
Response to Electrostimulation Is Impaired in Muscle Cells from Patients with Chronic Obstructive Pulmonary Disease

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Muscle Biopsy and Myoblast Purification
2.3. Cell Culture and Electrical Pulse Stimulation
2.4. Fluorescence Microscopy
2.5. SDS-PAGE and Immunoblotting
2.6. Quantitative Polymerase Chain Reaction
2.7. Statistical Analysis
3. Results
3.1. Characteristics of the Study Groups
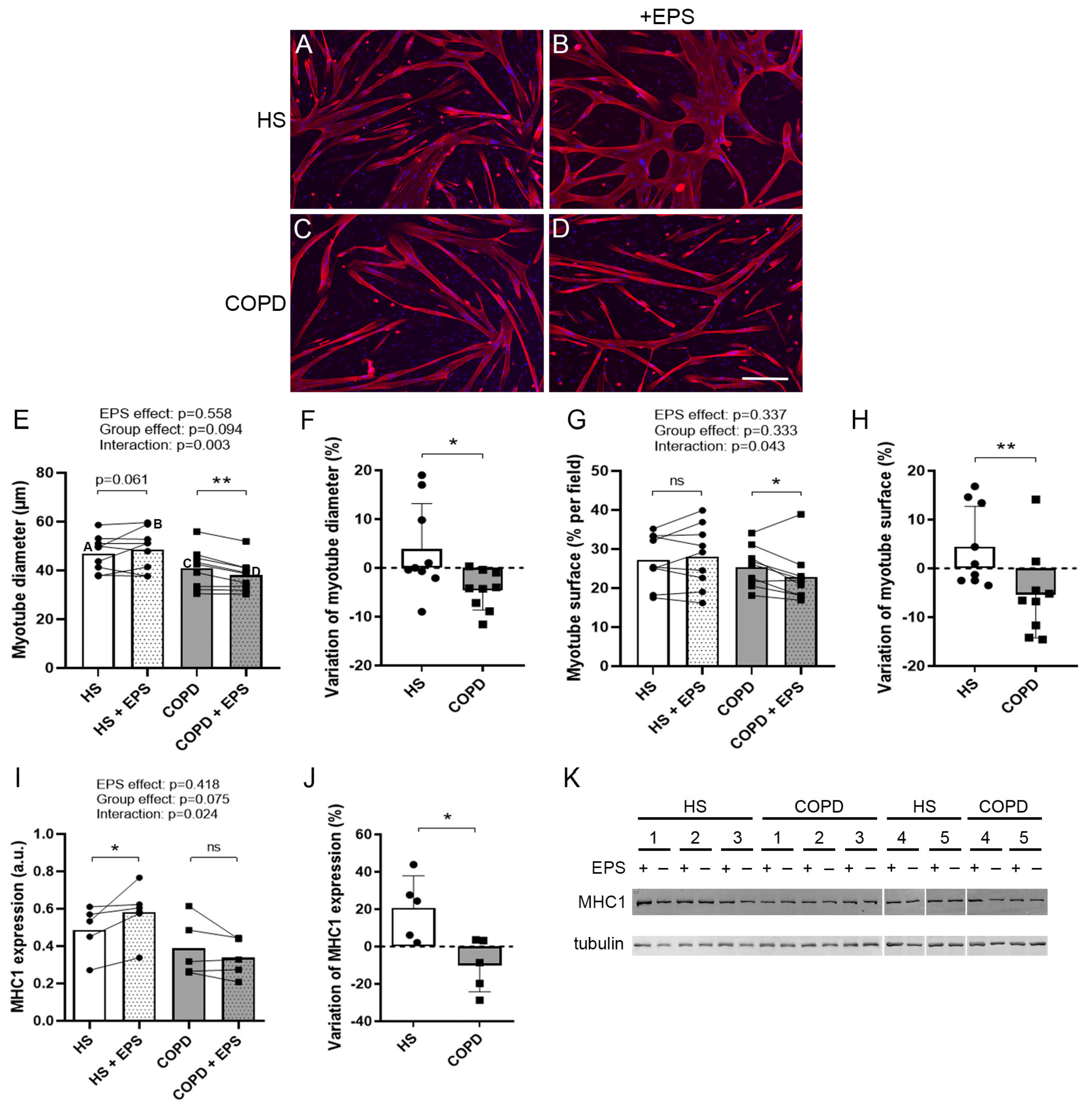
3.2. Effects of EPS on Myotube Morphology
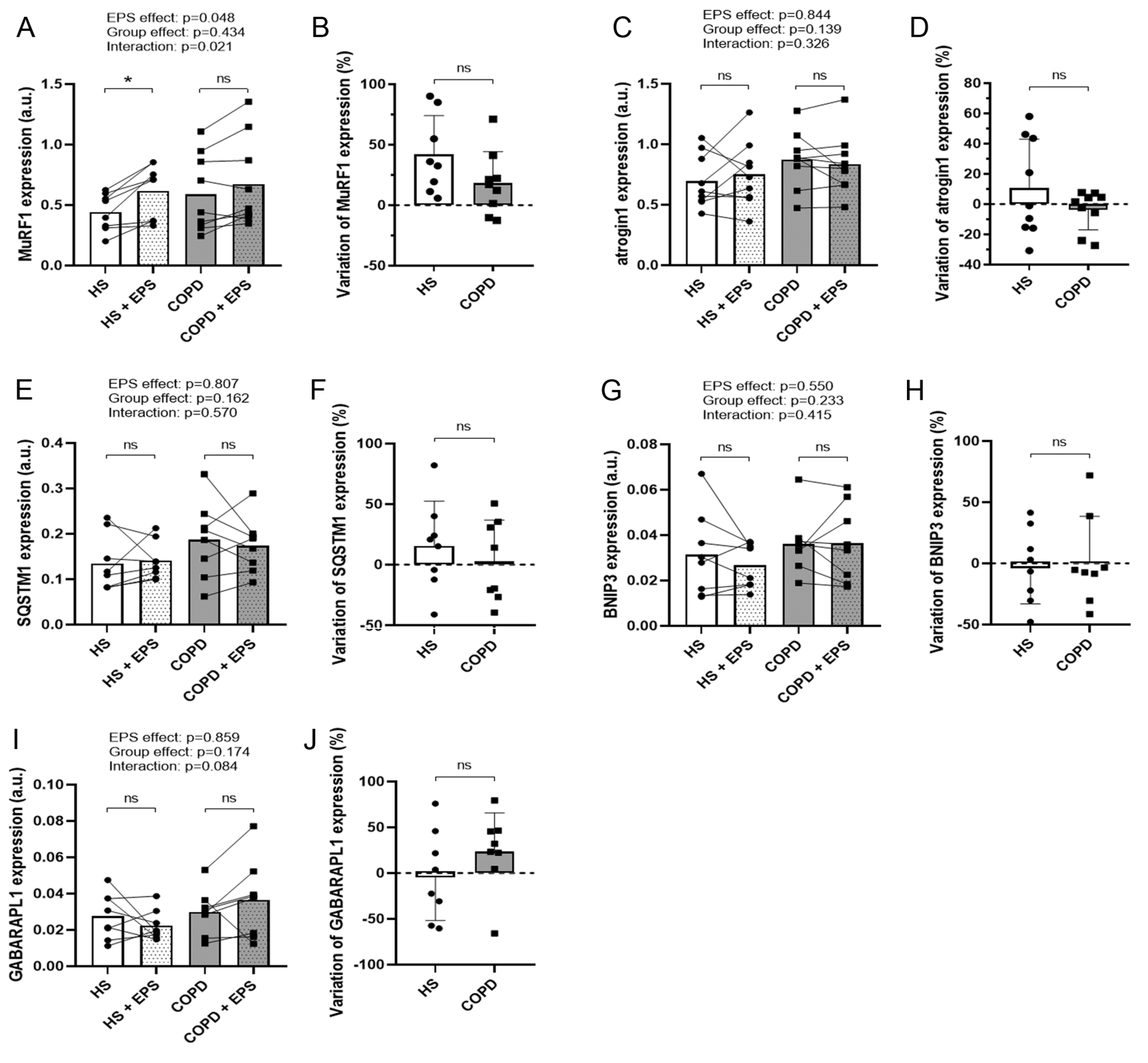
3.3. Effects of EPS on Protein Degradation and Synthesis Pathways
3.4. Effects of EPS on Differentiation Marker Expression
3.5. Effects of EPS on Mitochondrial Biogenesis
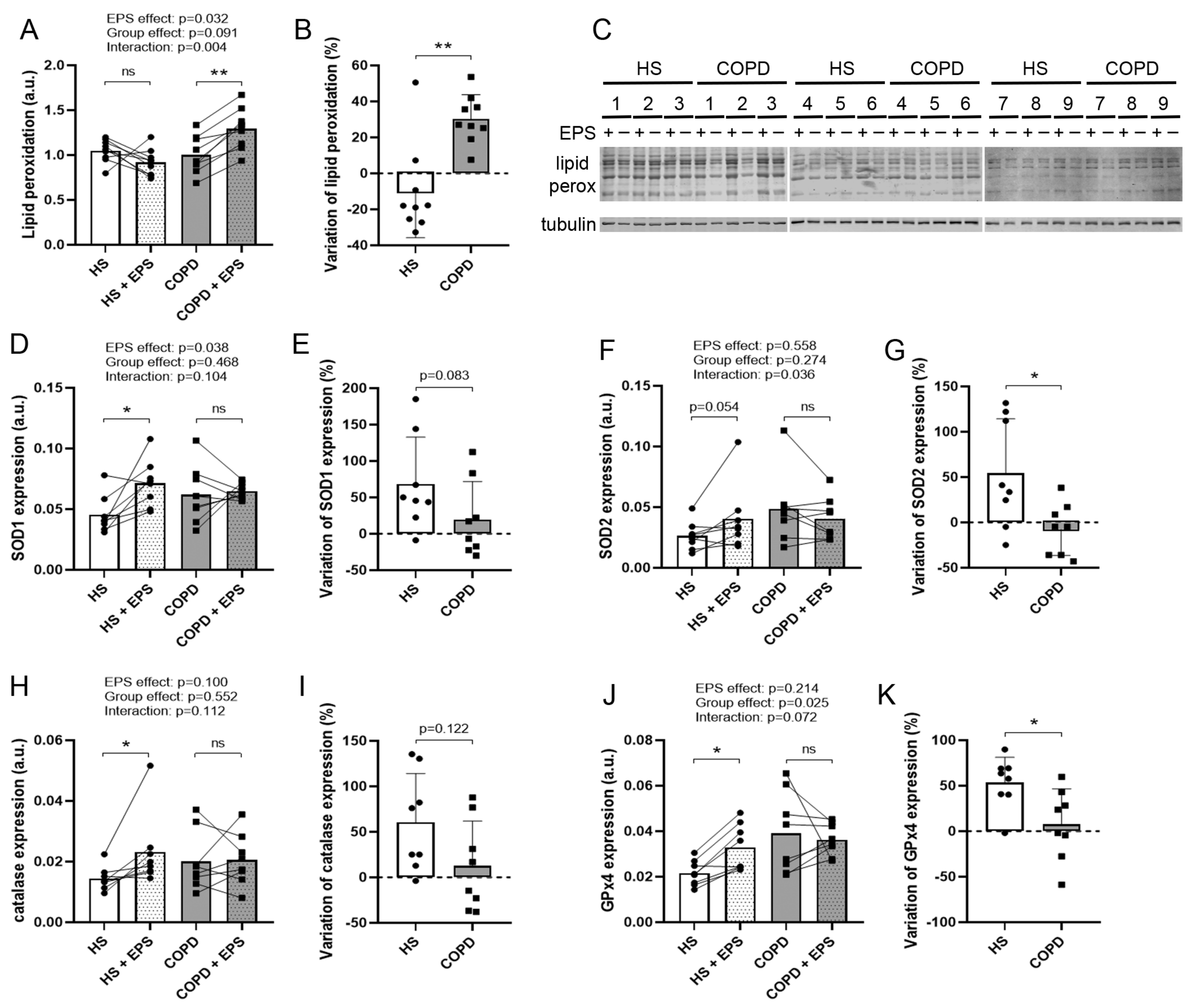
3.6. Effects of EPS on Oxidative Stress and Antioxidant Enzyme Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mostert, R.; Goris, A.; Weling-Scheepers, C.; Wouters, E.; Schols, A. Tissue depletion and health related quality of life in patients with chronic obstructive pulmonary disease. Respir. Med. 2000, 94, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Swallow, E.B.; Reyes, D.; Hopkinson, N.; Man, W.D.-C.; Porcher, R.; Cetti, E.J.; Moore, A.J.; Moxham, J.; Polkey, M.I. Quadriceps strength predicts mortality in patients with moderate to severe chronic obstructive pulmonary disease. Thorax 2007, 62, 115–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agustí, A.; Edwards, L.D.; Rennard, S.I.; MacNee, W.; Tal-Singer, R.; Miller, B.E.; Vestbo, J.; Lomas, D.A.; Calverley, P.M.A.; Wouters, E.; et al. Persistent Systemic Inflammation is Associated with Poor Clinical Outcomes in COPD: A Novel Phenotype. PLoS ONE 2012, 7, e37483. [Google Scholar] [CrossRef]
- Couillard, A.; Maltais, F.; Saey, D.; Debigaré, R.; Michaud, A.; Koechlin, C.; LeBlanc, P.; Préfaut, C. Exercise-induced Quadriceps Oxidative Stress and Peripheral Muscle Dysfunction in Patients with Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2003, 167, 1664–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blervaque, L.; Passerieux, E.; Pomiès, P.; Catteau, M.; Héraud, N.; Blaquière, M.; Bughin, F.; Ayoub, B.; Molinari, N.; Cristol, J.