
The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration


Abstract
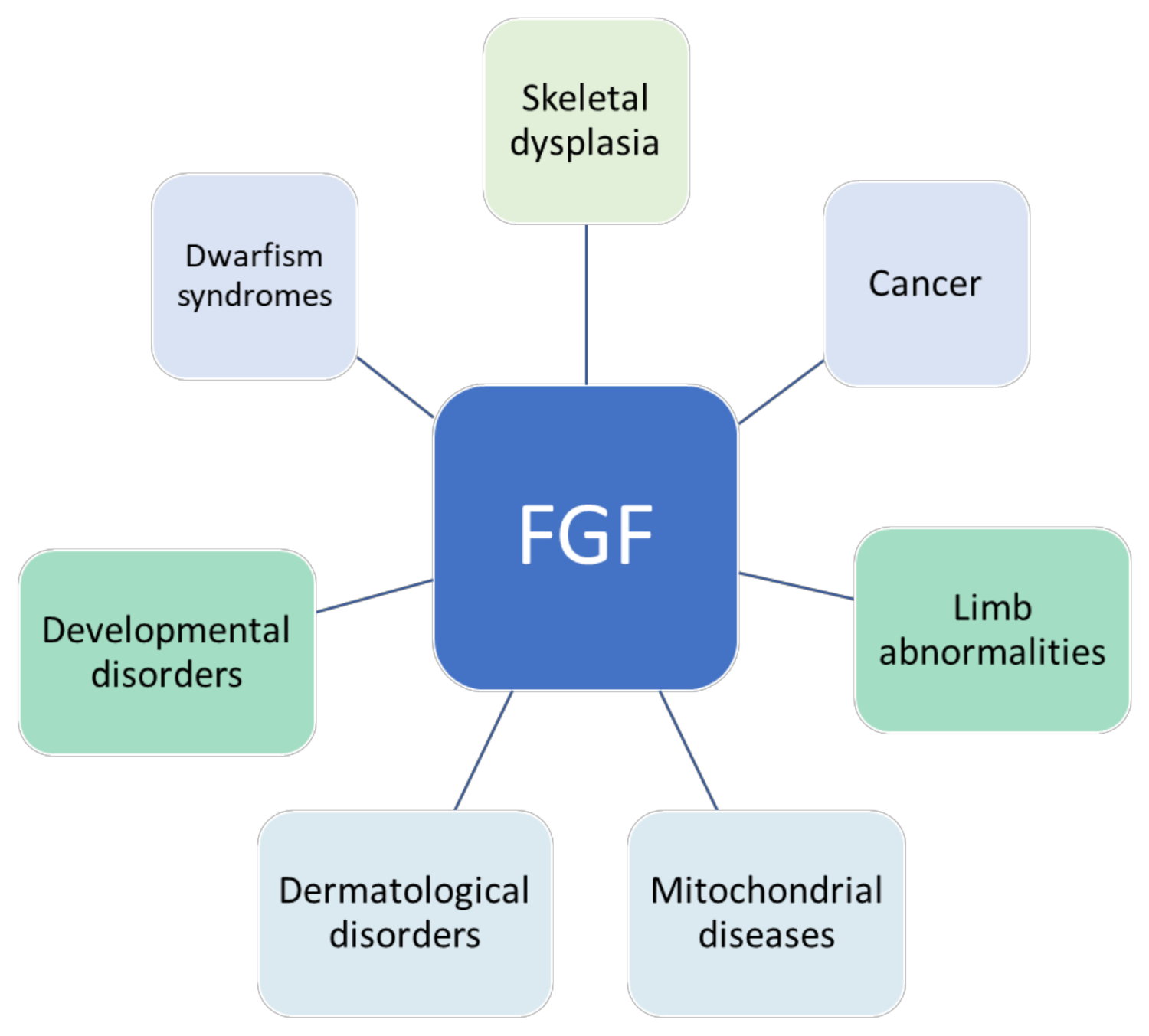
:1. Introduction
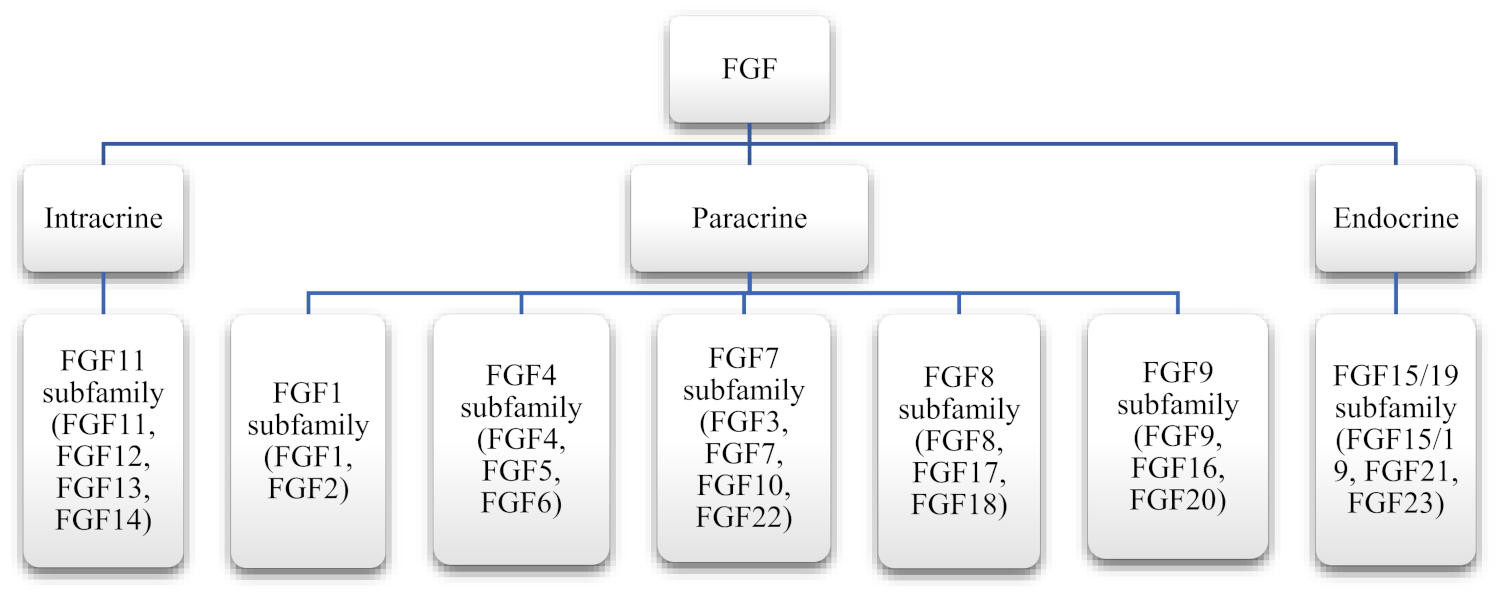
2. Fibroblast Growth Factor Family
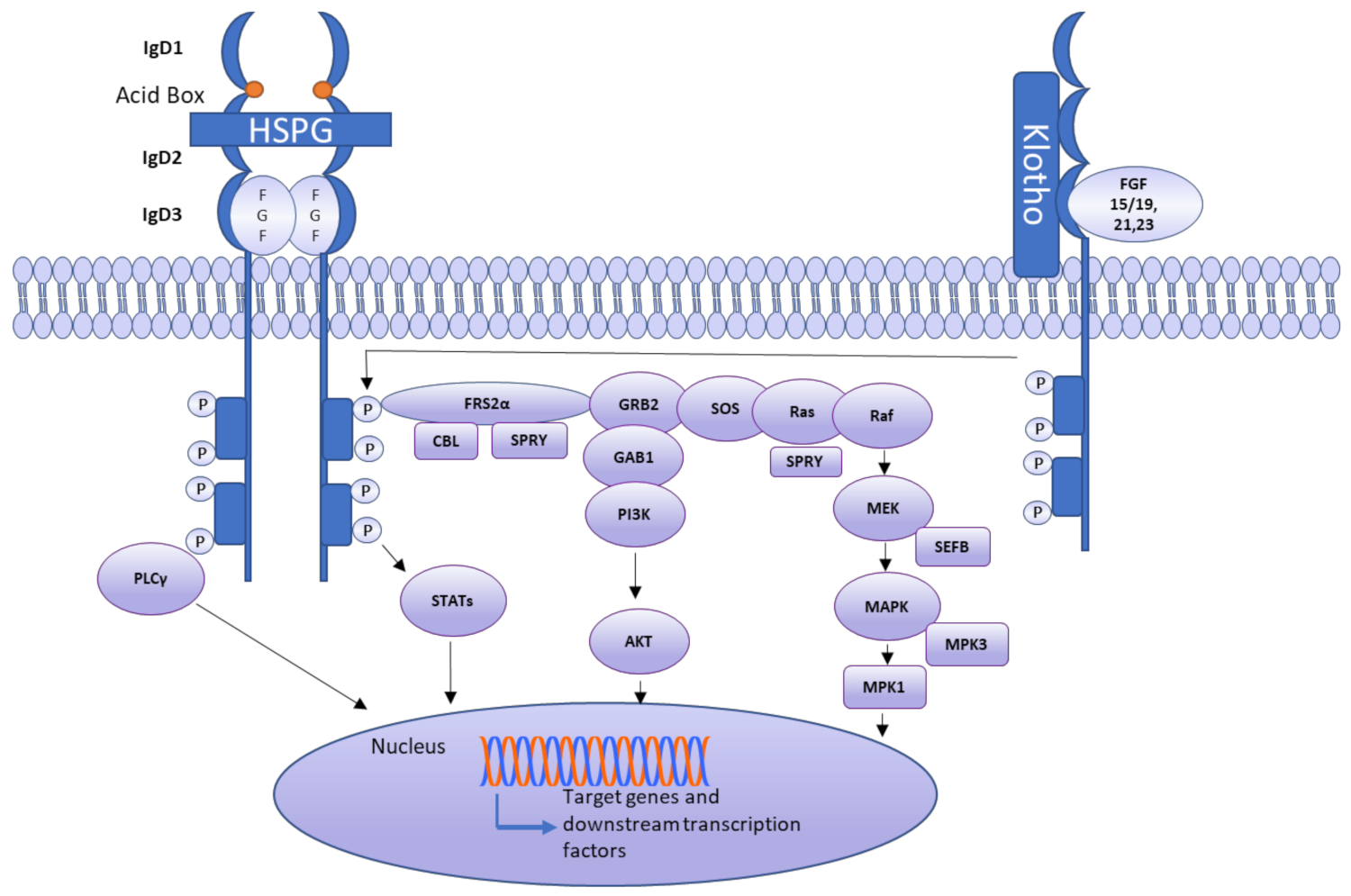
3. Receptor Activation Mechanism
Autocrine/Paracrine and Endocrine Signaling in Different FGF
4. FGF Mediation of Tissue Repair and Regeneration
4.1. General Mechanisms of Tissue Repair and Regeneration
4.2. Fibroblast Growth Factor in Regeneration
4.3. The Role of FGFs in Tissue Repair
4.4. Endocrine FGFs in Repair Processes
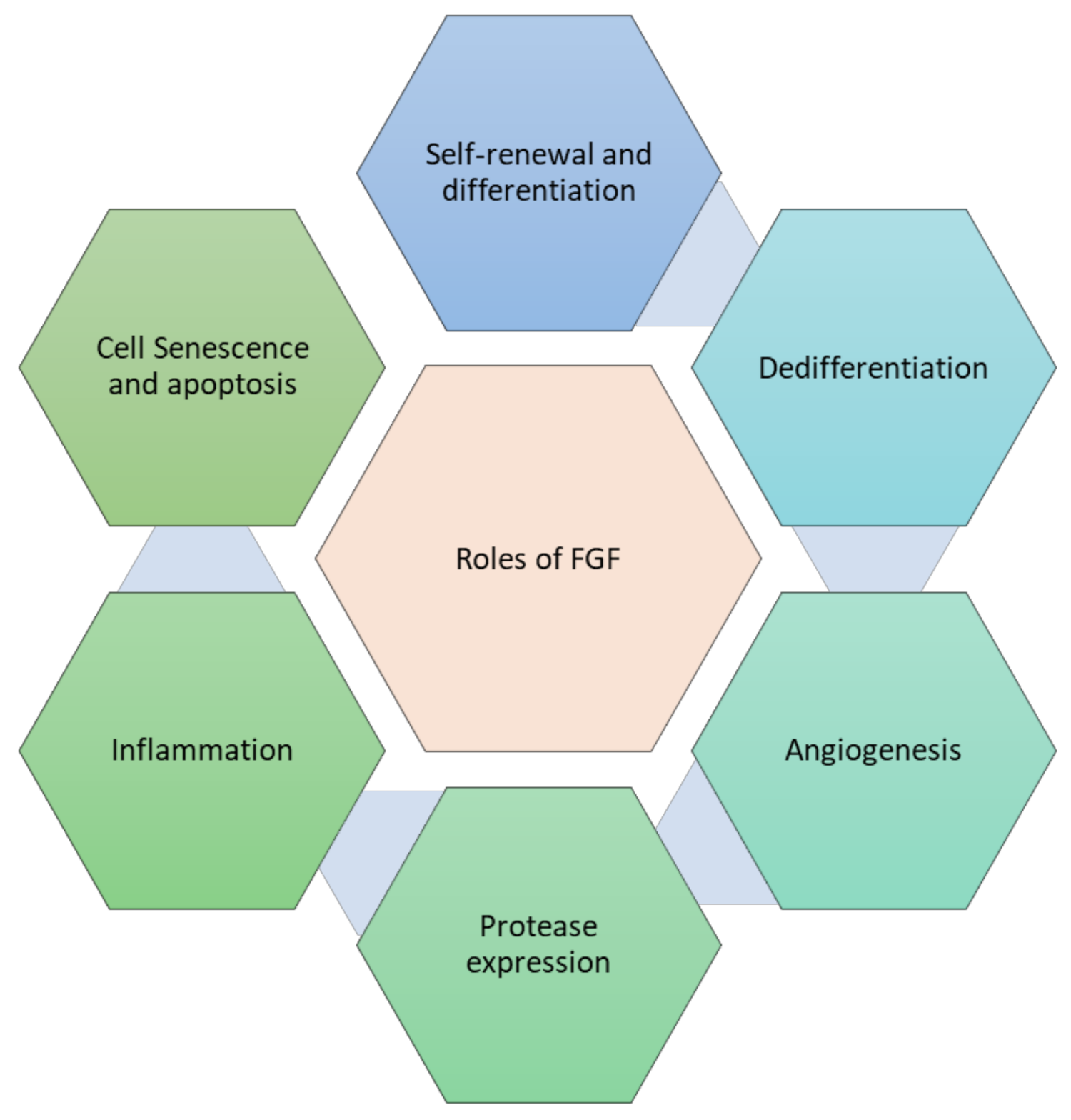
4.5. Cell Processes Underlying the Stimulation of Tissue Repair by Fibroblast Growth Factor
4.5.1. Self-Renewal and De-Differentiation
4.5.2. Effect on Proliferation
4.5.3. Suppression of Cell Senescence and Apoptosis
4.5.4. Regulation of Inflammation
4.5.5. Induction of Angiogenesis
4.5.6. FGF Impact on Protease Expression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| AP-1 | activating protein 1 |
| bFGF | basic fibroblast growth factor (FGF2) |
| EGF | epidermal growth factor |
| ERK | extracellular signal-regulated kinase |
| FGF | fibroblast growth factor |
| G-CSF | granulocyte colony-stimulating factor |
| GM-CSF | granulocyte macrophage colony-stimulating factor |
| HS | heparan sulfate |
| IL-1β | interleukin 1β |
| IL-6 | interleukin 6 |
| JNK | c-Jun N-terminal kinase |
| KGF | keratinocyte growth factor |
| MAPK | mitogen-activated protein kinase |
| MMP | matrix metalloproteinase |
| NF-κB | nuclear factor kappa B |
| PDGF | platelet-derived growth factor |
| PI3K | phosphoinositol-3 kinase |
| SEF | similar expression to FGF |
| STAT | signal transducer and activator of transcription |
| TGF | transforming growth factor |
| TNF-α | tumor necrosis factor alpha |
| VEGF | vascular endothelial growth factor |
References
- Ardizzone, A.; Scuderi, S.A.; Giuffrida, D.; Colarossi, C.; Puglisi, C.; Campolo, M.; Cuzzocrea, S.; Esposito, E.; Paterniti, I. Role of Fibroblast Growth Factors Receptors (FGFRs) in Brain Tumors, Focus on Astrocytoma and Glioblastoma. Cancers 2020, 12, 3825. [Google Scholar] [CrossRef]
- Litwack, G. Chapter 16—Steroid Hormones. In Human Biochemistry; Litwack, G., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 467–506. ISBN 978-0-12-383864-3. [Google Scholar]
- Gadaleta, R.M.; Moschetta, A. Metabolic Messengers: Fibroblast growth factor 15/19. Nat. Metab. 2019, 1, 588–594. [Google Scholar] [CrossRef]
- Sarabipour, S.; Hristova, K. Mechanism of FGF receptor dimerization and activation. Nat. Commun. 2016, 7, 10262. [Google Scholar] [CrossRef]
- Katoh, M. Fibroblast growth factor receptors as treatment targets in clinical oncology. Nat. Rev. Clin. Oncol. 2019, 16, 105–122. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Z.; Lin, X.; Beenken, A.; Eliseenkova, A.V.; Mohammadi, M.; Linhardt, R.J. Compositional analysis on heparin/heparan sulfate interacting with FGF•FGFR complexes. Biochemistry 2009, 48, 8379–8386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Su, N.; Yang, J.; Tan, Q.; Huang, S.; Jin, M.; Ni, Z.; Zhang, B.; Zhang, D.; Luo, F.; et al. FGF/FGFR signaling in health and disease. Signal Transduct. Target. Ther. 2020, 5, 1–38. [Google Scholar] [CrossRef]
- Katoh, M. Therapeutics Targeting FGF Signaling Network in Human Diseases. Trends Pharmacol. Sci. 2016, 37, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Scholle, L.M.; Lehmann, D.; Deschauer, M.; Kraya, T.; Zierz, S. FGF-21 as a Potential Biomarker for Mitochondrial Diseases. Curr. Med. Chem. 2018, 25, 2070–2081. [Google Scholar] [CrossRef]
- Stanier, P.; Pauws, E. Development of the Lip and Palate: FGF Signalling. Tooth Mov. Oral Biol. 2012, 16, 71–80. [Google Scholar] [CrossRef]
- Itoh, N.; Ohta, H.; Nakayama, Y.; Konishi, M. Roles of FGF Signals in Heart Development, Health, and Disease. Front. Cell Dev. Biol. 2016, 4, 110. [Google Scholar] [CrossRef] [Green Version]
- Fukagawa, M.; Kazama, J.J. FGF23: Its role in renal bone disease. Pediatr. Nephrol. 2006, 21, 1802–1806. [Google Scholar] [CrossRef]
- Guo, Y.-C.; Yuan, Q. Fibroblast growth factor 23 and bone mineralisation. Int. J. Oral Sci. 2015, 7, 8–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, E.; Yoshida, M.; Sasaki, S. Applicability of fibroblast growth factor 23 for evaluation of risk of vertebral fracture and chronic kidney disease-mineral bone disease in elderly chronic kidney disease patients. BMC Nephrol. 2012, 13, 122. [Google Scholar] [CrossRef] [Green Version]
- Kurpas, A.; Supeł, K.; Idzikowska, K.; Zielińska, M. FGF23: A Review of Its Role in Mineral Metabolism and Renal and Cardiovascular Disease. Dis. Markers 2021, 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ping, Q.; Yan, R.; Cheng, X.; Wang, W.; Zhong, Y.; Hou, Z.; Shi, Y.; Wang, C.; Li, R. Cancer-associated fibroblasts: Overview, progress, challenges, and directions. Cancer Gene Ther. 2021, 28, 984–999. [Google Scholar] [CrossRef]
- Bai, Y.; Shang, K.; Chen, H.; Ding, F.; Wang, Z.; Liang, C.; Xu, Y.; Sun, M.; Li, Y. FGF -1/-3/ FGFR 4 signaling in cancer-associated fibroblasts promotes tumor progression in colon cancer through Erk and MMP -7. Cancer Sci. 2015, 106, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Tovar, V.; Cornella, H.; Moeini, A.; Vidal, S.; Hoshida, Y.; Sia, D.; Peix, J.; Cabellos, L.; Alsinet, C.; Torrecilla, S.; et al. Tumour initiating cells and IGF/FGF signalling contribute to sorafenib resistance in hepatocellular carcinoma. Gut 2017, 66, 530–540. [Google Scholar] [CrossRef]
