
Metabolomics Intervention Towards Better Understanding of Plant Traits

 ,
,  ,
, 


 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Integrating Metabolomics with Genomics Study for Gene Characterization and Metabolomics-Assisted Breeding
2.1. Metabolomic Quantitative Trait Loci
2.2. Metabolic Genome-Wide Association Studies
2.3. Metabolic Analysis for Biotic Stress Tolerance in Crop Plants
3. Important Achievements through Metabolic Engineering
3.1. Fortification of Carotenoids and Flavonoids
3.2. Metabolic Engineering of Phytohormone Signaling and Biosynthetic Pathway to Improve Crop Performance
3.3. Engineering of Cell Wall Biosynthesis Pathway: Some Examples
3.4. Metabolic Engineering for Bio-Fortification of Phytonutrients
4. Study of Root Nodule Symbiosis (RNS) in Legumes
5. Addressing Symbiotic Nitrogen Fixation in Cereals and Non-Legume Crop Plants
6. Public Perception for the Metabolic Engineered Plants
7. Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| mQTLs | Metabolic Quantitative Trait Loci |
| mGWAS | Metabolic Genome-Wide Association Studies |
| NMR | Nuclear Magnetic Resonance |
| LC-MS | Liquid Chromatography–Mass Spectrometry |
| GC-MS | Gas Chromatography–Mass Spectrometry |
| PCA | Principal Component Analysis |
| PLS | Partial Least Squares |
| ABRC | ABA-Responsive Complex |
| DW | dry weight |
| FW | fresh weight |
| PSY | phytoene synthase |
| NUE | Nitrogen Use Efficiency |
| SYM | Symbiotic Signaling Pathway |
| AMSA | Arbuscular Mycorrhizal Symbiotic Associations |
| RNS | Root Nodule Symbiosis |
| PDS | phytoenedesaturase |
| LCYB | lycopene β-cyclase |
| HGGT | homogentisategeranylgeranyltransferase |
| DXS | 1-deoxy-D-xylulose-5-phosphate synthase |
| FIB | fibrillin |
| HMGR | 3-hydroxy-3-methylglutaryl-coenzyme A reductase |
| β-CHX | beta-carotene hydroxylase |
| ZDS | zeta-carotene desaturase |
| HYD | carotenoid hydroxylase |
| LCYE | lycopene ɛ-cyclase |
| crtB | phytoene synthase |
| crtI | phytoenedesaturase |
| crtY | lycopene β-cyclase |
| crtE | geranylgeranyldiphosphate synthase |
| crtW | beta-carotene ketolase |
References
- Kumar, R.; Bohra, A.; Pandey, A.K.; Pandey, M.K.; Kumar, A. Metabolomics for plant improvement: Status and prospects. Front. Plant Sci. 2017, 8, 1302. [Google Scholar] [CrossRef] [Green Version]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.T.; Ikeda, K.; Kanazawa, M.; Gheynst, J.S.V.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Xie, L.J.; Chen, Q.F.; Chen, M.X.; Yu, L.J.; Huang, L.; Chen, L.; Wang, F.Z.; Xia, F.N.; Zhu, T.R.; Wu, J.X.; et al. Unsaturation of very-long-chain ceramides protects plant from hypoxia-induced damages by modulating ethylene signaling in Arabidopsis. PLoS Genet. 2015, 11, e1005143. [Google Scholar] [CrossRef] [PubMed]
- Gundaraniya, S.A.; Ambalam, P.S.; Tomar, R.S. Metabolomic Profiling of Drought-Tolerant and Susceptible Peanut (Arachis hypogaea L.) Genotypes in Response to Drought Stress. ACS Omega 2020, 5, 31209–31219. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, Y.H.; Liu, J.X.; Feng, K.; Xu, Z.S.; Xiong, A.S. Advances in genomic, transcriptomic, proteomic, and metabolomic approaches to study biotic stress in fruit crops. Crit. Rev. Biotechnol. 2019, 39, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Sawada, Y.; Ochiai, K.; Sato, M.; Inaba, J.; Hirai, M.Y. Identification of a Unique Type of Isoflavone O-Methyltransferase, GmIOMT1, Based on Multi-Omics Analysis of Soybean under Biotic Stress. Plant Cell Physiol. 2020, 61, 1974–1985. [Google Scholar] [CrossRef]
- Arbona, V.; Manzi, M.; De Ollas, C.; Gómez-Cadenas, A. Metabolomics as a Tool to Investigate Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2013, 14, 4885–4911. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Saito, K. Integrated metabolomics for abiotic stress responses in plants. Curr. Opin. Plant Biol. 2015, 24, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Ding, C.; Li, W.; Wang, D.; Cui, D. Applications of metabolomics in the research of soybean plant under abiotic stress. Food Chem. 2020, 310, 125914. [Google Scholar] [CrossRef]
- Gong, L.; Chen, W.; Gao, Y.; Liu, X.; Zhang, H.; Xu, C.; Yu, S.; Zhang, Q.; Luo, J. Genetic analysis of the metabolome exemplified using a rice population. Proc. Natl. Acad. Sci. USA 2013, 110, 20320–20325. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Shi, J.; Quan, S.; Cui, B.; Kleessen, S.; Nikoloski, Z.; Tohge, T.; Alexander, D.; Guo, L.; Lin, H.; et al. Metabolic variation between japonica and indica rice cultivars as revealed by non-targeted metabolomics. Sci. Rep. 2014, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Tohge, T.; Chan, S.-A.; Song, Y.; Rao, J.; Cui, B.; Lin, H.; Wang, L.; Fernie, A.R.; Zhang, D.; et al. Identification of Conserved and Diverse Metabolic Shifts during Rice Grain Development. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef]
- Kusano, M.; Yang, Z.; Okazaki, Y.; Nakabayashi, R.; Fukushima, A.; Saito, K. Using metabolomic approaches to explore chemical diversity in rice. Mol. Plant. 2015, 8, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Muscolo, A.; Junker, A.; Klukas, C.; Weigelt-Fischer, K.; Riewe, D.; Altmann, T. Phenotypic and metabolic responses to drought and salinity of four contrasting lentil accessions. J. Exp. Bot. 2015, 66, 5467–5480. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.; Rabara, R.C.; Shulaev, V.; Shen, Q.J.; Rushton, P.J. Understanding Water-Stress Responses in Soybean Using Hydroponics System—A Systems Biology Perspective. Front. Plant Sci. 2015, 6, 1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Wang, W.; Peng, M.; Gong, L.; Gao, Y.; Wan, J.; Wang, S.; Shi, L.; Zhou, B.; Li, Z.; et al. Comparative and parallel genome-wide association studies for metabolic and agronomic traits in cereals. Nat. Commun. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, Y.; Saito, K. Integrated metabolomics and phytochemical genomics approaches for studies on rice. GigaScience 2016, 5, 13742–137016. [Google Scholar] [CrossRef] [Green Version]
- Kosma, D.K.; Parsons, E.P.; Isaacson, T.; Lü, S.; Rose, J.K.C.; Jenks, M.A. Fruit cuticle lipid composition during development in tomato ripening mutants. Physiol. Plant. 2010, 139, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Burgos, A.; Szymanski, J.; Seiwert, B.; Degenkolbe, T.; Hannah, M.A.; Giavalisco, P.; Willmitzer, L. Analysis of short-term changes in the Arabidopsis thaliana glycerolipidome in response to temperature and light. Plant J. 2011, 66, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Hou, Q.; Ufer, G.; Bartels, D. Lipid signalling in plant responses to abiotic stress. Plant Cell Environ. 2016, 39, 1029–1048. [Google Scholar] [CrossRef]
- Tenenboim, H.; Burgos, A.; Willmitzer, L.; Brotman, Y. Using lipidomics for expanding the knowledge on lipid metabo-lism in plants. Biochimie 2016, 130, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Idso, S.B.; Idso, K.E. Effects of atmospheric CO2 enrichment on plant constituents related to animal and human health. Environ. Exp. Bot. 2001, 45, 179–199. [Google Scholar] [CrossRef]
- Högy, P.; Wieser, H.; Köhler, P.; Schwadorf, K.; Breuer, J.; Franzaring, J.; Muntifering, R.; Fangmeier, A. Effects of ele-vated CO2 on grain yield and quality of wheat: Results from a 3-year free-air CO2 enrichment experiment. Plant Biol. 2009, 11, 60–69. [Google Scholar] [CrossRef]
- Pal, M.; Chaturvedi, A.K.; Pandey, S.K.; Bahuguna, R.N.; Khetarpal, S.; Anand, A. Rising atmospheric CO2 may affect oil quality and seed yield of sunflower (Helianthus annuus L.). Acta Physiol. Plant. 2014, 36, 2853–2861. [Google Scholar] [CrossRef]
- Reich, M.; van den Meerakker, A.N.; Parmar, S.; Hawkesford, M.J.; De Kok, L.J. Temperature determines size and direc-tion of effects of elevated CO2 and nitrogen form on yield quantity and quality of Chinese cabbage. Plant Biol. 2016, 18, 63–75. [Google Scholar] [CrossRef]
- Kumar, R.; Tamboli, V.; Sharma, R.; Sreelakshmi, Y. NAC-NOR mutations in tomato Penjar accessions attenuate multiple metabolic processes and prolong the fruit shelf life. Food Chem. 2018, 259, 234–244. [Google Scholar] [CrossRef]
