
Taming, Domestication and Exaptation: Trajectories of Transposable Elements in Genomes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Short-Term Co-Evolution of Transposable Elements and Genomes: Taming
3. Long Term Co-Evolution of Transposable Elements and Genomes: Domestication and Exaptation
4. How to “Capture” a Transposable Element in a Genomic Position
5. Immobilization of TEs
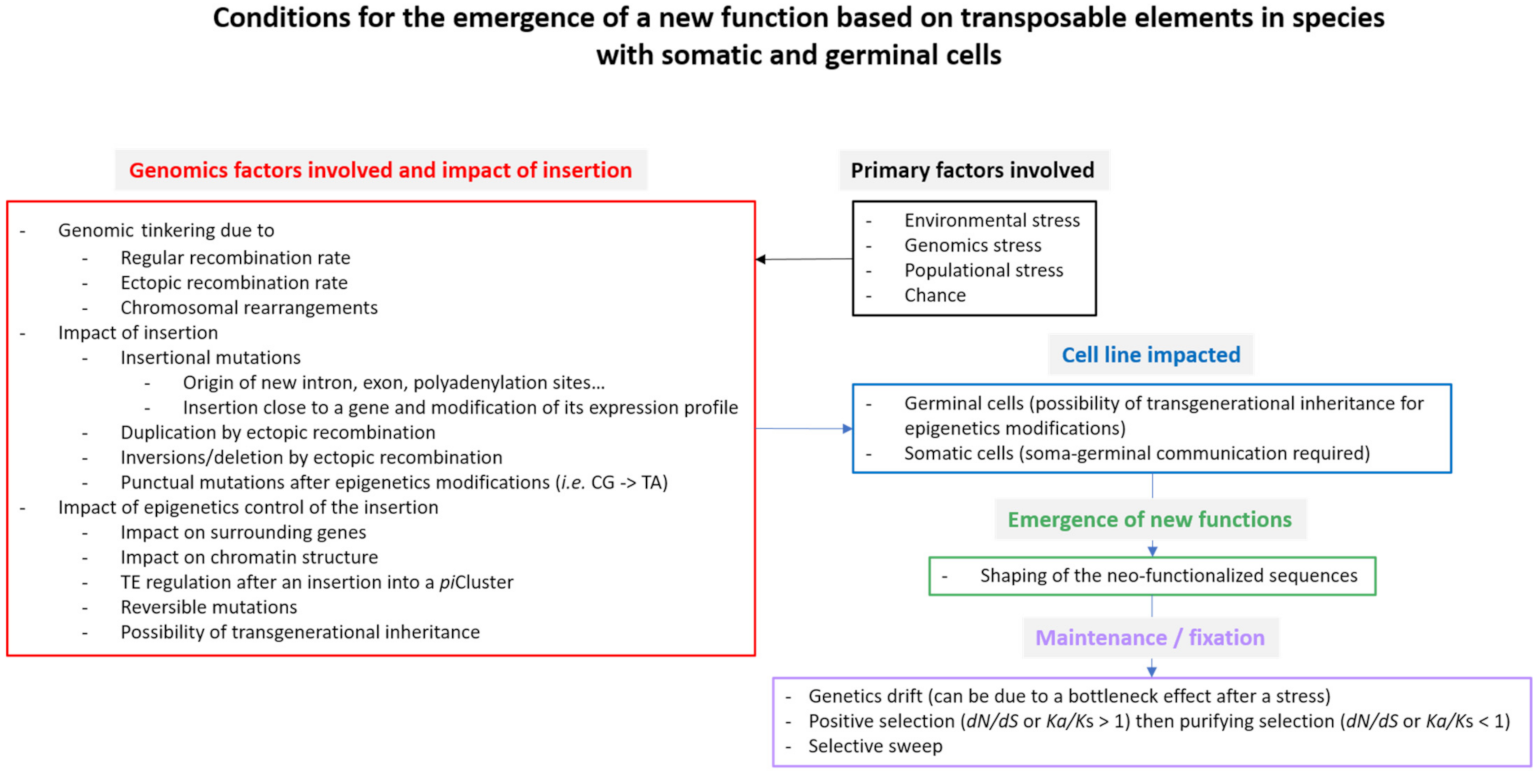
6. Fate of a “Captured” Copy and Emergence of New Functions
7. How to Detect Domesticated or Exapted Copies?
8. Conclusions and Perspectives
9. Remaining Questions
- (i)
- TEs (or viruses) are more frequently domesticated/exapted than any other type of sequence;
- (ii)
- DNA elements seem to be recruited more frequently than other types of TE (retrotransposons, etc.);
- (iii)
Funding
Conflicts of Interest
References
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 1976. [Google Scholar]
- Doolittle Sapienza, C. Selfish genes, the phenotype paradigm and genome evolution. Nature 1980, 284, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Graur, D.; Zheng, Y.; Azevedo, R.B.R. An Evolutionary Classification of Genomic Function. Genome Biol. Evol. 2015, 7, 642–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boissinot, S.; Bourgeois, Y.; Manthey, J.D.; Ruggiero, R.P. The Mobilome of Reptiles: Evolution, Structure, and Function. Cytogenet. Genome Res. 2019, 157, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Carotti, E.; Carducci, F.; Canapa, A.; Barucca, M.; Greco, S.; Gerdol, M.; Biscotti, M.A. Transposable Elements and Teleost Migratory Behaviour. Int. J. Mol. Sci. 2021, 22, 602. [Google Scholar] [CrossRef] [PubMed]
- El Baidouri, M.; Panaud, O. Comparative Genomic Paleontology across Plant Kingdom Reveals the Dynamics of TE-Driven Genome Evolution. Genome Biol. Evol. 2013, 5, 954–965. [Google Scholar] [CrossRef] [Green Version]
- Giraud, D.; Lima, O.; Huteau, V.; Coriton, O.; Boutte, J.; Kovarik, A.; Leitch, A.R.; Leitch, I.J.; Aïnouche, M.; Salmon, A. Evolutionary dynamics of transposable elements and satellite DNAs in polyploid Spartina species. Plant Sci. 2021, 302, 110671. [Google Scholar] [CrossRef]
- Horvath, V.; Merenciano, M.; Gonzalez, J. Revisiting the Relationship between Transposable Elements and the Eukaryotic Stress Response. Trends Genet. 2017, 33, 832–841. [Google Scholar] [CrossRef]
- Oliveira, D.S.; Rosa, M.T.; Vieira, C.; Loreto, E.L.S. Oxidative and radiation stress induces transposable element transcription in Drosophila melanogaster. J. Evol. Biol. 2021, 34, 628–638. [Google Scholar] [CrossRef]
- Gonzalez, J.; Macpherson, J.M.; Petrov, D.A. A Recent Adaptive Transposable Element Insertion Near Highly Conserved Developmental Loci in Drosophila melanogaster. Mol. Biol. Evol. 2009, 26, 1949–1961. [Google Scholar] [CrossRef] [Green Version]
- Merenciano, M.; Iacometti, C.; Gonzalez, J. A unique cluster of roo insertions in the promoter region of a stress response gene in Drosophila melanogaster. Mob. DNA 2019, 10, 10. [Google Scholar] [CrossRef]
- Petrov, D.A.; Fiston-Lavier, A.-S.; Lipatov, M.; Lenkov, K.; Gonzalez, J. Population Genomics of Transposable Elements in Drosophila melanogaster. Mol. Biol. Evol. 2011, 28, 1633–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wos, G.; Choudhury, R.R.; Kolář, F.; Parisod, C. Transcriptional activity of transposable elements along an elevational gradient in Arabidopsis arenosa. Mob. DNA 2021, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.J.; Vrba, E.S. Exaptation-A Missing Term in the Science of Form. Paleobiology 1982, 8, 4–15. [Google Scholar] [CrossRef]
- Jacob, F. Molecular tinkering in evolution. In Evolution from Molecules to Men; Bendall, D.S., Ed.; Cambridge University Press: London, UK, 1983; pp. 131–144. [Google Scholar]
- Schlötterer, C. Genes from scratch—The evolutionary fate of de novo genes. Trends Genet. 2015, 31, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Wissler, L.; Gadau, J.; Simola, D.F.; Helmkampf, M.; Bornberg-Bauer, E. Mechanisms and Dynamics of Orphan Gene Emergence in Insect Genomes. Genome Biol. Evol. 2013, 5, 439–455. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Makarova, K.S.; Aravind, L. Horizontal gene transfer in prokaryotes: Quantification and classification. Annu. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef]
- Crisp, A.; Boschetti, C.; Perry, M.; Tunnacliffe, A.; Micklem, G. Expression of multiple horizontally acquired genes is a hallmark of both vertebrate and invertebrate genomes. Genome Biol. 2015, 16, 50. [Google Scholar] [CrossRef] [Green Version]
- Dotto, B.R.; Carvalho, E.L.; Silva, A.F.; Duarte Silva, L.F.; Pinto, P.M.; Ortiz, M.F.; Wallau, G.L. HTT-DB: Horizontally transferred transposable elements database. Bioinformatics 2015, 31, 2915–2917. [Google Scholar] [CrossRef] [Green Version]
- Loreto, E.L.S.; Carareto, C.M.A.; Capy, P. Revisiting horizontal transfer of transposable elements in Drosophila. Heredity 2008, 100, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Peccoud, J.; Loiseau, V.; Cordaux, R.; Gilbert, C. Massive horizontal transfer of transposable elements in insects. Proc. Natl. Acad. Sci. USA 2017, 114, 4721–4726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallau, G.L.; Capy, P.; Loreto, E.; Le Rouzic, A.; Hua-Van, A. VHICA, a New Method to Discriminate between Vertical and Horizontal Transposon Transfer: Application to the Mariner Family within Drosophila. Mol. Biol. Evol. 2016, 33, 1094–1109. [Google Scholar] [CrossRef] [Green Version]