- Högler, W.; Ward, L.M. New developments in the management of achondroplasia. Wien. Med. Wochenschr. 2020, 170, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Hatzaki, A.; Sifakis, S.; Apostolopoulou, D.; Bouzarelou, D.; Konstantinidou, A.; Kappou, D.; Sideris, A.; Tzortzis, E.; Athanassiadis, A.; Florentin, L.; et al. FGFR3related skeletal dysplasias diagnosed prenatally by ultrasonography and molecular analysis: Presentation of 17 cases. Am. J. Med. Genet. Part A 2011, 155, 2426–2435. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Marie, P.J. Fibroblast growth factor signaling in skeletal development and disease. Genes Dev. 2015, 29, 1463–1486. [Google Scholar] [CrossRef] [Green Version]
- Presta, M.; Chiodelli, P.; Giacomini, A.; Rusnati, M.; Ronca, R. Fibroblast growth factors (FGFs) in cancer: FGF traps as a new therapeutic approach. Pharmacol. Ther. 2017, 179, 171–187. [Google Scholar] [CrossRef] [PubMed]
- DiPietro, L.A. Angiogenesis and wound repair: When enough is enough. J. Leukoc. Biol. 2016, 100, 979–984. [Google Scholar] [CrossRef]
- Chuma, H.; Mizuta, H.; Kudo, S.; Takagi, K.; Hiraki, Y. One day exposure to FGF-2 was sufficient for the regenerative repair of full-thickness defects of articular cartilage in rabbits. Osteoarthr. Cartil. 2004, 12, 834–842. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, J.; Xie, Y.; Wang, L.; Yang, L.; Yu, J.; Miyamoto, A.; Sun, F. Development of FGF-2-loaded electrospun waterborne polyurethane fibrous membranes for bone regeneration. Regen. Biomater. 2021, 8, rbaa046. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Huegel, J.; Taylor, B.L.; Nuss, C.A.; Weiss, S.N.; Soslowsky, L.J.; Mauck, R.L.; Kuntz, A.F. Biocompatibility and bioactivity of an FGF-loaded microsphere-based bilayer delivery system. Acta Biomater. 2020, 111, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Firoozi, N.; Kang, Y. Immobilization of FGF on Poly(xylitol dodecanedioic Acid) Polymer for Tissue Regeneration. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Trowell, O.A.; Willmer, E.N. Studies on the Growth of Tissues in vitro VI. The Effects of some Tissue Extracts on the Growth of Periosteal Fibroblasts. J. Exp. Biol. 1939, 16, 60–70. [Google Scholar] [CrossRef]
- Armelin, H.A. Pituitary Extracts and Steroid Hormones in the Control of 3T3 Cell Growth. Proc. Natl. Acad. Sci. USA 1973, 70, 2702–2706. [Google Scholar] [CrossRef] [Green Version]
- Gospodarowicz, D. Localisation of a fibroblast growth factor and its effect alone and with hydrocortisone on 3T3 cell growth. Nat. Cell Biol. 1974, 249, 123–127. [Google Scholar] [CrossRef]
- Burrus, L.W.; Olwin, B.B. Isolation of a Receptor for Acidic and Basic Fibroblast Growth Factor from Embryonic Chick. J. Biol. Chem. 1989, 264, 18647–18653. [Google Scholar] [CrossRef]
- Beenken, A.; Mohammadi, M. The FGF family: Biology, pathophysiology and therapy. Nat. Rev. Drug Discov. 2009, 8, 235–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, N.; Ornitz, D. Functional evolutionary history of the mouseFgf gene family. Dev. Dyn. 2008, 237, 18–27. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, N.; Ornitz, D. Fibroblast growth factors: From molecular evolution to roles in development, metabolism and disease. J. Biochem. 2010, 149, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antoine, M.; Wirz, W.; Tag, C.G.; Mavituna, M.; Emans, N.; Korff, T.; Stoldt, V.; Gressner, A.M.; Kiefer, P. Expression pattern of fibroblast growth factors (FGFs), their receptors and antagonists in primary endothelial cells and vascular smooth muscle cells. Growth Factors 2005, 23, 87–95. [Google Scholar] [CrossRef]
- Brewer, J.R.; Mazot, P.; Soriano, P. Genetic insights into the mechanisms of Fgf signaling. Genes Dev. 2016, 30, 751–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, M.; Wu, X.; Wang, F.; McKeehan, W.L. Specificity for Fibroblast Growth Factors Determined by Heparan Sulfate in a Binary Complex with the Receptor Kinase. J. Biol. Chem. 1999, 274, 15947–15952. [Google Scholar] [CrossRef] [Green Version]
- Chellaiah, A.; Yuan, W.; Chellaiah, M.; Ornitz, D.M. Mapping Ligand Binding Domains in Chimeric Fibroblast Growth Factor Receptor Molecules: Multiple regions determine ligand binding specificity. J. Biol. Chem. 1999, 274, 34785–34794. [Google Scholar] [CrossRef] [Green Version]
- Kharitonenkov, A.; Dunbar, J.D.; Bina, H.A.; Bright, S.; Moyers, J.S.; Zhang, C.; Ding, L.; Micanovic, R.; Mehrbod, S.F.; Knierman, M.; et al. FGF-21/FGF-21 receptor interaction and activation is determined by βKlotho. J. Cell. Physiol. 2008, 215, 1–7. [Google Scholar] [CrossRef]
- Pellegrini, L.; Burke, D.; Von Delft, F.; Mulloy, B.; Blundell, T.L. Crystal structure of fibroblast growth factor receptor ectodomain bound to ligand and heparin. Nat. Cell Biol. 2000, 407, 1029–1034. [Google Scholar] [CrossRef]
- Miura, K.; Oba, T.; Hamanaka, K.; Ito, K.-I. FGF2-FGFR1 pathway activation together with thymidylate synthase upregulation is induced in pemetrexed-resistant lung cancer cells. Oncotarget 2019, 10, 1171–1192. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, V.; Kinkl, N.; Léveillard, T.; Sahel, J.-A.; Hicks, D. Fibroblast Growth Factor Receptor 4 (FGFR4) Is Expressed in Adult Rat and Human Retinal Photoreceptors and Neurons. J. Mol. Neurosci. 1999, 13, 187–198. [Google Scholar] [CrossRef]
