
The Chloroplast Trans-Splicing RNA–Protein Supercomplex from the Green Alga Chlamydomonas reinhardtii


Abstract
:1. Introduction
2. Introns-Early or Introns-Late Debate
3. Group II Introns and Spliceosomal Introns Share Identical Splicing Mechanisms
4. Sequence Homologies and Mechanistic Similarities
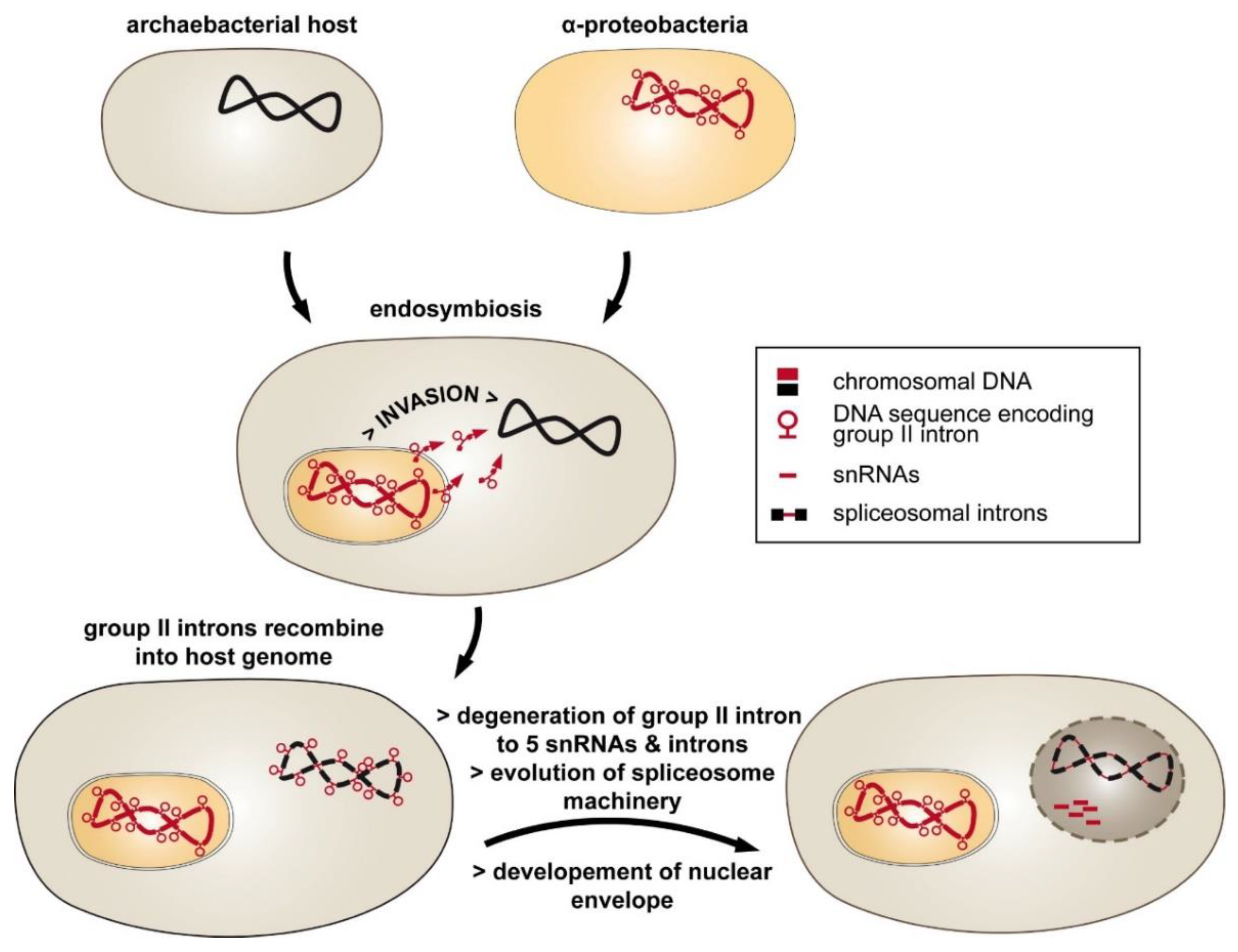
5. Trans-Spliced and Degenerated Group II Introns: A Step Toward Spliceosomal Introns?
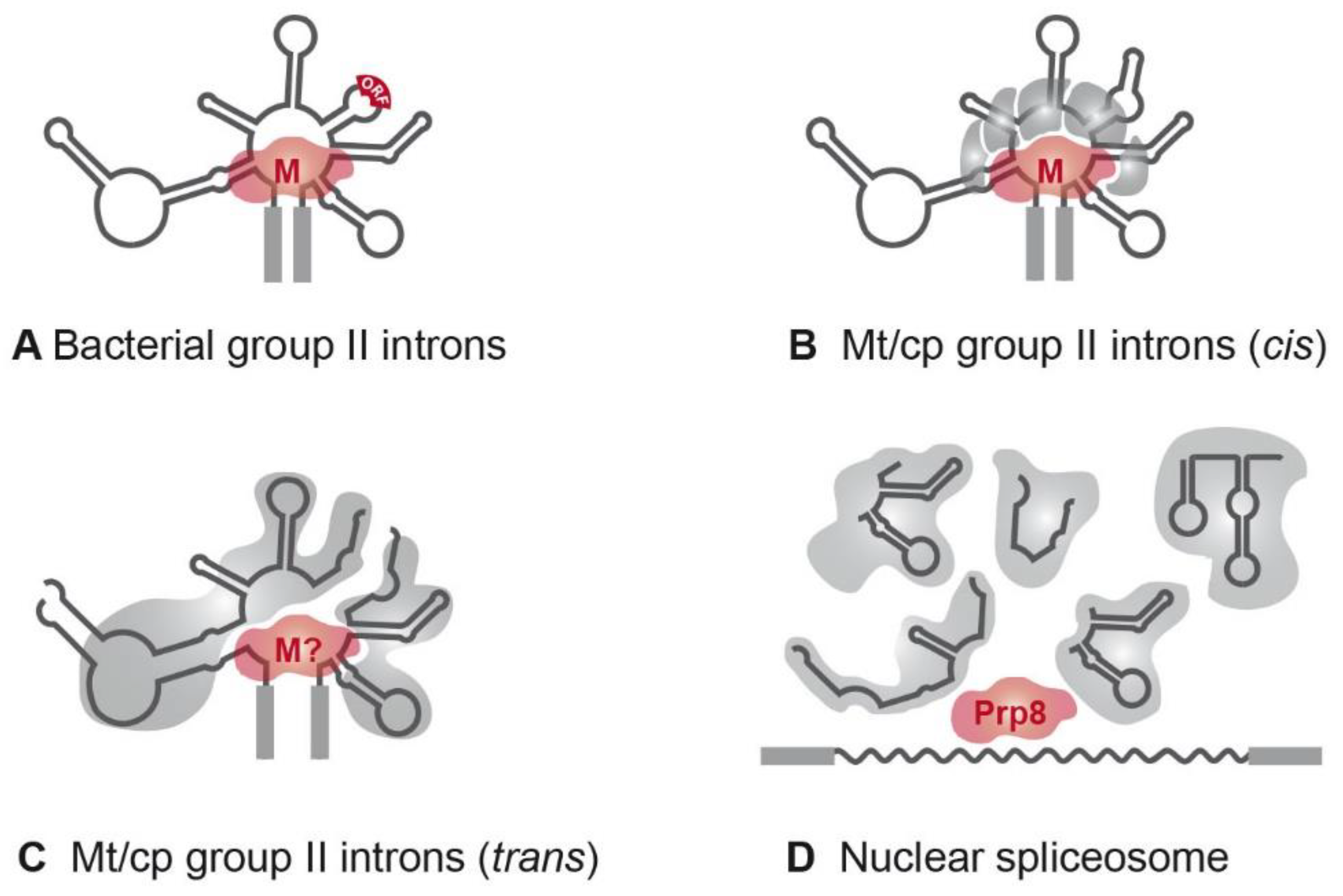
6. Splicing Factors: Ribozymes Need Facilitators
7. Evolution and Function of Nuclear Splicing Factors
8. Splicing Factors in Plastids and Mitochondria
9. Occurrence of trans-Splicing in Organellar Genomes
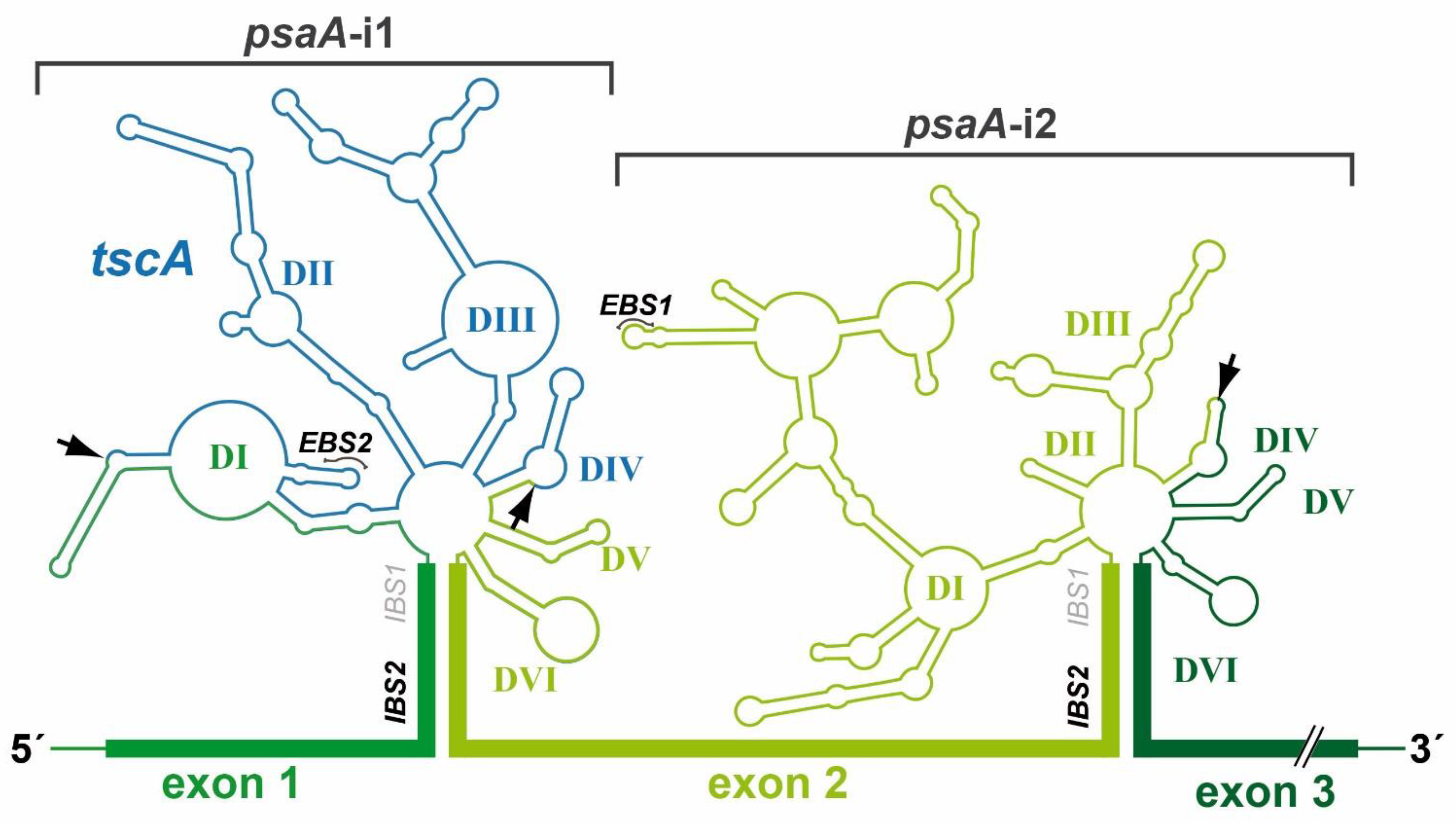
10. Two Group II Introns of the Chloroplast psaA Gene Are trans-Spliced in C. reinhardtii
11. Nuclear-Encoded Proteins Promote the Chloroplast trans-Splicing Process
12. The Chlamydomonas Chloroplast Spliceosome
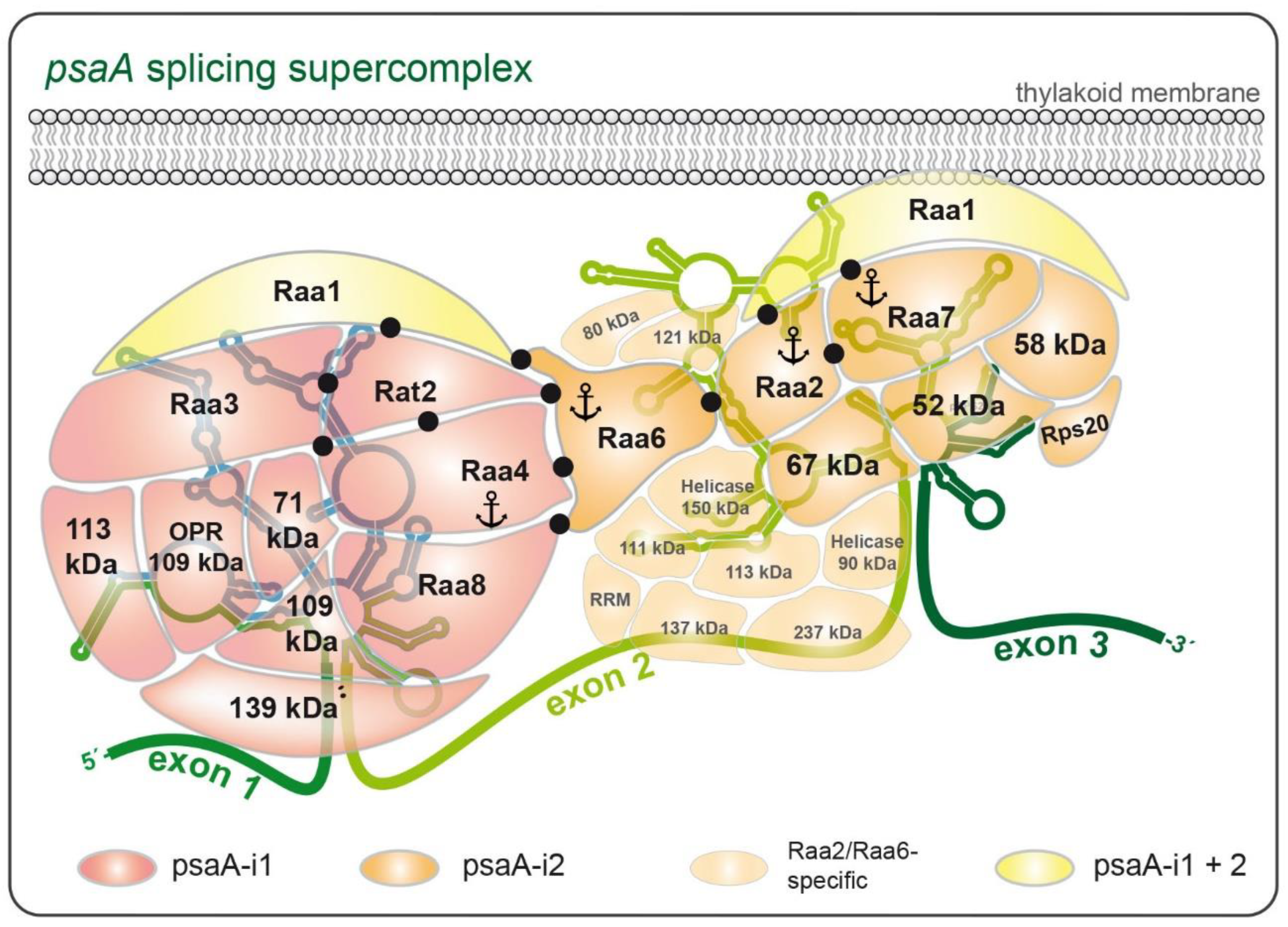
13. Evidence for the Formation of a Supercomplex During psaA Pre-mRNA Splicing
14. Splicing of psaA-i2 Is Mediated by a Membrane-Associated Complex
15. Putative Splicing Factors Identified by Mass Spectrometry
16. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| LC | liquid chromatography |
| MS | mass spectrometry |
| qRT-PCR | quantitative RT-PCR |
| RNP | ribonucleoprotein |
| SEC | size exclusion chromatography |
| TAP | tandem affinity purification |
References
- Berget, S.M.; Moore, C.; Sharp, P.A. Spliced Segments at the 5’ Terminus of Adenovirus 2 Late MRNA. Proc. Natl. Acad. Sci. USA 1977, 74, 3171–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, L.T.; Gelinas, R.E.; Broker, T.R.; Roberts, R.J. An Amazing Sequence Arrangement at the 5’ Ends of Adenovirus 2 Messenger RNA. Cell 1977, 12, 1–8. [Google Scholar] [CrossRef]
- Gilbert, W. Why Genes in Pieces? Nature 1978, 271, 501. [Google Scholar] [CrossRef]
- Chen, W.; Moore, M.J. Spliceosomes. Curr. Biol. 2015, 25, R181–R183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darnell, J.E.J. Implications of RNA-RNA Splicing in Evolution of Eukaryotic Cells. Science 1978, 202, 1257–1260. [Google Scholar] [CrossRef]
- Doolittle, W.F. Genes in Pieces: Were They Ever Together? Nature 1978, 272, 581–582. [Google Scholar] [CrossRef]
- Blake, C.C.F. Exons Encode Protein Functional Units. Nature 1979, 277, 598. [Google Scholar] [CrossRef]
- Gilbert, W.; de Souza, S.J.; Long, M. Origin of Genes. Proc. Natl. Acad. Sci. USA 1997, 94, 7698–7703. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.W. Recent Evidence for the Exon Theory of Genes. Genetica 2003, 118, 251–266. [Google Scholar] [CrossRef]
- Stoltzfus, A.; Spencer, D.F.; Zuker, M.; Logsdon, J.M.J.; Doolittle, W.F. Testing the Exon Theory of Genes: The Evidence from Protein Structure. Science 1994, 265, 202–207. [Google Scholar] [CrossRef]
- Cech, T.R. The Generality of Self-Splicing RNA: Relationship to Nuclear MRNA Splicing. Cell 1986, 44, 207–210. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Intron Phylogeny: A New Hypothesis. Trends Genet. 1991, 7, 145–148. [Google Scholar] [CrossRef]
- Bonen, L.; Vogel, J. The Ins and Outs of Group II Introns. Trends Genet. 2001, 17, 322–331. [Google Scholar] [CrossRef]
- Lambowitz, A.M.; Zimmerly, S. Group II Introns: Mobile Ribozymes That Invade DNA. Cold Spring Harb. Perspect. Biol. 2011, 3, a003616. [Google Scholar] [CrossRef]
- Sharp, P.A. Five Easy Pieces. Science 1991, 254, 663. [Google Scholar] [CrossRef]
- Rest, J.S.; Mindell, D.P. Retroids in Archaea: Phylogeny and Lateral Origins. Mol. Biol. Evol. 2003, 20, 1134–1142. [Google Scholar] [CrossRef] [Green Version]
- Martin, W.; Koonin, E.V. Introns and the Origin of Nucleus-Cytosol Compartmentalization. Nature 2006, 440, 41–45. [Google Scholar] [CrossRef]
- Sontheimer, E.J.; Gordon, P.M.; Piccirilli, J.A. Metal Ion Catalysis during Group II Intron Self-Splicing: Parallels with the Spliceosome. Genes Dev. 1999, 13, 1729–1741. [Google Scholar] [CrossRef] [Green Version]
- Wahl, M.C.; Will, C.L.; Lührmann, R. The Spliceosome: Design Principles of a Dynamic RNP Machine. Cell 2009, 136, 701–718. [Google Scholar] [CrossRef] [Green Version]
- de Longevialle, A.F.; Small, I.D.; Lurin, C. Nuclearly Encoded Splicing Factors Implicated in RNA Splicing in Higher Plant Organelles. Mol. Plant 2010, 3, 691–705. [Google Scholar] [CrossRef] [Green Version]
- Hegele, A.; Kamburov, A.; Grossmann, A.; Sourlis, C.; Wowro, S.; Weimann, M.; Will, C.L.; Pena, V.; Lührmann, R.; Stelzl, U. Dynamic Protein-Protein Interaction Wiring of the Human Spliceosome. Mol. Cell 2012, 45, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, F.; Umesono, K.; Ozeki, H. Comparative and Functional Anatomy of Group II Catalytic Introns—A Review. Gene 1989, 82, 5–30. [Google Scholar] [CrossRef]
