
Butyrate and Class I Histone Deacetylase Inhibitors Promote Differentiation of Neonatal Porcine Islet Cells into Beta Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Culture of NPICCs
2.2. Glucose Stimulated Insulin Release Assay (GSIS)
2.3. Insulin and DNA Content
2.4. Flow Cytometry Analysis
2.5. Proliferation Assay
2.6. Studies on GPCR and HDAC Mediated Signaling
2.7. Immunofluorescence
2.8. HDAC Activity Assays
2.9. RNA Extraction, Reverse Transcription and Quantitative Real-Time PCR
2.10. Statistics
3. Results
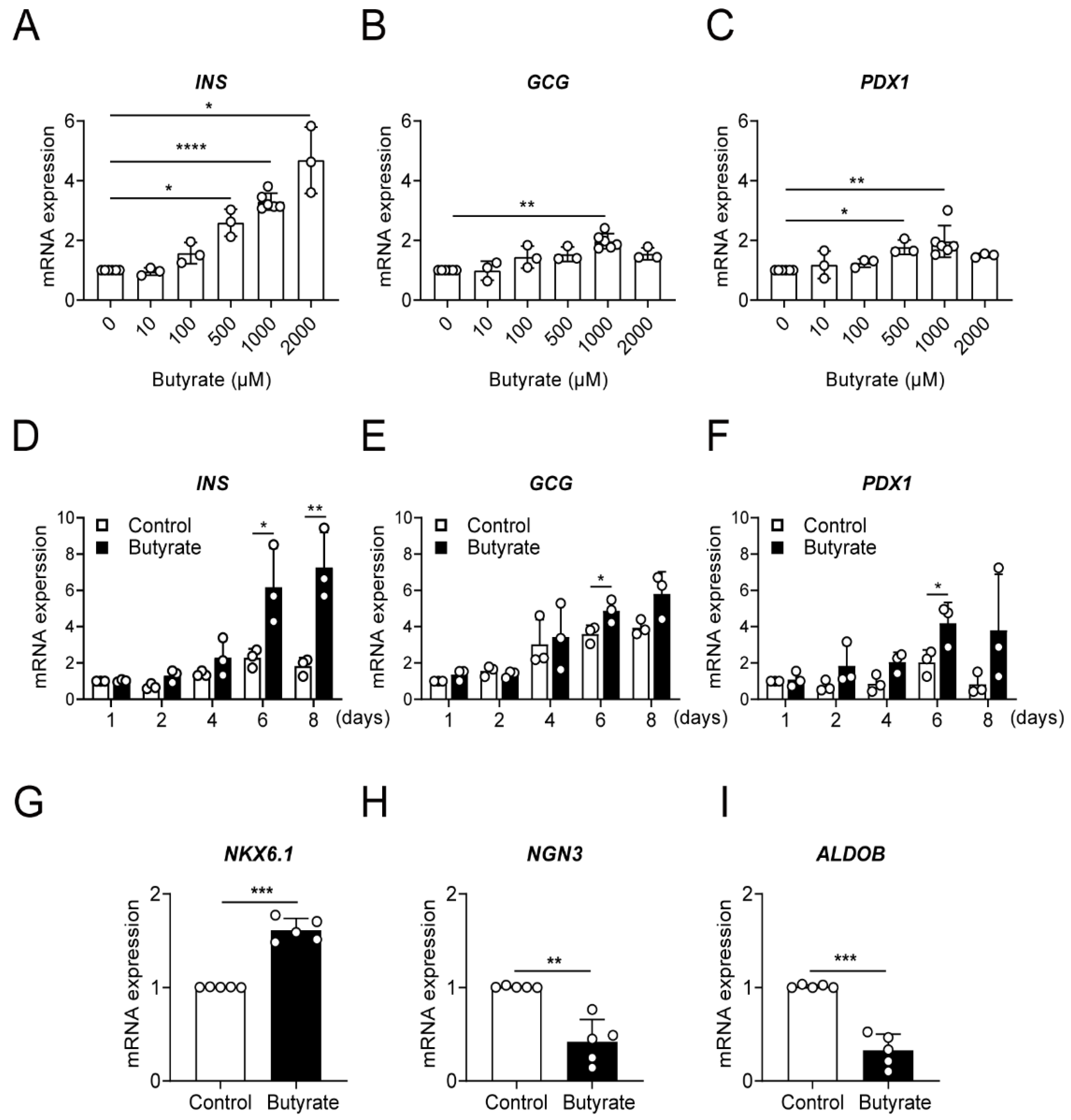
3.1. Butyrate Strongly Increases Insulin Gene Expression in a Dose- and Time-Dependent Manner
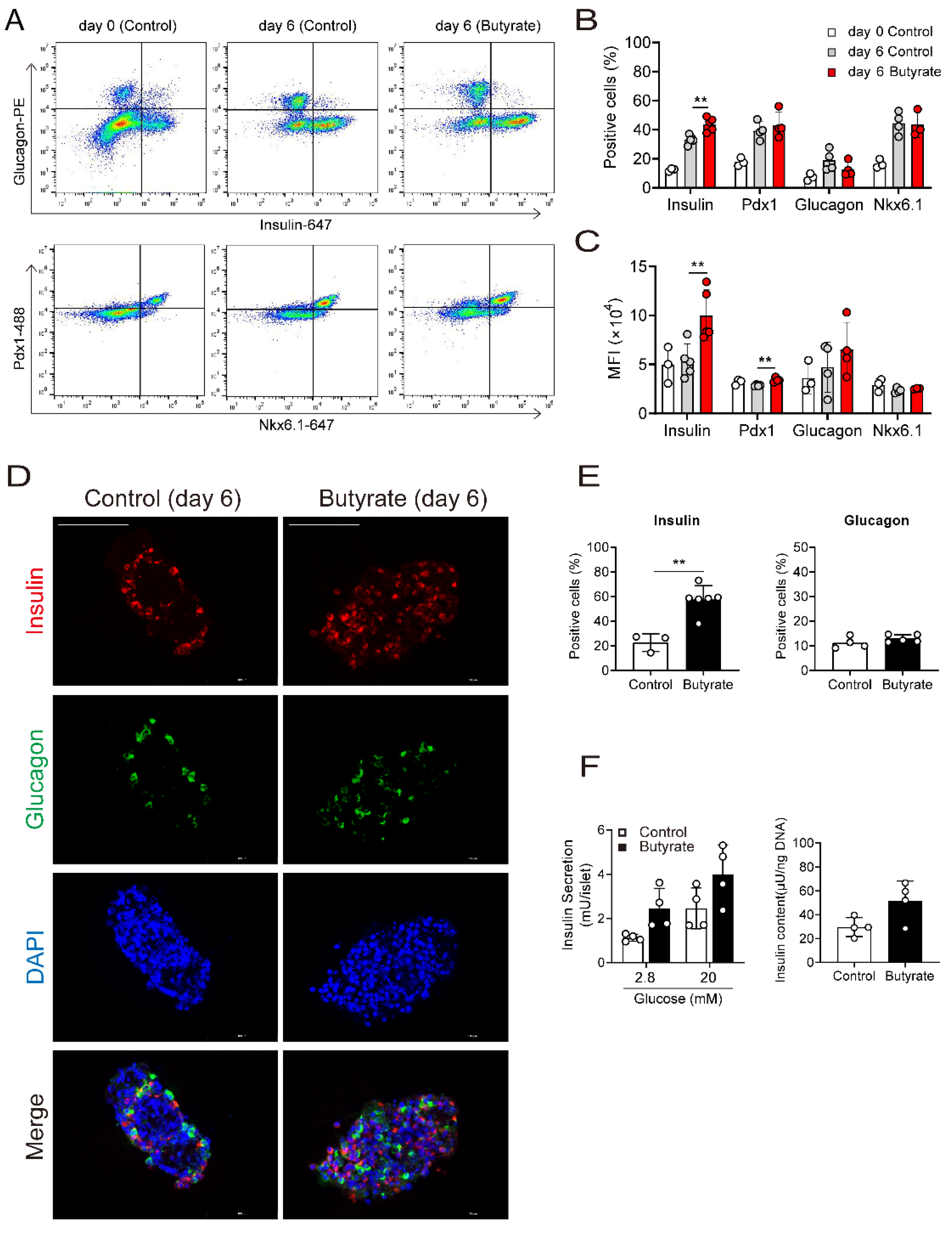
3.2. Butyrate Enhances Protein Expression of Beta Cell Maturation Markers
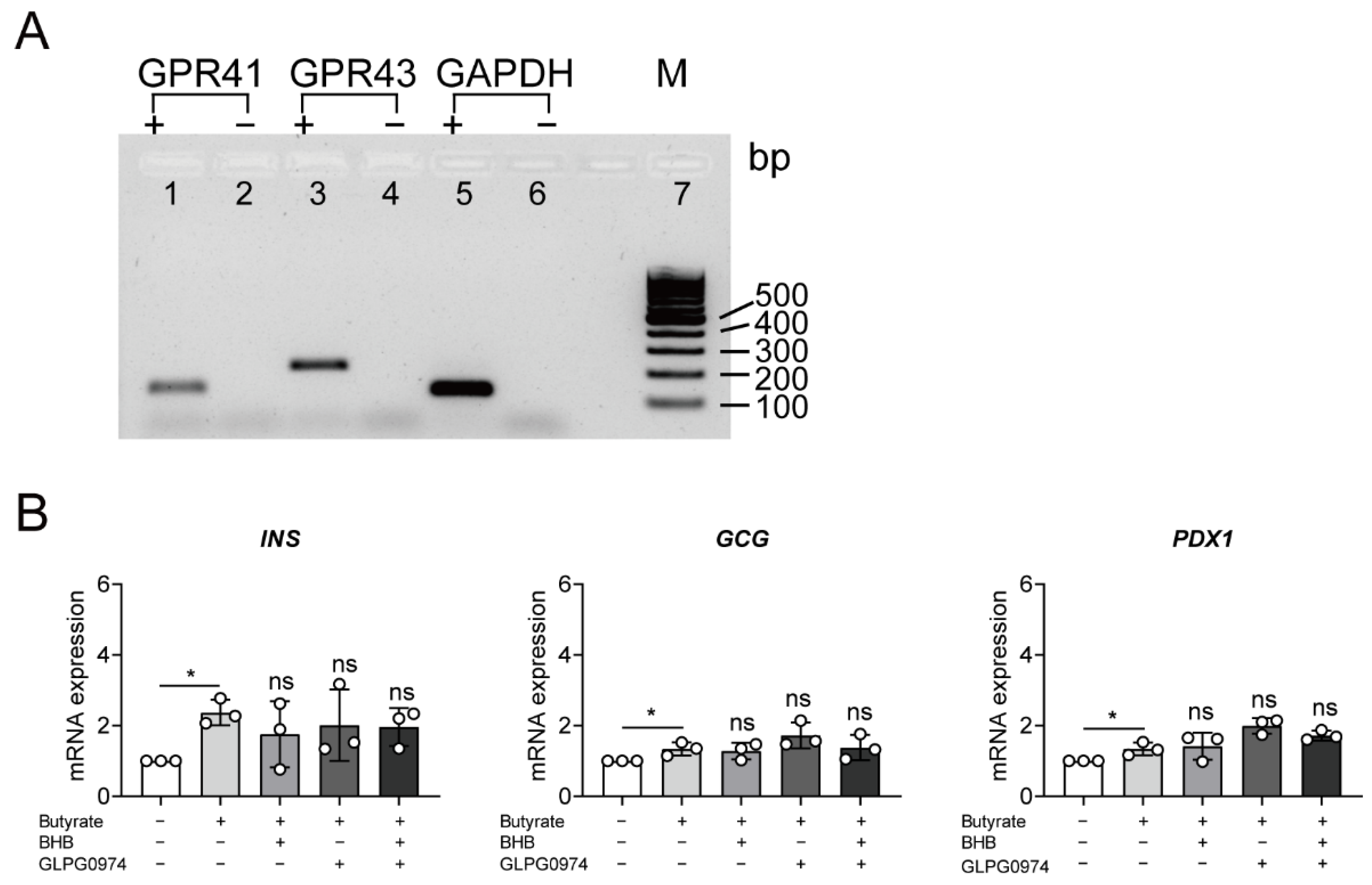
3.3. Butyrate Up-Regulated Insulin Expression Is Independent of Binding to G-Protein Coupled Receptors
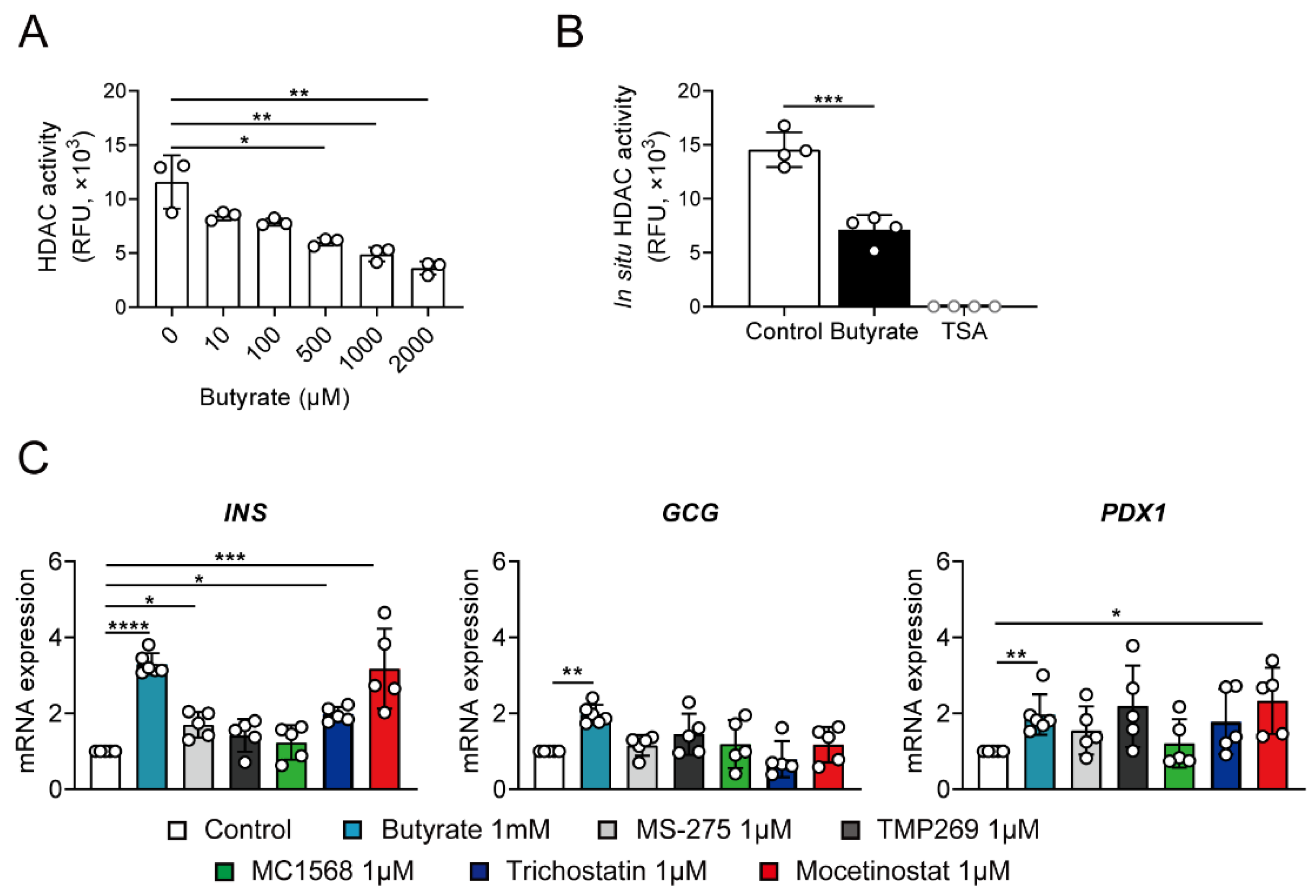
3.4. Butyrate Stimulates Insulin Transcription via Inhibition of HDAC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rickels, M.R.; Robertson, R.P. Pancreatic Islet Transplantation in Humans: Recent Progress and Future Directions. Endocr. Rev. 2019, 40, 631–668. [Google Scholar] [CrossRef] [Green Version]
- Reichart, B.; Niemann, H.; Chavakis, T.; Denner, J.; Jaeckel, E.; Ludwig, B.; Marckmann, G.; Schnieke, A.; Schwinzer, R.; Seissler, J.; et al. Xenotransplantation of porcine islet cells as a potential option for the treatment of type 1 diabetes in the future. Horm. Metab. Res. 2015, 47, 31–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemter, E.; Denner, J.; Wolf, E. Will Genetic Engineering Carry Xenotransplantation of Pig Islets to the Clinic? Curr. Diab. Rep. 2018, 18, 103. [Google Scholar] [CrossRef] [PubMed]