- Nayak, S.; Goel, M.M.; Makker, A.; Bhatia, V.; Chandra, S.; Kumar, S.; Agarwal, S.P. Fibroblast Growth Factor (FGF-2) and Its Receptors FGFR-2 and FGFR-3 May Be Putative Biomarkers of Malignant Transformation of Potentially Malignant Oral Lesions into Oral Squamous Cell Carcinoma. PLoS ONE 2015, 10, e0138801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fromme, J.E.; Schmitz, K.; Wachter, A.; Grzelinski, M.; Zielinski, D.; Koppel, C.; Conradi, L.-C.; Homayounfar, K.; Hugo, T.; Hugo, S.; et al. FGFR3 mRNA overexpression defines a subset of oligometastatic colorectal cancers with worse prognosis. Oncotarget 2018, 9, 32204–32218. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Sakamoto, H.; Terada, M.; Ochiya, T. Pleiotropic function of FGF-4: Its role in development and stem cells. Dev. Dyn. 2008, 238, 265–276. [Google Scholar] [CrossRef]
- Revest, J.-M.; Spencer-Dene, B.; Kerr, K.; De Moerlooze, L.; Rosewell, I.; Dickson, C. Fibroblast Growth Factor Receptor 2-IIIb Acts Upstream of Shh and Fgf4 and Is Required for Limb Bud Maintenance but Not for the Induction of Fgf8, Fgf10, Msx1, or Bmp4. Dev. Biol. 2001, 231, 47–62. [Google Scholar] [CrossRef] [Green Version]
- Holzmann, K.; Grunt, T.; Heinzle, C.; Sampl, S.; Steinhoff, H.; Reichmann, N.; Kleiter, M.; Hauck, M.; Marian, B. Alternative Splicing of Fibroblast Growth Factor Receptor IgIII Loops in Cancer. J. Nucleic Acids 2011, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becic, T.; Kero, D.; Vukojevic, K.; Mardesic, S.; Saraga-Babic, M. Growth factors FGF8 and FGF2 and their receptor FGFR1, transcriptional factors Msx-1 and MSX-2, and apoptotic factors p19 and RIP5 participate in the early human limb development. Acta Histochem. 2018, 120, 205–214. [Google Scholar] [CrossRef]
- Watson, J.; Francavilla, C. Regulation of FGF10 Signaling in Development and Disease. Front. Genet. 2018, 9, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, K.; Zhang, D.; Jia, Q.; Zhang, Q. Regulation of Fgf15 expression in the intestine by glucocorticoid receptor. Mol. Med. Rep. 2019, 19, 2953–2959. [Google Scholar] [CrossRef] [Green Version]
- Davidson, D.; Blanc, A.; Filion, D.; Wang, H.; Plut, P.; Pfeffer, G.; Buschmann, M.D.; Henderson, J.E. Fibroblast Growth Factor (FGF) 18 Signals through FGF Receptor 3 to Promote Chondrogenesis. J. Biol. Chem. 2005, 280, 20509–20515. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Cao, M.; Cai, Y.; Li, X.; Zhao, C.; Cui, R. Dissecting the Role of the FGF19-FGFR4 Signaling Pathway in Cancer Development and Progression. Front. Cell Dev. Biol. 2020, 8, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sleeman, I.J.; Boshoff, E.L.; Duty, S. Fibroblast growth factor-20 protects against dopamine neuron loss in vitro and provides functional protection in the 6-hydroxydopamine-lesioned rat model of Parkinson’s disease. Neuropharmacology 2012, 63, 1268–1277. [Google Scholar] [CrossRef]
- Zhang, X.; Ibrahimi, O.A.; Olsen, S.; Umemori, H.; Mohammadi, M.; Ornitz, D.M. Receptor Specificity of the Fibroblast Growth Factor Family: The complete mammalian fgf family. J. Biol. Chem. 2006, 281, 15694–15700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, Y.K.; Ranganath, K.; Hammerman, P.S.; Vaklavas, C.; Mohindra, N.; Kalyan, A.; Matsangou, M.; Costa, R.; Carneiro, B.; Villaflor, V.M.; et al. Inhibition of the fibroblast growth factor receptor (FGFR) pathway: The current landscape and barriers to clinical application. Oncotarget 2016, 8, 16052–16074. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, A.; Malvi, P.; Wajapeyee, N. Heparan Sulfate and Heparan Sulfate Proteoglycans in Cancer Initiation and Progression. Front. Endocrinol. 2018, 9, 483. [Google Scholar] [CrossRef] [PubMed]
- Mossahebi-Mohammadi, M.; Quan, M.; Zhang, J.-S.; Li, X. FGF Signaling Pathway: A Key Regulator of Stem Cell Pluripotency. Front. Cell Dev. Biol. 2020, 8, 79. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-P.; Kusche-Gullberg, M. Heparan Sulfate: Biosynthesis, Structure, and Function. Int. Rev. Cell Mol. Biol. 2016, 325, 215–273. [Google Scholar] [CrossRef]
- Khan, S.; Rodriguez, E.; Patel, R.; Gor, J.; Mulloy, B.; Perkins, S.J. Withdrawal: The solution structure of heparan sulfate differs from that of heparin. IMPLICATIONS FOR FUNCTION. J. Biol. Chem. 2013, 288, 21209. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.-J.; Akhtar, J.; Desai, P.; Shukla, D. A role for heparan sulfate in viral surfing. Biochem. Biophys. Res. Commun. 2010, 391, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Khurana, A.; Beleford, D.; He, X.; Chien, J.; Shridhar, V. Role of heparan sulfatases in ovarian and breast cancer. Am. J. Cancer Res. 2013, 3, 34–45. [Google Scholar]
- Knelson, E.H.; Nee, J.C.; Blobe, G.C. Heparan sulfate signaling in cancer. Trends Biochem. Sci. 2014, 39, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Denys, A.; Allain, F. The Emerging Roles of Heparan Sulfate 3-O-Sulfotransferases in Cancer. Front. Oncol. 2019, 9, 507. [Google Scholar] [CrossRef] [Green Version]
- Goetz, R.; Mohammadi, M. Exploring mechanisms of FGF signalling through the lens of structural biology. Nat. Rev. Mol. Cell Biol. 2013, 14, 166–180. [Google Scholar] [CrossRef] [Green Version]
- Kalinina, J.; Dutta, K.; Ilghari, D.; Beenken, A.; Goetz, R.; Eliseenkova, A.V.; Cowburn, D.; Mohammadi, M. The Alternatively Spliced Acid Box Region Plays a Key Role in FGF Receptor Autoinhibition. J. Struct. 2012, 20, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosravi, F.; Ahmadvand, N.; Bellusci, S.; Sauer, H. The Multifunctional Contribution of FGF Signaling to Cardiac Development, Homeostasis, Disease and Repair. Front. Cell Dev. Biol. 2021, 9, 1217. [Google Scholar] [CrossRef]
