
Effect of High Static Magnetic Fields on Biological Activities and Iron Metabolism in MLO-Y4 Osteocyte-like Cells

 ,
,  and
and 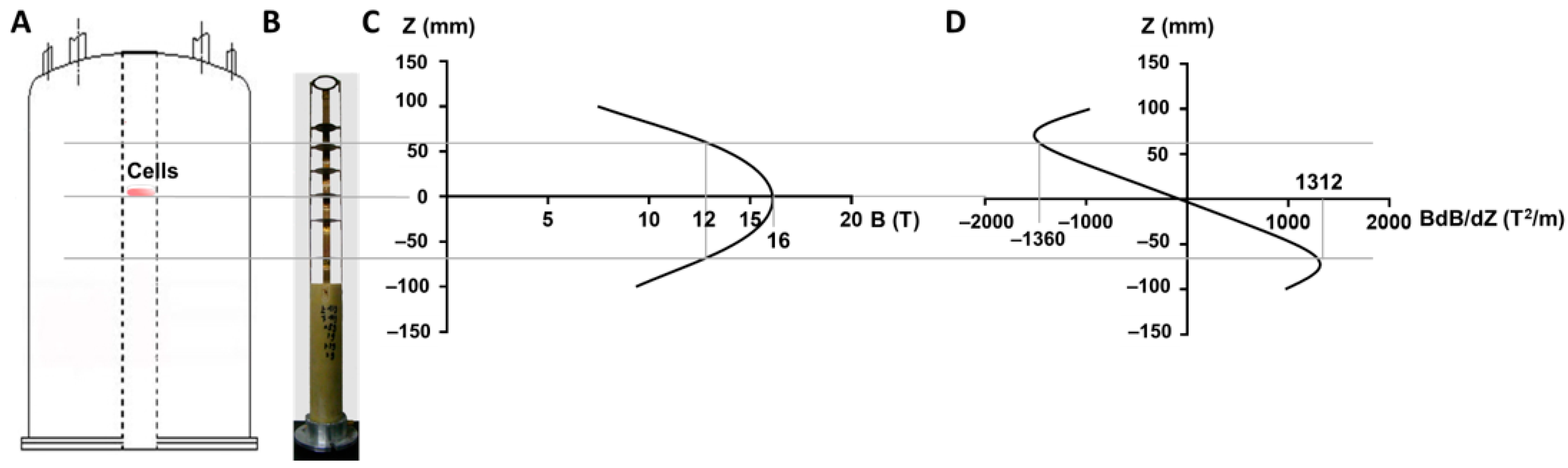{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Magnetic Field Exposure System
2.2. Cell Culture
2.3. Cell Viability Assay
2.4. Cell Morphology Observation
2.5. Cell Apoptosis Assay
2.6. Cytoskeleton Staining
2.7. Soluble Cytokines Assay
2.8. Cellular Iron Content Assay
2.9. Protein Expression Assay
2.10. Statistical Analysis
3. Results
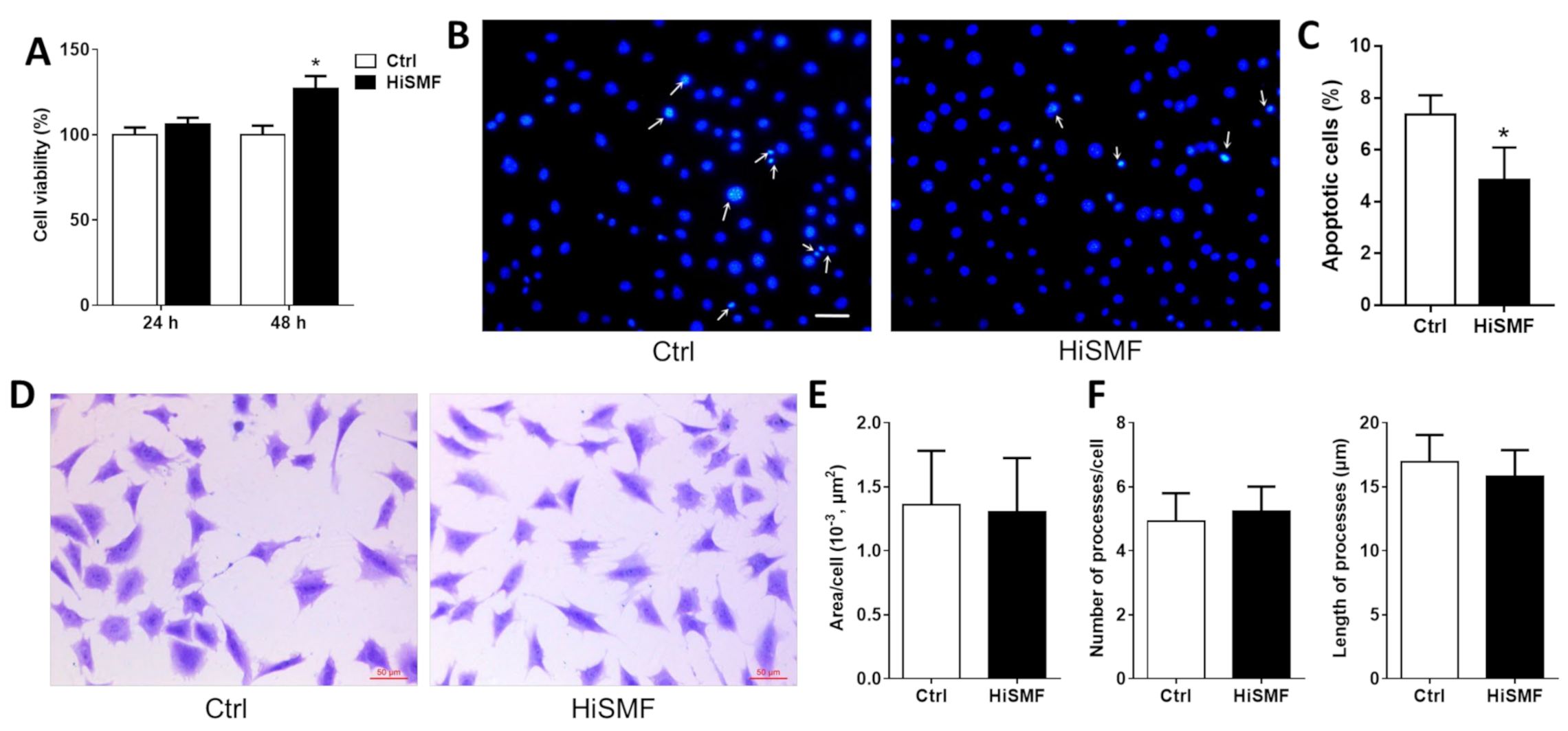
3.1. The Effect of HiSMF on Cell Growth
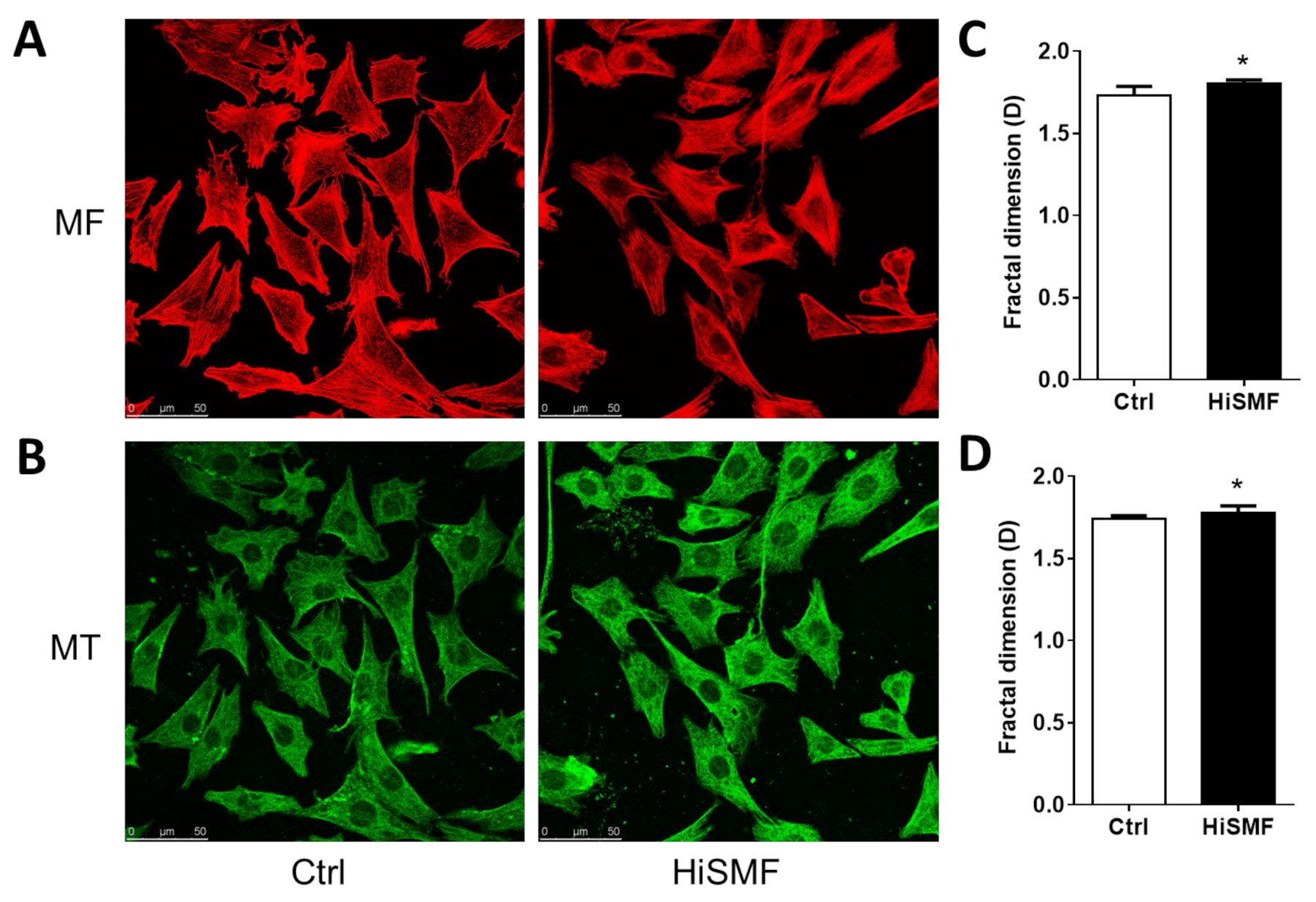
3.2. The Effect of HiSMF on Cytoskeleton
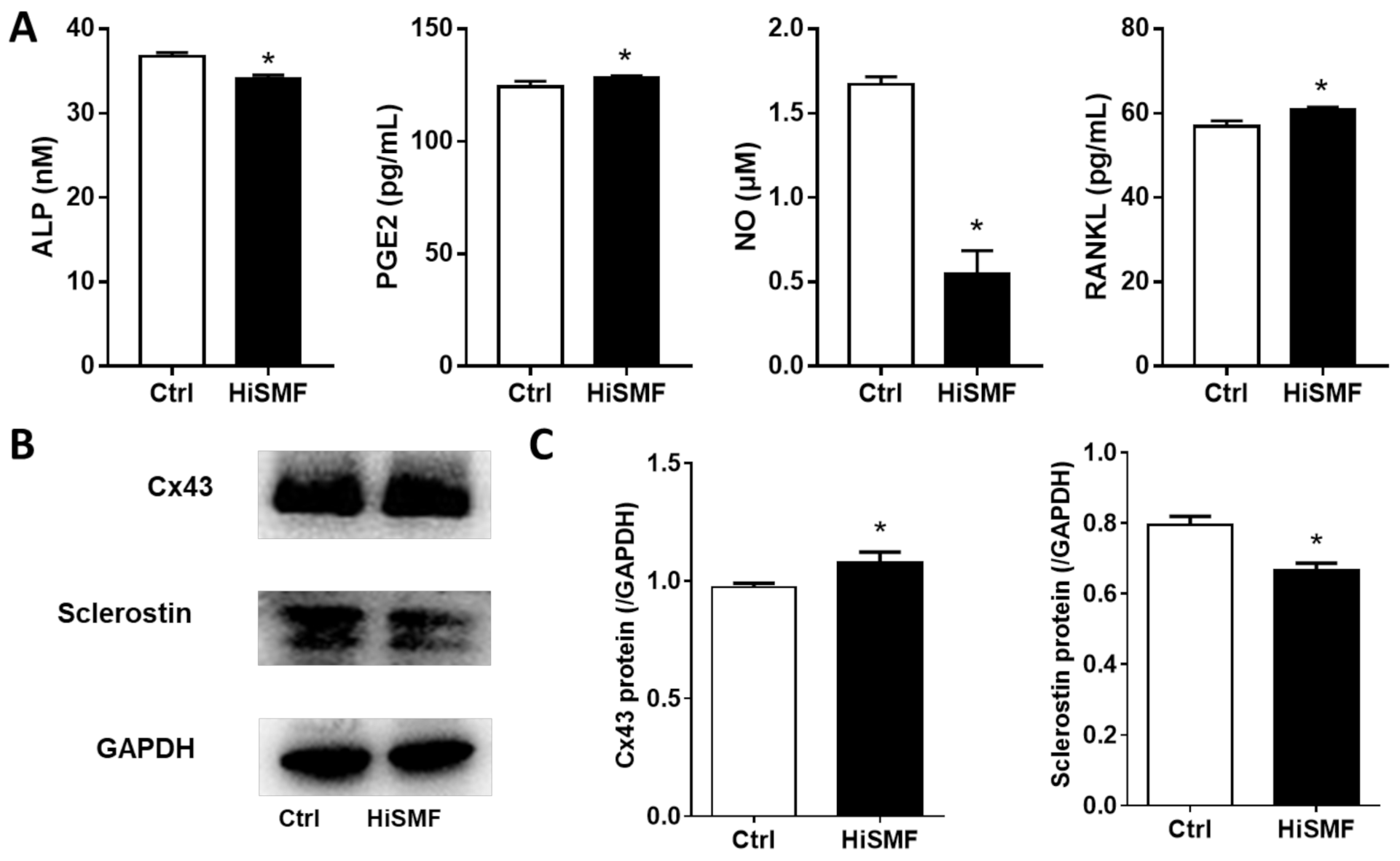
3.3. The Effect of HiSMF on the Secretion of Soluble Cytokines and the Expression of Functional Proteins
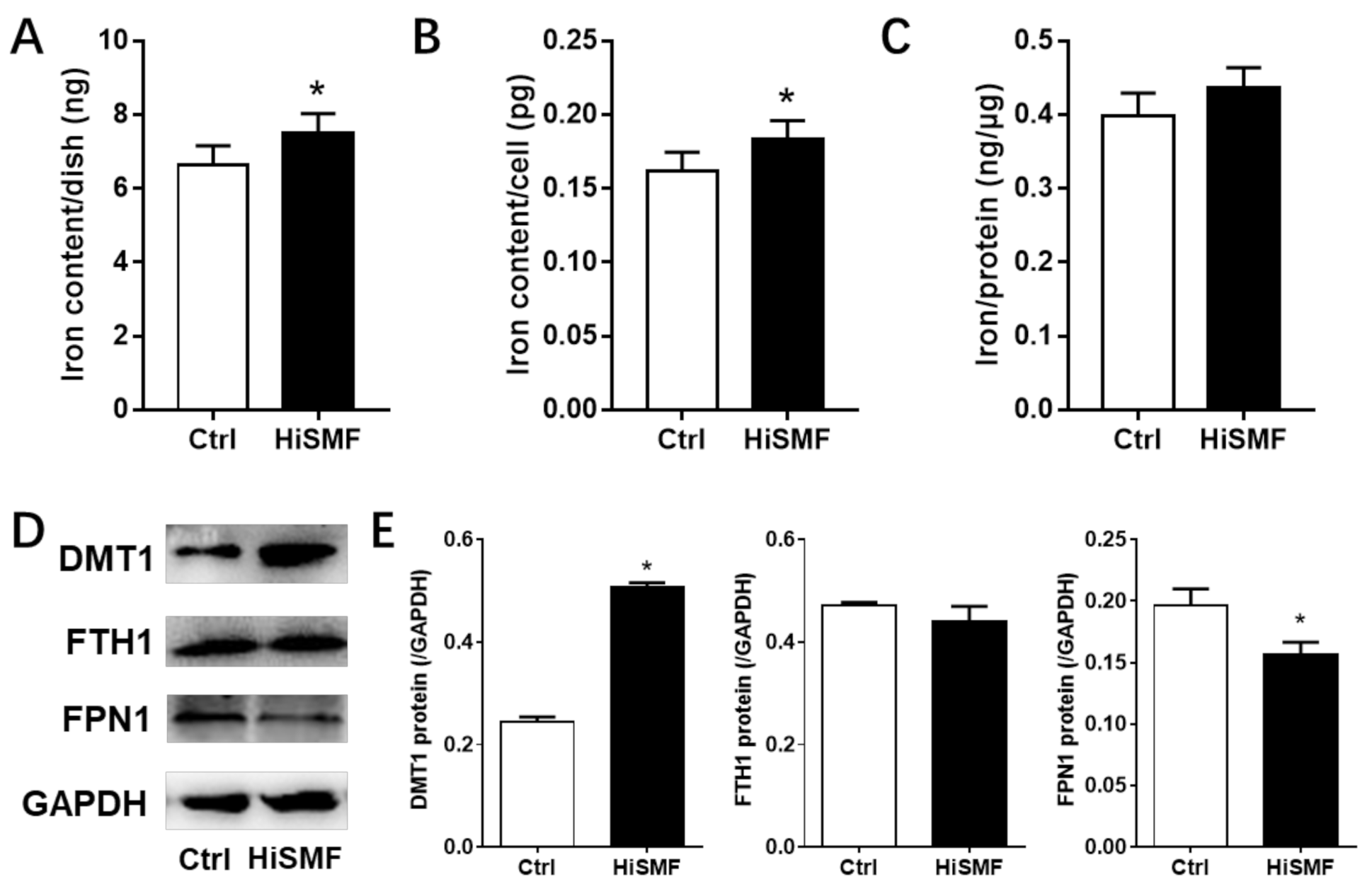
3.4. The Effect of HiSMF on Cellular Iron Metabolism
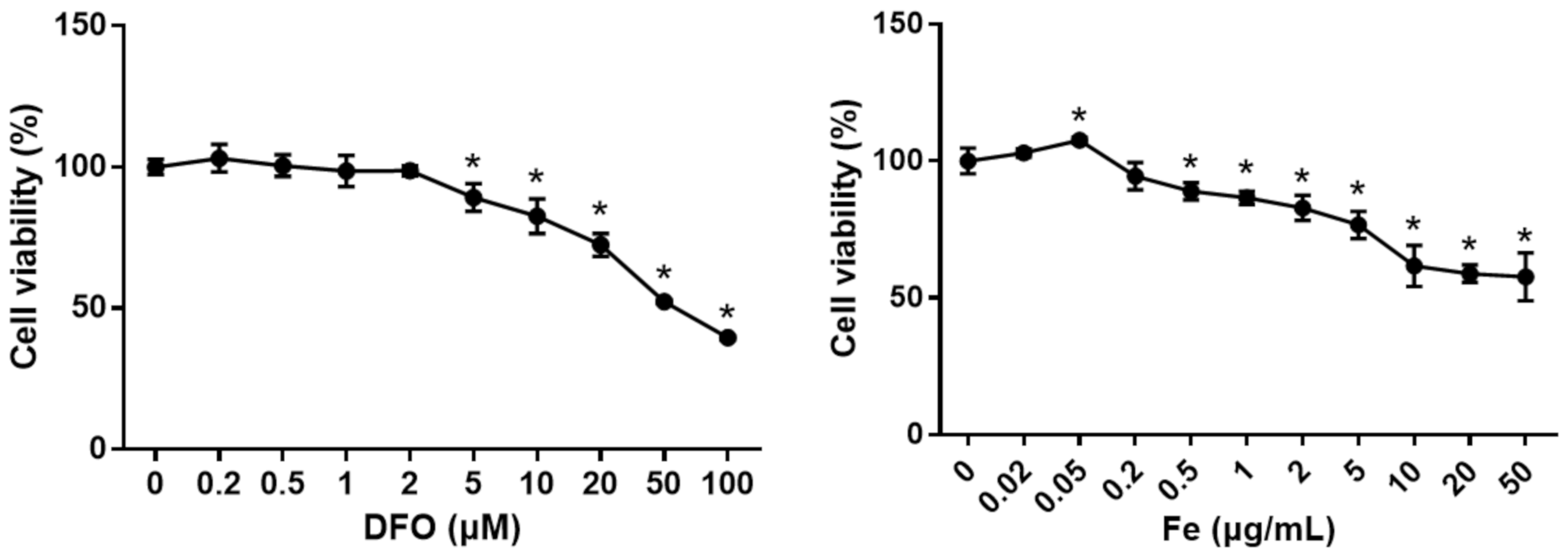
3.5. The Effect of Different Levels of Iron on Cell Viability
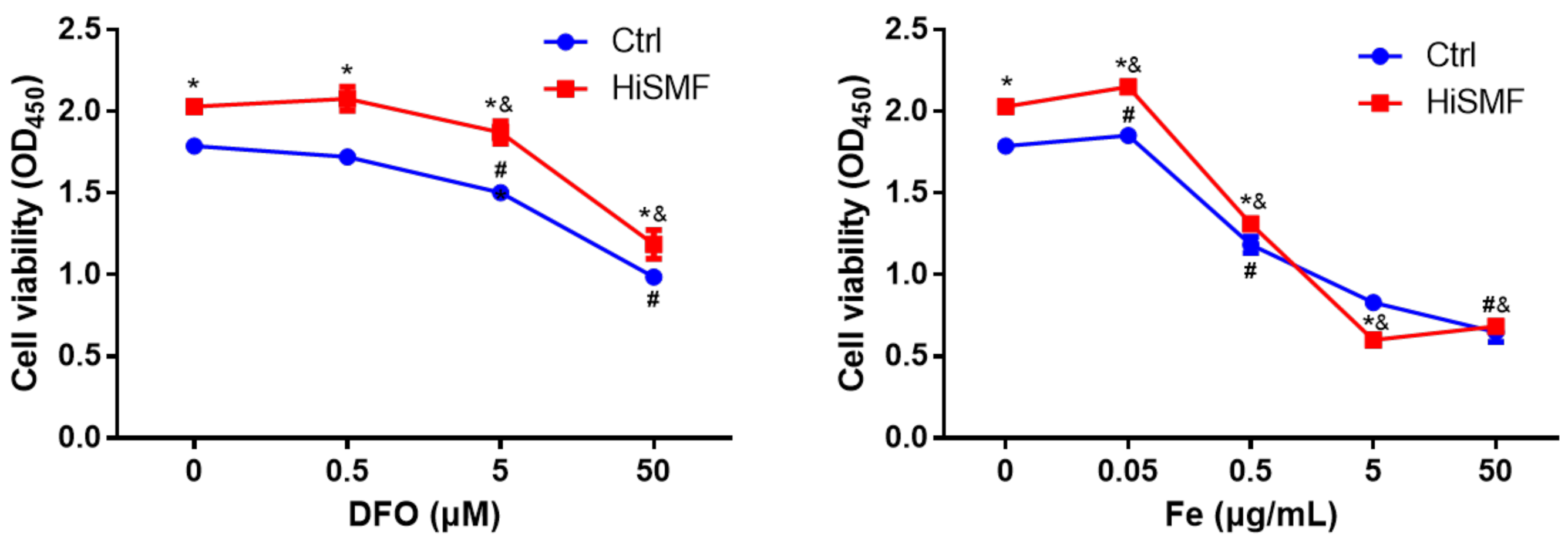
3.6. The Effect of Different Levels of Iron on Cell Viability Under HiSMF
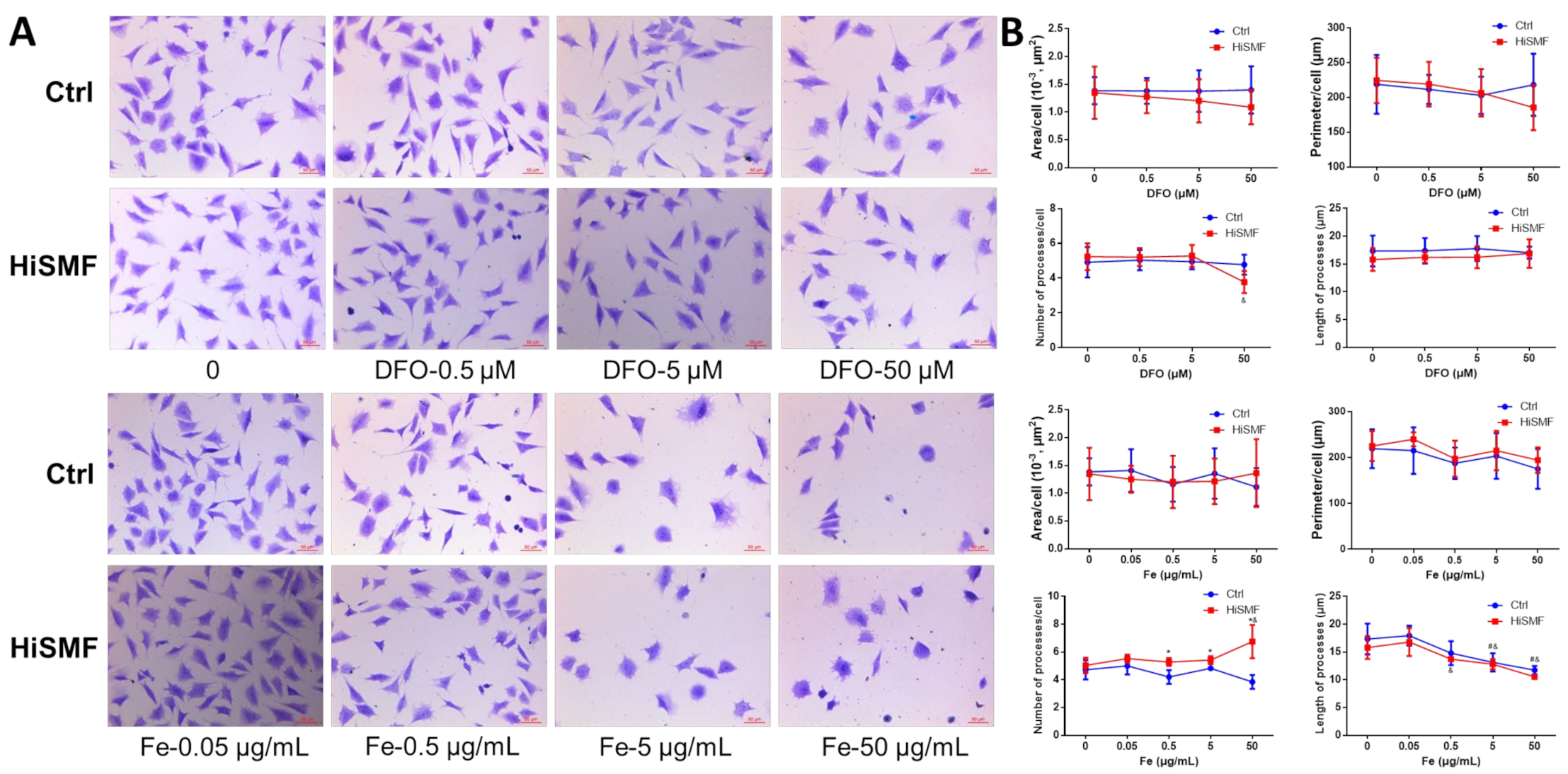
3.7. The Effect of Different Levels of Iron in HiSMF on Cell Morphology
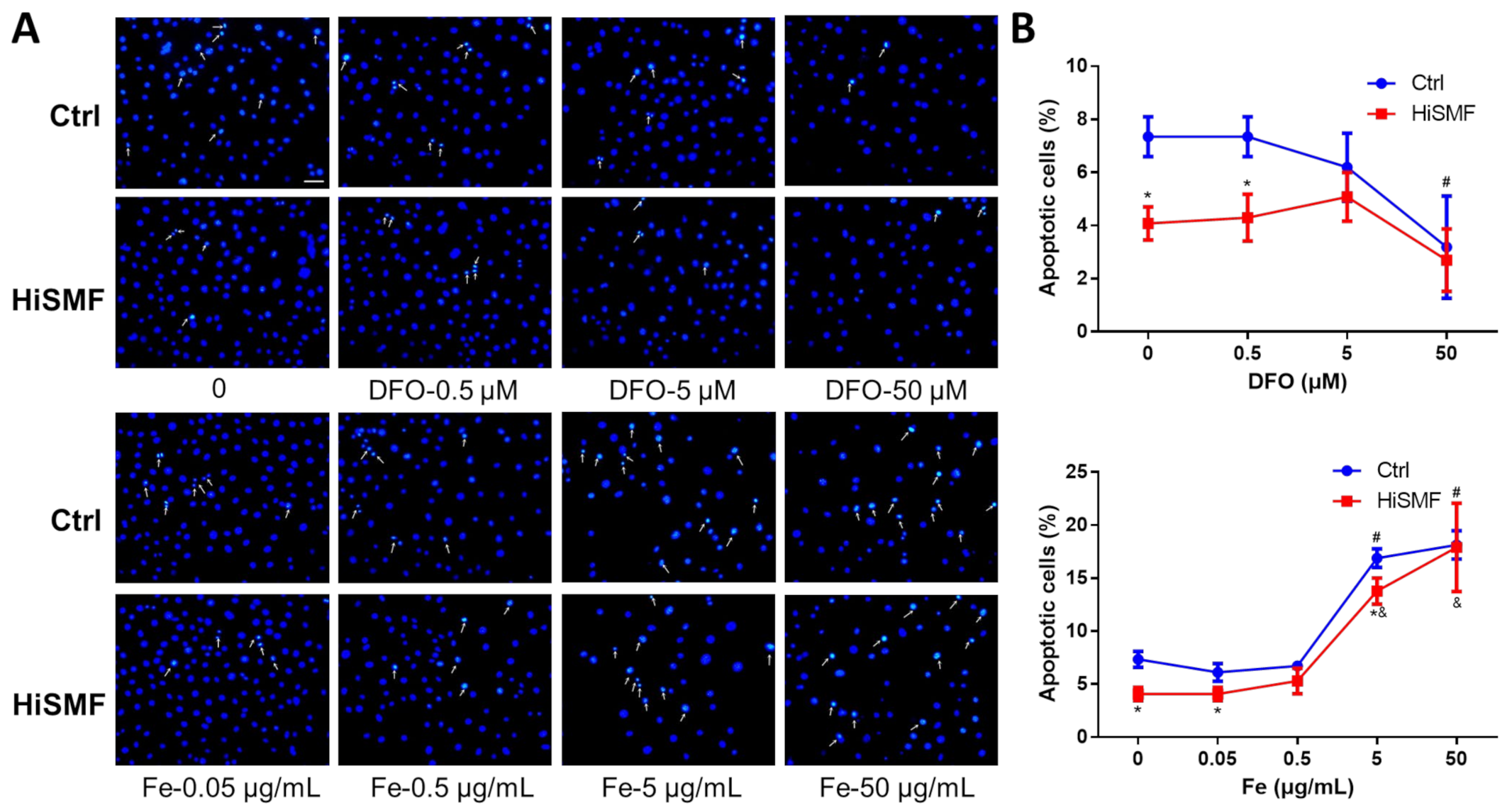
3.8. The Effect of Different Levels of Iron in HiSMF on Cell Apoptosis
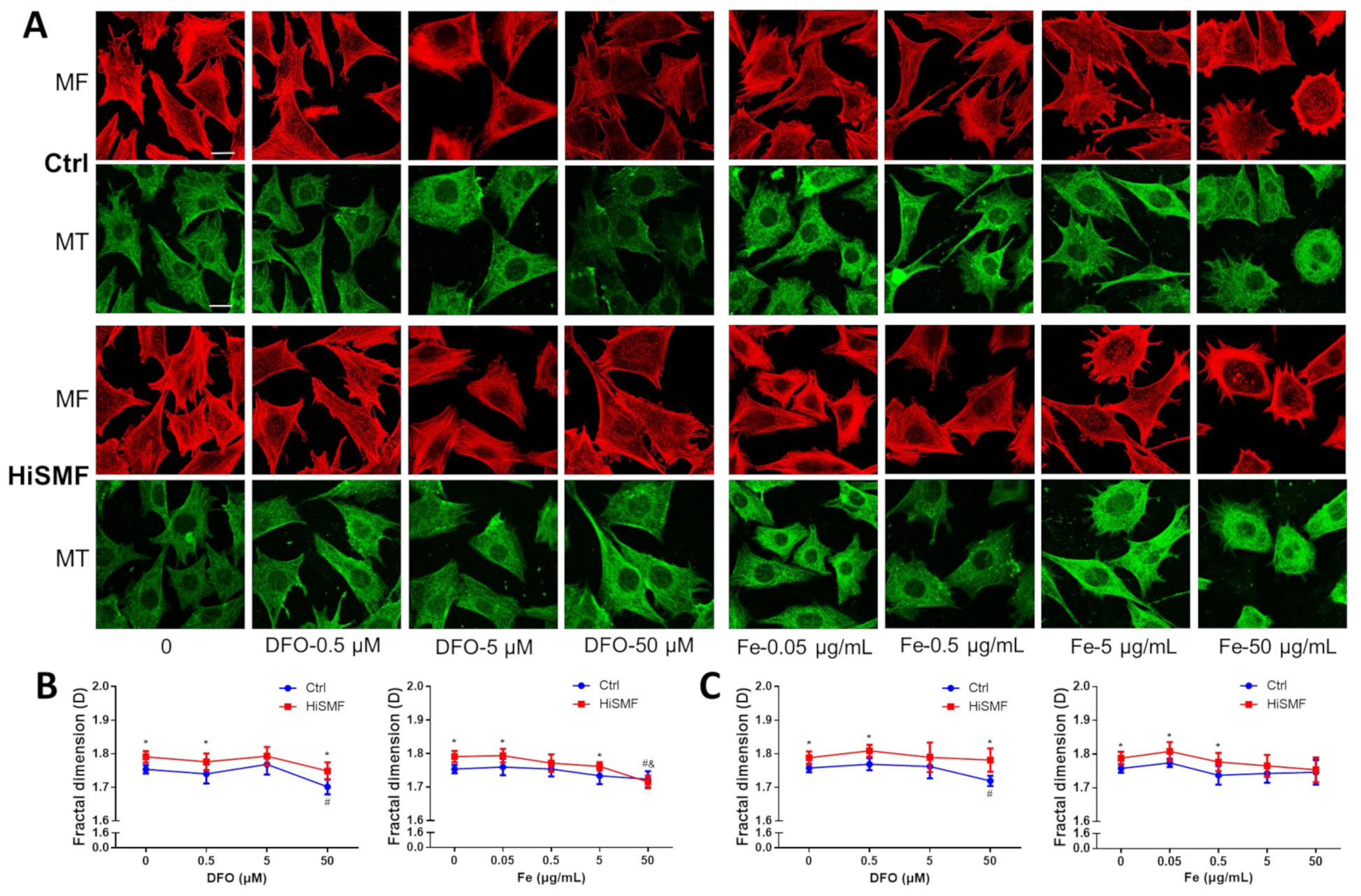
3.9. The Effect of Different Levels of Iron in HiSMF on the Cytoskeleton