- Barker, M.; Arrigo, N.; Baniaga, A.; Li, Z.; Levin, D. On the relative abundance of autopolyploids and allopolyploids. New Phytol. 2016, 210, 391–398. [Google Scholar] [CrossRef]
- Doolittle, W.F. Phylogenetic classification and the universal tree. Science 1999, 284, 2124–2129. [Google Scholar] [CrossRef] [PubMed]
- Cordaux, R.; Gilbert, C. Evolutionary Significance of Wolbachia-to-Animal Horizontal Gene Transfer: Female Sex Determination and the f Element in the Isopod Armadillidium vulgare. Genes 2017, 8, 186. [Google Scholar] [CrossRef]
- Schoenknecht, G.; Chen, W.-H.; Ternes, C.M.; Barbier, G.G.; Shrestha, R.P.; Stanke, M.; Braeutigam, A.; Baker, B.J.; Banfield, J.F.; Garavito, R.M.; et al. Gene Transfer from Bacteria and Archaea Facilitated Evolution of an Extremophilic Eukaryote. Science 2013, 339, 1207–1210. [Google Scholar] [CrossRef]
- Zimorski, V.; Ku, C.; Martin, W.F.; Gould, S.B. Endosymbiotic theory for organelle origins. Curr. Opin. Microbiol. 2014, 22, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Escudero, J.A.; Loot, C.; Nivina, A.; Mazel, D. The Integron: Adaptation On Demand. Microbiol. Spectr. 2015, 3, MDNA3-0019-2014. [Google Scholar] [CrossRef] [Green Version]
- San Millan, A.; Escudero, J.A.; Gifford, D.R.; Mazel, D.; MacLean, R.C. Multicopy plasmids potentiate the evolution of antibiotic resistance in bacteria. Nat. Ecol. Evol. 2017, 1, 0010. [Google Scholar] [CrossRef] [PubMed]
- García-Aljaro, C.; Ballesté, E.; Muniesa, M. Beyond the canonical strategies of horizontal gene transfer in prokaryotes. Curr. Opin. Microbiol. 2017, 38, 95–105. [Google Scholar] [CrossRef]
- von Wintersdorff, C.J.H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Ren, X.; Mason, A.S.; Liu, H.; Xiao, M.; Li, J.; Fu, D. Horizontal gene transfer in plants. Funct. Integr. Genomics 2014, 14, 23–29. [Google Scholar] [CrossRef]
- Gilbert, C.; Cordaux, R. Viruses as vectors of horizontal transfer of genetic material in eukaryotes. Curr. Opin. Virol. 2017, 25, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-H.; Jean Peccoud Xu, M.-R.-X.; Zhang, X.-G.; Gilbert, C. Horizontal transfer and evolution of transposable elements in vertebrates. Nat. Commun. 2020, 11, 1362. [Google Scholar] [CrossRef] [Green Version]
- Le Rouzic, A.; Capy, P. The first steps of transposable elements invasion: Parasitic strategy vs. genetic drift. Genetics 2005, 169, 1033–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.; Caizzi, R.; Viggiano, L.; Marsano, R.M. Does the Promoter Constitute a Barrier in the Horizontal Transposon Transfer Process? Insight from Bari Transposons. Genome Biol. Evol. 2017, 9, 1637–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.; Lorusso, P.; Miskey, C.; Walisko, O.; Gerbino, A.; Marobbio, C.M.T.; Ivics, Z.; Marsano, R.M. Transcriptionally promiscuous “blurry” promoters in Tc1/mariner transposons allow transcription in distantly related genomes. Mob. DNA 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokta, A.J. Elements of Physical Biology; Williams & Wilkins: Baltimore, MD, USA, 1925. [Google Scholar]
- Volterra, V. Fluctuations in the Abundance of a Species considered Mathematically. Nature 1926, 118, 558–560. [Google Scholar] [CrossRef]
- Le Rouzic, A.; Boutin, T.S.; Capy, P. Long-term evolution of transposable elements. Proc. Natl. Acad. Sci. USA 2007, 104, 19375–19380. [Google Scholar] [CrossRef] [Green Version]
- Robillard, É.; Le Rouzic, A.; Zhang, Z.; Capy, P.; Hua-Van, A. Experimental evolution reveals hyperparasitic interactions among transposable elements. Proc. Natl. Acad. Sci. USA 2016, 113, 14763–14768. [Google Scholar] [CrossRef] [Green Version]
- Bouyer, D.; Kramdi, A.; Kassam, M.; Heese, M.; Schnittger, A.; Roudier, F.; Colot, V. DNA methylation dynamics during early plant life. Genome Biol. 2017, 18, 179. [Google Scholar] [CrossRef]
- Brennecke, J.; Malone, C.D.; Aravin, A.A.; Sachidanandam, R.; Stark, A.; Hannon, G.J. An Epigenetic Role for Maternally Inherited piRNAs in Transposon Silencing. Science 2008, 322, 1387–1392. [Google Scholar] [CrossRef] [Green Version]
- Cappucci, U.; Noro, F.; Casale, A.M.; Fanti, L.; Berloco, M.; Alagia, A.A.; Grassi, L.; Le Pera, L.; Piacentini, L.; Pimpinelli, S. The Hsp70 chaperone is a major player in stress-induced transposable element activation. Proc. Natl. Acad. Sci. USA 2019, 116, 17943–17950. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, J.; Qi, H.; Liu, N.; Lin, H. Piwi Is a Key Regulator of Both Somatic and Germline Stem Cells in the Drosophila Testis. Cell Rep. 2015, 12, 150–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Leandre, B.; Capy, P.; Hua-Van, A.; Filee, J. piRNA and Transposon Dynamics in Drosophila: A Female Story. Genome Biol. Evol. 2020, 12, 931–947. [Google Scholar] [CrossRef] [PubMed]
- Saint-Leandre, B.; Clavereau, I.; Hua-Van, A.; Capy, P. Transcriptional polymorphism of pi RNA regulatory genes underlies the mariner activity in Drosophila simulans testes. Mol. Ecol. 2017, 26, 3715–3731. [Google Scholar] [CrossRef]