- Tritschler, H.; Fischer, K.; Seissler, J.; Fiedler, J.; Halbgebauer, R.; Huber-Lang, M.; Schnieke, A.; Brenner, R.E. New Insights into Xenotransplantation for Cartilage Repair: Porcine Multi-Genetically Modified Chondrocytes as a Promising Cell Source. Cells 2021, 10, 2152. [Google Scholar] [CrossRef]
- Ahrens, H.E.; Petersen, B.; Ramackers, W.; Petkov, S.; Herrmann, D.; Hauschild-Quintern, J.; Lucas-Hahn, A.; Hassel, P.; Ziegler, M.; Baars, W.; et al. Kidneys From alpha1,3-Galactosyltransferase Knockout/Human Heme Oxygenase-1/Human A20 Transgenic Pigs Are Protected From Rejection During Ex Vivo Perfusion With Human Blood. Transplant. Direct 2015, 1, e23. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.; Lyon, J.G.; Korbutt, G.S. Optimization and Scale-up Isolation and Culture of Neonatal Porcine Islets: Potential for Clinical Application. Cell Transpl. 2016, 25, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderschelden, R.; Sathialingam, M.; Alexander, M.; Lakey, J.R.T. Cost and Scalability Analysis of Porcine Islet Isolation for Islet Transplantation: Comparison of Juvenile, Neonatal and Adult Pigs. Cell Transpl. 2019, 28, 967–972. [Google Scholar] [CrossRef] [Green Version]