- Toubiana, D.; Semel, Y.; Tohge, T.; Beleggia, R.; Cattivelli, L.; Rosental, L.; Nikoloski, Z.; Zamir, D.; Fernie, A.R.; Fait, A. Metabolic Profiling of a Mapping Population Exposes New Insights in the Regulation of Seed Metabolism and Seed, Fruit, and Plant Relations. PLoS Genet. 2012, 8, e1002612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ainalidou, A.; Tanou, G.; Belghazi, M.; Samiotaki, M.; Diamantidis, G.; Molassiotis, A.; Karamanoli, K. Integrated analy-sis of metabolites and proteins reveal aspects of the tissue-specific function of synthetic cytokinin in kiwifruit development and ripening. J. Proteom. 2015, 143, 318–333. [Google Scholar] [CrossRef]
- Upadhyaya, P.; Tyagi, K.; Sarma, S.; Tamboli, V.; Sreelakshmi, Y.; Sharma, R. Natural variation in folate levels among tomato (Solanum lycopersicum) accessions. Food Chem. 2017, 217, 610–619. [Google Scholar] [CrossRef]
- Horgan, R.P.; Kenny, L.C. ‘Omic’ technologies: Genomics, transcriptomics, proteomics and metabolomics. Obstet. Gynaecol. 2011, 13, 189–195. [Google Scholar] [CrossRef]
- Raja, K.; Patrick, M.; Gao, Y.; Madu, D.; Yang, Y.; Tsoi, L.C. A Review of Recent Advancement in Integrating Omics Data with Literature Mining towards Biomedical Discoveries. Int. J. Genom. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Guijas, C.; Montenegro-Burke, J.R.; Warth, B.; Spilker, M.E.; Siuzdak, G. Metabolomics activity screening foridentifying metabolites that modulate phenotype. Nat. Biotechnol. 2018, 36, 316–320. [Google Scholar] [CrossRef]
- Fernie, A.R.; Schauer, N. Metabolomics-assisted breeding: A viable option for crop improvement? Trends Genet. 2009, 25, 39–48. [Google Scholar] [CrossRef]
- Chen, W.; Gao, Y.; Xie, W.; Gong, L.; Lu, K.; Wang, W.; Li, Y.; Liu, X.; Zhang, H.; Dong, H.; et al. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Genet. 2014, 46, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Li, K.; Alseekh, S.; Omranian, N.; Zhao, L.; Zhou, Y.; Xiao, Y.; Jin, M.; Yang, N.; Liu, H.; et al. Genetic determi-nants of the network of primary metabolism and their relationships to plant performance ina maize recombinant in-bred line population. Plant Cell 2015, 27, 1839–1856. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, O.; Urrutia, M.; Bernillon, S.; Giauffret, C.; Tardieu, F.; Le Gouis, J.; Langlade, N.; Charcosset, A.; Moing, A.; Gibon, Y. Fortune telling: Metabolic markers of plant performance. Metabolomics 2016, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.; Liu, H.; Zhou, Y.; Jin, M.; Yang, N.; Li, D.; Luo, J.; Xiao, Y.; Pan, Q.; Tohge, T.; et al. Combining Quantitative Genetics Approaches with Regulatory Network Analysis to Dissect the Complex Metabolism of the Maize Kernel. Plant Physiol. 2016, 170, 136–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scossa, F.; Brotman, Y.; Lima, F.D.A.E.; Willmitzer, L.; Nikoloski, Z.; Tohge, T.; Fernie, A.R. Genomics-based strategies for the use of natural variation in the improvement of crop metabolism. Plant Sci. 2016, 242, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Beleggia, R.; Rau, D.; Laidò, G.; Platani, C.; Nigro, F.; Fragasso, M.; De Vita, P.; Scossa, F.; Fernie, A.R.; Nikoloski, Z.; et al. Evolutionary Metabolomics Reveals Domestication-Associated Changes in Tetraploid Wheat Kernels. Mol. Biol. Evol. 2016, 33, 1740–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, F.; Okazaki, Y.; Oikawa, A.; Kusano, M.; Nakabayashi, R.; Kikuchi, J.; Yonemaru, J.I.; Ebana, K.; Yano, M.; Saito, K. Dissection of genotype–phenotype associations in rice grains using metabolomequantitative trait loci analysis. Plant J. 2012, 70, 624–636. [Google Scholar] [CrossRef]
- Riedelsheimer, C.; Lisec, J.; Czedik-Eysenberg, A.; Sulpice, R.; Flis, A.; Grieder, C.; Altmann, T.; Stitt, M.; Willmitzer, L.; Melchinger, A.E. Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize. Proc. Natl. Acad. Sci. USA 2012, 109, 8872–8877. [Google Scholar] [CrossRef] [Green Version]
- Piasecka, A.; Sawikowska, A.; Kuczynska, A.; Ogrodowicz, P.; Mikolajczak, K.; Krystkowiak, K.; Gudys, K.; Guzy-Wrobel-ska, J.; Krajewski, P.; Kachlicki, P. Drought-related econdary metabolites of barley (Hordeum vulgare L.) leaves and their metabolomic quantitative trait loci. Plant J. 2017, 89, 898–913. [Google Scholar] [CrossRef] [Green Version]
- Templer, S.E.; Ammon, A.; Pscheidt, D.; Ciobotea, O.; Schuy, C.; McCollum, C.; Sonnewald, U.; Hanemann, A.; Förster, J.; Ordon, F.; et al. Metabolite profiling of barley flag leaves under drought and combined heat anddrought stress reveals metabolic QTLs for metabolites associated with antioxidant defense. J. Exp. Bot. 2017, 68, 1697–1713. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Long, Y.; Shi, L.; Shi, J.; Barker, G.; Meng, J. Characterization of metabolite quantitative trait loci and metabolic networks that control glucosinolate concentration in the seeds and leaves of Brassica napus. New Phytol. 2011, 193, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Alseekh, S.; Tohge, T.; Wendenberg, R.; Scossa, F.; Omranian, N.; Tzili, P.; Kleessen, S.; Giavalisco, P.; Pleban, T.; Mueller-Roeber, B.; et al. Identification and Mode of Inheritance of Quantitative Trait Loci for Secondary Metabolite Abundance in Tomato. Plant Cell 2015, 27, 485–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alseekh, S.; Tong, H.; Scossa, F.; Brotman, Y.; Vigroux, F.; Tohge, T.; Ofner, I.; Zamir, D.; Nikoloski, Z.; Fernie, A.R. Can-alization of tomato fruit metabolism. Plant Cell 2017, 29, 2753–2765. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.B.; Taylor, J.D.; Edwards, J.; Mather, D.; Langridge, P.; Bacic, A.; Roessner, U. Detection of QTL for metabolic and agronomic traits in wheat with adjustments for variation at genetic loci that affect plant phenology. Plant Sci. 2015, 233, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Kazmi, R.H.; Willems, L.A.J.; Joosen, R.V.L.; Khan, N.; Ligterink, W.; Hilhorst, H.W.M. Metabolomic analysis of tomato seed germination. Metabolomics 2017, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, F.; Bradbury, P.J.; Brown, P.J.; Hung, H.; Sun, Q.; Flintgarcia, S.A.; Rocheford, T.R.; McMullen, M.D.; Holland, J.B.; Buckler, E.S. Genome-wide association study of leaf architecture in the maize nested association mapping population. Nat. Genet. 2011, 43, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Li, D.; Li, X.; Gao, Y.; Li, W.; Li, H.; Liu, J.; Liu, H.; Chen, W.; Luo, J. Metabolome-based genome-wide associa-tion study of maize kernel leads to novel biochemical insights. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Gao, Y.; Chen, W.; Wang, W.; Gong, L.; Liu, X.; Luo, J. Spatiotemporal distribution of phenolamides and the genetics of natural variation of hydroxycinnamoyl spermidine in rice. Mol. Plant 2015, 8, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sotelo, T.; Soengas, P.; Velasco, P.; Rodríguez, V.M.; Cartea, M.E. Identification of metabolic QTLs and candidate genes for glucosinolate synthesis in Brassica oleracealeaves, seeds and flower buds. PLoS ONE 2014, 9, e91428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, M.; Shahzad, R.; Gul, A.; Subthain, H.; Shen, S.; Lei, L.; Zheng, Z.; Zhou, J.; Lu, D.; Wang, S. Differentially evolved glucosyl transferases determine natural variation of rice flavone accumulation and UV-tolerance. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Nakabayashi, R.; Yang, Z.; Okazaki, Y.; Yonemaru, J.; Ebana, K.; Yano, M.; Saito, K. Metabo-lome-genome-wide association study (mGWAS) dissects genetic architecture for generating natural variation in rice secondary metabolism. Plant J. 2015, 81, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Matros, A.; Liu, G.; Hartmann, A.; Jiang, Y.; Zhao, Y.; Wang, H.; Ebmeyer, E.; Korzun, V.; Schachschneider, R.; Kazman, E.; et al. Genome–metabolite associations revealed low heritability, high genetic complexity, and causal relations for leaf metabolites in winter wheat (Triticum aestivum). J. Exp. Bot. 2016, 68, 415–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Zhu, A.; Jia, J.; Hu, X.; Chen, J.; Liu, W.; Ren, X.; Sun, D.; Fernie, A.R.; Cui, F.; et al. Metabolomics analysis and metabolite-agronomic trait associations using kernels of wheat (Triticum aestivum) recombinant inbred lines. Plant J. 2020, 103, 279–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Peng, Z.; Yang, X.; Wang, W.; Fu, J.; Wang, J.; Han, Y.; Chai, Y.; Guo, T.; Yang, N.; et al. Genome-wide association study dissects the genetic architecture of oil biosynthesis in maize kernels. Nat. Genet. 2013, 45, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Lipka, A.E.; Gore, M.A.; Magallanes-Lundback, M.; Mesberg, A.; Lin, H.; Tiede, T.; Chen, C.; Buell, C.R.; Buckler, E.S.; Rocheford, T.; et al. Genome-Wide Association Study and Pathway-Level Analysis of Tocochromanol Levels in Maize Grain. G3 Genes Genomes Genet. 2013, 3, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Owens, B.F.; Lipka, A.E.; Magallanes-Lundback, M.; Tiede, T.; Diepenbrock, C.H.; Kandianis, C.B.; Kim, E.; Cepela, J.; Mateos-Hernandez, M.; Buell, C.R.; et al. A foundation for provitamin A biofortification of maize: Genome-wide associ-ation and genomic prediction models of carotenoid levels. Genetics 2014, 198, 1699–1716. [Google Scholar] [CrossRef] [Green Version]