-P.; et al. Impaired training-induced angiogenesis process with loss of pericyte-endothelium interactions is associated with an abnormal capillary remodelling in the skeletal muscle of COPD patients. Respir. Res. 2019, 20, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandre, F.; Héraud, N.; Tremey, E.; Oliver, N.; Bourgouin, D.; Varray, A. Specific motor cortex hypoexcitability and hypoactivation in COPD patients with peripheral muscle weakness. BMC Pulm. Med. 2020, 20, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gundersen, K. Excitation-transcription coupling in skeletal muscle: The molecular pathways of exercise. Biol. Rev. 2010, 86, 564–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.; Chen, J. Mammalian Target of Rapamycin (mTOR) Signaling Network in Skeletal Myogenesis. J. Biol. Chem. 2012, 287, 43928–43935. [Google Scholar] [CrossRef] [Green Version]
- Zanou, B.N.; Gailly, P. Skeletal muscle hypertrophy and regeneration: Interplay between the myogenic regulatory factors (MRFs) and insulin-like growth factors (IGFs) pathways. Cell. Mol. Life Sci. 2013, 70, 4117–4130. [Google Scholar] [CrossRef]
- Casuso, R.A.; Huertas, J.R. The emerging role of skeletal muscle mitochondrial dynamics in exercise and ageing. Ageing Res. Rev. 2020, 58, 101025. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Mendoza, N.; Morales-González, Á.; Madrigal-Santillán, E.O.; Madrigal-Bujaidar, E.; Álvarez-González, I.; García-Melo, L.F.; Anguiano-Robledo, L.; Fregoso-Aguilar, T.; Morales-Gonzalez, J.A. Antioxidant and Adaptative Response Mediated by Nrf2 during Physical Exercise. Antioxidants 2019, 8, 196. [Google Scholar] [CrossRef] [Green Version]
- Nikolić, N.; Bakke, S.S.; Kase, E.T.; Rudberg, I.; Halle, I.F.; Rustan, A.C.; Thoresen, G.H.; Aas, V. Electrical Pulse Stimulation of Cultured Human Skeletal Muscle Cells as an In Vitro Model of Exercise. PLoS ONE 2012, 7, e33203. [Google Scholar] [CrossRef] [Green Version]