- Pyle, A.M. The Tertiary Structure of Group II Introns: Implications for Biological Function and Evolution. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 215–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcia, M.; Somarowthu, S.; Pyle, A.M. Now on Display: A Gallery of Group II Intron Structures at Different Stages of Catalysis. Mob. DNA 2013, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, J.L.; Boulanger, S.C.; Dib-Hajj, S.D.; Hebbar, S.K.; Perlman, P.S. Group II Introns Deleted for Multiple Substructures Retain Self-Splicing Activity. Mol. Cell. Biol. 1992, 12, 1950–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, P.Z.; Pyle, A.M. The Architectural Organization and Mechanistic Function of Group II Intron Structural Elements. Curr. Opin. Struct. Biol. 1998, 8, 301–308. [Google Scholar] [CrossRef]
- Toor, N.; Keating, K.S.; Fedorova, O.; Rajashankar, K.; Wang, J.; Pyle, A.M. Tertiary Architecture of the Oceanobacillus Iheyensis Group II Intron. RNA 2010, 16, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Schmitz-Linneweber, C.; Lampe, M.-K.; Sultan, L.D.; Ostersetzer-Biran, O. Organellar Maturases: A Window into the Evolution of the Spliceosome. Biochim. Biophys. Acta 2015, 1847, 798–808. [Google Scholar] [CrossRef] [Green Version]
- Will, C.L.; Lührmann, R. Spliceosome Structure and Function. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [Green Version]
- Valadkhan, S.; Jaladat, Y. The Spliceosomal Proteome: At the Heart of the Largest Cellular Ribonucleoprotein Machine. Proteomics 2010, 10, 4128–4141. [Google Scholar] [CrossRef] [Green Version]
- Smathers, C.M.; Robart, A.R. The Mechanism of Splicing as Told by Group II Introns: Ancestors of the Spliceosome. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 194390. [Google Scholar] [CrossRef] [PubMed]
- Vosseberg, J.; Snel, B. Domestication of Self-Splicing Introns during Eukaryogenesis: The Rise of the Complex Spliceosomal Machinery. Biol. Direct 2017, 12, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Query, C.C.; Moore, M.J.; Sharp, P.A. Branch Nucleophile Selection in Pre-MRNA Splicing: Evidence for the Bulged Duplex Model. Genes Dev. 1994, 8, 587–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berglund, J.A.; Rosbash, M.; Schultz, S.C. Crystal Structure of a Model Branchpoint-U2 SnRNA Duplex Containing Bulged Adenosines. RNA 2001, 7, 682–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robart, A.R.; Chan, R.T.; Peters, J.K.; Rajashankar, K.R.; Toor, N. Crystal Structure of a Eukaryotic Group II Intron Lariat. Nature 2014, 514, 193–197. [Google Scholar] [CrossRef] [Green Version]
- Keating, K.S.; Toor, N.; Perlman, P.S.; Pyle, A.M. A Structural Analysis of the Group II Intron Active Site and Implications for the Spliceosome. RNA 2010, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fica, S.M.; Mefford, M.A.; Piccirilli, J.A.; Staley, J.P. Evidence for a Group II Intron-like Catalytic Triplex in the Spliceosome. Nat. Struct. Mol. Biol. 2014, 21, 464–471. [Google Scholar] [CrossRef] [Green Version]
- Dayie, K.T.; Padgett, R.A. A Glimpse into the Active Site of a Group II Intron and Maybe the Spliceosome, Too. RNA 2008, 14, 1697–1703. [Google Scholar] [CrossRef] [Green Version]
- Fedorova, O.; Pyle, A.M. A Conserved Element That Stabilizes the Group II Intron Active Site. RNA 2008, 14, 1048–1056. [Google Scholar] [CrossRef] [Green Version]
- Swisher, J.; Duarte, C.M.; Su, L.J.; Pyle, A.M. Visualizing the Solvent-Inaccessible Core of a Group II Intron Ribozyme. EMBO J. 2001, 20, 2051–2061. [Google Scholar] [CrossRef] [Green Version]
- Toor, N.; Keating, K.S.; Taylor, S.D.; Pyle, A.M. Crystal Structure of a Self-Spliced Group II Intron. Science 2008, 320, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.T.; Maroney, P.A.; Darzynkiwicz, E.; Nilsen, T.W. U6 SnRNA Function in Nuclear Pre-MRNA Splicing: A Phosphorothioate Interference Analysis of the U6 Phosphate Backbone. RNA 1995, 1, 46–54. [Google Scholar] [PubMed]
- Seetharaman, M.; Eldho, N.V.; Padgett, R.A.; Dayie, K.T. Structure of a Self-Splicing Group II Intron Catalytic Effector Domain 5: Parallels with Spliceosomal U6 RNA. RNA 2006, 12, 235–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrizio, P.; Abelson, J. Point Mutations in Yeast U6 SnRNA Can Specifically Block the First or Second Step of Pre-MRNA Splicing in Vitro. Mol. Biol. Rep. 1990, 14, 135. [Google Scholar] [CrossRef] [PubMed]
- Madhani, H.D.; Bordonné, R.; Guthrie, C. Multiple Roles for U6 SnRNA in the Splicing Pathway. Genes Dev. 1990, 4, 2264–2277. [Google Scholar] [CrossRef] [PubMed]
- Datta, B.; Weiner, A.M. The Phylogenetically Invariant ACAGAGA and AGC Sequences of U6 Small Nuclear RNA Are More Tolerant of Mutation in Human Cells than in Saccharomyces cerevisiae. Mol. Cell. Biol. 1993, 13, 5377–5382. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.G.; Colas des Francs-Small, C.; Ostersetzer-Biran, O. Group II Intron Splicing Factors in Plant Mitochondria. Front. Plant Sci. 2014, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- Doolittle, W.F. The Trouble with (Group II) Introns. Proc. Natl. Acad. Sci. USA 2014, 111, 6536–6537. [Google Scholar] [CrossRef] [Green Version]
- Chalamcharla, V.R.; Curcio, M.J.; Belfort, M. Nuclear Expression of a Group II Intron Is Consistent with Spliceosomal Intron Ancestry. Genes Dev. 2010, 24, 827–836. [Google Scholar] [CrossRef] [Green Version]