- Donate-Correa, J.; Martín-Núñez, E.; Delgado, N.P.; de Fuentes, M.M.; Arduan, A.O.; Mora-Fernández, C.; González, J.F.N. Implications of Fibroblast growth factor/Klotho system in glucose metabolism and diabetes. Cytokine Growth Factor Rev. 2016, 28, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Degirolamo, C.; Sabbà, C.; Moschetta, A. Therapeutic potential of the endocrine fibroblast growth factors FGF19, FGF21 and FGF23. Nat. Rev. Drug Discov. 2016, 15, 51–69. [Google Scholar] [CrossRef]
- Quarles, L.D. Role of FGF23 in vitamin D and phosphate metabolism: Implications in chronic kidney disease. Exp. Cell Res. 2012, 318, 1040–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erben, R.G. Physiological Actions of Fibroblast Growth Factor-23. Front. Endocrinol. 2018, 9, 267. [Google Scholar] [CrossRef]
- Kuro-o, M. The Klotho proteins in health and disease. Nat. Rev. Nephrol. 2018, 15, 27–44. [Google Scholar] [CrossRef]
- Nakamizo, S.; Egawa, G.; Doi, H.; Natsuaki, Y.; Miyachi, Y.; Kabashima, K. Topical Treatment with Basic Fibroblast Growth Factor Promotes Wound Healing and Barrier Recovery Induced by Skin Abrasion. Ski. Pharmacol. Physiol. 2013, 26, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Robson, M.C.; Hill, D.P.; Smith, P.D.; Wang, X.; Meyer-Siegler, K.; Ko, F.; VandeBerg, J.S.; Payne, W.G.; Ochs, D.; Robson, L.E. Sequential Cytokine Therapy for Pressure Ulcers: Clinical and Mechanistic Response. Ann. Surg. 2000, 231, 600–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohura, T.; Nakajo, T.; Moriguchi, T.; Oka, H.; Tachi, M.; Ohura, N.; Nogami, R.; Murayama, S. Clinical efficacy of basic fibroblast growth factor on pressure ulcers: Case-control pairing study using a new evaluation method. Wound Repair Regen. 2011, 19, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Bing, M.; Da-Sheng, C.; Zhao-Fan, X.; Dao-Feng, B.; Wei, L.; Zhi-Fang, C.; Qiang, W.; Jia, H.; Jia-Ke, C.; Chuan-An, S.; et al. Randomized, multicenter, double-blind, and placebo-controlled trial using topical recombinant human acidic fibroblast growth factor for deep partial-thickness burns and skin graft donor site. Wound Repair Regen. 2007, 15, 795–799. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. PERSPECTIVE ARTICLE: Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Wilkinson, H.N.; Hardman, M.J. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10. [Google Scholar] [CrossRef]
- Zarei, F.; Soleimaninejad, M. Role of growth factors and biomaterials in wound healing. Artif. Cells Nanomed. Biotechnol. 2018, 46, 906–911. [Google Scholar] [CrossRef] [Green Version]
- Cundell, J. Diabetic Foot Ulcers: Assessment, Treatment, and Management. Smart Bandage Technol. 2016, 2016, 37–61. [Google Scholar] [CrossRef]
- Gonzalez, A.C.D.O.; Andrade, Z.D.A.; Costa, T.F.; Medrado, A.R.A.P. Wound Healing—A Literature Review. An. Bras. De Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Hwang, S.R.; Yoon, I.-S. Advanced Growth Factor Delivery Systems in Wound Management and Skin Regeneration. Molecules 2017, 22, 1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chester, D.; Marrow, E.A.; Daniele, M.A.; Brown, A.C. Wound Healing and the Host Response in Regenerative Engineering. Encycl. Biomed. Eng. 2019, 1–3, 1–12. [Google Scholar] [CrossRef]
- Petreaca, M.; Martinsgreen, M. The Dynamics of Cell-ECM Interactions, with Implications for Tissue Engineering. Princ. Tissue Eng. 2014, 161–187. [Google Scholar] [CrossRef]
- Chicharro-Alcántara, D.; Rubio-Zaragoza, M.; Damiá-Giménez, E.; Carrillo-Poveda, J.M.; Cuervo-Serrato, B.; Peláez-Gorrea, P.; Sopena-Juncosa, J.J. Platelet Rich Plasma: New Insights for Cutaneous Wound Healing Management. J. Funct. Biomater. 2018, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.M.; Ross, K.G.; Zayas, R.M. Regeneration in Invertebrates: Model Systems. eLS 2016, 1–9. [Google Scholar] [CrossRef]
- Noro, Y.; Yum, S.; Nishimiya-Fujisawa, C.; Busse, C.; Shimizu, H.; Mineta, K.; Zhang, X.; Holstein, T.; David, C.N.; Gojobori, T.; et al. Regionalized nervous system in Hydra and the mechanism of its development. Gene Expr. Patterns 2019, 31, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Lange, E.; Bertrand, S.; Holz, O.; Rebscher, N.; Hassel, M. Dynamic expression of a Hydra FGF at boundaries and termini. Dev. Genes Evol. 2014, 224, 235–244. [Google Scholar] [CrossRef]
- Krishnapati, L.-S.; Ghaskadbi, S. Identification and characterization of VEGF and FGF from Hydra. Int. J. Dev. Biol. 2013, 57, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Turwankar, A.; Ghaskadbi, S. VEGF and FGF signaling during head regeneration in hydra. Gene 2019, 717, 144047. [Google Scholar] [CrossRef]
- Luisetto, M.; Naseer, A.; Abdul, H.G.; Ibrahim, G.; Nili, A.B.; Yesvi, R.A.; Rasool, M.G.; Prince, G.T.; Yurievich, L.O. Regeneration Abilities of Vertebrates and Invertebrates and Relationship with Pharmacological Research: Hypothesis of Genetic Evolution Work and Microenvironment Inhibition Role. Int. J. Cancer Clin. Res. 2020, 7. [Google Scholar] [CrossRef]
- Auwal, M.A.; Kashima, M.; Nishimura, O.; Hosoda, K.; Motoishi, M.; Kamimura, A.; Okumura, A.; Agata, K.; Umesono, Y. Identification and characterization of a fibroblast growth factor gene in the planarian Dugesia japonica. Dev. Growth Differ. 2020, 62, 527–539. [Google Scholar] [CrossRef]
- Oh, G.-W.; Ko, S.-C.; Lee, D.H.; Heo, S.-J.; Jung, W.-K. Biological activities and biomedical potential of sea cucumber (Stichopus japonicus): A review. Fish. Aquat. Sci. 2017, 20, 28. [Google Scholar] [CrossRef]