3.10. The Effect of Different Concentrations of Iron in HiSMF on the Secretion of Soluble Cytokines
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, C.A.; Kang, C.K.; Kim, Y.B.; Cho, Z.H. Advances in MR Angiography with 7 T MRI: From Microvascular Imaging to Functional Angiography. Neuroimage 2017, 168, 269–278. [Google Scholar] [CrossRef]
- Nowogrodzki, A. The world’s strongest MRI machines are pushing human imaging to new limits. Nature 2018, 563, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Nagel, A.M.; Umathum, R.; Rösler, M.B.; Ladd, M.E.; Litvak, I.; Gor’kov, P.L.; Brey, W.W.; Schepkin, V.D. (39) K and (23) Na relaxation times and MRI of rat head at 21.1 T. NMR Biomed. 2016, 29, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Windle, J.J.; Koop, B.A.; Mundy, G.R.; Bonewald, L.F. Establishment of an osteocyte-like cell line, MLO-Y4. J. Bone Miner. Res. 1997, 12, 2014–2023. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Z.H.; Yin, C.; Ma, J.H.; Li, D.J.; Zhao, F.; Sun, Y.L.; Hu, L.F.; Shang, P.; Qian, A.R. GeneChip expression profiling reveals the alterations of energy metabolism related genes in osteocytes under large gradient high magnetic fields. PLoS ONE 2015, 10, e0116359. [Google Scholar]
- Xu, H.; Ning, D.; Zhao, D.; Chen, Y.; Zhao, D.; Gu, S.; Jiang, J.; Shang, P. Blockage of hemichannels alters gene expression in osteocytes in a high magneto-gravitational environment. Front. Biosci. Landmark Ed. 2017, 22, 783–794. [Google Scholar]