- Qu, G.; Dong, X.; Piazza, C.L.; Chalamcharla, V.R.; Lutz, S.; Curcio, M.J.; Belfort, M. RNA–RNA Interactions and Pre-MRNA Mislocalization as Drivers of Group II Intron Loss from Nuclear Genomes. Proc. Natl. Acad. Sci. USA 2014, 111, 6612–6617. [Google Scholar] [CrossRef] [Green Version]
- Belhocine, K.; Mak, A.B.; Cousineau, B. Trans-Splicing Versatility of the Ll.LtrB Group II Intron. RNA 2008, 14, 1782–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glanz, S.; Kück, U. Trans-Splicing of Organelle Introns--a Detour to Continuous RNAs. Bioessays 2009, 31, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Bonen, L. Tran-Splicing of Pre-MRNA in Plants, Animals, and Protists. FASEB J. 1993, 7, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Fukuzawa, H.; Kohchi, T.; Shirai, H.; Ohyama, K.; Umesono, K.; Inokucri, H.; Ozeki, H. Coding Sequences for Chloroplast Ribosomal Protein S12 from the Liverwort, Marchantia polymorpha, Are Separated Far Apart on the Different DNA Strands. FEBS Lett. 1986, 198, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Kück, U.; Choquet, Y.; Schneider, M.; Dron, M.; Bennoun, P. Structural and Transcription Analysis of Two Homologous Genes for the P700 Chlorophyll A-Apoproteins in Chlamydomonas reinhardii: Evidence for in Vivo Trans-Splicing. EMBO J. 1987, 6, 2185–2195. [Google Scholar] [CrossRef] [PubMed]
- Zaita, N.; Torazawa, K.; Shinozaki, K.; Sugiura, M. Trans Splicing in Vivo: Joining of Transcripts from the ‘Divided’ Gene for Ribosomal Protein S12 in the Chloroplasts of Tobacco. FEBS Lett. 1987, 210, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Michel, F.; Ferat, J.L. Structure and Activities of Group II Introns. Annu. Rev. Biochem. 1995, 64, 435–461. [Google Scholar] [CrossRef]
- Bonen, L. Cis- and Trans-Splicing of Group II Introns in Plant Mitochondria. Mitochondrion 2008, 8, 26–34. [Google Scholar] [CrossRef]
- Qiu, Y.-L.; Palmer, J.D. Many Independent Origins of Trans Splicing of a Plant Mitochondrial Group II Intron. J. Mol. Evol. 2004, 59, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Dlakić, M.; Mushegian, A. Prp8, the Pivotal Protein of the Spliceosomal Catalytic Center, Evolved from a Retroelement-Encoded Reverse Transcriptase. RNA 2011, 17, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Fromm, H.; Edelman, M.; Koller, B.; Goloubinoff, P.; Galun, E. The Enigma of the Gene Coding for Ribosomal Protein S12 in the Chloroplasts of Nicotiana. Nucleic Acids Res. 1986, 14, 883–898. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, A.-S.; Brouard, J.-S.; Charlebois, P.; Otis, C.; Lemieux, C.; Turmel, M. Distinctive Architecture of the Chloroplast Genome in the Chlorophycean Green Alga Stigeoclonium helveticum. Mol. Genet. Genom. 2006, 276, 464–477. [Google Scholar] [CrossRef]
- Brouard, J.-S.; Otis, C.; Lemieux, C.; Turmel, M. The Exceptionally Large Chloroplast Genome of the Green Alga Floydiella terrestris Illuminates the Evolutionary History of the Chlorophyceae. Genome Biol. Evol. 2010, 2, 240–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohchi, T.; Umesono, K.; Ogura, Y.; Komine, Y.; Nakahigashi, K.; Komano, T.; Yamada, Y.; Ozeki, H.; Ohyama, K. A Nicked Group II Intron and Trans-Splicing in Liverwort, Marchantia polymorpha, Chloroplasts. Nucleic Acids Res. 1988, 16, 10025–10036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouard, J.-S.; Otis, C.; Lemieux, C.; Turmel, M. Chloroplast DNA Sequence of the Green Alga Oedogonium cardiacum (Chlorophyceae): Unique Genome Architecture, Derived Characters Shared with the Chaetophorales and Novel Genes Acquired through Horizontal Transfer. BMC Genom. 2008, 9, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoop, V.; Altwasser, M.; Brennicke, A. A Tripartite Group II Intron in Mitochondria of an Angiosperm Plant. Mol. Gen. Genet. 1997, 255, 269–276. [Google Scholar] [CrossRef]
- Goldschmidt-Clermont, M.; Choquet, Y.; Girard-Bascou, J.; Michel, F.; Schirmer-Rahire, M.; Rochaix, J.D. A Small Chloroplast RNA May Be Required for Trans-Splicing in Chlamydomonas reinhardtii. Cell 1991, 65, 135–143. [Google Scholar] [CrossRef]
- Belhocine, K.; Mak, A.B.; Cousineau, B. Trans-Splicing of the Ll.LtrB Group II Intron in Lactococcus lactis. Nucleic Acids Res. 2007, 35, 2257–2268. [Google Scholar] [CrossRef]
- Ritlop, C.; Monat, C.; Cousineau, B. Isolation and Characterization of Functional Tripartite Group II Introns Using a Tn5-Based Genetic Screen. PLoS ONE 2012, 7, e41589. [Google Scholar] [CrossRef]
- McNeil, B.A.; Simon, D.M.; Zimmerly, S. Alternative Splicing of a Group II Intron in a Surface Layer Protein Gene in Clostridium tetani. Nucleic Acids Res. 2014, 42, 1959–1969. [Google Scholar] [CrossRef] [Green Version]
- Leoni, G.; Le Pera, L.; Ferrè, F.; Raimondo, D.; Tramontano, A. Coding Potential of the Products of Alternative Splicing in Human. Genome Biol. 2011, 12, R9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative Isoform Regulation in Human Tissue Transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, C.J.; Graveley, B.R. RNA Structure and the Mechanisms of Alternative Splicing. Curr. Opin. Genet. Dev. 2011, 21, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkan, A. Intron Splicing in Plant Organelles. In Molecular Biology and Biotechnology of Plant Organelles; Springer: Dordrecht, The Netherlands, 2004; pp. 295–322. [Google Scholar]