- Zhang, Y.; Song, S.; Song, D.; Liang, H.; Wang, W.; Ji, A. Proliferative effects on neural stem/progenitor cells of a sulfated polysaccharide purified from the sea cucumber Stichopus japonicus. J. Biosci. Bioeng. 2010, 109, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Cancre, I.; Van Wormhoudt, A.; le Gal, Y. Heparin-binding molecules with growth factor activities in regenerating-tissues of the starfish Asterias rubens. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1999, 123, 285–292. [Google Scholar] [CrossRef]
- Czarkwiani, A.; Dylus, D.V.; Carballo, L.; Oliveri, P. FGF signalling plays similar roles in development and regeneration of the skeleton in the brittle star Amphiura filiformis. bioRxiDev 2021, 148, dev180760. [Google Scholar] [CrossRef]
- Zhao, A.; Qin, H.; Fu, X. What Determines the Regenerative Capacity in Animals? Bioscience 2016, 66, 735–746. [Google Scholar] [CrossRef]
- Poss, K.; Shena, J.; Nechiporuka, A.; Mc Mahonb, G.; Thissec, B.; Thissec, C.; Keating, M.T. Roles for Fgf Signaling during Zebrafish Fin Regeneration. Dev. Biol. 2000, 222, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Duszynski, R.J.; Topczewski, J.; LeClair, E.E. Divergent requirements for fibroblast growth factor signaling in zebrafish maxillary barbel and caudal fin regeneration. Dev. Growth Differ. 2013, 55, 282–300. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Kidd, A.R.; Thomas, J.L.; Poss, K.D.; Hyde, D.R.; Raymond, P.A.; Thummel, R. FGF signaling regulates rod photoreceptor cell maintenance and regeneration in zebrafish. Exp. Eye Res. 2011, 93, 726–734. [Google Scholar] [CrossRef] [Green Version]
- Saera-Vila, A.; Kish, P.E.; Kahana, A. Fgf regulates dedifferentiation during skeletal muscle regeneration in adult zebrafish. Cell. Signal. 2016, 28, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Kakebeen, A.D.; Wills, A.E. More Than Just a Bandage: Closing the Gap Between Injury and Appendage Regeneration. Front. Physiol. 2019, 10, 81. [Google Scholar] [CrossRef]
- Han, M.-J.; An, J.-Y.; Kim, W.-S. Expression patterns ofFgf-8 during development and limb regeneration of the axolotl. Dev. Dyn. 2001, 220, 40–48. [Google Scholar] [CrossRef]
- Christen, B.; Slack, J.M. FGF-8Is Associated with Anteroposterior Patterning and Limb Regeneration in Xenopus. Dev. Biol. 1997, 192, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Müller, A.-K.; Yang, J.; Moik, D.; Ponzio, G.; Ornitz, D.; Grose, R.; Werner, S. FGF receptors 1 and 2 are key regulators of keratinocyte migration in vitro and in wounded skin. J. Cell Sci. 2012, 125, 5690–5701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narla, S.T.; Bushnell, D.S.; Schaefer, C.M.; Nouraie, M.; Tometich, J.T.; Hand, T.W.; Bates, C.M. Loss of Fibroblast Growth Factor Receptor 2 (FGFR2) Leads to Defective Bladder Urothelial Regeneration after Cyclophosphamide Injury. Am. J. Pathol. 2021, 191, 631–651. [Google Scholar] [CrossRef]
- Guzy, R.D.; Stoilov, I.; Elton, T.J.; Mecham, R.P.; Ornitz, D.M. Fibroblast Growth Factor 2 Is Required for Epithelial Recovery, but Not for Pulmonary Fibrosis, in Response to Bleomycin. Am. J. Respir. Cell Mol. Biol. 2015, 52, 116–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, H.Y.; El-Baz, L.M.; House, S.L.; Cilvik, S.; Dorry, S.J.; Shoukry, N.M.; Salem, M.L.; Hafez, H.S.; Dulin, N.O.; Ornitz, D.M.; et al. Fibroblast growth factor 2 decreases bleomycin-induced pulmonary fibrosis and inhibits fibroblast collagen production and myofibroblast differentiation. J. Pathol. 2018, 246, 54–66. [Google Scholar] [CrossRef]
- Morscheid, Y.P.; Venkatesan, J.K.; Schmitt, G.; Orth, P.; Zurakowski, D.; Speicher-Mentges, S.; Menger, M.D.; Laschke, M.W.; Cucchiarini, M.; Madry, H. rAAV-Mediated Human FGF-2 Gene Therapy Enhances Osteochondral Repair in a Clinically Relevant Large Animal Model Over Time In Vivo. Am. J. Sports Med. 2021, 49, 958–969. [Google Scholar] [CrossRef]
- Yun, Y.-R.; Won, J.E.; Jeon, E.; Lee, S.; Kang, W.; Jo, H.; Jang, J.H.; Shin, U.S.; Kim, H.W. Fibroblast Growth Factors: Biology, Function, and Application for Tissue Regeneration. J. Tissue Eng. 2010, 2010, 218142. [Google Scholar] [CrossRef]
- Akita, S.; Akino, K.; Imaizumi, T.; Hirano, A. Basic fibroblast growth factor accelerates and improves second-degree burn wound healing. Wound Repair Regen. 2008, 16, 635–641. [Google Scholar] [CrossRef]
- Callaghan, M.J.; Chang, E.I.; Seiser, N.; Aarabi, S.; Ghali, S.; Kinnucan, E.R.; Simon, B.J.; Gurtner, G.C. Pulsed Electromagnetic Fields Accelerate Normal and Diabetic Wound Healing by Increasing Endogenous FGF-2 Release. Plast. Reconstr. Surg. 2008, 121, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Blaber, S.I.; Díaz, J.; Blaber, M. Accelerated healing in NONcNZO10/LtJ type 2 diabetic mice by FGF-1. Wound Repair Regen. 2015, 23, 538–549. [Google Scholar] [CrossRef]
- Schmid, G.J.; Kobayashi, C.; Sandell, L.J.; Ornitz, D.M. Fibroblast growth factor expression during skeletal fracture healing in mice. Dev. Dyn. 2009, 238, 766–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, Q.; Jin, Z.; Li, X.; Liu, C.; Wang, X. FGF Family: From Drug Development to Clinical Application. Int. J. Mol. Sci. 2018, 19, 1875. [Google Scholar] [CrossRef] [Green Version]
- Itoh, N. Hormone-like (endocrine) Fgfs: Their evolutionary history and roles in development, metabolism, and disease. Cell Tissue Res. 2010, 342, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Yim, H.; Shin, J.; Lee, C.; Lee, J.-H.; Jeong, J. FGF11 induced by hypoxia interacts with HIF-1α and enhances its stability. FEBS Lett. 2017, 591, 348–357. [Google Scholar] [CrossRef]