- Qian, A.; Wang, L.; Gao, X.; Zhang, W.; Hu, L.; Han, J.; Li, J.; Di, S.; Shang, P. Diamagnetic Levitation Causes Changes in the Morphology, Cytoskeleton, and Focal Adhesion Proteins Expression in Osteocytes. IEEE Trans. Biomed. Eng. 2012, 59, 68–77. [Google Scholar] [CrossRef]
- Crielaard, B.J.; Lammers, T.; Rivella, S. Targeting iron metabolism in drug discovery and delivery. Nat. Rev. Drug. Discov. 2017, 16, 400–423. [Google Scholar] [CrossRef] [Green Version]
- Dietz, J.V.; Fox, J.L.; Khalimonchuk, O. Down the Iron Path: Mitochondrial Iron Homeostasis and Beyond. Cells 2021, 10, 2198. [Google Scholar] [CrossRef]
- Blesia, V.; Patel, V.B.; Al-Obaidi, H.; Renshaw, D.; Zariwala, M.G. Excessive Iron Induces Oxidative Stress Promoting Cellular Perturbations and Insulin Secretory Dysfunction in MIN6 Beta Cells. Cells 2021, 10, 1141. [Google Scholar] [CrossRef]
- Ishii, K.A.; Fumoto, T.; Iwai, K.; Takeshita, S.; Ito, M.; Shimohata, N.; Aburatani, H.; Taketani, S.; Lelliott, C.J.; Vidal-Puig, A.; et al. Coordination of PGC-1beta and iron uptake in mitochondrial biogenesis and osteoclast activation. Nat. Med. 2009, 15, 259–266. [Google Scholar] [CrossRef]
- Wang, L.; Fang, B.; Fujiwara, T.; Krager, K.; Gorantla, A.; Li, C.; Feng, J.Q.; Jennings, M.L.; Zhou, J.; Aykin-Burns, N.; et al. Deletion of ferroportin in murine myeloid cells increases iron accumulation and stimulates osteoclastogenesis in vitro and in vivo. J. Biol. Chem. 2018, 293, 9248–9264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Jian, J.; Abramson, S.B.; Huang, X. Inhibitory effects of iron on bone