CrossRef]
- Rey, C.; Delpech, J.; Madore, C.; Nadjar, A.; Greenhalgh, A.; Amadieu, C.; Aubert, A.; Pallet, V.; Vaysse, C.; Layé, S.; et al. Dietary n-3 long chain PUFA supplementation promotes a pro-resolving oxylipin profile in the brain. Brain Behav. Immun. 2018, 76, 17–27. [Google Scholar] [CrossRef]
- Cohen, G.; Riahi, Y.; Sunda, V.; Deplano, S.; Chatgilialoglu, C.; Ferreri, C.; Kaiser, N.; Sasson, S. Signaling properties of 4-hydroxyalkenals formed by lipid peroxidation in diabetes. Free. Radic. Biol. Med. 2013, 65, 978–987. [Google Scholar] [CrossRef]
- Yang, B.; Li, R.; Michael Greenlief, C.; Fritsche, K.L.; Gu, Z.; Cui, J.; Lee, J.C.; Beversdorf, D.Q.; Sun, G.Y. Unveiling anti-oxidative and anti-inflammatory effects of docosahexaenoic acid and its lipid peroxidation product on lipo-polysaccharide-stimulated BV-2 microglial cells. J. Neuroinflamm. 2018, 15, 202. [Google Scholar] [CrossRef] [Green Version]
- Cutuli, D.; De Bartolo, P.; Caporali, P.; Laricchiuta, D.; Foti, F.; Ronci, M.; Rossi, C.; Neri, C.; Spalletta, G.; Caltagirone, C.; et al. n-3 polyunsaturated fatty acids supplementation enhances hippocampal functionality in aged mice. Front. Aging Neurosci. 2014, 6, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labrousse, V.F.; Nadjar, A.; Joffre, C.; Costes, L.; Aubert, A.; Gregoire, S.; Bretillon, L.; Laye, S. Short-term long chain omega3 diet protects from neuroinflammatory processes and memory impairment in aged mice. PLoS ONE 2012, 7, e36861. [Google Scholar] [CrossRef]
- Calzada, C.; Colas, R.; Guillot, N.; Guichardant, M.; Laville, M.; Véricel, E.; Lagarde, M. Subgram daily supplementation with docosahexaenoic acid protects low-density lipoproteins from oxidation in healthy men. Atherosclerosis 2010, 208, 467–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishikado, A.; Nishio, Y.; Morino, K.; Ugi, S.; Kondo, H.; Makino, T.; Kashiwagi, A.; Maegawa, H. Low concentration of 4-hydroxy hexenal increases heme oxygenase-1 expression through activation of Nrf2 and anti-oxidative activity in vascular endothelial cells. Biochem. Biophys. Res. Commun. 2010, 402, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, F.; Morino, K.; Ugi, S.; Ishikado, A.; Kondo, K.; Sato, D.; Konno, S.; Nemoto, K.; Kusunoki, C.; Sekine, O.; et al. 4-Hydroxy hexenal derived from dietary n-3 polyunsaturated fatty acids induces anti-oxidative enzyme heme oxy-genase-1 in multiple organs. Biochem. Biophys. Res. Commun. 2014, 443, 991–996. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.-L.; Qi, S.-H. ALDH2 Protects Against Ischemic Stroke in Rats by Facilitating 4-HNE Clearance and AQP4 DownRegulation. Neurochem. Res. 2018, 43, 1339–1347. [Google Scholar] [CrossRef]
- Hall, E.D.; Wang, J.A.; Miller, D.M.; Cebak, J.E.; Hill, R.L. Newer pharmacological approaches for antioxidant neuroprotection in traumatic brain injury. Neuropharmacology 2018, 145, 247–258. [Google Scholar] [CrossRef]
- Kuda, O. Bioactive metabolites of docosahexaenoic acid. Biochimie 2017, 136, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Galano, J.-M.; Lee, Y.Y.; Oger, C.; Vigor, C.; Vercauteren, J.; Durand, T.; Giera, M.; Lee, J.C.-Y. Isoprostanes, neuroprostanes and phytoprostanes: An overview of 25 years of research in chemistry and biology. Prog. Lipid Res. 2017, 68, 83–108. [Google Scholar] [CrossRef] [PubMed]
- Stark, D.T.; Bazan, N.G. Neuroprotectin D1 Induces Neuronal Survival and Downregulation of Amyloidogenic Processing in Alzheimer’s Disease Cellular Models. Mol. Neurobiol. 2011, 43, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Fauconnier, J.; Oger, C.; Farah, C.; Angebault-Prouteau, C.; Thireau, J.; Bideaux, P.; Scheuermann, V.; Bultel-Ponce, V.; Demion, M.; et al. Non-enzymatic oxidized metabolite of DHA, 4(RS)-4-F4t-neuroprostane protects the heart against reperfusion injury. Free Radic. Biol. Med. 2017, 102, 229–239. [Google Scholar] [CrossRef]