- Sauvage, C.; Segura, V.; Bauchet, G.; Stevens, R.; Do, P.T.; Nikoloski, Z.; Fernie, A.R.; Causse, M. Genome-wideassociation in tomato reveals 44 candidate loci for fruit metabolic traits. Plant Physiol. 2014, 165, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Solomon, P.S.; Tan, K.-C.; Oliver, R.P. The nutrient supply of pathogenic fungi; a fertile field for study. Mol. Plant Pathol. 2003, 4, 203–210. [Google Scholar] [CrossRef]
- Divon, H.H.; Fluhr, R. Nutrition acquisition strategies during fungal infection of plants. FEMS Microbiol. Lett. 2007, 266, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Caldo, R.A.; Nettleton, D.; Wise, R.P. Interaction-dependent gene expression in Mla-specified response to barley pow-dery mildew. Plant Cell 2004, 16, 2514–2528. [Google Scholar] [CrossRef] [Green Version]
- Both, M.; Csukai, M.; Stumpf, M.P.; Spanu, P.D. Gene Expression Profiles of Blumeria graminis Indicate Dynamic Changes to Primary Metabolism during Development of an Obligate Biotrophic Pathogen. Plant Cell 2005, 17, 2107–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doehlemann, G.; Wahl, R.; Horst, R.J.; Voll, L.M.; Usadel, B.; Poree, F.; Stitt, M.; Pons-Kühnemann, J.; Sonnewald, U.; Kahmann, R.; et al. Reprogramming a maize plant: Transcriptional and metabolic changes induced by the fungal biotroph Ustilago maydis. Plant J. 2008, 56, 181–195. [Google Scholar] [CrossRef]
- Shi, H.; Ye, T.; Zhong, B.; Liu, X.; Chan, Z. Comparative proteomic and metabolomic analyses reveal mechanisms of improved cold stress tolerance in bermudagrass (Cynodon dactylon (L.) Pers.) by exogenous calcium. J. Integr. Plant Biol. 2014, 56, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Jorge, T.F.; Rodrigues, J.A.; Caldana, C.; Schmidt, R.; van Dongen, J.T.; Thomas-Oates, J.; António, C. Mass spectrometry-based plant metabolomics: Metabolite responses to abiotic stress. Mass Spectrom. Rev. 2016, 35, 620–649. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Shahid, M.A.; Kharabian-Masouleh, A. Advances in detection of stress tolerance in plants through metabolomics approaches. Plant Omics 2017, 10, 153–163. [Google Scholar] [CrossRef]
- Moradi, P.; Ford-Lloyd, B.; Pritchard, J. Metabolomic approach reveals the biochemical mechanisms underlying drought stress tolerance in thyme. Anal. Biochem. 2017, 527, 49–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Xin, Z.; Yang, T.; Ma, X.; Zhang, Y.; Wang, Z.; Ren, Y.; Lin, T. Metabolomics response for drought stress toler-ance in chinese wheat genotypes (Triticum aestivum). Plants 2020, 9, 520. [Google Scholar] [CrossRef] [Green Version]
- Min, K.; Chen, K.; Arora, R. A metabolomics study of ascorbic acid-induced in situ freezing tolerance in spinach (Spinacia oleracea L.). Plant Direct 2020, 4, e00202. [Google Scholar] [CrossRef] [Green Version]
- Danan, S.; Veyrieras, J.B.; Lefebvre, V. Construction of a potato consensus map and QTL meta-analysis offer new in-sights into the genetic architecture of late blight resistance and plant maturity traits. BMC Plant Biol. 2011, 11, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ballini, E.; Morel, J.-B.; Droc, G.; Price, A.; Courtois, B.; Notteghem, J.-L.; Tharreau, D. A Genome-Wide Meta-Analysis of Rice Blast Resistance Genes and Quantitative Trait Loci Provides New Insights into Partial and Complete Resistance. Mol. Plant-Microbe Interact. 2008, 21, 859–868. [Google Scholar] [CrossRef]
- Qi, X.; Niks, R.E.; Stam, P.; Lindhout, P. Identification of QTLs for partial resistance to leaf rust (Puccinia hordei) in barley. Theor. Appl. Genet. 1998, 96, 1205–1215. [Google Scholar] [CrossRef]
- Lemmens, M.; Scholz, U.; Berthiller, F.; Dall’Asta, C.; Koutnik, A.; Schuhmacher, R.; Adam, G.; Buerstmayr, H.; Mesterházy, Á.; Krska, R.; et al. The Ability to Detoxify the Mycotoxin Deoxynivalenol Colocalizes with a Major Quantitative Trait Locus for Fusarium Head Blight Resistance in Wheat. Mol. Plant-Microbe Interact. 2005, 18, 1318–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunnaiah, R.; Kushalappa, A.C.; Duggavathi, R.; Fox, S.; Somers, D.J. Integrated metabolo-proteomic approach to deci-pher the mechanisms by which wheat QTL (Fhb1) contributes to resistance against Fusarium graminearum. PLoS ONE 2012, 7, e40695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sana, T.R.; Fischer, S.; Wohlgemuth, G.; Katrekar, A.; Jung, K.-H.; Ronald, P.C.; Fiehn, O. Metabolomic and transcriptomic analysis of the rice response to the bacterial blight pathogen Xanthomonas oryzae pv. oryzae. Metabolomics 2010, 6, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Spencer, P.A.; Towers, G. Restricted occurrence of acetophenone signal compounds. Phytochemistry 1991, 30, 2933–2937. [Google Scholar] [CrossRef]
- Luo, H.-Z.; Guan, Y.; Yang, R.; Qian, G.-L.; Yang, X.-H.; Wang, J.-S.; Jia, A.-Q. Growth inhibition and metabolomic analysis of Xanthomonas oryzae pv. oryzae treated with resveratrol. BMC Microbiol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Koduru, L.; Kim, H.Y.; Lakshmanan, M.; Mohanty, B.; Lee, Y.Q.; Lee, C.H.; Lee, D.Y. Genome-scale metabolic reconstruc-tion and in silico analysis of the rice leaf blight pathogen, Xanthomonas oryzae. Mol. Plant Pathol. 2020, 21, 527–540. [Google Scholar] [CrossRef] [Green Version]
- Agarrwal, R.; Bentur, J.S.; Nair, S. Gas chromatography mass spectrometry based metabolic profiling reveals bi-omarkers involved in rice-gall midge interactions. J. Integr. Plant Biol. 2014, 56, 837–848. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Li, Y.; Zhang, J.; Xiao, Y.; Yue, Y.; Duan, L.; Zhang, M.; Li, Z. Overexpression of Arabidopsis molybdenum cofac-tor sulfurase gene confers drought tolerance in maize (Zea mays L.). PLoS ONE 2013, 8, e52126. [Google Scholar]
- Zhang, J.; Yu, H.; Zhang, Y.; Wang, Y.; Li, M.; Zhang, J.; Duan, L.; Zhang, M.; Li, Z. Increased abscisic acid levels in transgenic maize overexpressing AtLOS5 mediated root ion fluxes and leaf water status under salt stress. J. Exp. Bot. 2016, 67, 1339–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, Y.; Chang, L.; Zhang, T.; An, J.; Liu, Y.; Cao, Y.; Zhao, X.; Sha, X.; Hu, T.; et al. MsZEP, a novel ze-axanthin epoxidase gene from alfalfa (Medicago sativa), confers drought and salt tolerance in transgenic tobacco. Plant Cell Rep. 2016, 14, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Zhang, H.; Tian, S.; Chang, X.; Jing, R. TaSnRK2.4, an SNF1-type serine/threonine protein kinase of wheat (Trit-icum aestivum L.), confers enhanced multi stress tolerance in Arabidopsis. J. Exp. Bot. 2010, 61, 683–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.I.; Baek, D.; Park, H.C.; Chun, H.J.; Oh, D.H.; Lee, M.K.; Cha, J.Y.; Kim, W.Y.; Kim, M.C.; Chung, W.S.; et al. Overexpression of Arabidopsis YUCCA6 in potato results in high-auxin developmental phenotypes and enhanced re-sistance to water deficit. Mol. Plant 2013, 6, 337–349. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Lee, D.-K.; Choi, Y.D.; Kim, J.-K. OsIAA6, a member of the rice Aux/IAA gene family, is involved in drought tolerance and tiller outgrowth. Plant Sci. 2015, 236, 304–312. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Albacete, A.; Smigocki, A.C.; Frébort, I.; Pospíšilová, H.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Lutts, S.; Dodd, I.C.; et al. Root-synthesized cytokinins improve shoot growth and fruit yield in sali-nized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2011, 62, 125–140. [Google Scholar] [CrossRef] [PubMed]
