
Plant Viruses Can Alter Aphid-Triggered Calcium Elevations in Infected Leaves

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Aphids, Plants, and Viruses
2.2. Image Acquisition
2.3. Image Analysis
2.4. Transcriptome Analysis
2.5. CaMV Transmission Tests
2.6. Figures and Statistical Analysis
3. Results
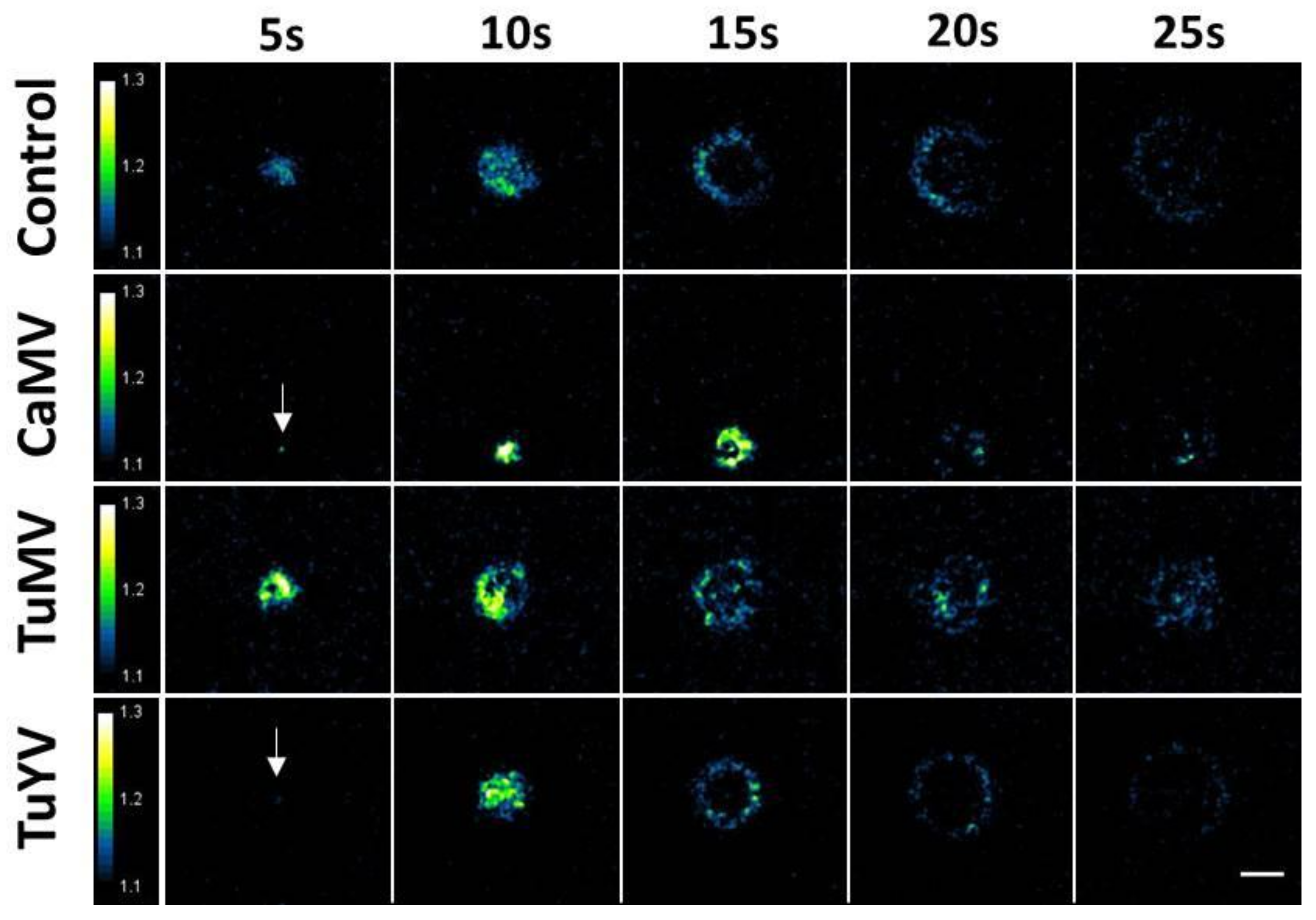
3.1. Aphids Trigger Ring-Like Calcium Waves
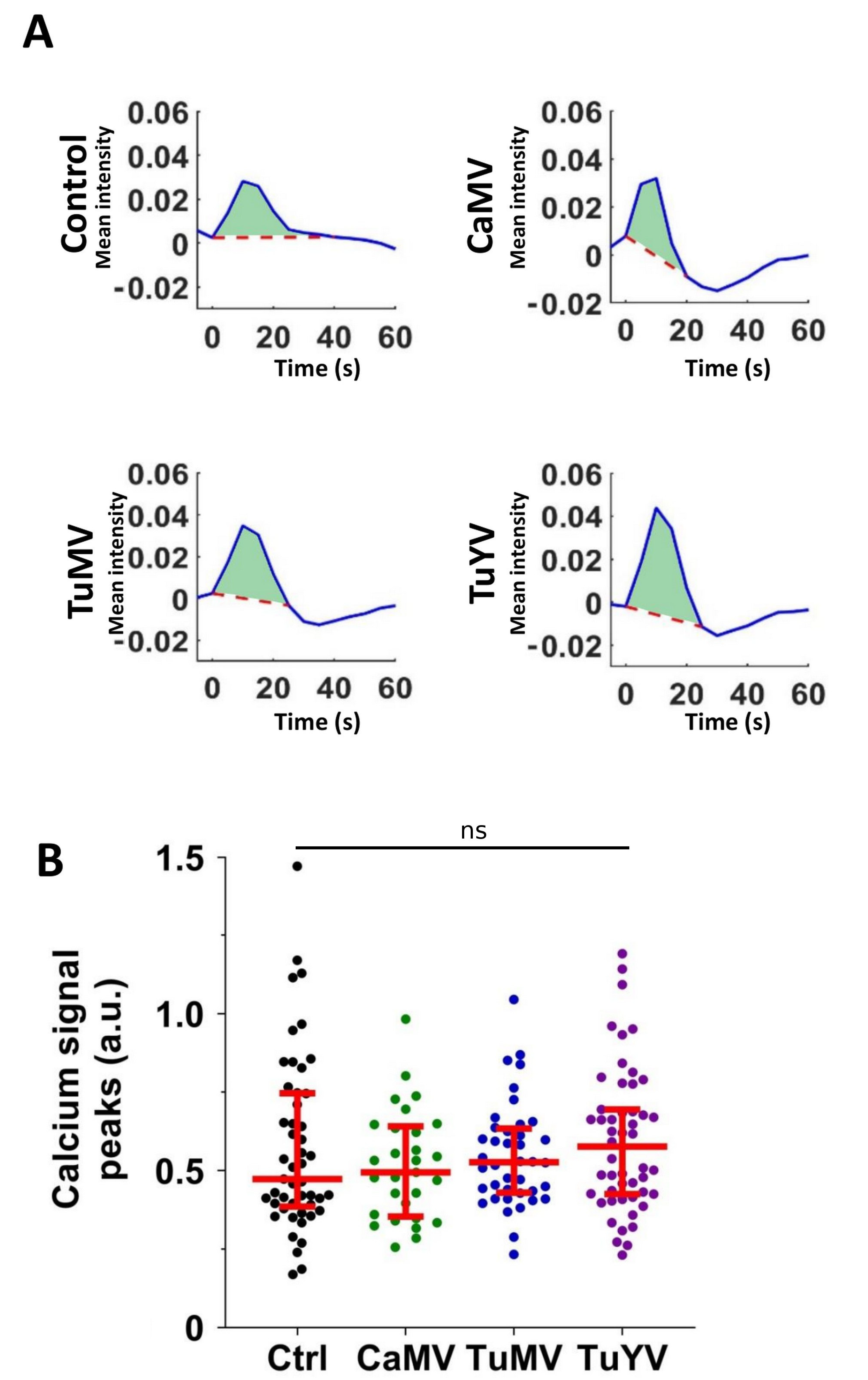
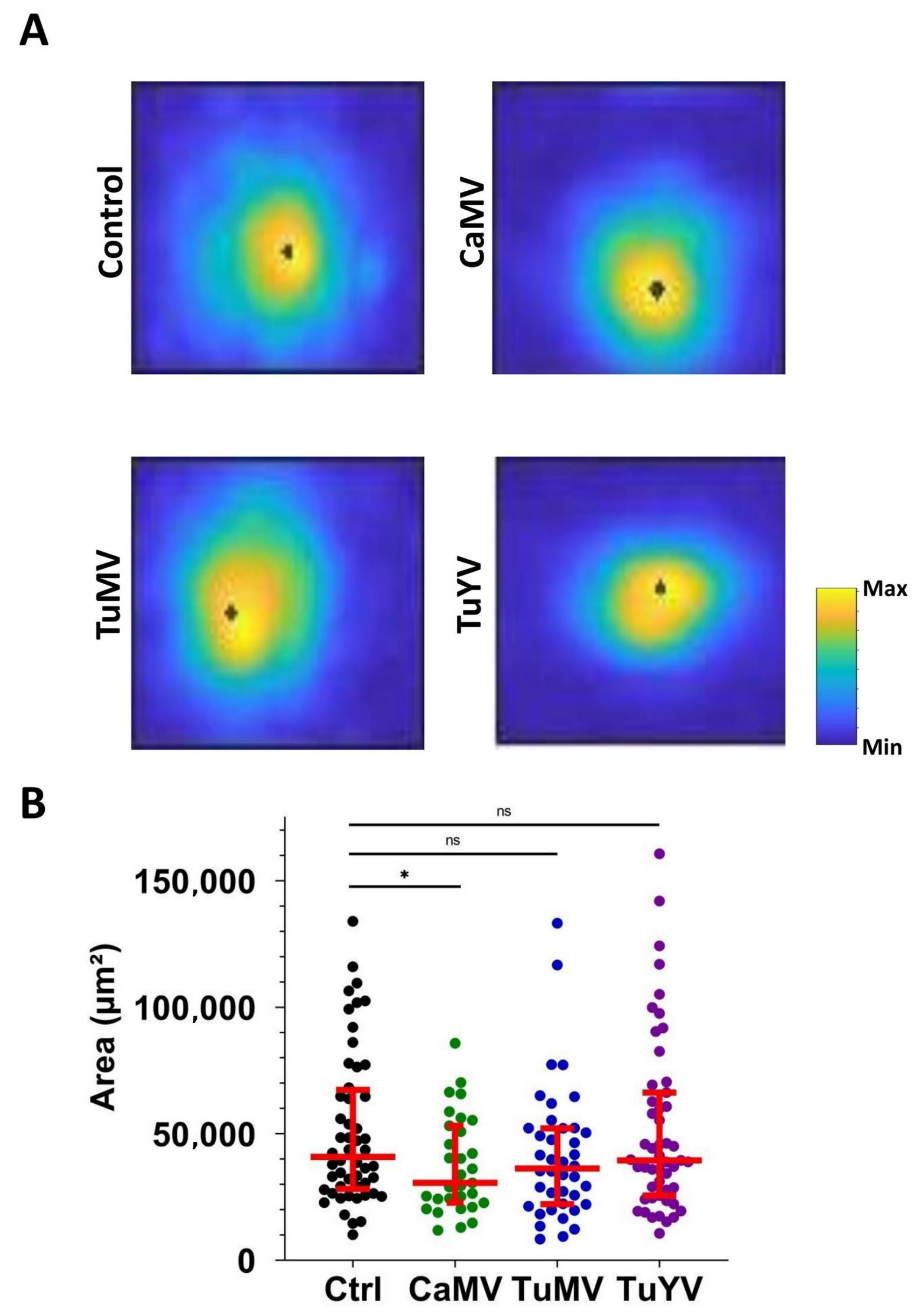
3.2. Characterisation of the Calcium Waves
3.3. CaMV Transmission from Arabidopsis Mutants Impaired in Pathogen Recognition or Signalling
3.4. Screen for Genes Potentially Involved in Aphid Perception
4. Discussion
4.1. Comparison between Our Results and Those Obtained by Vincent et al.
4.2. Calcium Propagation Is Slower in CaMV- and TuMV-Infected Leaves
4.3. Impact of Slower Calcium Propagation on Plant–Aphid Interactions
4.4. Role of BAK1 in Calcium-Mediated Aphid–Plant Interactions
4.5. Involvement of Different Ion Channels in Aphid-Triggered Calcium Elevations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaudhary, R.; Atamian, H.S.; Shen, Z.; Briggs, S.P.; Kaloshian, I. GroEL from the Endosymbiont Buchnera Aphidicola Betrays the Aphid by Triggering Plant Defense. Proc. Natl. Acad. Sci. USA 2014, 111, 8919–8924. [Google Scholar] [CrossRef] [Green Version]
- Prince, D.C.; Drurey, C.; Zipfel, C.; Hogenhout, S.A. The Leucine-Rich Repeat Receptor-like Kinase BRASSINOSTEROID INSENSITIVE1-ASSOCIATED KINASE1 and the Cytochrome P450 PHYTOALEXIN DEFICIENT3 Contribute to Innate Immunity to Aphids in Arabidopsis. Plant Physiol. 2014, 164, 2207–2219. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Nalam, V.; Louis, J.; Shah, J. Plant Defense against Aphids, the Pest Extraordinaire. Plant Sci. 2019, 279, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Prado, E.; Tjallingii, W.F. Aphid Activities during Sieve Element Punctures. Entomol. Exp. Appl. 1994, 72, 157–165. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Hogen Esch, T. Fine Structure of Aphid Stylet Routes in Plant Tissues in Correlation with EPG Signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Escudero-Martinez, C.; Rodriguez, P.A.; Liu, S.; Santos, P.A.; Stephens, J.; Bos, J.I.B. An Aphid Effector Promotes Barley Susceptibility through Suppression of Defence Gene Expression. J. Exp. Bot. 2020, 71, 2796–2807. [Google Scholar] [CrossRef] [Green Version]
- MacWilliams, J.R.; Dingwall, S.; Chesnais, Q.; Sugio, A.; Kaloshian, I. AcDCXR Is a Cowpea Aphid Effector with Putative Roles in Altering Host Immunity and Physiology. Front. Plant Sci. 2020, 11, 605. [Google Scholar] [CrossRef]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.-P. Calcium Signalling in Plant Biotic Interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Feng, B.; Zhou, J.-M.; Tang, D. Plant Immune Signaling: Advancing on Two Frontiers. J. Integr. Plant Biol. 2020, 62, 2–24. [Google Scholar] [CrossRef] [Green Version]
- Vincent, T.R.; Avramova, M.; Canham, J.; Higgins, P.; Bilkey, N.; Mugford, S.T.; Pitino, M.; Toyota, M.; Gilroy, S.; Miller, A.J.; et al. Interplay of Plasma Membrane and Vacuolar Ion Channels, Together with BAK1, Elicits Rapid Cytosolic Calcium Elevations in Arabidopsis during Aphid Feeding. Plant Cell 2017, 29, 1460–1479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Verrall, S.R.; Hancock, R.D. Systematic Analysis of Phloem-Feeding Insect-Induced Transcriptional Reprogramming in Arabidopsis Highlights Common Features and Reveals Distinct Responses to Specialist and Generalist Insects. J. Exp. Bot. 2015, 66, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Dáder, B.; Then, C.; Berthelot, E.; Ducousso, M.; Ng, J.C.K.; Drucker, M. Insect Transmission of Plant Viruses: Multilayered Interactions Optimize Viral Propagation. Insect Sci. 2017, 24, 929–946. [Google Scholar] [CrossRef]