morphogenetic protein 2-induced osteoblastogenesis. J. Bone Miner. Res. 2011, 26, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Xu, F.; Cao, Y.; Xu, L.; Yu, C.; Yang, F.; Zhang, P.; Wang, L.; Shen, G.; Wang, J.; et al. Iron Accumulation Leads to Bone Loss by Inducing Mesenchymal Stem Cell Apoptosis Through the Activation of Caspase3. Biol. Trace Elem. Res. 2018, 187, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Li, X.; Zhu, Z.; Wang, H.; Bai, X. Iron Overload Induces Apoptosis and Cytoprotective Autophagy Regulated by ROS Generation in Mc3t3-E1 Cells. Biol. Trace Elem. Res. 2021, 199, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Baschant, U.; Rauner, M.; Bulycheva, E.; Weidner, H.; Roetto, A.; Platzbecker, U.; Hofbauer, L.C. Wnt5a is a key target for the pro-osteogenic effects of iron chelation on osteoblast progenitors. Haematologica 2016, 101, 1499–1507. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hu, W.; Ding, C.; Yao, G.; Zhao, H.; Wu, S. Deferoxamine inhibits iron-uptake stimulated osteoclast differentiation by suppressing electron transport chain and MAPKs signaling. Toxicol. Lett. 2019, 313, 50–59. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Ding, C.; Dong, D.; Shang, P. Regulation of Osteoblast Differentiation and Iron Content in MC3T3-E1 Cells by Static Magnetic Field with Different Intensities. Biol. Trace Elem. Res. 2018, 184, 214–225. [Google Scholar] [CrossRef] [Green Version]