- Sun, G.Y.; Lin, T.-N. Time course for labeling of brain membrane phosphoinositides and other phospholipids after intracerebral injection of [32P]-ATP. Evaluation by an improved HPTLC procedure. Life Sci. 1989, 44, 689–696. [Google Scholar] [CrossRef]
- Wang, M.; Han, X. Advanced Shotgun Lipidomics for Characterization of Altered Lipid Patterns in Neurodegenerative Diseases and Brain Injury. Methods Mol. Biol. 2016, 1303, 405–422. [Google Scholar] [CrossRef] [Green Version]
- Huguenard, C.J.C.; Cseresznye, A.; Evans, J.E.; Oberlin, S.; Langlois, H.; Ferguson, S.; Darcey, T.; Nkiliza, A.; Dretsch, M.; Mullan, M.; et al. Plasma Lipidomic Analyses in Cohorts With mTBI and/or PTSD Reveal Lipids Differentially Associated With Diag-nosis and APOE epsilon4 Carrier Status. Front. Physiol. 2020, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Dray, F.; Charbonnel, B.; Maclouf, J. Radioimmunoassay of prostaglandins Falpha, E1 and E2 in human plasma. Eur. J. Clin. Investig. 1975, 5, 311–318. [Google Scholar] [CrossRef]
- Quinn, J.V.; Bilgrami, S.; Seidel, G.J.; Slotman, G.J. Evaluation of enzyme-linked immunosorbent assays for quantitation of eicosanoid mediators of sepsis syndrome. Shock 1996, 6, 142–149. [Google Scholar] [CrossRef]
- Wang, D.; DuBois, R.N. Measurement of Eicosanoids in Cancer Tissues. Methods Enzymol. 2007, 433, 27–50. [Google Scholar] [CrossRef]
- Liakh, I.; Pakiet, A.; Sledzinski, T.; Mika, A. Methods of the Analysis of Oxylipins in Biological Samples. Molecules 2020, 25, 349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinz, C.; Liggi, S.; Mocciaro, G.; Jung, S.; Induruwa, I.; Pereira, M.; Bryant, C.E.; Meckelmann, S.W.; O’Donnell, V.B.; Farndale, R.W.; et al. Comprehensive UHPLC Ion Mobility Quadrupole Time-of-Flight Method for Profiling and Quantification of Eicosanoids, Other Oxylipins, and Fatty Acids. Anal. Chem. 2019, 91, 8025–8035. [Google Scholar] [CrossRef] [PubMed]
- Willenberg, I.; Ostermann, A.I.; Schebb, N.H. Targeted metabolomics of the arachidonic acid cascade: Current state and challenges of LC–MS analysis of oxylipins. Anal. Bioanal. Chem. 2015, 407, 2675–2683. [Google Scholar] [CrossRef]
- Wang, Y.; Armando, A.M.; Quehenberger, O.; Yan, C.; Dennis, E.A. Comprehensive ultra-performance liquid chromatographic separation and mass spectrometric analysis of eicosanoid metabolites in human samples. J. Chromatogr. A 2014, 1359, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watrous, J.D.; Niiranen, T.J.; Lagerborg, K.A.; Henglin, M.; Xu, Y.J.; Rong, J.; Sharma, S.; Vasan, R.S.; Larson, M.G.; Armando, A.; et al. Directed Non-targeted Mass Spectrometry and Chemical Networking for Discovery of Eicosanoids and Related Oxylipins. Cell Chem. Biol. 2019, 26, 433–442 e4. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Title | Activities and Functions | References | |
|---|---|---|---|
| A. cPLA2 | 1. | Phosphorylation by mitogen-activated protein kinases (MAPK) | [8,13,14,15] |
| 2. | Translocation from cytoplasm to different intracellular components | [9] | |
| 3. | Neuronal excitation and glial activation | [13,14,15,16] | |
| 4. | Preference for release of ARA and inflammatory pathways | [17] | |
| 5. | Involvement in stroke, spinal cord and traumatic brain injury | [27,28,31,32] | |
| 6. | Involvement in AD | [33,34,35,36,37,38,39,40,41,42] | |
| 7. | Activation by oligomeric Aβ | [46,48,49,50,51,52,53] | |
| 8. | Inhibitors—arachidonyl trifluoromethyl ketone (AACOCF3 or ATK) | [17,23,24,53] | |
| B. iPLA2 | |||