- Korbutt, G.S.; Elliott, J.F.; Ao, Z.; Smith, D.K.; Warnock, G.L.; Rajotte, R.V. Large scale isolation, growth, and function of porcine neonatal islet cells. J. Clin. Investig. 1996, 97, 2119–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf-van Buerck, L.W.; Schuster, M.; Oduncu, F.S.; Baehr, A.; Mayr, T.; Guethoff, S.; Abicht, J.; Reichart, B.; Klymiuk, N.; Wolf, E.; et al. LEA29Y expression in transgenic neonatal porcine islet-like cluster promotes long-lasting xenograft survival in humanized mice without immunosuppressive therapy. Sci. Rep. 2017, 7, 3572. [Google Scholar] [CrossRef]
- Trivedi, N.; Hollister-Lock, J.; Lopez-Avalos, M.D.; O’Neil, J.J.; Keegan, M.; Bonner-Weir, S.; Weir, G.C. Increase in beta-cell mass in transplanted porcine neonatal pancreatic cell clusters is due to proliferation of beta-cells and differentiation of duct cells. Endocrinology 2001, 142, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Hassouna, T.; Seeberger, K.L.; Salama, B.; Korbutt, G.S. Functional Maturation and In Vitro Differentiation of Neonatal Porcine Islet Grafts. Transplantation 2018, 102, e413–e423. [Google Scholar] [CrossRef] [PubMed]
- Hardikar, A.A.; Wang, X.Y.; Williams, L.J.; Kwok, J.; Wong, R.; Yao, M.; Tuch, B.E. Functional maturation of fetal porcine beta-cells by glucagon-like peptide 1 and cholecystokinin. Endocrinology 2002, 143, 3505–3514. [Google Scholar] [CrossRef] [Green Version]
- Lau, H.; Corrales, N.; Rodriguez, S.; Luong, C.; Zaldivar, F.; Alexander, M.; Lakey, J.R.T. An islet maturation media to improve the development of young porcine islets during in vitro culture. Islets 2020, 12, 41–58. [Google Scholar] [CrossRef]
- Lopez-Avalos, M.D.; Tatarkiewicz, K.; Sharma, A.; Bonner-Weir, S.; Weir, G.C. Enhanced maturation of porcine neonatal pancreatic cell clusters with growth factors fails to improve transplantation outcome. Transplantation 2001, 71, 1154–1162. [Google Scholar] [CrossRef]