- Yamauchi, M.; Hirohashi, Y.; Torigoe, T.; Matsumoto, Y.; Yamashita, K.; Kayama, M.; Sato, N.; Yotsuyanagi, T. Wound healing delays in α-Klotho -deficient mice that have skin appearance similar to that in aged humans—Study of delayed wound healing mechanism. Biochem. Biophys. Res. Commun. 2016, 473, 845–852. [Google Scholar] [CrossRef]
- Zhu, S.; Ying, Y.; Ye, L.; Ying, W.; Ye, J.; Wu, Q.; Chen, M.; Zhu, H.; Li, X.; Dou, H.; et al. Systemic Administration of Fibroblast Growth Factor 21 Improves the Recovery of Spinal Cord Injury (SCI) in Rats and Attenuates SCI-Induced Autophagy. Front. Pharmacol. 2021, 11, 628369. [Google Scholar] [CrossRef]
- Kong, B.; Huang, J.; Zhu, Y.; Li, G.; Williams, J.; Shen, S.; Aleksunes, L.M.; Richardson, J.R.; Apte, U.; Rudnick, D.A.; et al. Fibroblast growth factor 15 deficiency impairs liver regeneration in mice. Am. J. Physiol. Liver Physiol. 2014, 306, G893–G902. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Wang, C. Dedifferentiation: Inspiration for devising engineering strategies for regenerative medicine. NPJ Regen. Med. 2020, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kleiderman, S.; Gutbier, S.; Tufekci, K.U.; Ortega, F.; Sá, J.V.; Teixeira, A.; Brito, C.; Glaab, E.; Berninger, B.; Alves, P.; et al. Conversion of Nonproliferating Astrocytes into Neurogenic Neural Stem Cells: Control by FGF2 and Interferon-γ. Stem Cells 2016, 34, 2861–2874. [Google Scholar] [CrossRef] [Green Version]
- Koike, Y.; Yozaki, M.; Utani, A.; Murota, H. Fibroblast growth factor 2 accelerates the epithelial–mesenchymal transition in keratinocytes during wound healing process. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Qin, L.; Lingfeng, Q.; Tellides, G.; Simons, M. Fibroblast growth factor (FGF) signaling regulates transforming growth factor beta (TGFβ)-dependent smooth muscle cell phenotype modulation. Sci. Rep. 2016, 6, 33407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kottakis, F.; Polytarchou, C.; Foltopoulou, P.; Sanidas, I.; Kampranis, S.; Tsichlis, P.N. FGF-2 Regulates Cell Proliferation, Migration, and Angiogenesis through an NDY1/KDM2B-miR-101-EZH2 Pathway. Mol. Cell 2011, 43, 285–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlikowski, B.; Vogler, T.O.; Gadek, K.; Olwin, B.B. Regulation of skeletal muscle stem cells by fibroblast growth factors. Dev. Dyn. 2017, 246, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Galimov, A.; Merry, T.L.; Luca, E.; Rushing, E.J.; Mizbani, A.; Turcekova, K.; Hartung, A.; Croce, C.M.; Ristow, M.; Krützfeldt, J. MicroRNA-29a in Adult Muscle Stem Cells Controls Skeletal Muscle Regeneration During Injury and Exercise Downstream of Fibroblast Growth Factor-2. Stem Cells 2016, 34, 768–780. [Google Scholar] [CrossRef] [Green Version]
- Whittemore, K.; Vera, E.; Martínez-Nevado, E.; Sanpera, C.; Blasco, M.A. Telomere shortening rate predicts species life span. Proc. Natl. Acad. Sci. USA 2019, 116, 15122–15127. [Google Scholar] [CrossRef] [Green Version]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Calado, R.T.; Dumitriu, B. Telomere Dynamics in Mice and Humans. Semin. Hematol. 2013, 50, 165–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Beck, S.; Sohn, Y.-W.; Kim, J.-K.; Kim, S.-H.; Yin, J.; Pian, X.; Kim, S.-C.; Choi, Y.-J.; Kim, H. Human telomerase catalytic subunit (hTERT) suppresses p53-mediated anti-apoptotic response via induction of basic fibroblast growth factor. Exp. Mol. Med. 2010, 42, 574–582. [Google Scholar] [CrossRef] [PubMed]
- Coutu, D.; Galipeau, J. Roles of FGF signaling in stem cell self-renewal, senescence and aging. Aging 2011, 3, 920–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Tong, H.J.; Li, M.; Tan, K.S.; Cao, T. Telomere length is regulated by FGF-2 in human embryonic stem cells and affects the life span of its differentiated progenies. Biogerontology 2016, 18, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Erusalimsky, J.D.; Skene, C. Mechanisms of endothelial senescence. Exp. Physiol. 2009, 94, 299–304. [Google Scholar] [CrossRef]
- Fernández, C.S.; Lorda-Diez, C.I.; García-Porrero, J.A.; Montero, J.A.; Hurlé, J.M. UHRF genes regulate programmed interdigital tissue regression and chondrogenesis in the embryonic limb. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Page, R.L.; Ambady, S.; Holmes, W.F.; Vilner, L.; Kole, D.; Kashpur, O.; Huntress, V.; Vojtic, I.; Whitton, H.; Dominko, T. Induction of Stem Cell Gene Expression in Adult Human Fibroblasts without Transgenes. Cloning Stem Cells 2009, 11, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Coutu, D.; Francois, M.; Galipeau, J. Inhibition of cellular senescence by developmentally regulated FGF receptors in mesenchymal stem cells. Blood 2011, 117, 6801–6812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorda-Diez, C.I.; Garcia-Riart, B.; Montero, J.A.; Leon, J.M.R.; Garcia-Porrero, J.; Hurle, J.M. Apoptosis during embryonic tissue remodeling is accompanied by cell senescence. Aging 2015, 7, 974–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.H.; Lee, H.C.; Yu, D.-M.; Kim, B.C.; Park, S.M.; Lee, Y.-S.; Park, H.J.; Ko, Y.-G.; Lee, J.-S. Heparan sulfation is essential for the prevention of cellular senescence. Cell Death Differ. 2016, 23, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Chow, R.; Roux, G.; Roghani, M.; Palmer, M.; Rifkin, D.; Moscatelli, D.; Lang, R. FGF suppresses apoptosis and induces differentiation of fibre cells in the mouse lens. Development 1995, 121, 4383–4393. [Google Scholar] [CrossRef]