| 1. | Multiple strings of ankyrin motif—binding with other proteins | [2,56,57] | |
| 2. | iPLA2β—most prominent phospholipase playing a house-keeping role-regulation of mitochondrial function | [58,60,61] | |
| 3. | iPLA2γ—has motif for mitochondrial localization | [67] | |
| 4. | Preference for the release of DHA | [63,64,65] | |
| 5. | Action on cardiolipins | [68,69] | |
| 6. | Regulation of AMPA neurons | [72,73] | |
| 7. | Specific inhibitor, bromoenol lactone (BEL) | [62] | |
| 8. | Mutations of GVI PLA2 gene—infantile neuroaxonal dystrophy (INAD) and neurodegeneration with brain iron accumulation (NBIA) | [57,60,63,74] | |
| C. sPLA2 | |||
| 1. | Small molecular weight proteins (20–40 kDa)—more than 10 isoforms | [77] | |
| 2. | Transcriptionally induced upon challenge by toxins and pro-inflammatory cytokines -inflammation | [76,80,83] | |
| 3. | Require high levels of calcium for activity in extracellular milieu | [78] | |
| 4. | Presence in extracellular vesicles secreted from astrocytes | [84,89,90] | |
| 5. | Many mouse strains lack the sPLA2-IIA gene | [85] | |
| 6. | Diseases—AD, cerebral ischemia, spinal cord injury | [93,94,95,96,97,98,99] | |
| 7. | Inhibitors: thioetheramide-phosphorylcholine | [100] | |
| D. Lysophospholipids | |||
| 1. | DHA-LPC—effectively transported to brain | [101,102] | |
| 2. | DHA-lysophospholipids—transported to brain through a specific Na-dependent transporter “Mfsd2A” | [103,104,105] | |
| 3. | Different isoforms of lysophospholipid acyltransferases engage in the “diacylation-reacylation” cycle | [108,109,110] | |
| 4. | LPA—pathways for synthesis | [114,115,116,117] | |
| 5. | LPA—interaction with G-protein-coupled receptors leading to different downstream signaling pathways | [120] | |
| E. ARE, DHA, oxylipins, 4-HNE and 4-HHE | |||
| 1. | Yin-Yang mechanism for the release of ARE and DHA | [122] | |
| 2. | Enzymatic and non-enzymatic free-radical oxidation reactions for production of oxylipins. | ||
| 3. | ARA—interaction with cyclooxygenase (COX), lipoxygenase (LOX) and cyctochrome P450 (CYP) for synthesis of prostaglandins, leukotrienes and thromboxanes, oxylipins that are inflammatory | [123,124] | |
| 4. | DHA—interact with different forms of LOX to form protective mediators such as resolvins, maresins and neuroprotection D1 | [125] | |
| 5. | Non-enzymatic free radical reactions—4-hydroxyhexenal (4-HHE) from DHA and 4-hydroxynonenal (4-HNE) from ARA | [122] | |
| 6. | Neuroprostanes | [155] | |
| 7. | Alkenyl aldehydes—bioactive and form adducts with proteins, phospholipids and nucleic acids | [143] | |
| 8. | 4-HNE—downstream from stimulation of cPLA2 and ARA | [144] | |
| 9. | 4-HHE—increase in brain upon dietary DHA | [139,144] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, G.Y.; Geng, X.; Teng, T.; Yang, B.; Appenteng, M.K.; Greenlief, C.M.; Lee, J.C. Dynamic Role of Phospholipases A2 in Health and Diseases in the Central Nervous System. Cells 2021, 10, 2963. https://doi.org/10.3390/cells10112963
Sun GY, Geng X, Teng T, Yang B, Appenteng MK, Greenlief CM, Lee JC. Dynamic Role of Phospholipases A2 in Health and Diseases in the Central Nervous System. Cells. 2021; 10(11):2963. https://doi.org/10.3390/cells10112963
Chicago/Turabian StyleSun, Grace Y., Xue Geng, Tao Teng, Bo Yang, Michael K. Appenteng, C. Michael Greenlief, and James C. Lee. 2021. "Dynamic Role of Phospholipases A2 in Health and Diseases in the Central Nervous System" Cells 10, no. 11: 2963. https://doi.org/10.3390/cells10112963
APA StyleSun, G. Y., Geng, X., Teng, T., Yang, B., Appenteng, M. K., Greenlief, C. M., & Lee, J. C. (2021). Dynamic Role of Phospholipases A2 in Health and Diseases in the Central Nervous System. Cells, 10(11), 2963. https://doi.org/10.3390/cells10112963