- Werner, T.; Nehnevajova, E.; Köllmer, I.; Novák, O.; Strnad, M.; Krämer, U.; Schmülling, T. Root-Specific Reduction of Cytokinin Causes Enhanced Root Growth, Drought Tolerance, and Leaf Mineral Enrichment in Arabidopsis and Tobacco. Plant Cell 2010, 22, 3905–3920. [Google Scholar] [CrossRef] [Green Version]
- Pospíšilová, H.; Jiskrová, E.; Vojta, P.; Mrízová, K.; Kokáš, F.; Čudejková, M.M.; Bergougnoux, V.; Plíhal, O.; Klimešová, J.; Novák, O.; et al. Transgenic barley overexpressing a cytokinin dehydrogenase gene shows greater tolerance to drought stress. New Biotechnol. 2016, 33, 692–705. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, F.; Li, D.; Zhang, H.; Huang, R. Expression of ethylene response factor JERF1 in rice improves tolerance to drought. Planta 2010, 232, 765–774. [Google Scholar] [CrossRef]
- Habben, J.E.; Bao, X.; Bate, N.J.; De Bruin, J.L.; Dolan, D.; Hasegawa, D.; Helentjaris, T.G.; Lafitte, H.R.; Lovan, N.; Mo, H.; et al. Transgenic alteration of ethylene biosynthesis increases grain yield in maize under field drought-stress conditions. Plant Biotechnol. J. 2014, 12, 685–693. [Google Scholar] [CrossRef]
- Shi, J.; Habben, J.E.; Archibald, R.L.; Drummond, B.J.; Chamberlin, M.A.; Williams, R.W.; Lafitte, H.R.; Weers, B.P. Overexpression of ARGOS Genes Modifies Plant Sensitivity to Ethylene, Leading to Improved Drought Tolerance in Both Arabidopsis and Maize. Plant Physiol. 2015, 169, 266–282. [Google Scholar] [CrossRef]
- Koh, S.; Lee, S.-C.; Kim, M.-K.; Koh, J.H.; Lee, S.; An, G.; Choe, S.; Kim, S.-R. T-DNA tagged knockout mutation of rice OsGSK1, an orthologue of Arabidopsis BIN2, with enhanced tolerance to various abiotic stresses. Plant Mol. Biol. 2007, 65, 453–466. [Google Scholar] [CrossRef]
- Li, F.; Asami, T.; Wu, X.; Tsang, E.W.; Cutler, A.J. A Putative Hydroxysteroid Dehydrogenase Involved in Regulating Plant Growth and Development. Plant Physiol. 2007, 145, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Yin, Y.; Fei, S. Down-regulation of BdBRI1, a putative brassinosteroid receptor gene produces a dwarf phenotype with enhanced drought tolerance in Brachypodium distachyon. Plant Sci. 2015, 234, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Kong, W.; Wong, G.; Fu, L.; Peng, R.; Li, Z.; Yao, Q. AtMYB12 regulates flavonoids accumulation and abiotic stress tolerance in transgenic Arabidopsis thaliana. Mol. Genet. Genom. 2016, 291, 1545–1559. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Ahn, Y.O.; Ahn, M.-J.; Lee, H.-S.; Kwak, S.-S. Down-regulation of β-carotene hydroxylase increases β-carotene and total carotenoids enhancing salt stress tolerance in transgenic cultured cells of sweetpotato. Phytochemistry 2012, 74, 69–78. [Google Scholar] [CrossRef]
- Nakabayashi, R.; Yonekura-Sakakibara, K.; Urano, K.; Suzuki, M.; Yamada, Y.; Nishizawa, T.; Matsuda, F.; Kojima, M.; Sakakibara, H.; Shinozaki, K.; et al. Enhancement of oxidative and drought tolerance in Arabidopsis by over accumulation of antioxidant flavonoids. Plant J. 2014, 77, 367–379. [Google Scholar] [CrossRef]
- Shi, Y.; Guo, J.; Zhang, W.; Jin, L.; Liu, P.; Chen, X.; Li, F.; Wei, P.; Li, Z.; Li, W.; et al. Cloning of the Lycopene β-cyclase Gene in Nicotiana tabacum and Its Overexpression Confers Salt and Drought Tolerance. Int. J. Mol. Sci. 2015, 16, 30438–30457. [Google Scholar] [CrossRef]
- Chen, X.; Han, H.; Jiang, P.; Nie, L.; Bao, H.; Fan, P.; Lv, S.; Feng, J.; Li, Y. Transformation of b-lycopene cyclase genes from Salicornia europaea and Arabidopsis conferred salt tolerance in Arabidopsis and Tobacco. Plant Cell Physiol. 2011, 52, 909–921. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; He, S.; Liu, D.; Patil, G.B.; Zhai, H.; Wang, F.; Stephenson, T.J.; Wang, Y.; Wang, B.; Valliyodan, B.; et al. A Sweetpotato Geranylgeranyl Pyrophosphate Synthase Gene, IbGGPS, Increases Carotenoid Content and Enhances Osmotic Stress Tolerance in Arabidopsis thaliana. PLoS ONE 2015, 10, e0137623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [Green Version]
- Głowacka, K.; Kromdijk, J.; Kucera, K.; Xie, J.; Cavanagh, A.P.; Leonelli, L.; Leakey, A.D.B.; Ort, D.R.; Niyogi, K.K.; Long, S.P. Photosystem II Subunit S overexpression increases the efficiency of water use in a field-grown crop. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Han, Y.; Liu, G.; An, B.; Yang, J.; Yang, G.; Li, Y.; Zhu, Y. Overexpression of sedoheptulose-1,7-bisphosphatase enhances photosynthesis and growth under salt stress in transgenic rice plants. Funct. Plant Biol. 2007, 34, 822–834. [Google Scholar] [CrossRef]
- Simkin, A.J.; McAusland, L.; Headland, L.R.; Lawson, T.; Raines, C.A. Multigene manipulation of photosynthetic carbon assimilation increases CO2 fixation and biomass yield in tobacco. J. Exp. Bot. 2015, 66, 4075–4090. [Google Scholar] [CrossRef] [PubMed]
- Driever, S.M.; Simkin, A.J.; Alotaibi, S.; Fisk, S.J.; Madgwick, P.J.; Sparks, C.A.; Jones, H.D.; Lawson, T.; Parry, M.A.J.; Raines, C.A. Increased SBPase activity improves photosynthesis and grain yield in wheat grown in greenhouse conditions. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160384. [Google Scholar] [CrossRef] [Green Version]
- López-Calcagno, P.E.; Fisk, S.J.; Brown, K.; Bull, S.E.; South, P.F.; Raines, C.A. Overexpressing the H-protein of the gly-cine cleavage system increases biomass yield in glasshouse and field grown transgenic tobacco plants. Plant Biotechnol. J. 2018, 17, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Timm, S.; Wittmiß, M.; Gamlien, S.; Ewald, R.; Florian, A.; Frank, M.; Wirtz, M.; Hell, R.; Fernie, A.R.; Bauwe, H. Mitochondrial dihydrolipoyl dehydrogenase activity shapes photosynthesis and photorespiration of Arabidopsis thaliana. Plant Cell 2015, 27, 1968–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timm, S.; Giese, J.; Engel, N.; Wittmiß, M.; Florian, A.; Fernie, A.R.; Bauwe, H. T-protein is present in large excess over the other proteins of the glycine cleavage system in leaves of Arabidopsis. Planta 2017, 247, 41–51. [Google Scholar] [CrossRef]
- Chida, H.; Nakazawa, A.; Akazaki, H.; Hirano, T.; Suruga, K.; Ogawa, M.; Satoh, T.; Kadokura, K.; Yamada, S.; Hakamata, W.; et al. Expression of the Algal Cytochrome c6 Gene in Arabidopsis Enhances Photosynthesis and Growth. Plant Cell Physiol. 2007, 48, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Khatri, K.; Rathore, M.S.; Jha, B. Introgression of UfCyt c6, a thylakoid lumen protein from a green sea-weed Ulva fasciata Delile enhanced photosynthesis and growth in tobacco. Mol. Biol. Rep. 2018, 45, 1745–1758. [Google Scholar] [CrossRef]
- Simkin, A.J.; McAusland, L.; Lawson, T.; Raines, C.A. Overexpression of the RieskeFeS Protein Increases Electron Transport Rates and Biomass Yield. Plant Physiol. 2017, 175, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Lieman-Hurwitz, J.; Rachmilevitch, S.; Mittler, R.; Marcus, Y.; Kaplan, A. Enhanced photosynthesis and growth of trans-genic plants that express ictB, a gene involved in HCO3–accumulation in cyanobacteria. Plant Biotechnol. J. 2003, 1, 43–50. [Google Scholar] [CrossRef]
- Lieman-Hurwitz, J.; Asipov, L.; Rachmilevitch, S.; Marcus, Y.; Kaplan, A. Expression of cyanobacterial ictB in higher plants enhanced photosynthesis and growth. In Plant Responses to Air Pollution and Global Change; Omasa, K., Nouchi, I., De Kok, L.J., Eds.; Springer: Tokyo, Japan, 2005; pp. 133–139. [Google Scholar]
- Gong, H.Y.; Li, Y.; Fang, G.; Hu, D.H.; Jin, W.B.; Wang, Z.H.; Li, Y.S. Transgenic rice expressing IctB and FBP/SBPase de-rived from cyanobacteria exhibits enhanced photosynthesis and mesophyll conductance to CO2. PLoS ONE 2015, 10, e0140928. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Du, H.; Wang, J.; Pu, Y.; Yang, C.; Yan, R.; Yang, H.; Cheng, H.; Yu, D. Multiplex CRISPR/Cas9-mediated met-abolic engineering increases soya bean isoflavone content and resistance to soya bean mosaic virus. Plant Biotechnol. J. 2020, 18, 1384–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, A.; Chen, H.; Chen, L.; Chen, S.; Hao, Q.; Guo, W.; Qiu, D.; Shan, Z.; Yang, Z.; Yuan, S.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmSPL9 genes alters plant architecture in soybean. BMC Plant Biol. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, Y.; Chen, S.; Tian, H.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. Lycopene Is Enriched in Tomato Fruit by CRISPR/Cas9-Mediated Multiplex Genome Editing. Front. Plant Sci. 2018, 9, 559. [Google Scholar] [CrossRef]