- Bak, A.; Gargani, D.; Macia, J.-L.; Malouvet, E.; Vernerey, M.-S.; Blanc, S.; Drucker, M. Virus Factories of Cauliflower Mosaic Virus Are Virion Reservoirs That Engage Actively in Vector Transmission. J. Virol. 2013, 87, 12207–12215. [Google Scholar] [CrossRef] [Green Version]
- Berthelot, E.; Macia, J.-L.; Martinière, A.; Morisset, A.; Gallet, R.; Blanc, S.; Khelifa, M.; Drucker, M. Pharmacological Analysis of Transmission Activation of Two Aphid-Vectored Plant Viruses, Turnip Mosaic Virus and Cauliflower Mosaic Virus. Sci. Rep. 2019, 9, 9374. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, E.; Ducousso, M.; Macia, J.L.; Bogaert, F.; Baecker, V.; Thébaud, G.; Gallet, R.; Yvon, M.; Blanc, S.; Khelifa, M.; et al. Turnip Mosaic Virus Is a Second Example of a Virus Using Transmission Activation for Plant-to-Plant Propagation by Aphids. J. Virol. 2019, 93, e01822-18. [Google Scholar] [CrossRef] [Green Version]
- Martinière, A.; Bak, A.; Macia, J.-L.; Lautredou, N.; Gargani, D.; Doumayrou, J.; Garzo, E.; Moreno, A.; Fereres, A.; Blanc, S.; et al. A Virus Responds Instantly to the Presence of the Vector on the Host and Forms Transmission Morphs. eLife 2013, 2, e00183. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, Q.; Wu, Y.; Huang, F.; Ismayil, A.; Zhang, D.; Li, H.; Gu, H.; Ludman, M.; Fátyol, K.; et al. A Calmodulin-Binding Transcription Factor Links Calcium Signaling to Antiviral RNAi Defense in Plants. Cell Host Microbe 2021, 29, 1393–1406.e7. [Google Scholar] [CrossRef]
- Casciano, J.C.; Duchemin, N.J.; Lamontagne, R.J.; Steel, L.F.; Bouchard, M.J. Hepatitis B Virus Modulates Store-Operated Calcium Entry to Enhance Viral Replication in Primary Hepatocytes. PLoS ONE 2017, 12, e0168328. [Google Scholar] [CrossRef] [PubMed]