- Ahlert, D.; Piepenburg, K.; Kudla, J.; Bock, R. Evolutionary Origin of a Plant Mitochondrial Group II Intron from a Reverse Transcriptase/Maturase-Encoding Ancestor. J. Plant Res. 2006, 119, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.-C.; Kim, J.-H.; Kim, N.-S. Comprehensive Genomic Analyses with 115 Plastomes from Algae to Seed Plants: Structure, Gene Contents, GC Contents, and Introns. Genes Genom. 2020, 42, 553–570. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Chapdelaine, Y.; Bonen, L. Variation in Sequence and RNA Editing within Core Domains of Mitochondrial Group II Introns among Plants. Mol. Gen. Genet. MGG 2001, 264, 595–603. [Google Scholar] [CrossRef]
- Turmel, M.; Otis, C.; Lemieux, C. The Chloroplast Genome Sequence of Chara Vulgaris Sheds New Light into the Closest Green Algal Relatives of Land Plants. Mol. Biol. Evol. 2006, 23, 1324–1338. [Google Scholar] [CrossRef] [PubMed]
- Peebles, C.L.; Perlman, P.S.; Mecklenburg, K.L.; Petrillo, M.L.; Tabor, J.H.; Jarrell, K.A.; Cheng, H.L. A Self-Splicing RNA Excises an Intron Lariat. Cell 1986, 44, 213–223. [Google Scholar] [CrossRef]
- van der Veen, R.; Arnberg, A.C.; van der Horst, G.; Bonen, L.; Tabak, H.F.; Grivell, L.A. Excised Group II Introns in Yeast Mitochondria Are Lariats and Can Be Formed by Self-Splicing in Vitro. Cell 1986, 44, 225–234. [Google Scholar] [CrossRef]
- Costa, M.; Fontaine, J.-M.; Goër, S.L.; Michel, F. A Group II Self-Splicing Intron from the Brown Alga Pylaiella pittoralis is Active at Unusually Low Magnesium Concentrations and Forms Populations of Molecules with a Uniform Conformation11Edited by M. Yaniv. J. Mol. Biol. 1997, 274, 353–364. [Google Scholar] [CrossRef]
- Valadkhan, S.; Mohammadi, A.; Wachtel, C.; Manley, J.L. Protein-Free Spliceosomal SnRNAs Catalyze a Reaction That Resembles the First Step of Splicing. RNA 2007, 13, 2300–2311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadkhan, S.; Mohammadi, A.; Jaladat, Y.; Geisler, S. Protein-Free Small Nuclear RNAs Catalyze a Two-Step Splicing Reaction. Proc. Natl. Acad. Sci. USA 2009, 106, 11901–11906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fica, S.M.; Tuttle, N.; Novak, T.; Li, N.-S.; Lu, J.; Koodathingal, P.; Dai, Q.; Staley, J.P.; Piccirilli, J.A. RNA Catalyses Nuclear Pre-MRNA Splicing. Nature 2013, 503, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, I.A.; Norman, C.M.; Churcher, M.J.; Newman, A.J. Dissection of Prp8 Protein Defines Multiple Interactions with Crucial RNA Sequences in the Catalytic Core of the Spliceosome. RNA 2006, 12, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grainger, R.J.; Beggs, J.D. Prp8 Protein: At the Heart of the Spliceosome. RNA 2005, 11, 533–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veretnik, S.; Wills, C.; Youkharibache, P.; Valas, R.E.; Bourne, P.E. Sm/Lsm Genes Provide a Glimpse into the Early Evolution of the Spliceosome. PLoS Comput. Biol. 2009, 5, e1000315. [Google Scholar] [CrossRef]
- Kambach, C.; Walke, S.; Young, R.; Avis, J.M.; de la Fortelle, E.; Raker, V.A.; Lührmann, R.; Li, J.; Nagai, K. Crystal Structures of Two Sm Protein Complexes and Their Implications for the Assembly of the Spliceosomal SnRNPs. Cell 1999, 96, 375–387. [Google Scholar] [CrossRef]
- Matera, A.G.; Wang, Z. A Day in the Life of the Spliceosome. Nat. Rev. Mol. Cell Biol. 2014, 15, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Beggs, J.D. Lsm Proteins and RNA Processing. Biochem. Soc. Trans. 2005, 33, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Törö, I.; Thore, S.; Mayer, C.; Basquin, J.; Séraphin, B.; Suck, D. RNA Binding in an Sm Core Domain: X-Ray Structure and Functional Analysis of an Archaeal Sm Protein Complex. EMBO J. 2001, 20, 2293–2303. [Google Scholar] [CrossRef] [Green Version]
- Murina, V.N.; Nikulin, A.D. RNA-Binding Sm-like Proteins of Bacteria and Archaea. Similarity and Difference in Structure and Function. Biochemistry 2011, 76, 1434–1449. [Google Scholar] [CrossRef] [PubMed]
- Sun, C. The SF3b Complex: Splicing and Beyond. Cell. Mol. Life Sci. 2020, 77, 3583–3595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.C.; Caceres, J.F. The SR Protein Family of Splicing Factors: Master Regulators of Gene Expression. Biochem. J. 2009, 417, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidaver, R.M.; Fortner, D.M.; Loos-Austin, L.S.; Brow, D.A. Multiple Functions of Saccharomyces cerevisiae Splicing Protein Prp24 in U6 RNA Structural Rearrangements. Genetics 1999, 153, 1205–1218. [Google Scholar]
- Newman, A.J.; Nagai, K. Structural Studies of the Spliceosome: Blind Men and an Elephant. Curr. Opin. Struct. Biol. 2010, 20, 82–89. [Google Scholar] [CrossRef]
- Staley, J.P.; Guthrie, C. Mechanical Devices of the Spliceosome: Motors, Clocks, Springs, and Things. Cell 1998, 92, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Cordin, O.; Hahn, D.; Beggs, J.D. Structure, Function and Regulation of Spliceosomal RNA Helicases. Curr. Opin. Cell Biol. 2012, 24, 431–438. [Google Scholar] [CrossRef]