- Lou, D.; Wang, H.; Liang, G.; Yu, D. OsSAPK2 Confers Abscisic Acid Sensitivity and Tolerance to Drought Stress in Rice. Front. Plant Sci. 2017, 8, 993. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Gao, H.; Wang, H.; Lafitte, H.R.; Archibald, R.L.; Yang, M.; Hakimi, S.M.; Mo, H.; Habben, J.E. ARGOS8 variants generated by CRISPR-Cas9 improve maize grain yield under field drought stress conditions. Plant Biotechnol. J. 2016, 15, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, L.; Li, R.; Zhao, R.; Yang, M.; Sheng, J.; Shen, L. Reduced Drought Tolerance by CRISPR/Cas9-Mediated SlMAPK3 Mutagenesis in Tomato Plants. J. Agric. Food Chem. 2017, 65, 8674–8682. [Google Scholar] [CrossRef] [PubMed]
- Paine, J.A.; Shipton, C.A.; Chaggar, S.; Howells, R.M.; Kennedy, M.J.; Vernon, G.; Wright, S.Y.; Hinchliffe, E.; Adams, J.L.; Silverstone, A.L.; et al. Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nat. Biotechnol. 2005, 23, 482. [Google Scholar] [CrossRef]
- Cong, L.; Wang, C.; Chen, L.; Liu, H.; Yang, G.; He, G. Expression of phytoene synthase1 and carotene desaturase crtI genes result in an increase in the total carotenoids content in transgenic elite wheat (Triticum aestivum L.). J. Agric. Food Chem. 2009, 57, 8652–8660. [Google Scholar] [CrossRef]
- Goto, F.; Yoshihara, T.; Shigemoto, N.; Toki, S.; Takaiwa, F. Iron fortification of rice seed by the soybean ferritin gene. Nat. Biotechnol. 1999, 17, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.A.T.; Kyriacou, B.; Callahan, D.L.; Carruthers, L.; Stangoulis, J.; Lombi, E.; Tester, M. Constitutive overex-pression of the OsNAS gene family reveals single gene strategies for effective iron- and zinc-biofortification of rice en-dosperm. PLoS ONE 2011, 6, e24476. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Usuda, K.; Kobayashi, T.; Ishimaru, Y.; Kakei, Y.; Takahashi, M.; Higuchi, K.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Overexpression of the Barley Nicotianamine Synthase Gene HvNAS1 Increases Iron and Zinc Concentrations in Rice Grains. Rice 2009, 2, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yin, M.; Li, Y.; Fan, C.; Yang, Q.; Wu, J.; Zhang, C.; Wang, H.; Zhou, Y. Expression of Brassica napus TTG2, a regulator of trichome development, increases plant sensitivity to salt stress by suppressing the expression of auxin biosynthesis genes. J. Exp. Bot. 2015, 66, 5821–5836. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Fu, S.; Chen, S.; Zhang, W.; Qi, C. Ethylene response factor BnERF2-like (ERF2.4) from Brassica napus L. enhances submergence tolerance and alleviates oxidative damage caused by submergence in Arabidopsis thaliana. Crop J. 2016, 4, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Guo, X.; Wang, C.; Ma, J.; Niu, F.; Zhang, H.; Yang, B.; Liang, W.; Han, F.; Jiang, Y.Q. Identification and charac-terization of plant-specific NAC gene family in canola (Brassica napus L.) reveal novel members involved in cell death. Plant Mol. Biol. 2015, 87, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Liu, X.; Xue, H.; Li, X.; Wang, X. Functional characterization of BnHSFA4a as a heat shock transcription factor in controlling the re-establishment of desiccation tolerance in seeds. J. Exp. Bot. 2017, 68, 2361–2375. [Google Scholar] [CrossRef]
- Xu, J.; Dai, H. Brassica napus Cycling Dof Factor1 (BnCDF1) is involved in flowering time and freezing tolerance. Plant Growth Regul. 2016, 80, 315–322. [Google Scholar] [CrossRef]
- Li, L.; Ye, C.; Zhao, R.; Li, X.; Liu, W.Z.; Wu, F.; Yan, J.; Jiang, Y.Q.; Yang, B. Mitogen-activated protein kinase kinase kinase (MAPKKK) 4 from rapeseed (Brassica napus L.) is a novel member inducing ROS accumulation and cell death. Biochem. Biophys. Res. Commun. 2015, 467, 792–797. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, C.; Yang, B.; Wu, F.; Hao, X.; Liang, W.; Niu, F.; Yan, J.; Zhang, H.; Wang, B.; et al. Identification and functional analysis of mitogen-activated protein kinase kinase kinase (MAPKKK) genes in canola (Brassica napus L.). J. Exp. Bot. 2014, 65, 2171–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Zhang, H.; Wei, X.; Yang, L.; Yang, B.; Zhang, L.; Li, J.; Jiang, Y.Q. Functional characterization of calci-um-dependent protein kinase (CPK) 2 gene from oilseed rape (Brassica napus L.) in regulating reactive oxygen species signaling and cell death control. Gene 2018, 651, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Zhang, L.; Chen, C.; Li, J.; Ye, S.; Liu, G.; Mei, X.; Tang, K.; Luo, L. Isolation and characterization of BnMKK1 responsive to multiple stresses and affecting plant architecture in tobacco. Acta Physiol. Plant. 2014, 36, 1313–1324. [Google Scholar] [CrossRef]
- Jian, H.; Lu, K.; Yang, B.; Wang, T.; Zhang, L.; Zhang, A.; Wang, J.; Liu, L.; Qu, C.; Li, J. Genome-wide analysis and ex-pression profiling of the SUC and SWEET gene families of sucrose transporters in oilseed rape (Brassica napus L.). Front. Plant Sci. 2016, 7, 1464. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Xiao, H.; Sun, J.; Wang, S.; Wang, J.; Chang, P.; Zhou, X.; Lei, B.; Lu, K.; Luo, F.; et al. Genome-wide analysis and expression profiling of the HMA gene family in Brassica napus under cd stress. Plant Soil 2018, 426, 365–381. [Google Scholar] [CrossRef]
- Zhang, X.D.; Zhao, K.X.; Yang, Z.M. Identification of genomic ATP binding cassette (ABC) transporter genes and Cd-responsive ABCs in Brassica napus. Gene 2018, 664, 139–151. [Google Scholar] [CrossRef]
- Hu, W.; Yuan, Q.; Wang, Y.; Cai, R.; Deng, X.; Wang, J.; Zhou, S.; Chen, M.; Chen, L.; Huang, C.; et al. Overexpression of a Wheat Aquaporin Gene, TaAQP8, Enhances Salt Stress Tolerance in Transgenic Tobacco. Plant Cell Physiol. 2012, 53, 2127–2141. [Google Scholar] [CrossRef] [Green Version]
- Lücker, J.; Bouwmeester, H.J.; Schwab, W.; Blaas, J.; Van Der Plas, L.H.; Verhoeven, H.A. Expression of Clarkia S-linalool synthase in transgenic petunia plants results in the accumulation of S-linalyl-β-d-glucopyranoside. Plant J. 2001, 27, 315–324. [Google Scholar] [CrossRef]
- Lewinsohn, E.; Schalechet, F.; Wilkinson, J.; Matsui, K.; Tadmor, Y.; Nam, K.H.; Amar, O.; Lastochkin, E.; Larkov, O.; Ravid, U.; et al. Enhanced levels of the aroma and flavor compound S-linalool by metabolic engineering of the ter-penoid pathway in tomato fruits. Plant Physiol. 2001, 127, 1256–1265. [Google Scholar] [CrossRef]
- Diemer, F.; Caissard, J.C.; Moja, S.; Chalchat, J.C.; Jullien, F. Altered monoterpene composition in transgenic mint fol-lowing the introduction of 4S-limonene synthase. Plant Physiol. Biochem. 2012, 39, 603–614. [Google Scholar] [CrossRef]
- Wei, S.; Marton, I.; Dekel, M.; Shalitin, D.; Lewinsohn, E.; Bravdo, B.A.; Shoseyov, O. Manipulating volatile emission in tobacco leaves by expressing Aspergillus niger beta-glucosidase in different subcellular compartments. Plant Biotechnol. J. 2004, 2, 341–350. [Google Scholar] [CrossRef]
- Hohn, T.M.; Ohlrogge, J.B. Expression of a Fungal Sesquiterpene Cyclase Gene in Transgenic Tobacco. Plant Physiol. 1991, 97, 460–462. [Google Scholar] [CrossRef] [Green Version]
- Davidovich-Rikanati, R.; Lewinsohn, E.; Bar, E.; Iijima, Y.; Pichersky, E.; Sitrit, Y. Overexpression of the lemon basil α-zingiberene synthase gene (ZIS) increases both mono- and sesquiterpene contents in tomato fruit. Plant J. 2008, 56, 228–238. [Google Scholar] [CrossRef] [Green Version]
- Aharoni, A.; Giri, A.P.; Deuerlein, S.; Griepink, F.; De Kogel, W.-J.; Verstappen, F.W.A.; Verhoeven, H.A.; Jongsma, M.A.; Schwab, W.; Bouwmeester, H.J. Terpenoid Metabolism in Wild-Type and Transgenic Arabidopsis Plants. Plant Cell 2003, 15, 2866–2884. [Google Scholar] [CrossRef] [Green Version]
- Besumbes, Ó.; Sauret-Güeto, S.; Phillips, M.A.; Imperial, S.; Rodríguez-Concepción, M.; Boronat, A. Metabolic engineer-ing of isoprenoid biosynthesis in Arabidopsis for the production of taxadiene, the first committed precursor of Taxol. Biotechnol. Bioeng. 2004, 88, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Boschi, F.; Schvartzman, C.; Murchio, S.; Ferreira, V.; Siri, M.I.; Galván, G.A.; Smoker, M.; Stransfeld, L.; Zipfel, C.; Vilaró, F.L.; et al. Enhanced bacterial wilt resistance in potato through expression of Arabidopsis EFR and introgression of quantitative resistance from Solanum commersonii. Front. Plant Sci. 2017, 8, 1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, D.M.; Stall, R.E.; Jones, J.B.; Pauly, M.H.; Vallad, G.E.; Dahlbeck, D.; Staskawicz, B.J.; Scott, J.W. Transgenic resistance confers effective field level control of bacterial spot disease in tomato. PLoS ONE 2012, 7, e42036. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, B.; Spalding, M.H.; Weeks, D.P.; Yang, B. High-efficiency TALEN-based gene editing produces disease-resistant rice. Nat. Biotechnol. 2012, 30, 390–392. [Google Scholar] [CrossRef]