- Dong, D.; Yang, J.; Zhang, G.; Huyan, T.; Shang, P. 16 T high static magnetic field inhibits receptor activator of nuclear factor kappa-Β ligand-induced osteoclast differentiation by regulating iron metabolism in Raw264.7 cells. J. Tissue Eng. Regen. Med. 2019, 13, 2181–2190. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, Z.; Ding, C.; Wang, Y.; Shang, P.; Qian, A.R. Experimental Conditions Optimization for Cell Culture in Large Gradient High Magnetic Field. Space Med. Med. Eng. 2015, 28, 79–84. [Google Scholar]
- Yang, J.; Meng, X.; Dong, D.; Xue, Y.; Chen, X.; Wang, S.; Shen, Y.; Zhang, G.; Shang, P. Iron overload involved in the enhancement of unloading-induced bone loss by hypomagnetic field. Bone 2018, 114, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, T.; Dehghani, F. The Cytoskeleton-A Complex Interacting Meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, D.A.; Mullins, R.D. Cell mechanics and the cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franke, W.W.; Hergt, M.; Grund, C. Rearrangement of the vimentin cytoskeleton during adipose conversion: Formation of an intermediate filament cage around lipid globules. Cell 1987, 49, 131–141. [Google Scholar] [CrossRef]
- Thomason, D.B.; Anderson, O., III; Menon, V. Fractal analysis of cytoskeleton rearrangement in cardiac muscle during head-down tilt. J. Appl. Physiol. 1996, 81, 1522–1527. [Google Scholar] [CrossRef]