- Jimenez-Vera, E.; Davies, S.; Phillips, P.; O’Connell, P.J.; Hawthorne, W.J. Long-term cultured neonatal islet cell clusters demonstrate better outcomes for reversal of diabetes: In vivo and molecular profiles. Xenotransplantation 2015, 22, 114–123. [Google Scholar] [CrossRef]
- Pingitore, A.; Gonzalez-Abuin, N.; Ruz-Maldonado, I.; Huang, G.C.; Frost, G.; Persaud, S.J. Short chain fatty acids stimulate insulin secretion and reduce apoptosis in mouse and human islets in vitro: Role of free fatty acid receptor 2. Diabetes Obes. Metab. 2019, 21, 330–339. [Google Scholar] [CrossRef] [Green Version]
- Pingitore, A.; Chambers, E.S.; Hill, T.; Maldonado, I.R.; Liu, B.; Bewick, G.; Morrison, D.J.; Preston, T.; Wallis, G.A.; Tedford, C.; et al. The diet-derived short chain fatty acid propionate improves beta-cell function in humans and stimulates insulin secretion from human islets in vitro. Diabetes Obes. Metab. 2017, 19, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Ximenes, H.M.; Hirata, A.E.; Rocha, M.S.; Curi, R.; Carpinelli, A.R. Propionate inhibits glucose-induced insulin secretion in isolated rat pancreatic islets. Cell Biochem. Funct. 2007, 25, 173–178. [Google Scholar] [CrossRef]
- Tang, C.; Ahmed, K.; Gille, A.; Lu, S.; Gröne, H.J.; Tunaru, S.; Offermanns, S. Loss of FFA2 and FFA3 increases insulin secretion and improves glucose tolerance in type 2 diabetes. Nat. Med. 2015, 21, 173–177. [Google Scholar] [CrossRef]