- Hummel, A.W.; Doyle, E.; Bogdanove, A.J. Addition of transcription activator-like effector binding sites to a pathogen strain-specific rice bacterial blight resistance gene makes it effective against additional strains and against bacterial leaf streak. New Phytol. 2012, 195, 883–893. [Google Scholar] [CrossRef]
- Xu, G.; Yuan, M.; Ai, C.; Liu, L.; Zhuang, E.; Karapetyan, S.; Wang, S.; Dong, X. uORF-mediated translation allows engi-neered plant disease resistance without fitness costs. Nature 2017, 545, 491–494. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, L.; Shi, H.; Chern, M.; Yu, H.; Yi, H.; He, M.; Yin, J.; Zhu, X.; Li, Y.; et al. A single transcription factor promotes both yield and immunity in rice. Science 2018, 361, 1026–1028. [Google Scholar] [CrossRef] [Green Version]
- Kusch, S.; Panstruga, R. mlo-based resistance: An apparently universal ‘weapon’ to defeat powdery mildew disease. Mol. Plant-Microbe Interact. 2017, 30, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iliescu, E.C.; Balogh, M.; Szabo, Z.; Kiss, G.B. Identification of a Xanthomonas Euvesicatoria Resistance Gene from Pep-Per (Capsicum annuum) and Method for Generating Plants with Resistance. International (PCT) Patent Application. Budapest (HU) WO/2014/068346A2, 30 October 2013. [Google Scholar]
- Huang, H.E.; Ger, M.J.; Yip, M.K.; Chen, C.Y.; Pandey, A.K.; Feng, T.Y. A hypersensitive response was induced by viru-lent bacteria in transgenic tobacco plants overexpressing a plant ferredoxin-like protein (PFLP). Physiol. Mol. Plant Pathol. 2004, 64, 103–110. [Google Scholar] [CrossRef]
- Schnippenkoetter, W.; Lo, C.; Liu, G.; Dibley, K.; Chan, W.L.; White, J.; Milne, R.; Zwart, A.; Kwong, E.; Keller, B.; et al. The wheat Lr34 multi pathogen resistance gene confers resistance to anthracnose and rust in sorghum. Plant Biotechnol. J. 2017, 15, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Ji, R.; Guo, X.; Foster, S.J.; Chen, H.; Dong, C.; Liu, Y.; Hu, Q.; Liu, S. Expressing a gene encoding wheat oxalate oxidase enhances resistance to Sclerotinia sclerotiorum in oilseed rape (Brassica napus). Planta 2008, 228, 331–340. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, M.; Zhang, Z.; Ren, L.; Du, L.; Zhang, B.; Xu, H.; Xin, Z. Expression of a radish defensin in transgenic wheat confers increased resistance to Fusarium graminearum and Rhizoctonia cerealis. Funct. Integr. Genom. 2011, 11, 63–70. [Google Scholar] [CrossRef]
- Quijano, C.D.; Wichmann, F.; Schlaich, T.; Fammartino, A.; Huckauf, J.; Schmidt, K.; Unger, C.; Broer, I.; Sautter, C. KP4 to control Ustilago tritici in wheat: Enhanced greenhouse resistance to loose smut and changes in transcript abundance of pathogen related genes in infected KP4 plants. Biotechnol. Rep. 2016, 11, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Rustagi, A.; Kumar, D.; Shekhar, S.; Yusuf, M.A.; Misra, S.; Sarin, N.B. Transgenic Brassica juncea Plants Expressing MsrA1, a Synthetic Cationic Antimicrobial Peptide, Exhibit Resistance to Fungal Phytopathogens. Mol. Biotechnol. 2014, 56, 535–545. [Google Scholar] [CrossRef]
- Bonfim, K.; Faria, J.C.; Nogueira, E.O.P.L.; Mendes, É.A.; Aragão, F.J.L. RNAi-Mediated Resistance to Bean golden mosaic virus in Genetically Engineered Common Bean (Phaseolus vulgaris). Mol. Plant-Microbe Interact. 2007, 20, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Lawson, C.; Kaniewski, W.; Haley, L.; Rozman, R.; Newell, C.; Sanders, P.; Tumer, N.E. Engineering Resistance to Mixed Virus Infection in a Commercial Potato Cultivar: Resistance to Potato Virus X and Potato Virus Y in Transgenic Russet Burbank. Nat. Biotechnol. 1990, 8, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. Oxidative DNA damage, antioxidants, and cancer. BioEssays 1999, 21, 238–246. [Google Scholar] [CrossRef]
- Krinsky, N.I. Overview of lycopene, carotenoids, and disease prevention. Proc. Soc. Exp. Biol. Med. 1998, 218, 95–97. [Google Scholar] [CrossRef] [PubMed]
- Hotz, C.; Loechl, C.; Lubowa, A.; Tumwine, J.K.; Ndeezi, G.; Nandutu Masawi, A.; Baingana, R.; Carriquiry, A.; de Brauw, A.; Meenakshi, J.V.; et al. Introduction of β-carotene–rich orange sweet potato in rural Uganda resulted in in-creased vitamin A intakes among children and women and improved vitamin A status among children. J. Nutr. 2012, 142, 1871–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pixley, K.; Rojas, N.P.; Babu, R.; Mutale, R.; Surles, R.; Simpungwe, E. Biofortification of maize with provitamin A ca-rotenoids. In Carotenoids and Human Health; Springer: Berlin, Germany, 2013; pp. 271–292. [Google Scholar]
- Ceballos, H.; Morante, N.; Sánchez, T.; Ortiz, D.; Aragón, I.; Chávez, A.; Pizarro, M.; Calle, F.; Dufour, D. Rapid Cycling Recurrent Selection for Increased Carotenoids Content in Cassava Roots. Crop Sci. 2013, 53, 2342–2351. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Al-Babili, S.; Klöti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (β-carotene) bio-synthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, C.; Aquilani, R.; Dharmapuri, S.; Pallara, P.; Marusic, C.; Tavazza, R.; Bouvier, F.; Camara, B.; Giuliano, G. Meta-bolic engineering of beta-carotene and lycopene content in tomato fruit. Plant J. 2000, 24, 413–420. [Google Scholar] [CrossRef]
- D’Ambrosio, C.; Giorio, G.; Marino, I.; Merendino, A.; Petrozza, A.; Salfi, L.; Stigliani, A.L.; Cellini, F. Virtually com-plete conversion of lycopene into β-carotene in fruits of tomato plants transformed with the tomato lycopene β-cyclase (tlcy-b) cDNA. Plant Sci. 2004, 166, 207–214. [Google Scholar] [CrossRef]
- Adalid, A.M.; Roselló, S.; Nuez, F. Evaluation and selection of tomato accessions (Solanum section Lycopersicon) for content of lycopene, β-carotene and ascorbic acid. J. Food Compos. Anal. 2010, 23, 613–618. [Google Scholar] [CrossRef]
- Orchard, C. Naturally Occurring Variation in the Promoter of the Chromoplast-Specific Cyc-B Gene in Tomato Can Be Used to Modulate Levels of ß-Carotene in Ripe Tomato Fruit. Ph.D. Thesis, The Ohio State University, Columbus, OH, USA, 2014. [Google Scholar]
- Tzuri, G.; Zhou, X.; Chayut, N.; Yuan, H.; Portnoy, V.; Meir, A.; Sa’ar, U.; Baumkoler, F.; Mazourek, M.; Lewinsohn, E.; et al. A ‘golden’SNP in CmOr governs the fruit flesh color of melon (Cucumis melo). Plant J. 2015, 82, 267–279. [Google Scholar] [CrossRef]
- Lopez, A.B.; Van Eck, J.; Conlin, B.J.; Paolillo, D.J.; O’Neill, J.; Li, L. Effect of the cauliflower or transgene on carotenoid accumulation and chromoplast formation in transgenic potato tubers. J. Exp. Bot. 2008, 59, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, B.; Natoli, S.; Liew, G.; Flood, V.M. Lutein and Zeaxanthin—Food Sources, Bioavailability and Dietary Variety in Age-Related Macular Degeneration Protection. Nutrients 2017, 9, 120. [Google Scholar] [CrossRef] [PubMed]
- Flaxman, S.R.; Bourne, R.R.; Resnikoff, S.; Ackland, P.; Braithwaite, T.; Cicinelli, M.V.; Das, A.; Jonas, J.B.; Keeffe, J.; Kempen, J.H.; et al. Global causes of blindness and distance vision impairment 1990–2020: A systematic review and meta-analysis. Lancet Glob. Health 2017, 5, e1221–e1234. [Google Scholar] [CrossRef] [Green Version]
- Karniel, U.; Koch, A.; Zamir, D.; Hirschberg, J. Development of zeaxanthin-rich tomato fruit through genetic manipulations of carotenoid biosynthesis. Plant Biotechnol. J. 2020, 18, 2292–2303. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zeng, J.; Li, Y.; Hu, W.; Chen, L.; Miao, Y.; Deng, P.; Yuan, C.; Ma, C.; Chen, X.; et al. Enrichment of provitamin A content in wheat (Triticum aestivum L.) by introduction of the bacterial carotenoid biosynthetic genes CrtB and CrtI. J. Exp. Bot. 2014, 65, 2545–2556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Wang, X.; Miao, Y.; Wang, C.; Zang, M.; Chen, X.; Li, M.; Li, X.; Wang, Q.; Li, K.; et al. Metabolic engineer-ing of wheat provitamin A by simultaneously overexpressing CrtB and silencing carotenoid hydroxylase (TaHYD). J. Agric. Food Chem. 2015, 63, 9083–9092. [Google Scholar] [CrossRef]