- De Bortoli, F.; Espinosa, S.; Zhao, R. DEAH-Box RNA Helicases in Pre-MRNA Splicing. Trends Biochem. Sci. 2020. [Google Scholar] [CrossRef]
- Neuhaus, H.; Link, G. The Chloroplast TRNALys(UUU) Gene from Mustard (Sinapis alba) Contains a Class II Intron Potentially Coding for a Maturase-Related Polypeptide. Curr. Genet. 1987, 11, 251–257. [Google Scholar] [CrossRef]
- Wahleithner, J.A.; MacFarlane, J.L.; Wolstenholme, D.R. A Sequence Encoding a Maturase-Related Protein in a Group II Intron of a Plant Mitochondrial Nad1 Gene. Proc. Natl. Acad. Sci. USA 1990, 87, 548–552. [Google Scholar] [CrossRef] [Green Version]
- Barthet, M.M.; Hilu, K.W. Expression of MatK: Functional and Evolutionary Implications. Am. J. Bot. 2007, 94, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Zoschke, R.; Nakamura, M.; Liere, K.; Sugiura, M.; Börner, T.; Schmitz-Linneweber, C. An Organellar Maturase Associates with Multiple Group II Introns. Proc. Natl. Acad. Sci. USA 2010, 107, 3245–3250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turmel, M.; Otis, C.; Lemieux, C. The Complete Chloroplast DNA Sequences of the Charophycean Green Algae Staurastrum and Zygnema Reveal That the Chloroplast Genome Underwent Extensive Changes during the Evolution of the Zygnematales. BMC Biol. 2005, 3, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNeal, J.R.; Arumugunathan, K.; Kuehl, J.V.; Boore, J.L.; DePamphilis, C.W. Systematics and Plastid Genome Evolution of the Cryptically Photosynthetic Parasitic Plant Genus Cuscuta (Convolvulaceae). BMC Biol. 2007, 5, 55. [Google Scholar] [CrossRef] [Green Version]
- Mohr, G.; Lambowitz, A.M. Putative Proteins Related to Group II Intron Reverse Transcriptase/Maturases are Encoded by Nuclear Genes in Higher Plants. Nucleic Acids Res. 2003, 31, 647–652. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Mower, J.P. Evolution of Plant Mitochondrial Intron-Encoded Maturases: Frequent Lineage-Specific Loss and Recurrent Intracellular Transfer to the Nucleus. J. Mol. Evol. 2013, 77, 43–54. [Google Scholar] [CrossRef]
- Keren, I.; Bezawork-Geleta, A.; Kolton, M.; Maayan, I.; Belausov, E.; Levy, M.; Mett, A.; Gidoni, D.; Shaya, F.; Ostersetzer-Biran, O. AtnMat2, a Nuclear-Encoded Maturase Required for Splicing of Group-II Introns in Arabidopsis Mitochondria. RNA 2009, 15, 2299–2311. [Google Scholar] [CrossRef] [Green Version]
- Malik, S.; Upadhyaya, K.C.; Khurana, S.M.P. Phylogenetic Analysis of Nuclear-Encoded RNA Maturases. Evol. Bioinform. 2017, 13. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, B.D.; Barkan, A. Recruitment of a Peptidyl-TRNA Hydrolase as a Facilitator of Group II Intron Splicing in Chloroplasts. EMBO J. 2001, 20, 872–879. [Google Scholar] [CrossRef]
- Merendino, L.; Perron, K.; Rahire, M.; Howald, I.; Rochaix, J.-D.; Goldschmidt-Clermont, M. A Novel Multifunctional Factor Involved in Trans-Splicing of Chloroplast Introns in Chlamydomonas. Nucleic Acids Res. 2006, 34, 262–274. [Google Scholar] [CrossRef] [Green Version]
- Watkins, K.P.; Kroeger, T.S.; Cooke, A.M.; Williams-Carrier, R.E.; Friso, G.; Belcher, S.E.; van Wijk, K.J.; Barkan, A. A Ribonuclease III Domain Protein Functions in Group II Intron Splicing in Maize Chloroplasts. Plant Cell 2007, 19, 2606–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkan, A. Expression of Plastid Genes: Organelle-Specific Elaborations on a Prokaryotic Scaffold. Plant Physiol. 2011, 155, 1520–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perron, K.; Goldschmidt-Clermont, M.; Rochaix, J.D. A Factor Related to Pseudouridine Synthases Is Required for Chloroplast Group II Intron Trans-Splicing in Chlamydomonas reinhardtii. EMBO J. 1999, 18, 6481–6490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, J.; Glanz, S.; Bunse-Grassmann, A.; Kruse, O.; Kück, U. RNA Trans-Splicing: Identification of Components of a Putative Chloroplast Spliceosome. Eur. J. Cell Biol. 2010, 89, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Till, B.; Schmitz-Linneweber, C.; Williams-Carrier, R.; Barkan, A. CRS1 Is a Novel Group II Intron Splicing Factor That Was Derived from a Domain of Ancient Origin. RNA 2001, 7, 1227–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostheimer, G.J.; Williams-Carrier, R.; Belcher, S.; Osborne, E.; Gierke, J.; Barkan, A. Group II Intron Splicing Factors Derived by Diversification of an Ancient RNA-Binding Domain. EMBO J. 2003, 22, 3919–3929. [Google Scholar] [CrossRef] [Green Version]
- Barkan, A.; Klipcan, L.; Ostersetzer, O.; Kawamura, T.; Asakura, Y.; Watkins, K.P. The CRM Domain: An RNA Binding Module Derived from an Ancient Ribosome-Associated Protein. RNA 2007, 13, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Keren, I.; Klipcan, L.; Bezawork-Geleta, A.; Kolton, M.; Shaya, F.; Ostersetzer-Biran, O. Characterization of the Molecular Basis of Group II Intron RNA Recognition by CRS1-CRM Domains. J. Biol. Chem. 2008, 283, 23333–23342. [Google Scholar] [CrossRef] [Green Version]