- Lewis, S.H.; Quarles, K.A.; Yang, Y.; Tanguy, M.; Frézal, L.; Smith, S.A.; Sharma, P.P.; Cordaux, R.; Gilbert, C.; Giraud, I.; et al. Pan-arthropod analysis reveals somatic piRNAs as an ancestral defence against transposable elements. Nat. Ecol. Evol. 2018, 2, 174–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCue, A.D.; Nuthikattu, S.; Reeder, S.H.; Slotkin, R.K. Gene Expression and Stress Response Mediated by the Epigenetic Regulation of a Transposable Element Small RNA. PLoS Genet. 2012, 8, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Qu, Z.; Yang, L.; Zhang, Q.; Liu, Z.-H.; Do, T.; Adelson, D.L.; Wang, Z.-Y.; Searle, I.; Zhu, J.-K. Transposable elements (TEs) contribute to stress-related long intergenic noncoding RNAs in plants. Plant J. 2017, 90, 133–146. [Google Scholar] [CrossRef] [Green Version]
- Josse, T.; Boivin, A.; Anxolabehere, D.; Ronsseray, S. P element-encoded regulatory products enhance Repeat-Induced Gene Silencing (RIGS) of P-lacZ-white clusters in Drosophila melanogaster. Mol. Genet. Genom. 2002, 268, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.; Marconi, S.; Specchia, V.; Bozzetti, M.P.; Ivics, Z.; Caizzi, R.; Marsano, R.M. Functional Characterization of the Bari1 Transposition System. PLoS ONE 2013, 8, e79385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirochika, H.; Okamoto, H.; Kakutani, T. Silencing of Retrotransposons in Arabidopsis and Reactivation by the ddm1 Mutation. Plant Cell 2000, 12, 357–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukahara, S.; Kobayashi, A.; Kawabe, A.; Mathieu, O.; Miura, A.; Kakutani, T. Bursts of retrotransposition reproduced in Arabidopsis. Nature 2009, 461, 423–426. [Google Scholar] [CrossRef]
- Choi, J.Y.; Lee, Y.C.G. Double-edged sword: The evolutionary consequences of the epigenetic silencing of transposable elements. PLoS Genet. 2020, 16, e1008872. [Google Scholar] [CrossRef]
- Hollister, J.D.; Gaut, B.S. Epigenetic silencing of transposable elements: A trade-off between reduced transposition and deleterious effects on neighboring gene expression. Genome Res. 2009, 19, 1419–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippman, Z.; Gendrel, A.V.; Black, M.; Vaughn, M.W.; Dedhia, N.; McCombie, W.R.; Lavine, K.; Mittal, V.; May, B.; Kasschau, K.D.; et al. Role of transposable elements in heterochromatin and epigenetic control. Nature 2004, 430, 471–476. [Google Scholar] [CrossRef]
- Quadrana, L.; Silveira, A.B.; Mayhew, G.F.; LeBlanc, C.; Martienssen, R.A.; Jeddeloh, J.A.; Colot, V. The Arabidopsis thaliana mobilome and its impact at the species level. eLife 2016, 5, e15716. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.I.; Agrawal, N.; Bureau, T.E. Effects of Recombination Rate and Gene Density on Transposable Element Distributions in Arabidopsis thaliana. Genome Res. 2003, 13, 1897–1903. [Google Scholar] [CrossRef] [PubMed]
- Grover, D.; Majumder, P.P.; Rao, C.B.; Brahmachari, S.K.; Mukerji, M. Nonrandom distribution of Alu elements in genes of various functional categories: Insight from analysis of human chromosomes 21 and 22. Mol. Biol. Evol. 2003, 20, 1420–1424. [Google Scholar] [CrossRef] [Green Version]
- Sultana, T.; van Essen, D.; Siol, O.; Bailly-Bechet, M.; Philippe, C.; Zine El Aabidine, A.; Pioger, L.; Nigumann, P.; Saccani, S.; Andrau, J.-C.; et al. The Landscape of L1 Retrotransposons in the Human Genome Is Shaped by Pre-insertion Sequence Biases and Post-insertion Selection. Mol. Cell 2019, 74, 555–570.e7. [Google Scholar] [CrossRef] [PubMed]
- Cridland, J.M.; Macdonald, S.J.; Long, A.D.; Thornton, K.R. Abundance and Distribution of Transposable Elements in Two Drosophila QTL Mapping Resources. Mol. Biol. Evol. 2013, 30, 2311–2327. [Google Scholar] [CrossRef] [Green Version]
- Favarato, R.M.; Ribeiro, L.B.; Feldberg, E.; Matoso, D.A. Chromosomal mapping of transposable elements of the Rex family in the bristlenose catfish, Ancistrus (Siluriformes, Loricariidae), from the Amazonian region. J. Hered. 2017, 108, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Bertocchi, N.A.; de Oliveira, T.D.; del Garnero, A.V.; Buogo Coan, R.L.; Gunski, R.J.; Martins, C.; Torres, F.P. Distribution of CR1-like transposable element in woodpeckers (Aves Piciformes): Z sex chromosomes can act as a refuge for transposable elements. Chromosome Res. 2018, 26, 333–343. [Google Scholar] [CrossRef]
- Charlesworth, B.; Langley, C.H. The population genetics of Drosophila transposable elements. Annu. Rev. Genet. 1989, 23, 251–287. [Google Scholar] [CrossRef] [PubMed]
- Langley, C.H.; Montgomery, E.; Hudson, R.; Kaplan, N.; Charlesworth, B. On the role of unequal exchange in the containment of transposable element copy number. Genet. Res. 1988, 52, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Fontanillas, P.; Hartl, D.L.; Reuter, M. Genome Organization and Gene Expression Shape the Transposable Element Distribution in the Drosophila melanogaster Euchromatin. PLoS Genet. 2007, 3, e210. [Google Scholar] [CrossRef]
- Sultana, T.; Zamborlini, A.; Cristofari, G.; Lesage, P. Integration site selection by retroviruses and transposable elements in eukaryotes. Nat. Rev. Genet. 2017, 18, 292–308. [Google Scholar] [CrossRef] [PubMed]