- Priyadarshini, M.; Layden, B.T. FFAR3 modulates insulin secretion and global gene expression in mouse islets. Islets 2015, 7, e1045182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ørgaard, A.; Jepsen, S.L.; Holst, J.J. Short-chain fatty acids and regulation of pancreatic endocrine secretion in mice. Islets 2019, 11, 103–111. [Google Scholar] [CrossRef]
- Philippe, J.; Drucker, D.J.; Chick, W.L.; Habener, J.F. Transcriptional regulation of genes encoding insulin, glucagon, and angiotensinogen by sodium butyrate in a rat islet cell line. Mol. Cell. Biol. 1987, 7, 560–563. [Google Scholar] [PubMed] [Green Version]
- Powers, A.C.; Philippe, J.; Hermann, H.; Habener, J.F. Sodium butyrate increases glucagon and insulin gene expression by recruiting immunocytochemically negative cells to produce hormone. Diabetes 1988, 37, 1405–1410. [Google Scholar] [CrossRef]
- Hu, S.; Kuwabara, R.; de Haan, B.J.; Smink, A.M.; de Vos, P. Acetate and Butyrate Improve β-cell Metabolism and Mitochondrial Respiration under Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otonkoski, T.; Ustinov, J.; Rasilainen, S.; Kallio, E.; Korsgren, O.; Häyry, P. Differentiation and maturation of porcine fetal islet cells in vitro and after transplantation. Transplantation 1999, 68, 1674–1683. [Google Scholar] [CrossRef]
- Li, W.C.; Chen, C.Y.; Kao, C.W.; Huang, P.C.; Hsieh, Y.T.; Kuo, T.Y.; Chen, T.Y.; Chia, H.Y.; Juang, J.H. Porcine Neonatal Pancreatic Cell Clusters Maintain Their Multipotency in Culture and After Transplantation. Sci. Rep. 2018, 8, 8212. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Hao, J.; Pan, Y.; Luo, B.; Westgard, B.; Heremans, Y.; Sutherland, D.E.; Hering, B.J.; Guo, Z. Increasing donor chimerism and inducing tolerance to islet allografts by post-transplant donor lymphocyte infusion. Am. J. Transpl. 2006, 6, 933–946. [Google Scholar] [CrossRef]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef]
- Lorza-Gil, E.; Kaiser, G.; Rexen Ulven, E.; König, G.M.; Gerst, F.; Oquendo, M.B.; Birkenfeld, A.L.; Häring, H.U.; Kostenis, E.; Ulven, T.; et al. FFA2-, but not FFA3-agonists inhibit GSIS of human pseudoislets: A comparative study with mouse islets and rat INS-1E cells. Sci. Rep. 2020, 10, 16497. [Google Scholar] [CrossRef] [PubMed]
- Dewanjee, S.; Vallamkondu, J.; Kalra, R.S.; Chakraborty, P.; Gangopadhyay, M.; Sahu, R.; Medala, V.; John, A.; Reddy, P.H.; De Feo, V.; et al. The Emerging Role of HDACs: Pathology and Therapeutic Targets in Diabetes Mellitus. Cells 2021, 10, 1340. [Google Scholar] [CrossRef]
- Liu, J.L.; Segovia, I.; Yuan, X.L.; Gao, Z.H. Controversial Roles of Gut Microbiota-Derived Short-Chain Fatty Acids (SCFAs) on Pancreatic β-Cell Growth and Insulin Secretion. Int. J. Mol. Sci. 2020, 21, 910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haumaitre, C.; Lenoir, O.; Scharfmann, R. Histone deacetylase inhibitors modify pancreatic cell fate determination and amplify endocrine progenitors. Mol. Cell. Biol. 2008, 28, 6373–6383. [Google Scholar] [CrossRef] [Green Version]