- Negash, A.A.; Olson, R.M.; Griffin, S.; Gale, M. Modulation of Calcium Signaling Pathway by Hepatitis C Virus Core Protein Stimulates NLRP3 Inflammasome Activation. PLoS Pathog 2019, 15, e1007593. [Google Scholar] [CrossRef]
- Krebs, M.; Held, K.; Binder, A.; Hashimoto, K.; Herder, G.D.; Parniske, M.; Kudla, J.; Schumacher, K. FRET-Based Genetically Encoded Sensors Allow High-Resolution Live Cell Imaging of Ca2+ Dynamics. Plant J. 2012, 69, 181–192. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Occhipinti, A.; Maffei, M.E. Calcium Imaging Perspectives in Plants. Int. J. Mol. Sci. 2014, 15, 3842–3859. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Yamada, S.; Tominaga, T.; Ichikawa, M.; Miyawaki, A. Expanded Dynamic Range of Fluorescent Indicators for Ca2+ by Circularly Permuted Yellow Fluorescent Proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 10554–10559. [Google Scholar] [CrossRef] [Green Version]
- Delseny, M.; Hull, R. Isolation and Characterization of Faithful and Altered Clones of the Genomes of Cauliflower Mosaic Virus Isolates Cabb B-JI, CM4-184, and Bari I. Plasmid 1983, 9, 31–41. [Google Scholar] [CrossRef]
- Ohshima, K.; Yamaguchi, Y.; Hirota, R.; Hamamoto, T.; Tomimura, K.; Tan, Z.; Sano, T.; Azuhata, F.; Walsh, J.A.; Fletcher, J.; et al. Molecular Evolution of Turnip Mosaic Virus: Evidence of Host Adaptation, Genetic Recombination and Geographical Spread. J. Gen. Virol. 2002, 83, 1511–1521. [Google Scholar] [CrossRef]
- Bortolamiol-Bécet, D.; Monsion, B.; Chapuis, S.; Hleibieh, K.; Scheidecker, D.; Alioua, A.; Bogaert, F.; Revers, F.; Brault, V.; Ziegler-Graff, V. Phloem-Triggered Virus-Induced Gene Silencing Using a Recombinant Polerovirus. Front. Microbiol. 2018, 9, 2449. [Google Scholar] [CrossRef]
- Xiong, T.C.; Ronzier, E.; Sanchez, F.; Corratgé-Faillie, C.; Mazars, C.; Thibaud, J.-B. Imaging Long Distance Propagating Calcium Signals in Intact Plant Leaves with the BRET-Based GFP-Aequorin Reporter. Front. Plant Sci. 2014, 5, 43. [Google Scholar] [CrossRef]