- Auvinet, J.; Graça, P.; Ghigliotti, L.; Pisano, E.; Dettaï, A.; Ozouf-Costaz, C.; Higuet, D. Insertion Hot Spots of DIRS1 Retrotransposon and Chromosomal Diversifications among the Antarctic Teleosts Nototheniidae. Int. J. Mol. Sci. 2019, 20, 701. [Google Scholar] [CrossRef] [Green Version]
- Biemont, C.; Vieira, C. Genetics—Junk DNA as an evolutionary force. Nature 2006, 443, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Mazzuchelli, J.; Martins, C. Genomic organization of repetitive DNAs in the cichlid fish Astronotus ocellatus. Genetica 2009, 136, 461–469. [Google Scholar] [CrossRef]
- Pardue, M.L.; DeBaryshe, P.G. Drosophila telomeres: Two transposable elements with important roles in chromosomes. Genetica 1999, 107, 189–196. [Google Scholar] [CrossRef]
- Pimpinelli, S.; Berloco, M.; Fanti, L.; Dimitri, P.; Bonaccorsi, S.; Marchetti, E.; Caizzi, R.; Caggese, C.; Gatti, M. Transposable elements are stable structural components of Drosophila melanogaster heterochromatin. Proc. Natl. Acad. Sci. USA 1995, 92, 3804–3808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganko, E.W.; Greene, C.S.; Lewis, J.A.; Bhattacharjee, V.; McDonald, J.F. LTR retrotransposon-gene associations in Drosophila melanogaster. J. Mol. Evol. 2006, 62, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Bogwitz, M.R.; McCart, C.; Andrianopoulos, A.; Ffrench-Constant, R.H.; Batterham, P.; Daborn, P.J. Cis-regulatory elements in the Accord retrotransposon result in tissue-specific expression of the Drosophila melanogaster insecticide resistance gene Cyp6g1. Genetics 2007, 175, 1071–1077. [Google Scholar] [CrossRef] [Green Version]
- Daborn, P.J.; Yen, J.L.; Bogwitz, M.R.; Le Goff, G.; Feil, E.; Jeffers, S.; Tijet, N.; Perry, T.; Heckel, D.; Batterham, P.; et al. A single P450 allele associated with insecticide resistance in Drosophila. Science 2002, 297, 2253–2256. [Google Scholar] [CrossRef] [PubMed]
- Mateo, L.; Ullastres, A.; González, J. A Transposable Element Insertion Confers Xenobiotic Resistance in Drosophila. PLoS Genet. 2014, 10, e1004560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villanueva-Cañas, J.L.; Horvath, V.; Aguilera, L.; González, J. Diverse families of transposable elements affect the transcriptional regulation of stress-response genes in Drosophila melanogaster. Nucleic Acids Res. 2019, 47, 6842–6857. [Google Scholar] [CrossRef] [Green Version]
- DeBarry, J.D.; Ganko, E.W.; McCarthy, E.M.; McDonald, J.F. The contribution of LTR retrotransposon sequences to gene evolution in Mus musculus. Mol. Biol. Evol. 2006, 23, 479–481. [Google Scholar] [CrossRef] [Green Version]
- Wicker, T.; Gundlach, H.; Spannagl, M.; Uauy, C.; Borrill, P.; Ramirez-Gonzalez, R.H.; De Oliveira, R.; Mayer, K.F.X.; Paux, E.; Choulet, F. Impact of transposable elements on genome structure and evolution in bread wheat. Genome Biol. 2018, 19, 103. [Google Scholar] [CrossRef] [PubMed]
- SanMiguel, P.; Tikhonov, A.; Jin, Y.-K.; Motchoulskaia, N.; Zakharov, D.; Melake-Berhan, A.; Springer, P.S.; Edwards, K.J.; Lee, M.; Avramova, Z.; et al. Nested Retrotransposons in the Intergenic Regions of the Maize Genome. Science 1996, 274, 765–768. [Google Scholar] [CrossRef] [Green Version]
- SanMiguel, P.; Gaut, B.S.; Tikhonov, A.; Nakajima, Y.; Bennetzen, J.L. The paleontology of intergene retrotransposons of maize. Nat. Genet. 1998, 20, 43–45. [Google Scholar] [CrossRef]
- Kofler, R. Dynamics of Transposable Element Invasions with piRNA Clusters. Mol. Biol. Evol. 2019, 36, 1457–1472. [Google Scholar] [CrossRef] [Green Version]
- Kofler, R. piRNA Clusters Need a Minimum Size to Control Transposable Element Invasions. Genome Biol. Evol. 2020, 12, 736–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mevel-Ninio, M.; Pelisson, A.; Kinder, J.; Campos, A.R.; Bucheton, A. The flamenco locus controls the gypsy and ZAM retroviruses and is required for Drosophila oogenesis. Genetics 2007, 175, 1615–1624. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Pointer, B.; Kelleher, E.S. Rapid evolution of piRNA-mediated silencing of an invading transposable element was driven by abundant de novo mutations. Genome Res. 2020, 30, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Chuong, E.B.; Elde, N.C.; Feschotte, C. Regulatory activities of transposable elements: From conflicts to benefits. Nat. Rev. Genet. 2017, 18, 71–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cost, G.J.; Golding, A.; Schlissel, M.S.; Boeke, J.D. Target DNA chromatinization modulates nicking by L1 endonuclease. Nucleic Acids Res. 2001, 29, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Levin, H.L.; Moran, J.V. Dynamic interactions between transposable elements and their hosts. Nat. Rev. Genet. 2011, 12, 615–627. [Google Scholar] [CrossRef]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [Green Version]
- Genzor, P.; Konstantinidou, P.; Stoyko, D.; Manzourolajdad, A.; Andrews, C.M.; Elchert, A.R.; Stathopoulos, C.; Haase, A.D. Cellular abundance shapes function in piRNA-guided genome defense 2 (Running title: The functional piRNA sequence space). BioRxiv 2021, 31, 2058–2068. [Google Scholar]