- Qian, A.R.; Li, D.; Han, J.; Gao, X.; Di, S.M.; Zhang, W.; Hu, L.F.; Shang, P. Fractal dimension as a measure of altered actin cytoskeleton in MC3T3-E1 cells under simulated microgravity using 3-D/2-D clinostats. IEEE Trans. Biomed. Eng. 2012, 59, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Fuseler, J.W.; Millette, C.F.; Davis, J.M.; Carver, W. Fractal and image analysis of morphological changes in the actin cytoskeleton of neonatal cardiac fibroblasts in response to mechanical stretch. Microsc. Microanal. 2007, 13, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Calle, J.M.; Bellido, T. The Osteocyte as a Signaling Cell. Physiol. Rev. 2021, 102, 379–410. [Google Scholar] [CrossRef]
- Winkler, D.G.; Sutherland, M.K.; Geoghegan, J.C.; Yu, C.; Hayes, T.; Skonier, J.E.; Shpektor, D.; Jonas, M.; Kovacevich, B.R.; Staehling-Hampton, K.; et al. Osteocyte control of bone formation via sclerostin, a novel BMP antagonist. Embo J. 2003, 22, 6267–6276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.; Jiang, X.; Dai, Z.; Guo, X.; Weng, T.; Wang, J.; Li, Y.; Feng, G.; Gao, X.; He, L. Sclerostin mediates bone response to mechanical unloading through antagonizing Wnt/beta-catenin signaling. J. Bone Miner. Res. 2009, 24, 1651–1661. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, H.; Pilbeam CCHarrison, J.R.; Raisz, L.G. The role of prostaglandins in the regulation of bone metabolism. Clin. Orthop. Relat. Res. 1995, 313, 36–46. [Google Scholar]
- Zhang, J.; Ding, C.; Meng, X.; Shang, P. Nitric oxide modulates the responses of osteoclast formation to static magnetic fields. Electromagn. Biol. Med. 2017, 37, 23–34. [Google Scholar] [CrossRef]