- Hammani, K.; Barkan, A. An MTERF Domain Protein Functions in Group II Intron Splicing in Maize Chloroplasts. Nucleic Acids Res. 2014, 42, 5033–5042. [Google Scholar] [CrossRef] [Green Version]
- Schmitz-Linneweber, C.; Small, I. Pentatricopeptide Repeat Proteins: A Socket Set for Organelle Gene Expression. Trends Plant Sci. 2008, 13, 663–670. [Google Scholar] [CrossRef]
- Rahire, M.; Laroche, F.; Cerutti, L.; Rochaix, J.-D. Identification of an OPR Protein Involved in the Translation Initiation of the PsaB Subunit of Photosystem I. Plant J. 2012, 72, 652–661. [Google Scholar] [CrossRef] [PubMed]
- Marx, C.; Wünsch, C.; Kück, U. The Octatricopeptide Repeat Protein Raa8 Is Required for Chloroplast Trans Splicing. Eukaryot. Cell 2015, 14, 998–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostersetzer, O.; Cooke, A.M.; Watkins, K.P.; Barkan, A. CRS1, a Chloroplast Group II Intron Splicing Factor, Promotes Intron Folding through Specific Interactions with Two Intron Domains. Plant Cell 2005, 17, 241–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Köhler, D.; Schmidt-Gattung, S.; Binder, S. The DEAD-Box Protein PMH2 Is Required for Efficient Group II Intron Splicing in Mitochondria of Arabidopsis thaliana. Plant Mol. Biol. 2010, 72, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Rivier, C.; Goldschmidt-Clermont, M.; Rochaix, J.D. Identification of an RNA-Protein Complex Involved in Chloroplast Group II Intron Trans-Splicing in Chlamydomonas reinhardtii. EMBO J. 2001, 20, 1765–1773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perron, K.; Goldschmidt-Clermont, M.; Rochaix, J.-D. A Multiprotein Complex Involved in Chloroplast Group II Intron Splicing. RNA 2004, 10, 704–711. [Google Scholar] [CrossRef] [Green Version]
- Matthes, A.; Schmidt-Gattung, S.; Köhler, D.; Forner, J.; Wildum, S.; Raabe, M.; Urlaub, H.; Binder, S. Two DEAD-Box Proteins May Be Part of RNA-Dependent High-Molecular-Mass Protein Complexes in Arabidopsis Mitochondria. Plant Physiol. 2007, 145, 1637–1646. [Google Scholar] [CrossRef] [Green Version]
- Blocker, F.J.H.; Mohr, G.; Conlan, L.H.; Qi, L.; Belfort, M.; Lambowitz, A.M. Domain Structure and Three-Dimensional Model of a Group II Intron-Encoded Reverse Transcriptase. RNA 2005, 11, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Shaikh, T.R.; Gupta, K.; Contreras-Martin, L.M.; Grassucci, R.A.; Van Duyne, G.D.; Frank, J.; Belfort, M. The Group II Intron Ribonucleoprotein Precursor Is a Large, Loosely Packed Structure. Nucleic Acids Res. 2011, 39, 2845–2854. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, M.; Noah, J.W.; Lambowitz, A.M. Mechanism of Maturase-Promoted Group II Intron Splicing. EMBO J. 2001, 20, 7259–7270. [Google Scholar] [CrossRef]
- Noah, J.W.; Lambowitz, A.M. Effects of Maturase Binding and Mg2+ Concentration on Group II Intron RNA Folding Investigated by UV Cross-Linking. Biochemistry 2003, 42, 12466–12480. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, M.; Saldanha, R.; Ma, H.; Wank, H.; Yang, J.; Mohr, G.; Cavanagh, S.; Dunny, G.M.; Belfort, M.; Lambowitz, A.M. A Bacterial Group II Intron Encoding Reverse Transcriptase, Maturase, and DNA Endonuclease Activities: Biochemical Demonstration of Maturase Activity and Insertion of New Genetic Information within the Intron. Genes Dev. 1997, 11, 2910–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldanha, R.; Chen, B.; Wank, H.; Matsuura, M.; Edwards, J.; Lambowitz, A.M. RNA and Protein Catalysis in Group II Intron Splicing and Mobility Reactions Using Purified Components. Biochemistry 1999, 38, 9069–9083. [Google Scholar] [CrossRef] [PubMed]
- Malek, O.; Knoop, V. Trans-Splicing Group II Introns in Plant Mitochondria: The Complete Set of Cis-Arranged Homologs in Ferns, Fern Allies, and a Hornwort. RNA 1998, 4, 1599–1609. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Zhu, A.; Fan, W.; Adams, R.P.; Mower, J.P. Extensive Shifts from Cis- to Trans-Splicing of Gymnosperm Mitochondrial Introns. Mol. Biol. Evol. 2020, 37, 1615–1620. [Google Scholar] [CrossRef]
- Mower, J.P. Variation in Protein Gene and Intron Content among Land Plant Mitogenomes. Mitochondrion 2020, 53, 203–213. [Google Scholar] [CrossRef]
- Marande, W.; Burger, G. Mitochondrial DNA as a Genomic Jigsaw Puzzle. Science 2007, 318, 415. [Google Scholar] [CrossRef]
- Kaur, B.; Záhonová, K.; Valach, M.; Faktorová, D.; Prokopchuk, G.; Burger, G.; Lukeš, J. Gene Fragmentation and RNA Editing without Borders: Eccentric Mitochondrial Genomes of Diplonemids. Nucleic Acids Res. 2020, 48, 2694–2708. [Google Scholar] [CrossRef]
- Gallaher, S.D.; Fitz-Gibbon, S.T.; Strenkert, D.; Purvine, S.O.; Pellegrini, M.; Merchant, S.S. High-Throughput Sequencing of the Chloroplast and Mitochondrion of Chlamydomonas reinhardtii to Generate Improved de Novo Assemblies, Analyze Expression Patterns and Transcript Speciation, and Evaluate Diversity among Laboratory Strains and Wild Isolates. Plant J. 2018, 93, 545–565. [Google Scholar] [CrossRef] [Green Version]
- Goldschmidt-Clermont, M.; Girard-Bascou, J.; Choquet, Y.; Rochaix, J.D. Trans-Splicing Mutants of Chlamydomonas reinhardtii. Mol. Gen. Genet. 1990, 223, 417–425. [Google Scholar] [CrossRef]