- Kjellman, C.; Sjogren, H.; Widegren, B. The Y-Chromosome—A Graveyard For Endogenous Retroviruses. Gene 1995, 161, 163–170. [Google Scholar] [CrossRef]
- Sadeq, S.; Al-Hashimi, S.; Cusack, C.M.; Werner, A. Endogenous Double-Stranded RNA. Non-Coding RNA 2021, 7, 15. [Google Scholar] [CrossRef]
- Bhattacharyya, M.; Smith, A.; Ellis, T.; Hedley, C.; Martin, C. The wrinkled-seed character of pea described by Mendel is caused by a transposon-like insertion in a gene coding starch-branching enzyme. Cell 1990, 60, 115–122. [Google Scholar] [CrossRef]
- van’t Hof, A.E.; Campagne, P.; Rigden, D.J.; Yung, C.J.; Lingley, J.; Quail, M.A.; Hall, N.; Darby, A.C.; Saccheri, I.J. The industrial melanism mutation in British peppered moths is a transposable element. Nature 2016, 534, 102–105. [Google Scholar]
- Bogwitz, M.R.; Chung, H.; Magoc, L.; Rigby, S.; Wong, W.; O’Keefe, M.; McKenzie, J.A.; Batterham, P.; Daborn, P.J. Cyp12a4 confers lufenuron resistance in a natural population of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2005, 102, 12807–12812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsano, R.M.; Caizzi, R.; Moschetti, R.; Junakovic, N. Evidence for a functional interaction between the Bari1 transposable element and the cytochrome P450 cyp12a4 gene in Drosophila melanogaster. Gene 2005, 357, 122–128. [Google Scholar] [CrossRef]
- Barron, M.G.; Fiston-Lavier, A.-S.; Petrov, D.M.; Gonzalez, J. Population Genomics of Transposable Ele-ments in Drosophila. Annu. Rev. Genet. 2014, 48, 561–581. [Google Scholar] [CrossRef] [Green Version]
- Garavís, M.; González, C.; Villasante, A. On the Origin of the Eukaryotic Chromosome: The Role of Noncanonical DNA Structures in Telomere Evolution. Genome Biol. Evol. 2013, 5, 1142–1150. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Krupovic, M. Evolution of adaptive immunity from transposable elements combined with innate immune systems. Nat. Rev. Genet. 2015, 16, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Lavialle, C.; Cornelis, G.; Dupressoir, A.; Esnault, C.; Heidmann, O.; Vernochet, C.; Heidmann, T. Paleovirology of ‘syncytins’, retroviral env genes exapted for a role in placentation. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120507. [Google Scholar] [CrossRef] [Green Version]
- Etchegaray, E.; Naville, M.; Volff, J.-N.; Haftek-Terreau, Z. Transposable element-derived sequences in vertebrate development. Mob. DNA 2021, 12, 1. [Google Scholar] [CrossRef]
- Joly-Lopez, Z.; Bureau, T.E. Exaptation of transposable element coding sequences. Curr. Opin. Genet. Dev. 2018, 49, 34–42. [Google Scholar] [CrossRef]
- Danilevskaya, O.N.; Lofsky, A.; Pardue, M.L. Het-A a transposable element with an apparent role in the telomeres of Drosophila. Molec. Biol. Cell 1992, 3, 83A. [Google Scholar]
- George, J.A.; DeBaryshe, P.G.; Traverse, K.L.; Celniker, S.E.; Pardue, M.-L. Genomic organization of the Drosophila telomere retrotransposable elements. Genome Res. 2006, 16, 1231–1240. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Vanguri, S.; Boeke, J.D.; Gabriel, A.; Voytas, D.F. Transposable elements and genome organization: A comprehensive survey of retrotransposons revealed by the complete Saccharomyces cerevisiae genome sequence. Genome Res. 1998, 8, 464–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
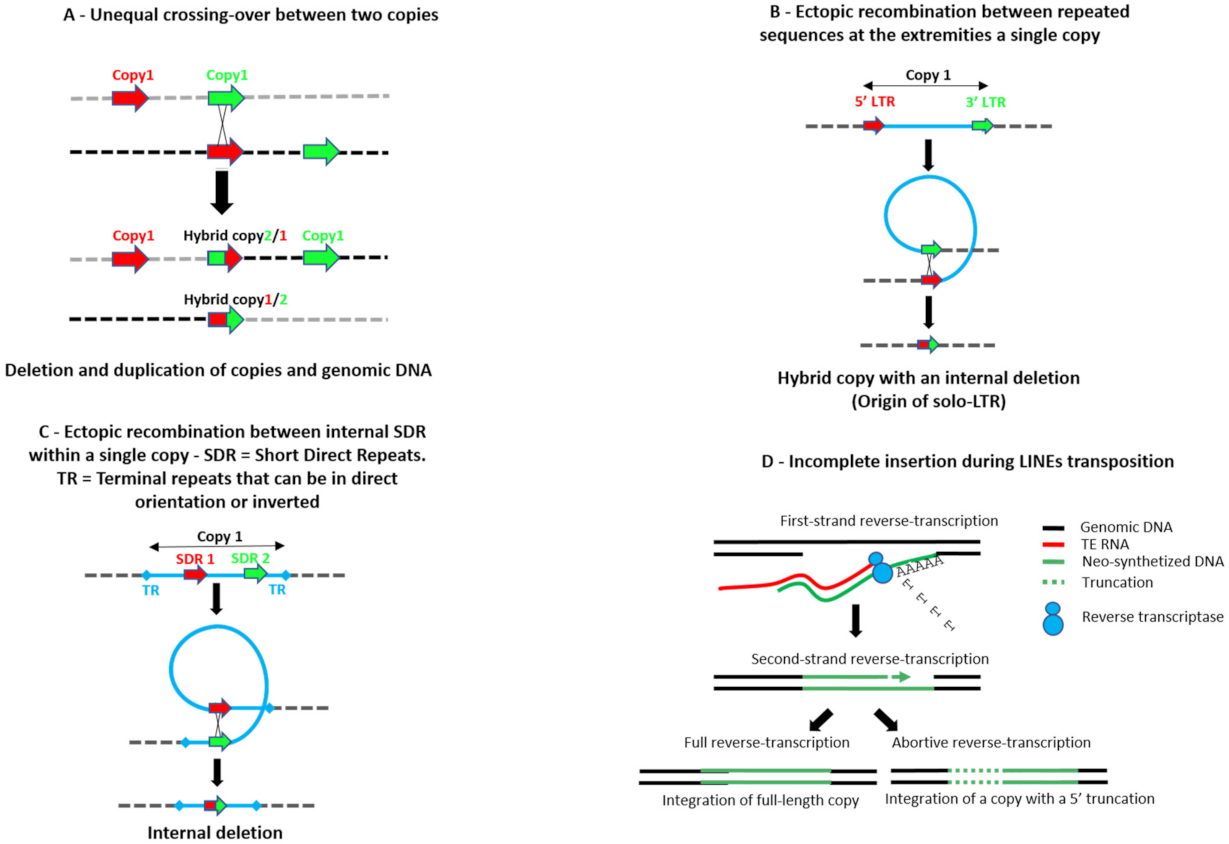
- Vitte, C.; Panaud, O. Formation of solo-LTRs through unequal homologous recombination counterbalances amplifications of LTR retrotransposons in rice Oryza sativa L. Mol. Biol. Evol. 2003, 20, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; DeWoody, J.A. Genomic Landscape of Long Terminal Repeat Retrotransposons (LTR-RTs) and Solo LTRs as Shaped by Ectopic Recombination in Chicken and Zebra Finch. J. Mol. Evol. 2016, 82, 251–263. [Google Scholar] [CrossRef]
- Merenciano, M.; Ullastres, A.; de Cara, M.A.R.; Barrón, M.G.; González, J. Multiple Independent Retroelement Insertions in the Promoter of a Stress Response Gene Have Variable Molecular and Functional Effects in Drosophila. PLoS Genet. 2016, 12, e1006249. [Google Scholar] [CrossRef]
- Luan, D.D.; Korman, M.H.; Jakubczak, J.L.; Eickbush, T.H. Reverse transcription of R2Bm RNA is primed by a nick at the chromosomal traget site: A mechanism for non-LTR retrotransposition. Cell 1993, 72, 595–605. [Google Scholar] [CrossRef]
- Petrov, D.A.; Lozovskaya, E.R.; Hartl, D.L. High intrinsic: Rate of DNA loss in Drosophila. Nature 1996, 384, 346–349. [Google Scholar] [CrossRef]
- Brunet, F.; Giraud, T.; Godin, F.; Capy, P. Do deletions of the Mos1-like elements occur randomly in the Drosophilidae family? J. Mol. Evol. 2002, 54, 227–234. [Google Scholar] [CrossRef]
- Negoua, A.; Rouault, J.-D.; Chakir, M.; Capy, P. Internal deletions of transposable elements: The case of Lemi elements. Genetica 2013, 141, 369–379. [Google Scholar] [CrossRef] [PubMed]
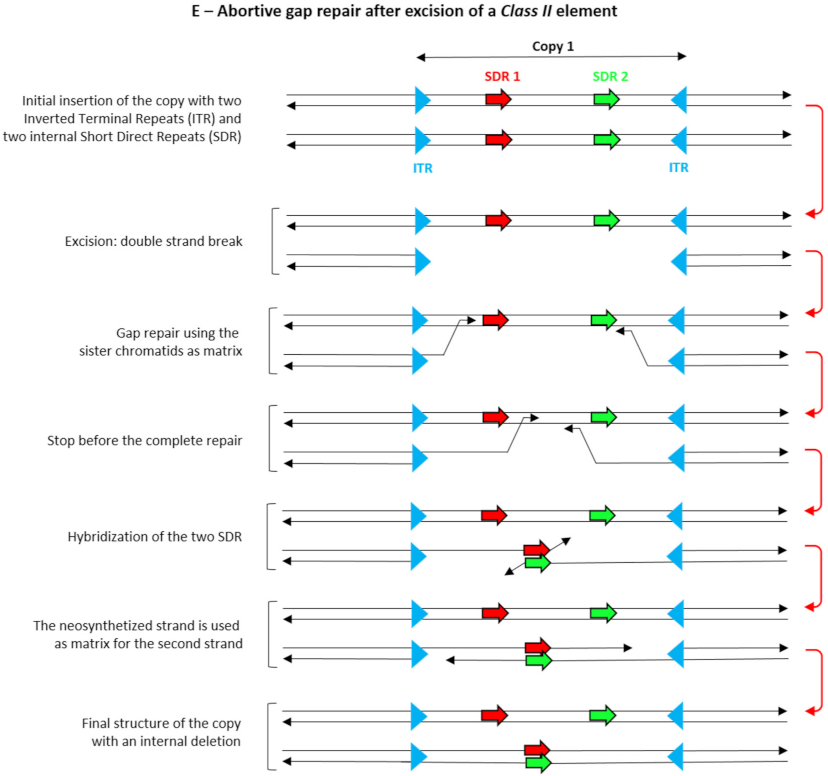
- Rubin, E.; Levy, A.A. Abortive gap repair: Underlying mechanism for Ds element formation. Mol. Cell. Biol. 1997, 17, 6294–6302. [Google Scholar] [CrossRef] [Green Version]
- McGurk, M.P.; Dion-Cote, A.-M.; Barbash, D.A. Rapid evolution at the Drosophila telomere: Transposable element dynamics at an intrinsically unstable locus. Genetics 2021, 217, iyaa027. [Google Scholar] [CrossRef] [PubMed]