- Burch, N.; Arnold, A.-S.; Item, F.; Summermatter, S.; Santos, G.B.S.; Christe, M.; Boutellier, U.; Toigo, M.; Handschin, C. Electric Pulse Stimulation of Cultured Murine Muscle Cells Reproduces Gene Expression Changes of Trained Mouse Muscle. PLoS ONE 2010, 5, e10970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, C.S.; Christensen, D.P.; Lundh, M.; Dahllöf, M.S.; Haase, T.N.; Velasquez, J.M.; Laye, M.J.; Mandrup-Poulsen, T.; Solomon, T.P.J. Skeletal Muscle to Pancreatic β-Cell Cross-talk: The Effect of Humoral Mediators Liberated by Muscle Contraction and Acute Exercise on β-Cell Apoptosis. J. Clin. Endocrinol. Metab. 2015, 100, E1289–E1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheler, M.; Irmler, M.; Lehr, S.; Hartwig, S.; Staiger, H.; Al-Hasani, H.; Beckers, J.; de Angelis, M.H.; Häring, H.-U.; Weigert, C. Cytokine response of primary human myotubes in an in vitro exercise model. Am. J. Physiol. Physiol. 2013, 305, C877–C886. [Google Scholar] [CrossRef]
- Tarum, J.; Folkesson, M.; Atherton, P.J.; Kadi, F. Electrical pulse stimulation: Anin vitroexercise model for the induction of human skeletal muscle cell hypertrophy. A proof-of-concept study. Exp. Physiol. 2017, 102, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Gouzi, F.; Blaquière, M.; Catteau, M.; Bughin, F.; Maury, J.; Passerieux, E.; Ayoub, B.; Mercier, J.; Hayot, M.; Pomiès, P. Oxidative stress regulates autophagy in cultured muscle cells of patients with chronic obstructive pulmonary disease. J. Cell. Physiol. 2018, 233, 9629–9639. [Google Scholar] [CrossRef] [PubMed]
- Pomiès, P.; Rodriguez, J.; Blaquière, M.; Sedraoui, S.; Gouzi, F.; Carnac, G.; Laoudj-Chenivesse, D.; Mercier, J.; Préfaut, C.; Hayot, M. Reduced myotube diameter, atrophic signalling and elevated oxidative stress in cultured satellite cells from COPD patients. J. Cell. Mol. Med. 2014, 19, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Pomiès, P.; Blaquière, M.; Maury, J.; Mercier, J.; Gouzi, F.; Hayot, M. Involvement of the FoxO1/MuRF1/Atrogin-1 Signaling Pathway in the Oxidative Stress-Induced Atrophy of Cultured Chronic Obstructive Pulmonary Disease Myotubes. PLoS ONE 2016, 11, e0160092. [Google Scholar] [CrossRef] [Green Version]