- Prasch, C.M.; Sonnewald, U. Simultaneous Application of Heat, Drought, and Virus to Arabidopsis Plants Reveals Significant Shifts in Signaling Networks. Plant Physiol. 2013, 162, 1849–1866. [Google Scholar] [CrossRef] [PubMed]
- Basu, D.; Haswell, E.S. Plant Mechanosensitive Ion Channels: An Ocean of Possibilities. Curr. Opin. Plant Biol. 2017, 40, 43–48. [Google Scholar] [CrossRef]
- Mugford, S.T.; Barclay, E.; Drurey, C.; Findlay, K.C.; Hogenhout, S.A. An Immuno-Suppressive Aphid Saliva Protein Is Delivered into the Cytosol of Plant Mesophyll Cells During Feeding. Mol. Plant-Microbe Interact. 2016, 29, 854–861. [Google Scholar] [CrossRef] [Green Version]
- Chesnais, Q.; Verdier, M.; Burckbuchler, M.; Brault, V.; Pooggin, M.; Drucker, M. Cauliflower Mosaic Virus Protein P6-TAV Plays a Major Role in Alteration of Aphid Vector Feeding Behaviour but Not Performance on Infected Arabidopsis. Mol. Plant Pathol. 2021, 22, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Love, A.J.; Yun, B.W.; Laval, V.; Loake, G.J.; Milner, J.J. Cauliflower Mosaic Virus, a Compatible Pathogen of Arabidopsis, Engages Three Distinct Defense-Signaling Pathways and Activates Rapid Systemic Generation of Reactive Oxygen Species. Plant Physiol. 2005, 139, 935–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, A.J.; Geri, C.; Laird, J.; Carr, C.; Yun, B.-W.; Loake, G.J.; Tada, Y.; Sadanandom, A.; Milner, J.J. Cauliflower Mosaic Virus Protein P6 Inhibits Signaling Responses to Salicylic Acid and Regulates Innate Immunity. PLoS ONE 2012, 7, e47535. [Google Scholar] [CrossRef] [Green Version]