- Ducreux, L.J.; Morris, W.L.; Hedley, P.E.; Shepherd, T.; Davies, H.V.; Millam, S.; Taylor, M.A. Metabolic engineering of high carotenoid potato tubers containing enhanced levels of β-carotene and lutein. J. Exp. Bot. 2005, 56, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Diretto, G.; Tavazza, R.; Welsch, R.; Pizzichini, D.; Mourgues, F.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic engi-neering of potato tuber carotenoids through tuber-specific silencing of lycopene epsilon cyclase. BMC Plant Biol. 2006, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Diretto, G.; Al-Babili, S.; Tavazza, R.; Papacchioli, V.; Beyer, P.; Giuliano, G. Metabolic Engineering of Potato Carotenoid Content through Tuber-Specific Overexpression of a Bacterial Mini-Pathway. PLoS ONE 2007, 2, e350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Eck, J.; Conlin, B.; Garvin, D.F.; Mason, H.; Navarre, D.A.; Brown, C.R. Enhancing beta-carotene content in potato by rnai-mediated silencing of the beta-carotene hydroxylase gene. Am. J. Potato Res. 2007, 84, 331–342. [Google Scholar] [CrossRef]
- Zhu, C.; Naqvi, S.; Breitenbach, J.; Sandmann, G.; Christou, P.; Capell, T. Combinatorial genetic transformation gener-ates a library of metabolic phenotypes for the carotenoid pathway in maize. Proc. Natl. Acad. Sci. USA 2008, 105, 18232–18237. [Google Scholar] [CrossRef] [Green Version]
- Aluru, M.; Xu, Y.; Guo, R.; Wang, Z.; Li, S.; White, W.; Wang, K.; Rodermel, S. Generation of transgenic maize with enhanced provitamin A content. J. Exp. Bot. 2008, 59, 3551–3562. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, S.; Zhu, C.; Farre, G.; Ramessar, K.; Bassie, L.; Breitenbach, J.; Conesa, D.P.; Ros, G.; Sandmann, G.; Capell, T.; et al. Transgenic multivitamin corn through biofortification of endosperm with three vitamins representing three distinct metabolic pathways. Proc. Natl. Acad. Sci. USA 2009, 106, 7762–7767. [Google Scholar] [CrossRef] [Green Version]
- Römer, S.; Fraser, P.D.; Kiano, J.W.; Shipton, C.A.; Misawa, N.; Schuch, W.; Bramley, P.M. Elevation of the provitamin A content of transgenic tomato plants. Nat. Biotechnol. 2000, 18, 666–669. [Google Scholar] [CrossRef]
- Dharmapuri, S.; Rosati, C.; Pallara, P.; Aquilani, R.; Bouvier, F.; Camara, B.; Giuliano, G. Metabolic engineering of xan-thophyll content in tomato fruits. FEBS Lett. 2002, 519, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Fraser, P.D.; Romer, S.; Shipton, C.A.; Mills, P.B.; Kiano, J.W.; Misawa, N.; Drake, R.G.; Schuch, W.; Bramley, P.M. Evalu-ation of transgenic tomato plants expressing an additional phytoene synthase in a fruit-specific manner. Proc. Natl. Acad. Sci. USA 2002, 99, 1092–1097. [Google Scholar] [CrossRef] [Green Version]
- Enfissi, E.M.A.; Fraser, P.D.; Lois, L.-M.; Boronat, A.; Schuch, W.; Bramley, P.M. Metabolic engineering of the mevalonate and non-mevalonate isopentenyl diphosphate-forming pathways for the production of health-promoting isoprenoids in tomato. Plant Biotechnol. J. 2004, 3, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J.; Gaffé, J.; Alcaraz, J.P.; Carde, J.P.; Bramley, P.M.; Fraser, P.D.; Kuntz, M. Fibrillin influence on plastid ultra-structure and pigment content in tomato fruit. Phytochemistry 2007, 68, 1545–1556. [Google Scholar] [CrossRef]
- Apel, W.; Bock, R. Enhancement of carotenoid biosynthesis in transplastomic tomatoes by induced lycopene-to-provitamin A conversion. Plant Physiol. 2009, 151, 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.; Zhou, W.; Zhang, J.; Xu, Q.; Deng, X. Effect of the Citrus Lycopene β-Cyclase Transgene on Carotenoid Metabolism in Transgenic Tomato Fruits. PLoS ONE 2012, 7, e32221. [Google Scholar] [CrossRef] [Green Version]
- Failla, M.L.; Chitchumroonchokchai, C.; Siritunga, D.; De Moura, F.F.; Fregene, M.; Manary, M.J.; Sayre, R.T. Retention during Processing and Bioaccessibility of β-Carotene in High β-Carotene Transgenic Cassava Root. J. Agric. Food Chem. 2012, 60, 3861–3866. [Google Scholar] [CrossRef] [PubMed]
- Welsch, R.; Arango, J.; Bär, C.; Salazar, B.; Al-Babili, S.; Beltrán, J.; Chavarriaga, P.; Ceballos, H.; Tohme, J.; Beyer, P. Pro-vitamin A accumulation in cassava (Manihot esculenta) roots driven by a single nucleotide polymorphism in a phytoene synthase gene. Plant Cell 2010, 22, 3348–3356. [Google Scholar] [CrossRef] [Green Version]
- Sayre, R.; Beeching, J.R.; Cahoon, E.B.; Egesi, C.; Fauquet, C.; Fellman, J.; Fregene, M.; Gruissem, W.; Mallowa, S.; Manary, M.; et al. The BioCassava Plus Program: Biofortification of Cassava for Sub-Saharan Africa. Annu. Rev. Plant Biol. 2011, 62, 251–272. [Google Scholar] [CrossRef]
- Che, P.; Zhao, Z.Y.; Glassman, K.; Dolde, D.; Hu, T.X.; Jones, T.J.; Gruis, D.F.; Obukosia, S.; Wambugu, F.; Albertsen, M.C. Elevated vitamin E content improves all-trans β-carotene accumulation and stability in biofortified sorghum. Proc. Natl. Acad. Sci. USA 2016, 113, 11040–11045. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Paolillo, D.J.; Parthasarathy, M.V.; DiMuzio, E.M.; Garvin, D.F. A novel gene mutation that confers abnormal patterns of β-carotene accumulation in cauliflower (Brassica oleracea var. botrytis). Plant J. 2001, 26, 59–67. [Google Scholar] [CrossRef]
- Schijlen, E.; De Vos, C.R.; Jonker, H.; Broeck, H.V.D.; Molthoff, J.; Van Tunen, A.; Martens, S.; Bovy, A. Pathway engineering for healthy phytochemicals leading to the production of novel flavonoids in tomato fruit. Plant Biotechnol. J. 2006, 4, 433–444. [Google Scholar] [CrossRef]
- Muir, S.R.; Collins, G.J.; Robinson, S.; Hughes, S.; Bovy, A.; De Vos, C.R.; van Tunen, A.J.; Verhoeyen, M.E. Overexpres-sion of petunia chalcone isomerase in tomato results in fruit containing increased levels of flavonols. Nat. Biotechnol. 2001, 19, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Bovy, A.; De Vos, R.; Kemper, M.; Schijlen, E.; Pertejo, M.A.; Muir, S.; Collins, G.; Robinson, S.; Verhoeyen, M.; Hughes, S.; et al. High-Flavonol Tomatoes Resulting from the Heterologous Expression of the Maize Transcription Factor Genes LC and C1. Plant Cell 2002, 14, 2509–2526. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Butelli, E.; Alseekh, S.; Tohge, T.; Rallapalli, G.; Luo, J.; Kawar, P.G.; Hill, L.; Santino, A.; Fernie, A.R.; et al. Multi-level engineering facilitates the production of phenylpropanoid compounds in tomato. Nat. Commun. 2015, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Butelli, E.; Titta, L.; Giorgio, M.; Mock, H.-P.; Matros, A.; Peterek, S.; Schijlen, E.G.W.M.; Hall, R.D.; Bovy, A.G.; Luo, J.; et al. Enrichment of tomato fruit with health-promoting anthocyanins by expression of select transcription factors. Nat. Biotechnol. 2008, 26, 1301–1308. [Google Scholar] [CrossRef]
- Jian, W.; Cao, H.; Yuan, S.; Liu, Y.; Lu, J.; Lu, W.; Li, N.; Wang, J.; Zou, J.; Tang, N.; et al. SlMYB75, an MYB-type transcription factor, promotes anthocyanin accumulation and enhances volatile aroma production in tomato fruits. Hortic. Res. 2019, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef] [Green Version]
- Ashikari, M.; Sasaki, A.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Datta, S.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Loss-of-function of a Rice Gibberellin Biosynthetic Gene, GA20 oxidase (GA20ox-2), Led to the Rice ‘Green Revolution’. Breed. Sci. 2002, 52, 143–150. [Google Scholar] [CrossRef] [Green Version]
- Spielmeyer, W.; Ellis, M.H.; Chandler, P.M. Semidwarf (sd-1), “green revolution” rice, contains a defective gibberellin 20-oxidase gene. Proc. Natl. Acad. Sci. USA 2002, 99, 9043–9048. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S. Gibberellin Metabolism and its Regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Ha, S.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.P. Cytokinins: Metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci. 2012, 17, 172–179. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidop-sis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, R.; Sahoo, K.K.; Tripathi, A.K.; Kumar, R.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Knockdown of an inflo-rescence meristem-specific cytokinin oxidase–OsCKX2 in rice reduces yield penalty under salinity stress condition. Plant Cell Environ. 2018, 41, 936–946. [Google Scholar] [CrossRef]