- McDonald, M.M.; Khoo, W.H.; Ng, P.Y.; Xiao, Y.; Zamerli, J.; Thatcher, P.; Kyaw, W.; Pathmanandavel, K.; Grootveld, A.K.; Moran, I.; et al. Osteoclasts recycle via osteomorphs during RANKL-stimulated bone resorption. Cell 2021, 184, 1330–1347. [Google Scholar] [CrossRef]
- Li, G.; Zhang, L.; Ning, K.; Yang, B.; Acosta, F.M.; Shang, P.; Jiang, J.X.; Xu, H. Osteocytic Connexin43 Channels Regulate Bone-Muscle Crosstalk. Cells 2021, 10, 237. [Google Scholar] [CrossRef]
- De Vernejoul, M.C.; Pointillart, A.; Golenzer, C.C.; Morieux, C.; Bielakoff, J.; Modrowski, D.; Miravet, L. Effects of iron overload on bone remodeling in pigs. Am. J. Pathol. 1984, 116, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Balogh, E.; Paragh, G.; Jeney, V. Influence of Iron on Bone Homeostasis. Pharmaceuticals 2018, 11, E107. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Dong, D.; Luo, X.; Zhou, J.; Shang, P.; Zhang, H. Iron Overload-Induced Osteocyte Apoptosis Stimulates Osteoclast Differentiation Through Increasing Osteocytic RANKL Production In Vitro. Calcif. Tissue Int. 2020, 107, 499–509. [Google Scholar] [CrossRef]
- Robling, A.G.; Bonewald, L.F. The Osteocyte: New Insights. Annu. Rev. Physiol. 2020, 82, 485–506. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.S.; Kamath, T.; Mazur, C.M.; Mirzamohammadi, F.; Rotter, D.; Hojo, H.; Castro, C.D.; Tokavanich, N.; Patel, R.; Govea, N.; et al. Control of osteocyte dendrite formation by Sp7 and its target gene osteocrin. Nat. Commun. 2021, 12, 6271. [Google Scholar] [CrossRef]