- Casteel, C.L.; De Alwis, M.; Bak, A.; Dong, H.; Whitham, S.A.; Jander, G. Disruption of Ethylene Responses by Turnip Mosaic Virus Mediates Suppression of Plant Defense against the Green Peach Aphid Vector. Plant Physiol. 2015, 169, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Patton, M.F.; Bak, A.; Sayre, J.M.; Heck, M.L.; Casteel, C.L. A Polerovirus, Potato Leafroll Virus, Alters Plant–Vector Interactions Using Three Viral Proteins. Plant Cell Environ. 2020, 43, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Guzmán-Benito, I.; Donaire, L.; Amorim-Silva, V.; Vallarino, J.G.; Esteban, A.; Wierzbicki, A.T.; Ruiz-Ferrer, V.; Llave, C. The Immune Repressor BIR1 Contributes to Antiviral Defense and Undergoes Transcriptional and Post-Transcriptional Regulation during Viral Infections. New Phytol. 2019, 224, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.K.; Prajapati, R.; Krishna, D.; Divakaran, K.; Pandey, Y.; Reichelt, M.; Mathew, M.K.; Boland, W.; Mithöfer, A.; Vadassery, J. The Ca2+ Channel CNGC19 Regulates Arabidopsis Defense Against Spodoptera Herbivory. Plant Cell 2019, 31, 1539–1562. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.-I.; Ozawa, R.; Maffei, M.E. Recent Advances in Plant Early Signaling in Response to Herbivory. Int. J. Mol. Sci. 2011, 12, 3723–3739. [Google Scholar] [CrossRef] [Green Version]
- Hogenhout, S.A.; Bos, J.I. Effector Proteins That Modulate Plant–Insect Interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Haswell, E.S.; Peyronnet, R.; Barbier-Brygoo, H.; Meyerowitz, E.M.; Frachisse, J.-M. Two MscS Homologs Provide Mechanosensitive Channel Activities in the Arabidopsis Root. Curr. Biol. 2008, 18, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claudel, P.; Chesnais, Q.; Fouché, Q.; Krieger, C.; Halter, D.; Bogaert, F.; Meyer, S.; Boissinot, S.; Hugueney, P.; Ziegler-Graff, V.; et al. The Aphid-Transmitted Turnip Yellows Virus Differentially Affects Volatiles Emission and Subsequent Vector Behavior in Two Brassicaceae Plants. Int. J. Mol. Sci. 2018, 19, 2316. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Transmission ± SD * Using as Source | Plants Inoculated | |||||||