- Pardue, M.-L.; DeBaryshe, P.G. Retrotransposons that maintain chromosome ends. Proc. Natl. Acad. Sci. USA 2011, 108, 20317–20324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzohairy, A.M.; Gyulai, G.; Jansen, R.K.; Bahieldin, A. Transposable elements domesticated and neofunctionalized by eukaryotic genomes. Plasmid 2013, 69, 1–15. [Google Scholar] [CrossRef]
- Jangam, D.; Feschotte, C.; Betran, E. Transposable Element Domestication As an Adaptation to Evolutionary Conflicts. Trends Genet. 2017, 33, 817–831. [Google Scholar] [CrossRef]
- Volff, J.-N. Turning junk into gold: Domestication of transposable elements and the creation of new genes in eukaryotes. Bioessays 2006, 28, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Schartl, M.; Schories, S.; Wakamatsu, Y.; Nagao, Y.; Hashimoto, H.; Bertin, C.; Mourot, B.; Schmidt, C.; Wilhelm, D.; Centanin, L.; et al. Sox5 is involved in germ-cell regulation and sex determination in medaka following co-option of nested transposable elements. BMC Biol. 2018, 16, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tipney, H.J.; Hinsley, T.A.; Brass, A.; Metcalfe, K.; Donnai, D.; Tassabehji, M. Isolation and characterisation of GTF2IRD2, a novel fusion gene and member of the TFII-I family of transcription factors, deleted in Williams–Beuren syndrome. Eur. J. Hum. Genet. 2004, 12, 551–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, J.C.; Bailey, A.D.; Fan, H.-Y.; Pavelitz, T.; Weiner, A.M. An Abundant Evolutionarily Conserved CSB-PiggyBac Fusion Protein Expressed in Cockayne Syndrome. PLoS Genet. 2008, 4, e1000031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baudry, C.; Malinsky, S.; Restituito, M.; Kapusta, A.; Rosa, S.; Meyer, E.; Betermier, M. PiggyMac, a domesticated piggyBac transposase involved in programmed genome rearrangements in the ciliate Paramecium tetraurelia. Genes Dev. 2009, 23, 2478–2483. [Google Scholar] [CrossRef] [Green Version]
- Bischerour, J.; Bhullar, S.; Denby Wilkes, C.; Régnier, V.; Mathy, N.; Dubois, E.; Singh, A.; Swart, E.; Arnaiz, O.; Sperling, L.; et al. Six domesticated PiggyBac transposases together carry out programmed DNA elimination in Paramecium. eLife 2018, 7, e37927. [Google Scholar] [CrossRef] [PubMed]
- Yellan, I.; Yang, A.W.H.; Hughes, T.R. Diverse Eukaryotic CGG-Binding Proteins Produced by Independent Domestications of hAT Transposons. Mol. Biol. Evol. 2021, 38, 2070–2075. [Google Scholar] [CrossRef] [PubMed]
- Cosby, R.L.; Judd, J.; Zhang, R.; Zhong, A.; Garry, N.; Pritham, E.J.; Feschotte, C. Recurrent evolution of vertebrate transcription factors by transposase capture. Science 2021, 371, eabc6405. [Google Scholar] [CrossRef] [PubMed]
- Velanis, C.N.; Perera, P.; Thomson, B.; de Leau, E.; Liang, S.C.; Hartwig, B.; Förderer, A.; Thornton, H.; Arede, P.; Chen, J.; et al. The domesticated transposase ALP2 mediates formation of a novel Polycomb protein complex by direct interaction with MSI1, a core subunit of Polycomb Repressive Complex 2 (PRC2). PLoS Genet. 2020, 16, e1008681. [Google Scholar] [CrossRef]
- Abascal, F.; Tress, M.L.; Valencia, A. Alternative splicing and co-option of transposable elements: The case of TMPO/LAP2α and ZNF451 in mammals. Bioinformatics 2015, 31, 2257–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sela, N.; Kim, E.; Ast, G. The role of transposable elements in the evolution of non-mammalian vertebrates and invertebrates. Genome Biol. 2010, 11, R59. [Google Scholar] [CrossRef]
- Zhang, X.H.-F.; Chasin, L.A. Comparison of multiple vertebrate genomes reveals the birth and evolution of human exons. Proc. Natl. Acad. Sci. USA 2006, 103, 13427–13432. [Google Scholar] [CrossRef] [Green Version]
- McGurk, M.P.; Barbash, D.A. Double insertion of transposable elements provides a substrate for the evolution of satellite DNA. Genome Res. 2018, 28, 714–725. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Barbash, D.A.; Kelleher, E.S. Adaptive evolution among cytoplasmic piRNA proteins leads to decreased genomic auto-immunity. PLoS Genet. 2020, 16, e1008861. [Google Scholar] [CrossRef]
- Eaton, S.A.; Jayasooriah, N.; Buckland, M.E.; Martin, D.I.; Cropley, J.E.; Suter, C.M. Roll over Weismann: Extracellular vesicles in the transgenerational transmission of environmental effects. Epigenomics 2015, 7, 1165–1171. [Google Scholar] [CrossRef] [Green Version]
- Okabe, E.; Uno, M.; Kishimoto, S.; Nishida, E. Intertissue small RNA communication mediates the acquisition and inheritance of hormesis in Caenorhabditis elegans. Commun. Biol. 2021, 4, 207. [Google Scholar] [CrossRef]
- Shapiro, J.; Noble, D. What prevents mainstream evolutionists teaching the whole truth about how genomes evolve? Prog. Biophys. Mol. Biol. 2021, 165, 140–152. [Google Scholar] [CrossRef]
- Ashe, A.; Colot, V.; Oldroyd, B.P. How does epigenetics influence the course of evolution? Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200111. [Google Scholar] [CrossRef]
- Mendizabal, I.; Keller, T.E.; Zeng, J.; Yi, S.V. Epigenetics and Evolution. Integr. Comp. Biol. 2014, 54, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, S.V.; Goodisman, M.A.D. The impact of epigenetic information on genome evolution. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200114. [Google Scholar] [CrossRef] [PubMed]