- Vaahtokari, A.; Aberg, T.; Thesleff, I. Apoptosis in the developing tooth: Association with an embryonic signaling center and suppression by EGF and FGF-4. Development 1996, 122, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Pardo, O.; Wellbrock, C.; Khanzada, U.K.; Aubert, M.; Arozarena, I.; Davidson, S.; Bowen, F.; Parker, P.; Filonenko, V.; Gout, I.T.; et al. FGF-2 protects small cell lung cancer cells from apoptosis through a complex involving PKCɛ, B-Raf and S6K2. EMBO J. 2006, 25, 3078–3088. [Google Scholar] [CrossRef]
- Giacomini, A.; Taranto, S.; Rezzola, S.; Matarazzo, S.; Grillo, E.; Bugatti, M.; Scotuzzi, A.; Guerra, J.; Di Trani, M.; Presta, M.; et al. Inhibition of the FGF/FGFR System Induces Apoptosis in Lung Cancer Cells via c-Myc Downregulation and Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 9376. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Enkhjargal, B.; Travis, Z.D.; Ocak, U.; Tang, J.; Suzuki, H.; Zhang, J.H. FGF-2 Attenuates Neuronal Apoptosis via FGFR3/PI3k/Akt Signaling Pathway After Subarachnoid Hemorrhage. Mol. Neurobiol. 2019, 56, 8203–8219. [Google Scholar] [CrossRef]
- Motamedi, F.J.; Badro, D.; Clarkson, M.; Lecca, M.R.; Bradford, S.; Buske, F.A.; Saar, K.; Hübner, N.; Brändli, A.W.; Schedl, A. WT1 controls antagonistic FGF and BMP-pSMAD pathways in early renal progenitors. Nat. Commun. 2014, 5, 4444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, M.; Xin, S. FGF signaling contributes to atherosclerosis by enhancing the inflammatory response in vascular smooth muscle cells. Mol. Med. Rep. 2019, 20, 162–170. [Google Scholar] [CrossRef]
- Lou, D.; Han, J.; Zhou, L.; Ma, H.; Xv, J.; Shou, J.; Xu, Z.; Jiang, L.; Qian, Y. Fibroblast growth factor receptor 1 antagonism attenuates lipopolysaccharide-induced activation of hepatic stellate cells via suppressing inflammation. Exp. Ther. Med. 2018, 16, 2909–2916. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ke, Y.; Liu, S.; Pan, S.; Liu, Z.; Zhang, H.; Fan, Z.; Zhou, C.; Liu, J.; Wang, F. Ectopic fibroblast growth factor receptor 1 promotes inflammation by promoting nuclear factor-κB signaling in prostate cancer cells. J. Biol. Chem. 2018, 293, 14839–14849. [Google Scholar] [CrossRef] [Green Version]
- Pu, J.; Wang, R.; Zhang, G.; Wang, J. FGF-7 facilitates the process of psoriasis by inducing TNF-α expression in HaCaT cells. Acta Biochim. Biophys. Sin. 2019, 51, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Im, H.-J.; Li, X.; Muddasani, P.; Kim, G.-H.; Davis, F.; Rangan, J.; Forsyth, C.B.; Ellman, M.; Thonar, E.J. Basic fibroblast growth factor accelerates matrix degradation via a neuro-endocrine pathway in human adult articular chondrocytes. J. Cell. Physiol. 2008, 215, 452–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirov, A.; Duarte, M.; Guay, J.; Karolak, M.; Yan, C.; Oxburgh, L.; Prudovsky, I. Transgenic Expression of Nonclassically Secreted FGF Suppresses Kidney Repair. PLoS ONE 2012, 7, e36485. [Google Scholar] [CrossRef]
- Meij, J.T.A.; Sheikh, F.; Jimenez, S.K.; Nickerson, P.W.; Kardami, E.; Cattini, P.A. Exacerbation of myocardial injury in transgenic mice overexpressing FGF-2 is T cell dependent. Am. J. Physiol. Circ. Physiol. 2002, 282, H547–H555. [Google Scholar] [CrossRef] [Green Version]
- Bovolenta, R.; Zucchini, S.; Paradiso, B.; Rodi, D.; Merigo, F.; Mora, G.N.; Osculati, F.; Berto, E.; Marconi, P.; Marzola, A.; et al. Hippocampal FGF-2 and BDNF overexpression attenuates epileptogenesis-associated neuroinflammation and reduces spontaneous recurrent seizures. J. Neuroinflammation 2010, 7, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, H.-J.; Zhao, C.-F.; Chen, Z.-W.; Lin, W.; Jiang, Y.-C. Fibroblast Growth Factor (FGF) Signaling Protects Against Acute Pancreatitis-Induced Damage by Modulating Inflammatory Responses. Med Sci. Monit. 2020, 26, e920684–1. [Google Scholar] [CrossRef]
- Fan, L.; Ding, L.; Lan, J.; Niu, J.; He, Y.; Song, L. Fibroblast Growth Factor-1 Improves Insulin Resistance via Repression of JNK-Mediated Inflammation. Front. Pharmacol. 2019, 10, 1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czaya, B.; Faul, C. FGF23 and inflammation—A vicious coalition in CKD. Kidney Int. 2019, 96, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Egli-Spichtig, D.; Imenez Silva, P.H.; Glaudemans, B.; Gehring, N.; Bettoni, C.; Zhang, M.Y.H.; Pastor-Arroyo, E.M.; Schönenberger, D.; Rajski, M.; Hoogewijs, D.; et al. Tumor necrosis factor stimulates fibroblast growth factor 23 levels in chronic kidney disease and non-renal inflammation. Kidney Int. 2019, 96, 890–905. [Google Scholar] [CrossRef]
- Taylor, A.; Yanucil, C.; Musgrove, J.; Shi, M.; Ide, S.; Souma, T.; Faul, C.; Wolf, M.; Grabner, A. FGFR4 does not contribute to progression of chronic kidney disease. Sci. Rep. 2019, 9, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Grabner, A.; Yanucil, C.; Schramm, K.; Czaya, B.; Krick, S.; Czaja, M.J.; Bartz, R.; Abraham, R.; Di Marco, G.S.; et al. Fibroblast growth factor 23 directly targets hepatocytes to promote inflammation in chronic kidney disease. Kidney Int. 2016, 90, 985–996. [Google Scholar] [CrossRef] [Green Version]
- Bodnar, R.J. Chemokine Regulation of Angiogenesis During Wound Healing. Adv. Wound Care 2015, 4, 641–650. [Google Scholar] [CrossRef]
- Vailhé, B.; Vittet, D.; Feige, J.-J. In Vitro Models of Vasculogenesis and Angiogenesis. Lab. Investig. 2001, 81, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Javerzat, S.; Auguste, P.; Bikfalvi, A. The role of fibroblast growth factors in vascular development. Trends Mol. Med. 2002, 8, 483–489. [Google Scholar] [CrossRef]