|---|---|---|---|---|---|---|---|---|
| TAIR Locus | Mutant | Mutant | WT # | n | Mutant/WT | p-Value | Seeds from | References |
| Signal transduction | ||||||||
| AT4G33430 | bak1-5 | 45.0 ± 13.9 | 52.0 ± 18.2 | 7 | 138/130 | 0.39 | NASC N799997 | https://doi.org/10.1371/journal.pgen.1002046 (accessed on 4 March 2021) |
| AT3G21630 | lyk1=CERK1 | 36.2 ± 14.8 | 37.1 ± 15.9 | 12 | 221/240 | 0.85 | GABI-KAT 096F09 | https://doi.org/10.1073/pnas.0705147104 (accessed on 4 March 2021) |
| AT3G01840 AT1G51940 AT2G33580 | lyk2 lyk3 lyk5 | 48.3 ± 12.8 | 47.5 ± 12.0 | 11 | 214/217 | 0.85 | Gary Stacey | https://doi.org/10.1104/pp.112.201699 (accessed on 4 March 2021) |
| AT2G23770 | lyk4 | 50.4 ± 9.8 | 54.2 ± 14.7 | 12 | 233/227 | 0.46 | Gary Stacey | https://doi.org/10.1104/pp.112.201699 (accessed on 4 March 2021) |
| AT1G77630 | lyp3 | 32.6 ± 16.3 | 38.6 ± 19.1 | 11 | 216/220 | 0.19 | NASC SALK_132566 | https://doi.org/10.1073/pnas.1112862108 (accessed on 4 March 2021) |
| Calcium signal regulation | ||||||||
| AT4G03560 | tpc1.2 | 53.7 ± 14.2 | 50.2 ± 6.6 | 5 | 188/199 | 0.10 | Saskia Hogenhout | https://doi.org/10.1038/nature03381 (accessed on 4 March 2021) |
| AT4G03560 | fou2 | 46.9 ± 11.5 | 36.6 ± 7.9 | 4 | 96/93 | 0.19 | Edward Farmer | https://doi.org/10.1111/j.1365-313x.2006.03002.x (accessed on 4 March 2021) |
| AT1G53470 AT3G14810 AT1G78610 AT5G19520 AT5G12080 | M5 | 42.7 ± 27.2 | 47.2 ± 29.2 | 14 | 334/330 | 0.28 | Dominique Roby | https://doi.org/10.1016/j.cub.2008.04.039 (accessed on 4 March 2021) |
| AT4G35920 AT2G17780 | mca1 mca2 | 52.9 ± 13.9 | 50.9 ± 14.1 | 17 | 326/329 | 0.76 | Hidetoshi Iida | https://doi.org/10.1104/pp.109.147371 (accessed on 4 March 2021) |
| TAIR Locus ID | Name | CaMV (log2FC) | TuMV (log2FC) |
|---|---|---|---|
| Signal transduction regulator | |||