- Maury, J.; Gouzi, F.; De Rigal, P.; Heraud, N.; Pincemail, J.; Molinari, N.; Pomiès, P.; Laoudj-Chenivesse, D.; Mercier, J.; Prefaut, C.; et al. Heterogeneity of Systemic Oxidative Stress Profiles in COPD: A Potential Role of Gender. Oxidative Med. Cell. Longev. 2015, 2015, 201843. [Google Scholar] [CrossRef] [PubMed]
- Blervaque, L.; Pomiès, P.; Rossi, E.; Catteau, M.; Blandinières, A.; Passerieux, E.; Blaquière, M.; Ayoub, B.; Molinari, N.; Mercier, J.; et al. COPD is deleterious for pericytes: Implications during training-induced angiogenesis in skeletal muscle. Am. J. Physiol. Circ. Physiol. 2020, 319, H1142–H1151. [Google Scholar] [CrossRef] [PubMed]
- ATS. Committee on Proficiency Standards for Clinical Pulmonary Function Laboratories ATS Statement. Am. J. Respir. Crit. Care Med. 2002, 166, 111–117. [Google Scholar] [CrossRef]
- Koechlin, C.; Couillard, A.; Simar, D.; Cristol, J.P.; Bellet, H.; Hayot, M.; Prefaut, C. Does Oxidative Stress Alter Quadriceps Endurance in Chronic Obstructive Pulmonary Disease? Am. J. Respir. Crit. Care Med. 2004, 169, 1022–1027. [Google Scholar] [CrossRef] [PubMed]
- Hayot, M.; Michaud, A.; Koechlin, C.; Caron, M.-A.; Leblanc, P.; Prefaut, C.; Maltais, F. Skeletal muscle microbiopsy: A validation study of a minimally invasive technique. Eur. Respir. J. 2005, 25, 431–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catteau, M.; Gouzi, F.; Blervaque, L.; Passerieux, E.; Blaquière, M.; Ayoub, B.; Bughin, F.; Mercier, J.; Hayot, M.; Pomiès, P. Effects of a human microenvironment on the differentiation of human myoblasts. Biochem. Biophys. Res. Commun. 2020, 525, 968–973. [Google Scholar] [CrossRef]
- Hawke, T.J.; Meeson, A.P.; Jiang, N.; Graham, S.; Hutcheson, K.; DiMaio, J.M.; Garry, D.J. p21 is essential for normal myogenic progenitor cell function in regenerating skeletal muscle. Am. J. Physiol. Physiol. 2003, 285, C1019–C1027. [Google Scholar] [CrossRef] [PubMed]
- Urashima, M.; Teoh, G.; Akiyama, M.; Yuza, Y.; Anderson, K.C.; Maekawa, K. Restoration of p16INK4A protein induces myogenic differentiation in RD rhabdomyosarcoma cells. Br. J. Cancer 1999, 79, 1032–1036. [Google Scholar] [CrossRef] [Green Version]