- Glanz, S.; Jacobs, J.; Kock, V.; Mishra, A.; Kück, U. Raa4 Is a Trans-Splicing Factor That Specifically Binds Chloroplast TscA Intron RNA. Plant J. 2012, 69, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Reifschneider, O.; Marx, C.; Jacobs, J.; Kollipara, L.; Sickmann, A.; Wolters, D.; Kück, U. A Ribonucleoprotein Supercomplex Involved in Trans-Splicing of Organelle Group II Introns. J. Biol. Chem. 2016, 291, 23330–23342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefebvre-Legendre, L.; Reifschneider, O.; Kollipara, L.; Sickmann, A.; Wolters, D.; Kück, U.; Goldschmidt-Clermont, M. A Pioneer Protein Is Part of a Large Complex Involved in Trans-Splicing of a Group II Intron in the Chloroplast of Chlamydomonas reinhardtii. Plant J. 2016, 85, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balczun, C.; Bunse, A.; Hahn, D.; Bennoun, P.; Nickelsen, J.; Kück, U. Two Adjacent Nuclear Genes Are Required for Functional Complementation of a Chloroplast Trans-Splicing Mutant from Chlamydomonas reinhardtii. Plant J. 2005, 43, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Rigaut, G.; Shevchenko, A.; Rutz, B.; Wilm, M.; Mann, M.; Séraphin, B. A Generic Protein Purification Method for Protein Complex Characterization and Proteome Exploration. Nat. Biotechnol. 1999, 17, 1030–1032. [Google Scholar] [CrossRef] [PubMed]
- Puig, O.; Caspary, F.; Rigaut, G.; Rutz, B.; Bouveret, E.; Bragado-Nilsson, E.; Wilm, M.; Séraphin, B. The Tandem Affinity Purification (TAP) Method: A General Procedure of Protein Complex Purification. Methods 2001, 24, 218–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, I.-F. Mass Spectrometry-Based Proteomic Analysis of the Epitope-Tag Affinity Purified Protein Complexes in Eukaryotes. Proteomics 2006, 6, 6158–6166. [Google Scholar] [CrossRef]
- Van Leene, J.; Stals, H.; Eeckhout, D.; Persiau, G.; Van De Slijke, E.; Van Isterdael, G.; De Clercq, A.; Bonnet, E.; Laukens, K.; Remmerie, N.; et al. A Tandem Affinity Purification-Based Technology Platform to Study the Cell Cycle Interactome in Arabidopsis thaliana. Mol. Cell. Proteomics 2007, 6, 1226–1238. [Google Scholar] [CrossRef] [Green Version]
- Bayram, O.; Krappmann, S.; Ni, M.; Bok, J.W.; Helmstaedt, K.; Valerius, O.; Braus-Stromeyer, S.; Kwon, N.-J.; Keller, N.P.; Yu, J.-H.; et al. VelB/VeA/LaeA Complex Coordinates Light Signal with Fungal Development and Secondary Metabolism. Science 2008, 320, 1504–1506. [Google Scholar] [CrossRef]
- Bloemendal, S.; Bernhards, Y.; Bartho, K.; Dettmann, A.; Voigt, O.; Teichert, I.; Seiler, S.; Wolters, D.A.; Pöggeler, S.; Kück, U. A Homologue of the Human STRIPAK Complex Controls Sexual Development in Fungi. Mol. Microbiol. 2012, 84, 310–323. [Google Scholar] [CrossRef]
- Kroeger, T.S.; Watkins, K.P.; Friso, G.; van Wijk, K.J.; Barkan, A. A Plant-Specific RNA-Binding Domain Revealed through Analysis of Chloroplast Group II Intron Splicing. Proc. Natl. Acad. Sci. USA 2009, 106, 4537–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, Y.-W.; Wang, H.-J.; Hsieh, M.-H.; Hsieh, H.-L.; Jauh, G.-Y. Arabidopsis MTERF15 Is Required for Mitochondrial Nad2 Intron 3 Splicing and Functional Complex I Activity. PLoS ONE 2014, 9, e112360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reifschneider, O. Spliceosome-like Complexes in Chloroplasts of the Green Alga Chlamydomonas reinhardtii. Ph.D. Thesis, Ruhr-University Bochum, Bochum, Germany, 2015. [Google Scholar]
- Fourmy, D.; Yoshizawa, S. Protein-RNA Footprinting: An Evolving Tool. Wiley Interdiscip. Rev. RNA 2012, 3, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Ule, J.; Jensen, K.B.; Ruggiu, M.; Mele, A.; Ule, A.; Darnell, R.B. CLIP Identifies Nova-Regulated RNA Networks in the Brain. Science 2003, 302, 1212–1215. [Google Scholar] [CrossRef] [PubMed]
- Germain, A.; Hotto, A.M.; Barkan, A.; Stern, D.B. RNA Processing and Decay in Plastids. Wiley Interdiscip. Rev. RNA 2013, 4, 295–316. [Google Scholar] [CrossRef] [PubMed]
- Khrouchtchova, A.; Monde, R.-A.; Barkan, A. A Short PPR Protein Required for the Splicing of Specific Group II Introns in Angiosperm Chloroplasts. RNA 2012, 18, 1197–1209. [Google Scholar] [CrossRef] [Green Version]
- Kotzer-Nevo, H.; de Lima Alves, F.; Rappsilber, J.; Sperling, J.; Sperling, R. Supraspliceosomes at Defined Functional States Portray the Pre-Assembled Nature of the Pre-MRNA Processing Machine in the Cell Nucleus. Int. J. Mol. Sci. 2014, 15, 11637–11664. [Google Scholar] [CrossRef] [Green Version]
- Shefer, K.; Sperling, J.; Sperling, R. The Supraspliceosome—A Multi-Task Machine for Regulated Pre-MRNA Processing in the Cell Nucleus. Comput. Struct. Biotechnol. J. 2014, 11, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Margulies, M.M.; Michaels, A. Ribosomes Bound to Chloroplast Membranes in Chlamydomonas reinhardtii. J. Cell Biol. 1974, 60, 65–77. [Google Scholar] [CrossRef]
- Jagendorf, A.T.; Michaels, A. Rough Thylakoids: Translation on Photosynthetic Membranes. Plant Sci. 1990, 71, 137–145. [Google Scholar] [CrossRef]