- Tiede-Lewis, L.M.; Dallas, S.L. Changes in the osteocyte lacunocanalicular network with aging. Bone 2019, 122, 101–113. [Google Scholar] [CrossRef]
- Fowler, T.W.; Acevedo, C.; Mazur, C.M.; Hall-Glenn, F.; Fields, A.J.; Bale, H.A.; Ritchie, R.O.; Lotz, J.C.; Vail, T.P.; Alliston, T. Glucocorticoid suppression of osteocyte perilacunar remodeling is associated with subchondral bone degeneration in osteonecrosis. Sci. Rep. 2017, 7, 44618. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Zhang, G.; Li, Q.; Tang, Q.; Feng, Y.; Shang, P.; Zeng, Y. Effect of High Static Magnetic Fields on Biological Activities and Iron Metabolism in MLO-Y4 Osteocyte-like Cells. Cells 2021, 10, 3519. https://doi.org/10.3390/cells10123519
Yang J, Zhang G, Li Q, Tang Q, Feng Y, Shang P, Zeng Y. Effect of High Static Magnetic Fields on Biological Activities and Iron Metabolism in MLO-Y4 Osteocyte-like Cells. Cells. 2021; 10(12):3519. https://doi.org/10.3390/cells10123519
Chicago/Turabian StyleYang, Jiancheng, Gejing Zhang, Qingmei Li, Qinghua Tang, Yan Feng, Peng Shang, and Yuhong Zeng. 2021. "Effect of High Static Magnetic Fields on Biological Activities and Iron Metabolism in MLO-Y4 Osteocyte-like Cells" Cells 10, no. 12: 3519. https://doi.org/10.3390/cells10123519
APA StyleYang, J., Zhang, G., Li, Q., Tang, Q., Feng, Y., Shang, P., & Zeng, Y. (2021). Effect of High Static Magnetic Fields on Biological Activities and Iron Metabolism in MLO-Y4 Osteocyte-like Cells. Cells, 10(12), 3519. https://doi.org/10.3390/cells10123519