- Chiodelli, P.; Bugatti, A.; Urbinati, C.E.; Rusnati, M. Heparin/Heparan Sulfate Proteoglycans Glycomic Interactome in Angiogenesis: Biological Implications and Therapeutical Use. Molecules 2015, 20, 6342–6388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucuzian, A.A.; Gassman, A.A.; East, A.T.; Greisler, H.P. Molecular Mediators of Angiogenesis. J. Burn. Care Res. 2010, 31, 158–175. [Google Scholar] [CrossRef] [PubMed]
- LaVine, K.J.; White, A.; Park, C.; Smith, C.S.; Choi, K.; Long, F.; Hui, C.-C.; Ornitz, D.M. Fibroblast growth factor signals regulate a wave of Hedgehog activation that is essential for coronary vascular development. Genes Dev. 2006, 20, 1651–1666. [Google Scholar] [CrossRef] [Green Version]
- Murakami, M.; Simons, M. Fibroblast growth factor regulation of neovascularization. Curr. Opin. Hematol. 2008, 15, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Bao, L.; Zhao, M.; Cao, J.; Zheng, H. Progress in Research on the Role of FGF in the Formation and Treatment of Corneal Neovascularization. Front. Pharmacol. 2020, 11, 111. [Google Scholar] [CrossRef]
- Hori, Y.; Ito, K.; Hamamichi, S.; Ozawa, Y.; Matsui, J.; Umeda, I.O.; Fujii, H. Functional Characterization of VEGF- and FGF-induced Tumor Blood Vessel Models in Human Cancer Xenografts. Anticancer. Res. 2017, 37, 6629–6638. [Google Scholar] [CrossRef] [Green Version]
- Ramos, O.; Malcata, F. Food-Grade Enzymes. Compr. Biotechnol. 2017, 2017, 587–603. [Google Scholar] [CrossRef]
- Kondreddy, S.; Palli, S. Proteases and tissue repair: Peri operative role of chymotrypsin: Trypsin in surgical patients. Int. Surg. J. 2018, 6, 283–286. [Google Scholar] [CrossRef]
- Wang, C.; Li, Y.; Li, H.; Zhang, Y.; Ying, Z.; Wang, X.; Zhang, T.; Zhang, W.; Fan, Z.; Li, X.; et al. Disruption of FGF Signaling Ameliorates Inflammatory Response in Hepatic Stellate Cells. Front. Cell Dev. Biol. 2020, 8, 601. [Google Scholar] [CrossRef] [PubMed]
- Nummenmaa, E.; Hämäläinen, M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Effects of FGF-2 and FGF receptor antagonists on MMP enzymes, aggrecan, and type II collagen in primary human OA chondrocytes. Scand. J. Rheumatol. 2015, 44, 321–330. [Google Scholar] [CrossRef]
- Lungu, G.; Covaleda, L.; Mendes, O.; Martini-Stoica, H.; Stoica, G. FGF-1-induced matrix metalloproteinase-9 expression in breast cancer cells is mediated by increased activities of NF-κB and activating protein-1. Mol. Carcinog. 2008, 47, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-F.; Crépin, M.; Barritault, D.; Ledoux, D. FGF-2 and TPA induce matrix metalloproteinase-9 secretion in MCF-7 cells through PKC activation of the Ras/ERK pathway. Biochem. Biophys. Res. Commun. 2002, 293, 1174–1182. [Google Scholar] [CrossRef]
- Abdelhakim, M.; Lin, X.; Ogawa, R. The Japanese Experience with Basic Fibroblast Growth Factor in Cutaneous Wound Management and Scar Prevention: A Systematic Review of Clinical and Biological Aspects. Dermatol. Ther. 2020, 10, 569–587. [Google Scholar] [CrossRef] [PubMed]
- Jimi, S.; Jaguparov, A.; Nurkesh, A.; Sultankulov, B.; Saparov, A. Sequential Delivery of Cryogel Released Growth Factors and Cytokines Accelerates Wound Healing and Improves Tissue Regeneration. Front. Bioeng. Biotechnol. 2020, 8, 345. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Y.; Deng, J.; Li, W.; Nie, X. Fibroblast Growth Factor in Diabetic Foot Ulcer: Progress and Therapeutic Prospects. Front. Endocrinol. 2021, 12, 1348. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Members |
|---|---|
| FGF1 subfamily | FGF-1, FGF-2 |
| FGF4 subfamily | FGF-4, FGF-5, FGF-6 |
| FGF7 subfamily | FGF-3, FGF-7, FGF-10, FGF-22 |
| FGF8 subfamily | FGF-8, FGF-17, FGF-18 |
| FGF9 subfamily | FGF-9, FGF-16, FGF-20 |
| FGF11 subfamily | FGF-11, FGF-12, FGF-13, FGF-14 |
| FGF19 subfamily | FGF-19, FGF-21, FGF-23 |
| Growth Factor | Interacting with the Receptor | References |
|---|---|---|
| FGF1 | FGFR1 IIIb, FGFR1 IIIc, FGFR2 IIIb, FGFR2 IIIc, FGFR3 IIIb, FGFR3 IIIc, FGFR4 | [37,38,39,40] |
| FGF2 | FGFR1 IIIb, FGFR1 IIIc, FGFR2 IIIc, FGFR3 IIIc, FGFR4 | [40,41,42,43] |
| FGF3 | FGFR3 | [44] |
| FGF4 | FGFR2 IIIb, FGFR1 IIIc, FGFR2 IIIc, FGFR3 IIIc, FGFR4 | [45,46] |
| FGF5 | FGFR3 IIIc | [35] |
| FGF6 | FGFR1 IIIc, FGFR2 IIIc, FGFR3 IIIc, FGFR4 | [47] |
| FGF7 | FGFR2 IIIb | [47] |
| FGF8a | FGFR1 | [48] |
| FGF8b | FGFR2 IIIc, FGFR3 IIIc, FGFR4 | [2] |
| FGF8c | FGFR3 IIIc, FGFR4 | [2] |
| FGF8f | FGFR2 IIIc, FGFR3 IIIb, FGFR3 IIIc, FGFR4 | [2] |
| FGF9 | FGFR1 IIIb, FGFR2 IIIb, FGFR3 | [38] |
| FGF10 | FGFR2 | [49] |
| FGF15 | FGFR4 | [50] |
| FGF16 | FGFR1 IIIc, FGFR2 IIIc, FGFR3 IIIb, FGF3 IIIc | [47] |
| FGF17b | FGFR2IIIc, FGFR3IIIc, FGFR4 | [2] |
| FGF18 | FGFFR3 | [51] |
| FGF19 | FGFR4 | [52] |
| FGF20 | FGFR1 | [53] |
| FGF21 | FGFR1, FGFR2 | [39] |
| FGF22 | FGFR1 IIIb, FGFR2 IIIb | [47,54] |
| FGF23 | FGFR1 IIIc, FGFR2 IIIc | [47,54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, M.; Khan, A.W.; Kim, M.S.; Choi, S. The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration. Cells 2021, 10, 3242. https://doi.org/10.3390/cells10113242
Farooq M, Khan AW, Kim MS, Choi S. The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration. Cells. 2021; 10(11):3242. https://doi.org/10.3390/cells10113242
Chicago/Turabian StyleFarooq, Mariya, Abdul Waheed Khan, Moon Suk Kim, and Sangdun Choi. 2021. "The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration" Cells 10, no. 11: 3242. https://doi.org/10.3390/cells10113242
APA StyleFarooq, M., Khan, A. W., Kim, M. S., & Choi, S. (2021). The Role of Fibroblast Growth Factor (FGF) Signaling in Tissue Repair and Regeneration. Cells, 10(11), 3242. https://doi.org/10.3390/cells10113242