- Zamudio, N.; Bourc’his, D. Transposable elements in the mammalian germline: A comfortable niche or a deadly trap? Heredity 2010, 105, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Barau, J.; Teissandier, A.; Zamudio, N.; Roy, S.; Nalesso, V.; Herault, Y.; Guillou, F.; Bourc’his, D. The DNA methyltransferase DNMT3C protects male germ cells from transposon activity. Science 2016, 354, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.T.; Bonneau, A.R.; Giraldez, A.J. Zygotic Genome Activation During the Maternal-to-Zygotic Transition. Annu. Rev. Cell Dev. Biol. 2014, 30, 581–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landschaft, D. Gaps and barriers: Gap junctions as a channel of communication between the soma and the germline. Semin. Cell Dev. Biol. 2020, 97, 167–171. [Google Scholar] [CrossRef]
- Brasset, E.; Taddei, A.; Arnaud, F.; Faye, B.; Fausto, A.; Mazzini, M.; Giorgi, F.; Vaury, C. Viral particles of the endogenous retrovirus ZAM from Drosophila melanogaster use a pre-existing endosome/exosome pathway for transfer to the oocyte. Retrovirology 2006, 3, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalvet, F.; Teysset, L.; Terzian, C.; Prud’homme, N.; Santamaria, P.; Bucheton, A.; Pélisson, A. Proviral amplification of the Gypsy endogenous retrovirus of Drosophila melanogaster involves env-independent invasion of the female germline. EMBO J. 1999, 18, 2659–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touret, F.; Guiguen, F.; Terzian, C. Un rétrovirus endogène actif et infectieux: Le cas gypsy chez Drosophila melanogaster. Virologie 2012, 16, 9. [Google Scholar]
- Anxolabéhère, D.; Kidwell, M.G.; Periquet, G. Molecular characteristics of diverse populations are consistent with the hypothesis of a recent invasion of Drosophila melanogaster by mobile P elements. Mol. Biol. Evol. 1988, 5, 252–269. [Google Scholar] [PubMed] [Green Version]
- Daniels, S.B.; Peterson, K.R.; Strausbaugh, L.D.; Kidwell, M.G.; Chovnick, A. Evidence for horizontal transmission of the P transposable element between Drosophila species. Genetics 1990, 124, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Hermant, C.; Boivin, A.; Teysset, L.; Delmarre, V.; Asif-Laidin, A.; van den Beek, M.; Antoniewski, C.; Ronsseray, S. Paramutation in Drosophila Requires Both Nuclear and Cytoplasmic Actors of the piRNA Pathway and Induces Cis-spreading of piRNA Production. Genetics 2015, 201, 1381–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaitre, B.; Ronsseray, S.; Coen, D. Maternal repression of the P- element promoter in the germline of Drosophila melanogaster—A model for the P-cytotype. Genetics 1993, 135, 149–160. [Google Scholar] [CrossRef]
- Saito, K. The epigenetic regulation of transposable elements by PIWI-interacting RNAs in Drosophila. Genes Genet. Syst. 2013, 88, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conte, C.; Dastugue, B.; Vaury, C. Coupling of enhancer and insulator properties identified in two retrotransposons modulates their mutagenic impact on nearby genes. Mol. Cell. Biol. 2002, 22, 1767–1777. [Google Scholar] [CrossRef] [Green Version]
- Minervini, C.F.; Ruggieri, S.; Traversa, M.; D’Aiuto, L.; Marsano, R.M.; Leronni, D.; Centomani, I.; De Giovanni, C.; Viggiano, L. Evidences for insulator activity of the 5′UTR of the Drosophila melanogaster LTR-retrotransposon ZAM. Mol. Genet. Genom. 2010, 283, 503–509. [Google Scholar] [CrossRef]
- Parkhurst, S.M.; Corces, V.G. Interactions among the gypsy transposable element and the yellow and the Suppressor of Hairy-wing loci in Drosophila melanogaster. Mol. Cell. Biol. 1986, 6, 47–53. [Google Scholar] [PubMed] [Green Version]
- Ali, A.; Han, K.; Liang, P. Role of Transposable Elements in Gene Regulation in the Human Genome. Life 2021, 11, 118. [Google Scholar] [CrossRef] [PubMed]
- Candelli, T.; Gros, J.; Libri, D. Pervasive transcription fine-tunes replication origin activity. eLife 2018, 7, e40802. [Google Scholar] [CrossRef]
- Porrua, O.; Boudvillain, M.; Libri, D. Transcription Termination: Variations on Common Themes. Trends Genet. 2016, 32, 508–522. [Google Scholar] [CrossRef]
- Porrua, O.; Libri, D. Transcription termination and the control of the transcriptome: Why, where and how to stop. Nat. Rev. Mol. Cell Biol. 2015, 16, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Hurst, L. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486. [Google Scholar] [CrossRef]
- Bouallègue, M.; Rouault, J.-D.; Hua-Van, A.; Makni, M.; Capy, P. Molecular evolution of piggyBac superfamily: From selfishness to domestication. Genome Biol. Evol. 2017, 9, 323–339. [Google Scholar] [CrossRef] [Green Version]
- Joly-Lopez, Z.; Forczek, E.; Hoen, D.R.; Juretic, N.; Bureau, T.E. A Gene Family Derived from Transposable Elements during Early Angiosperm Evolution Has Reproductive Fitness Benefits in Arabidopsis thaliana. PLoS Genet. 2012, 8, e1002931. [Google Scholar] [CrossRef] [Green Version]
- Aminetzach, Y.T.; Macpherson, J.M.; Petrov, D.A. Pesticide resistance via transposition-mediated adaptive gene truncation in Drosophila. Science 2005, 309, 764–767. [Google Scholar] [CrossRef] [Green Version]