- Rochaix, J.-D.; Perron, K.; Dauvillée, D.; Laroche, F.; Takahashi, Y.; Goldschmidt-Clermont, M. Post-Transcriptional Steps Involved in the Assembly of Photosystem I in Chlamydomonas. Biochem. Soc. Trans. 2004, 32, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre-Legendre, L.; Choquet, Y.; Kuras, R.; Loubéry, S.; Douchi, D.; Goldschmidt-Clermont, M. A Nucleus-Encoded Helical-Repeat Protein Which Is Regulated by Iron Availability Controls Chloroplast PsaA MRNA Expression in Chlamydomonas. Plant Physiol. 2015, pp.114.253906. [Google Scholar] [CrossRef] [Green Version]
- Dauvillée, D.; Stampacchia, O.; Girard-Bascou, J.; Rochaix, J.-D. Tab2 Is a Novel Conserved RNA Binding Protein Required for Translation of the Chloroplast PsaB MRNA. EMBO J. 2003, 22, 6378–6388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breidenbach, E.; Leu, S.; Michaels, A.; Boschetti, A. Synthesis of EF-Tu and Distribution of Its MRNA between Stroma and Thylakoids during the Cell Cycle of Chlamydomonas reinhardii. Biochim. Biophys. Acta Gene Struct. Expr. 1990, 1048, 209–216. [Google Scholar] [CrossRef]
- Mühlbauer, S.K.; Eichacker, L.A. The Stromal Protein Large Subunit of Ribulose-1,5-Bisphosphate Carboxylase Is Translated by Membrane-Bound Ribosomes. Eur. J. Biochem. 1999, 261, 784–788. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Beligni, M.V.; Prieto, S.; Haynes, P.A.; McDonald, W.H.; Yates, J.R., 3rd; Mayfield, S.P. Proteomic Characterization of the Chlamydomonas reinhardtii Chloroplast Ribosome. Identification of Proteins Unique to Th E70 S Ribosome. J. Biol. Chem. 2003, 278, 33774–33785. [Google Scholar] [CrossRef] [Green Version]
- Chang, T.-H.; Tung, L.; Yeh, F.-L.; Chen, J.-H.; Chang, S.-L. Functions of the DExD/H-Box Proteins in Nuclear Pre-MRNA Splicing. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 764–774. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Hill, R.C.; Qiu, Y.; Zhang, W.; Hansen, K.C.; Zhao, R. Brr2 Plays a Role in Spliceosomal Activation in Addition to U4/U6 Unwinding. Nucleic Acids Res. 2015, 43, 3286–3297. [Google Scholar] [CrossRef] [Green Version]
- Cordin, O.; Beggs, J.D. RNA Helicases in Splicing. RNA Biol. 2013, 10, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Jarmoskaite, I.; Russell, R. RNA Helicase Proteins as Chaperones and Remodelers. Annu. Rev. Biochem. 2014, 83, 697–725. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, K.A.; Gallagher, J.E.G.; Mitchell, B.M.; Granneman, S.; Baserga, S.J. The Small-Subunit Processome Is a Ribosome Assembly Intermediate. Eukaryot. Cell 2004, 3, 1619–1626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, R.; Patena, W.; Gang, S.S.; Blum, S.R.; Ivanova, N.; Yue, R.; Robertson, J.M.; Lefebvre, P.A.; Fitz-Gibbon, S.T.; et al. An Indexed, Mapped Mutant Library Enables Reverse Genetics Studies of Biological Processes in Chlamydomonas reinhardtii. Plant Cell 2016, 28, 367–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Liu, G.; Ke, W.; Zhao, L.; Lv, B.; Ma, X.; Xu, N.; Xia, X.; Deng, X.; Zheng, C.; et al. Building a Multipurpose Insertional Mutant Library for Forward and Reverse Genetics in Chlamydomonas. Plant Methods 2017, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Dumas, L.; Zito, F.; Auroy, P.; Johnson, X.; Peltier, G.; Alric, J. Structure-Function Analysis of Chloroplast Proteins via Random Mutagenesis Using Error-Prone PCR. Plant Physiol. 2018, 177, 465–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Patena, W.; Fauser, F.; Jinkerson, R.E.; Saroussi, S.; Meyer, M.T.; Ivanova, N.; Robertson, J.M.; Yue, R.; Zhang, R.; et al. A Genome-Wide Algal Mutant Library and Functional Screen Identifies Genes Required for Eukaryotic Photosynthesis. Nat. Genet. 2019, 51, 627–635. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Functional Domain | Size (kDa) | Involved in Intron Trans-Splicing | References |
|---|---|---|---|---|
| Raa1 | OPR domains | 210 | psaA-i1 psaA-i2 | [111] |
| Raa2 | Pseudouridine synthase | 45 | psaA-i2 | [127] |
| Raa3 | No functional annotation | 180 | psaA-i1 | [126] |
| Raa4 | No functional annotation | 116 | psaA-i1 | [142] |
| Raa6 | No functional annotation | 113 | psaA-i2 | [143] |
| Raa7 | No functional annotation | 130 | psaA-i2 | [144] |
| Raa8 | OPR domains | 269 | psaA-i1 | [123] |
| Rat2 | OPR domains | 144 | psaA-i1 | [123,145] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kück, U.; Schmitt, O. The Chloroplast Trans-Splicing RNA–Protein Supercomplex from the Green Alga Chlamydomonas reinhardtii. Cells 2021, 10, 290. https://doi.org/10.3390/cells10020290
Kück U, Schmitt O. The Chloroplast Trans-Splicing RNA–Protein Supercomplex from the Green Alga Chlamydomonas reinhardtii. Cells. 2021; 10(2):290. https://doi.org/10.3390/cells10020290
Chicago/Turabian StyleKück, Ulrich, and Olga Schmitt. 2021. "The Chloroplast Trans-Splicing RNA–Protein Supercomplex from the Green Alga Chlamydomonas reinhardtii" Cells 10, no. 2: 290. https://doi.org/10.3390/cells10020290
APA StyleKück, U., & Schmitt, O. (2021). The Chloroplast Trans-Splicing RNA–Protein Supercomplex from the Green Alga Chlamydomonas reinhardtii. Cells, 10(2), 290. https://doi.org/10.3390/cells10020290