- Nikolić, N.; Görgens, S.W.; Thoresen, G.H.; Aas, V.; Eckel, J.; Eckardt, K. Electrical pulse stimulation of cultured skeletal muscle cells as a model forin vitroexercise - possibilities and limitations. Acta Physiol. 2016, 220, 310–331. [Google Scholar] [CrossRef] [PubMed]
- Lambernd, S.; Taube, A.; Schober, A.; Platzbecker, B.; Görgens, S.W.; Schlich, R.; Jeruschke, K.; Weiss, J.; Eckardt, K.; Eckel, J. Contractile activity of human skeletal muscle cells prevents insulin resistance by inhibiting pro-inflammatory signalling pathways. Diabetologia 2012, 55, 1128–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, Y.H.; Lee, S.-M.; Lee, S.H.; Yoon, J.H.; Kang, J.S.; Yang, Y.R.; Kwon, K.-S. Comparative molecular analysis of endurance exercise in vivo with electrically stimulated in vitro myotube contraction. J. Appl. Physiol. 2019, 127, 1742–1753. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Nedachi, T.; Kanzaki, M. Accelerated de novo sarcomere assembly by electric pulse stimulation in C2C12 myotubes. Exp. Cell Res. 2007, 313, 1853–1865. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liang, X.; Shan, T.; Jiang, Q.; Deng, C.; Zheng, R.; Kuang, S. mTOR is necessary for proper satellite cell activity and skeletal muscle regeneration. Biochem. Biophys. Res. Commun. 2015, 463, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.; Ji, L.L. Role of PGC-1α signaling in skeletal muscle health and disease. Ann. N. Y. Acad. Sci. 2012, 1271, 110–117. [Google Scholar] [CrossRef]
- Yan, Z. Exercise, PGC-1α, and metabolic adaptation in skeletal muscle. This paper article is one of a selection of papers published in this Special Issue, entitled 14th International Biochemistry of Exercise Conference—Muscles as Molecular and Metabolic Machines, and has undergone the Journal’s usual peer review process. Appl. Physiol. Nutr. Metab. 2009, 34, 424–427. [Google Scholar] [CrossRef] [Green Version]