- Lundh, M.; Christensen, D.P.; Rasmussen, D.N.; Mascagni, P.; Dinarello, C.A.; Billestrup, N.; Grunnet, L.G.; Mandrup-Poulsen, T. Lysine deacetylases are produced in pancreatic beta cells and are differentially regulated by proinflammatory cytokines. Diabetologia 2010, 53, 2569–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosley, A.L.; Ozcan, S. Glucose regulates insulin gene transcription by hyperacetylation of histone h4. J. Biol. Chem. 2003, 278, 19660–19666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosley, A.L.; Ozcan, S. The pancreatic duodenal homeobox-1 protein (Pdx-1) interacts with histone deacetylases Hdac-1 and Hdac-2 on low levels of glucose. J. Biol. Chem. 2004, 279, 54241–54247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Jena, G.B. Protective role of sodium butyrate, a HDAC inhibitor on beta-cell proliferation, function and glucose homeostasis through modulation of p38/ERK MAPK and apoptotic pathways: Study in juvenile diabetic rat. Chem. -Biol. Interact. 2014, 213, 1–12. [Google Scholar] [CrossRef]
- Lenoir, O.; Flosseau, K.; Ma, F.X.; Blondeau, B.; Mai, A.; Bassel-Duby, R.; Ravassard, P.; Olson, E.N.; Haumaitre, C.; Scharfmann, R. Specific control of pancreatic endocrine β- and δ-cell mass by class IIa histone deacetylases HDAC4, HDAC5, and HDAC9. Diabetes 2011, 60, 2861–2871. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence 5′-3′ | Accession No. |
|---|---|---|
| INS | Forward: CAGGCCTTCGTGAACCAG | NM_001109772.1 |
| Reverse: CTTGGGCGTGTAGAAGAAGC | ||
| GCG | Forward: GAATTCATTGCTTGGCTGGT | NM_214324.1 |
| Reverse: CATCTGAGAAGGAGCCATCAG | ||
| PDX1 | Forward: GTGGAAAAAGGAGGAGGACA | NM_001141984.3 |
| Reverse: CAGCTCCTCTCCCGAGGT | ||
| NKX6.1 | Forward: GCCTACCCCGTTTCAGTAGC | XM_021101796.1 |
| Reverse: GGGTGGACTCTGCATCACTC | ||