- Huda, A.; Bowen, N.J.; Conley, A.B.; Jordan, I.K. Epigenetic regulation of transposable element derived human gene promoters. Gene 2011, 475, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Capy, P.; Anxolabehere, D.; Langin, T. The strange phylogenies of transposable elements: Are horizontal transfers the only explantation? Trends Genet. 1994, 10, 7–12. [Google Scholar] [CrossRef]
- Casola, C.; Lawing, A.M.; Betran, E.; Feschotte, C. PIF-like Transposons are common in Drosophila and have been repeatedly domesticated to generate new host genes. Mol. Biol. Evol. 2007, 24, 1872–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baduel, P.; Quadrana, L. Jumpstarting evolution: How transposition can facilitate adaptation to rapid environmental changes. Curr. Opin. Plant Biol. 2021, 61, 102043. [Google Scholar] [CrossRef]
- Joly-Lopez, Z.; Hoen, D.R.; Blanchette, M.; Bureau, T.E. Phylogenetic and Genomic Analyses Resolve the Origin of Important Plant Genes Derived from Transposable Elements. Mol. Biol. Evol. 2016, 33, 1937–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Han, G.-Z. Unearthing LTR Retrotransposon gag Genes Co-opted in the Deep Evolution of Eukaryotes. Mol. Biol. Evol. 2021, 38, 3267–3278. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Wang, Y.; Diaby, M.; Zong, W.; Shen, D.; Wang, S.; Chen, C.; Wang, X.; Song, C. Evolution of pogo, a separate superfamily of IS630-Tc1-mariner transposons, revealing recurrent domestication events in vertebrates. Mob. DNA 2020, 11, 25. [Google Scholar] [CrossRef]
- Baduel, P.; Colot, V. The epiallelic potential of transposable elements and its evolutionary significance in plants. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200123. [Google Scholar] [CrossRef] [PubMed]
- Quesneville, H.; Nouaud, D.; Anxolabehere, D. Recurrent Recruitment of the THAP DNA-Binding Domain and Molecular Domestication of the P-Transposable Element. Mol. Biol. Evol. 2005, 22, 741–746. [Google Scholar] [CrossRef]
- Hayward, A.; Ghazal, A.; Andersson, G.; Andersson, L.; Jern, P. ZBED Evolution: Repeated Utilization of DNA Transposons as Regulators of Diverse Host Functions. PLoS ONE 2013, 8, e59940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusa, K. piggyBac Transposon. Microbiol. Spectr. 2015, 3, MDNA3-0028-2014. [Google Scholar] [CrossRef] [PubMed]
- Ivics, Z.; Hackett, P.B.; Plasterk, R.H.; Izsvak, Z. Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 1997, 91, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Ivics, Z.; Kaufman, C.D.; Zayed, H.; Csaba, M.; Walisko, O.; Izsvak, Z. The Sleeping Beauty Transposable Element: Evolution, Regulation and Genetic Applications. Curr. Issues Mol. Biol. 2004, 6, 43–56. Available online: https://www.caister.com/cimb/abstracts/v6/43.html (accessed on 19 December 2021).
- Almojil, D.; Bourgeois, Y.; Falis, M.; Hariyani, I.; Wilcox, J.; Boissinot, S. The Structural, Functional and Evolutionary Impact of Transposable Elements in Eukaryotes. Genes 2021, 12, 918. [Google Scholar] [CrossRef] [PubMed]
- Kidwell, M.G. Transposable elements and the evolution of genome size in eukaryotes. Genetica 2002, 115, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.S. The Origins of Genome Complexity. Science 2003, 302, 1401–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Mueller, R.L. Hellbender Genome Sequences Shed Light on Genomic Expansion at the Base of Crown Salamanders. Genome Biol. Evol. 2014, 6, 1818–1829. [Google Scholar] [CrossRef] [Green Version]
- Grandbastien, M.-A. LTR retrotransposons, handy hitchhikers of plant regulation and stress response. Biochim. Biophys. Acta-Gene Regul. Mech. 2015, 1849, 403–416. [Google Scholar] [CrossRef]
- Grandbastien, M.A.; Lucas, H.; Morel, J.B.; Mhiri, C.; Vernhettes, S.; Casacuberta, J.M. The expression of the tobacco Tnt1 retrotransposon is linked to plant defense responses. Genetica 1997, 100, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Mhiri, C.; Morel, J.B.; Vernhettes, S.; Casacuberta, J.M.; Lucas, H.; Grandbastien, M.A. The promoter of the tobacco Tnt1 retrotransposon is induced by wounding and by abiotic stress. Plant Mol. Biol. 1997, 33, 257–266. [Google Scholar] [CrossRef]
- Mager, D.L.; Stoye, J.P. Mammalian Endogenous Retroviruses. Microbiol. Spectr. 2015, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- Hermant, C.; Torres-Padilla, M.-E. TFs for TEs: The transcription factor repertoire of mammalian transposable elements. Genes Dev. 2021, 35, 22–39. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capy, P. Taming, Domestication and Exaptation: Trajectories of Transposable Elements in Genomes. Cells 2021, 10, 3590. https://doi.org/10.3390/cells10123590
Capy P. Taming, Domestication and Exaptation: Trajectories of Transposable Elements in Genomes. Cells. 2021; 10(12):3590. https://doi.org/10.3390/cells10123590
Chicago/Turabian StyleCapy, Pierre. 2021. "Taming, Domestication and Exaptation: Trajectories of Transposable Elements in Genomes" Cells 10, no. 12: 3590. https://doi.org/10.3390/cells10123590
APA StyleCapy, P. (2021). Taming, Domestication and Exaptation: Trajectories of Transposable Elements in Genomes. Cells, 10(12), 3590. https://doi.org/10.3390/cells10123590