- Remels, A.H.; Schrauwen, P.; Broekhuizen, R.; Willems, J.; Kersten, S.; Gosker, H.R.; Schols, A.M. Peroxisome proliferator-activated receptor expression is reduced in skeletal muscle in COPD. Eur. Respir. J. 2007, 30, 245–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittom, F.; Jobin, J.; Simard, P.M.; Leblanc, P.; Simard, C.; Bernard, S.; Belleau, R.; Maltais, F. Histochemical and morphological characteristics of the vastus lateralis muscle in patients with chronic obstructive pulmonary disease. Med. Sci. Sports Exerc. 1998, 30, 1467–1474. [Google Scholar] [CrossRef]
- Gouzi, F.; Préfaut, C.; Abdellaoui, A.; Roudier, E.; De Rigal, P.; Molinari, N.; Laoudj-Chenivesse, D.; Mercier, J.; Birot, O.; Hayot, M. Blunted muscle angiogenic training-response in COPD patientsversussedentary controls. Eur. Respir. J. 2012, 41, 806–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gifford, J.R.; Trinity, J.D.; Kwon, O.-S.; Layec, G.; Garten, R.S.; Park, S.-Y.; Nelson, A.D.; Richardson, R.S. Altered skeletal muscle mitochondrial phenotype in COPD: Disease vs. disuse. J. Appl. Physiol. 2018, 124, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Cole-Ezea, P.; Swan, D.; Shanley, D.; Hesketh, J. Glutathione peroxidase 4 has a major role in protecting mitochondria from oxidative damage and maintaining oxidative phosphorylation complexes in gut epithelial cells. Free. Radic. Biol. Med. 2012, 53, 488–497. [Google Scholar] [CrossRef]
- Lustgarten, M.S.; Jang, Y.C.; Liu, Y.; Muller, F.L.; Qi, W.; Steinhelper, M.; Brooks, S.V.; Larkin, L.; Shimizu, T.; Shirasawa, T.; et al. Conditional knockout of Mn-SOD targeted to type IIB skeletal muscle fibers increases oxidative stress and is sufficient to alter aerobic exercise capacity. Am. J. Physiol. Physiol. 2009, 297, C1520–C1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kneppers, A.; Verdijk, L.; De Theije, C.; Corten, M.; Gielen, E.; Van Loon, L.; Schols, A.; Langen, R. A novel in vitro model for the assessment of postnatal myonuclear accretion. Skelet. Muscle 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orfanos, Z.; Gödderz, M.P.O.; Soroka, E.; Gödderz, T.; Rumyantseva, A.; van der Ven, P.; Hawke, T.J.; Fürst, D.O. Breaking sarcomeres by in vitro exercise. Sci. Rep. 2016, 6, 19614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.Z.; Nikolić, N.; Bakke, S.S.; Kase, E.T.; Guderud, K.; Hjelmesæth, J.; Aas, V.; Rustan, A.C.; Thoresen, G.H. Myotubes from lean and severely obese subjects with and without type 2 diabetes respond differently to an in vitro model of exercise. Am. J. Physiol. Physiol. 