| NGN3 | Forward: GCCTGCGTCTCAGCTGAACTT | XM_021072424.1 |
| Reverse: AGCCAGAGGCAGGAGGAACAA | ||
| ALDOB | Forward: ATTTGGAGGGCACTCTGTTG | XM_021066854.1 |
| Reverse: AGGTTGATAGCATTGAGGTTGAG | ||
| GPR43 | Forward: TCATGGGTTTCGGCTTCTACAG | EU122439.1 |
| Reverse: GTACTGAACGATGAACACGACG | ||
| GPR41 | Forward: ACTACTTCTCATCCTCGGGGTT | JX566879.1 |
| Reverse: CTCCACTTCGCTCTTCTTCAGT | ||
| GAPDH | Forward: GTCGGTTGTGGATCTGACCT | NM_001206359.1 |
| Reverse: GTCCTCAGTGTAGCCCAGGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Lei, Y.; Honarpisheh, M.; Kemter, E.; Wolf, E.; Seissler, J. Butyrate and Class I Histone Deacetylase Inhibitors Promote Differentiation of Neonatal Porcine Islet Cells into Beta Cells. Cells 2021, 10, 3249. https://doi.org/10.3390/cells10113249
Zhang Y, Lei Y, Honarpisheh M, Kemter E, Wolf E, Seissler J. Butyrate and Class I Histone Deacetylase Inhibitors Promote Differentiation of Neonatal Porcine Islet Cells into Beta Cells. Cells. 2021; 10(11):3249. https://doi.org/10.3390/cells10113249
Chicago/Turabian StyleZhang, Yichen, Yutian Lei, Mohsen Honarpisheh, Elisabeth Kemter, Eckhard Wolf, and Jochen Seissler. 2021. "Butyrate and Class I Histone Deacetylase Inhibitors Promote Differentiation of Neonatal Porcine Islet Cells into Beta Cells" Cells 10, no. 11: 3249. https://doi.org/10.3390/cells10113249
APA StyleZhang, Y., Lei, Y., Honarpisheh, M., Kemter, E., Wolf, E., & Seissler, J. (2021). Butyrate and Class I Histone Deacetylase Inhibitors Promote Differentiation of Neonatal Porcine Islet Cells into Beta Cells. Cells, 10(11), 3249. https://doi.org/10.3390/cells10113249