| AT4G33430 | BAK1 | −0.084 | −0.236 |
| AT5G48380 | BIR1 | 2.764 | −1.145 |
| AT4G32910 | SBB1 | 1.414 | <1 |
| AT5G44585 | PROSCOOP12 | <1 | −1.027 |
| AT2G17120 | LYM2 | −1.099 | <1 |
| AT5G66210 | CPK28 | <1 | −1.204 |
| AT3G21630 | LYK1 | −1.296 | <1 |
| AT2G13790 | SERK4 | <1 | −1.722 |
| AT1G51850 | SIF2 | <1 | −2.394 |
| <1 | <1 | ||
| Calcium signal generation | |||
| AT4G03560 | TPC1 | −0.126 | 0.231 |
| AT1G42540 | GLR3.3 | −0.079 | 0.442 |
| AT3G51480 | GLR3.6 | −0.381 | 0.268 |
| AT2G29110 | GLR2.8 | 1.074 | −2.608 |
| AT2G29120 | GLR2.7 | <1 | −1.136 |
| AT2G32400 | GLR3.7 | −1.616 | <1 |
| AT5G54250 | ATCNGC4/ATDND2/ATHLM1 | 1.048 | <1 |
| AT2G46430 | ATCNGC3 | <1 | −1.025 |
| AT2G24610 | ATCNGC14 | <1 | −1.317 |
| AT1G01340 | ATCNGC10/ATACBK1 | <1 | −1.339 |
| AT3G17690 | ATCNGC19/ATCNBT2 | <1 | −2.266 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Then, C.; Bellegarde, F.; Schivre, G.; Martinière, A.; Macia, J.-L.; Xiong, T.C.; Drucker, M. Plant Viruses Can Alter Aphid-Triggered Calcium Elevations in Infected Leaves. Cells 2021, 10, 3534. https://doi.org/10.3390/cells10123534
Then C, Bellegarde F, Schivre G, Martinière A, Macia J-L, Xiong TC, Drucker M. Plant Viruses Can Alter Aphid-Triggered Calcium Elevations in Infected Leaves. Cells. 2021; 10(12):3534. https://doi.org/10.3390/cells10123534
Chicago/Turabian StyleThen, Christiane, Fanny Bellegarde, Geoffrey Schivre, Alexandre Martinière, Jean-Luc Macia, Tou Cheu Xiong, and Martin Drucker. 2021. "Plant Viruses Can Alter Aphid-Triggered Calcium Elevations in Infected Leaves" Cells 10, no. 12: 3534. https://doi.org/10.3390/cells10123534
APA StyleThen, C., Bellegarde, F., Schivre, G., Martinière, A., Macia, J. -L., Xiong, T. C., & Drucker, M. (2021). Plant Viruses Can Alter Aphid-Triggered Calcium Elevations in Infected Leaves. Cells, 10(12), 3534. https://doi.org/10.3390/cells10123534