2015, 308, C548–C556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.E.; Jones, D.E.; Walker, M.; Newton, J.L. Abnormalities of AMPK Activation and Glucose Uptake in Cultured Skeletal Muscle Cells from Individuals with Chronic Fatigue Syndrome. PLoS ONE 2015, 10, e0122982. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Subjects | Patients with COPD | p-Value | |
|---|---|---|---|
| n | 9 | 9 | - |
| Gender (M/F) | 7/2 | 6/3 | - |
| Age (yrs) | 62.3 ± 6.4 | 58.0 ± 6.5 | 0.180 |
| BMI (kg/m2) | 25.5 ± 3.6 | 20.5 ± 3.1 | 0.006 |
| FFMI (kg/m2) | 18.5 ± 2.0 | 16.7 ± 2.2 | 0.110 |
| FEV1 (% pred.) | 102.4 ± 14.7 | 36.3 ± 14.6 | <0.0001 |
| 6MWD (% pred.) | 93.4 ± 11.8 | 62.5 ± 16.3 | 0.0003 |
| qMVC (kg) | 36.5 ± 9.7 1 | 18.0 ± 6.9 2 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catteau, M.; Passerieux, E.; Blervaque, L.; Gouzi, F.; Ayoub, B.; Hayot, M.; Pomiès, P. Response to Electrostimulation Is Impaired in Muscle Cells from Patients with Chronic Obstructive Pulmonary Disease. Cells 2021, 10, 3002. https://doi.org/10.3390/cells10113002
Catteau M, Passerieux E, Blervaque L, Gouzi F, Ayoub B, Hayot M, Pomiès P. Response to Electrostimulation Is Impaired in Muscle Cells from Patients with Chronic Obstructive Pulmonary Disease. Cells. 2021; 10(11):3002. https://doi.org/10.3390/cells10113002
Chicago/Turabian StyleCatteau, Matthias, Emilie Passerieux, Léo Blervaque, Farés Gouzi, Bronia Ayoub, Maurice Hayot, and Pascal Pomiès. 2021. "Response to Electrostimulation Is Impaired in Muscle Cells from Patients with Chronic Obstructive Pulmonary Disease" Cells 10, no. 11: 3002. https://doi.org/10.3390/cells10113002
APA StyleCatteau, M., Passerieux, E., Blervaque, L., Gouzi, F., Ayoub, B., Hayot, M., & Pomiès, P. (2021). Response to Electrostimulation Is Impaired in Muscle Cells from Patients with Chronic Obstructive Pulmonary Disease. Cells, 10(11), 3002. https://doi.org/10.3390/cells10113002