- Meitzel, T.; Radchuk, R.; Nunes-Nesi, A.; Fernie, A.R.; Link, W.; Weschke, W.; Weber, H. Hybrid embryos of Vicia faba develop enhanced sink strength, which is established during early development. Plant J. 2010, 65, 517–531. [Google Scholar] [CrossRef] [PubMed]
- Merewitz, E.B.; Gianfagna, T.; Huang, B. Effects of SAG12-ipt and HSP18.2-ipt Expression on Cytokinin Production, Root Growth, and Leaf Senescence in Creeping Bentgrass Exposed to Drought Stress. J. Am. Soc. Hortic. Sci. 2010, 135, 230–239. [Google Scholar] [CrossRef]
- Qin, H.; Gu, Q.; Zhang, J.; Sun, L.; Kuppu, S.; Zhang, Y.; Burow, M.; Payton, P.; Blumwald, E.; Zhang, H. Regulated ex-pression of an isopentenyltransferase gene (IPT) in peanut significantly improves drought tolerance and increases yield under field conditions. Plant Cell Physiol. 2011, 52, 1904–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleg, Z.; Reguera, M.; Tumimbang, E.; Walia, H.; Blumwald, E. Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotechnol. J. 2011, 9, 747–758. [Google Scholar] [CrossRef]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-X.; Yang, J.-S.; Li, X.-M.; Yu, M.; Wang, J. Effects of Irrigation Water Quality and Drip Tape Arrangement on Soil Salinity, Soil Moisture Distribution, and Cotton Yield (Gossypium hirsutum L.) Under Mulched Drip Irrigation in Xinjiang, China. J. Integr. Agric. 2012, 11, 502–511. [Google Scholar] [CrossRef]
- Ke, Q.; Wang, Z.; Ji, C.Y.; Jeong, J.C.; Lee, H.S.; Li, H.; Xu, B.; Deng, X.; Kwak, S.S. Transgenic poplar expressing Arabidop-sis YUCCA6 exhibits auxin-overproduction phenotypes and increased tolerance to abiotic stress. Plant Physiol. Biochem. 2015, 94, 19–27. [Google Scholar] [CrossRef]
- Miao, C.; Xiao, L.; Hua, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J.-K. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. USA 2018, 115, 6058–6063. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.T.; Prasad, V.; Yang, P.T.; Wu, J.F.; David Ho, T.H.; Charng, Y.Y.; Chan, M.T. Expression of Arabidopsis CBF1 regu-lated by an ABA/stress inducible promoter in transgenic tomato confers stress tolerance without affecting yield. Plant Cell Environ. 2003, 26, 1181–1190. [Google Scholar] [CrossRef]
- Wan, X.-R.; Li, L. Regulation of ABA level and water-stress tolerance of Arabidopsis by ectopic expression of a peanut 9-cis-epoxycarotenoid dioxygenase gene. Biochem. Biophys. Res. Commun. 2006, 347, 1030–1038. [Google Scholar] [CrossRef]
- Wagner, A.; Donaldson, L.; Kim, H.; Phillips, L.; Flint, H.; Steward, D.; Torr, K.; Koch, G.; Schmitt, U.; Ralph, J. Suppression of 4-coumarate-CoA ligase in the coniferous gymnosperm Pinus radiata. Plant Physiol. 2009, 149, 370–383. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wang, L.; Zhang, W.; Takechi, K.; Takano, H.; Lin, X. Overexpression of UDP-glucose pyrophosphorylase from Larix gmelinii enhances vegetative growth in transgenic Arabidopsis thaliana. Plant Cell Rep. 2014, 33, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, C.; Ketelaar, K.; Schneider, R.; Villalobos, J.A.; Somerville, C.R.; Persson, S.; Wallace, I.S. BRASSI-NOSTEROID INSENSITIVE2 negatively regulates cellulose synthesis in Arabidopsis by phosphorylating cellulose syn-thase 1. Proc. Natl. Acad. Sci. USA 2017, 114, 3533–3538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Zhao, J.; Li, X.; Qin, L.; Yan, X.; Liao, H. A soybean β-expansin gene GmEXPB2 intrinsically involved in root system architecture responses to abiotic stresses. Plant J. 2011, 66, 541–552. [Google Scholar] [CrossRef]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription fac-tors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambavaram, M.M.; Krishnan, A.; Trijatmiko, K.R.; Pereira, A. Coordinated activation of cellulose and repression of lig-nin biosynthesis pathways in rice. Plant Physiol. 2011, 155, 916–931. [Google Scholar] [CrossRef] [Green Version]
- Dai, F.; Zhang, C.; Jiang, X.; Kang, M.; Yin, X.; Lü, P.; Zhang, X.; Zheng, Y.; Gao, J. RhNAC2 and RhEXPA4 Are Involved in the Regulation of Dehydration Tolerance during the Expansion of Rose Petals. Plant Physiol. 2012, 160, 2064–2082. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Feng, S.; Huang, J.; Wang, Y.; Wu, L.; Li, X.; Wang, L.; Tu, Y.; Xia, T.; Li, J.; et al. AtCesA8-driven OsSUS3 ex-pression leads to largely enhanced biomass saccharification and lodging resistance by distinctively altering lignocellulose features in rice. Biotechnol. Biofuels 2017, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- De La Garza, R.D.; Quinlivan, E.P.; Klaus, S.M.J.; Basset, G.J.C.; Gregory, J.F.; Hanson, A.D. Folate biofortification in tomatoes by engineering the pteridine branch of folate synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 13720–13725. [Google Scholar] [CrossRef] [Green Version]
- Storozhenko, S.; De Brouwer, V.; Volckaert, M.; Navarrete, O.; Blancquaert, D.; Zhang, G.-F.; Lambert, W.E.; Van Der Straeten, D. Folate fortification of rice by metabolic engineering. Nat. Biotechnol. 2007, 25, 1277–1279. [Google Scholar] [CrossRef]
- Nunes, A.C.S.; Kalkmann, D.C.; Aragão, F.J.L. Folate biofortification of lettuce by expression of a codon optimized chicken GTP cyclohydrolase I gene. Transgenic Res. 2009, 18, 661–667. [Google Scholar] [CrossRef]
- Dong, W.; Cheng, Z.J.; Lei, C.L.; Wang, J.L.; Wang, J.; Wu, F.Q.; Zhang, X.; Guo, X.P.; Zhai, H.Q.; Wan, J.M. Overexpres-sion of folate biosynthesis genes in rice (Oryza sativa L.) and evaluation of their impact on seed folate content. Plant Food Hum. Nutr. 2014, 69, 379–385. [Google Scholar] [CrossRef]
- Hirsch, A.M. Brief History of the Discovery of Nitrogen-Fixing Organisms. Advance Access Published 2009. Available online: http://www.mcdb.ucla.edu/Research/Hirsch/imagesb/HistoryDiscoveryN2fixingOrganisms.pdf (accessed on 25 October 2020).
- Desbrosses, G.J.; Stougaard, J. Root nodulation: A paradigm for how plant-microbe symbiosis influences host develop-mental pathways. Cell Host Microbe 2011, 10, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Bhattacharyya, S.; Kumar, R.; Kumar, A.; Ibañez, F.J.; Wang, J.; Guo, B.; Sudini, H.K.; Gopalakrishnan, S.; Dasgupta, M.; et al. Molecular Basis of Root Nodule Symbiosis between Bradyrhizobium and ‘Crack-Entry’ Legume Groundnut (Arachis hypogaea L.). Plants 2020, 9, 276. [Google Scholar] [CrossRef] [Green Version]
- Maunoury, N.; Redondo-Nieto, M.; Bourcy, M.; Van De Velde, W.; Alunni, B.; Laporte, P.; Durand, P.; Agier, N.; Marisa, L.; Vaubert, D.; et al. Differentiation of Symbiotic Cells and Endosymbionts in Medicago truncatula Nodulation Are Coupled to Two Transcriptome-Switches. PLoS ONE 2010, 5, e9519. [Google Scholar] [CrossRef] [PubMed]
- Takanashi, K.; Takahashi, H.; Sakurai, N.; Sugiyama, A.; Suzuki, H.; Shibata, D.; Nakazono, M.; Yazaki, K. Tissue-Specific Transcriptome Analysis in Nodules of Lotus japonicus. Mol. Plant-Microbe Interact. 2012, 25, 869–876. [Google Scholar] [CrossRef] [Green Version]
- Demina, I.V.; Persson, T.; Santos, P.; Plaszczyca, M.; Pawlowski, K. Comparison of the Nodule vs. Root Transcriptome of the Actinorhizal Plant Datisca glomerata: Actinorhizal Nodules Contain a Specific Class of Defensins. PLoS ONE 2013, 8, e72442. [Google Scholar] [CrossRef] [Green Version]
- Limpens, E.; Moling, S.; Hooiveld, G.; Pereira, P.A.; Bisseling, T.; Becker, J.D.; Küster, H. Cell and tissue-specific tran-scriptome analyses of Medicago truncatula root nodules. PLoS ONE 2013, 8, e64377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breakspear, A.; Liu, C.; Roy, S.; Stacey, N.J.; Rogers, C.; Trick, M.; Morieri, G.; Mysore, K.S.; Wen, J.; Oldroyd, G.E.; et al. The Root Hair “Infectome” of Medicago truncatula Uncovers Changes in Cell Cycle Genes and Reveals a Requirement for Auxin Signaling in Rhizobial Infection. Plant Cell 2014, 26, 4680–4701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, B.; Rodde, N.; Jardinaud, M.F.; Timmers, T.; Sauviac, L.; Cottret, L.; Carrere, S.; Sallet, E.; Courcelle, E.; Moreau, S.; et al. An integrated analysis of plant and bacterial gene expression in symbiotic root nodules using laser-capture mi-crodissection coupled to RNA sequencing. Plant J. 2014, 77, 817–837. [Google Scholar] [CrossRef]
- Ligeza, B.O.-; Parizot, B.; Gantet, P.; Beeckman, T.; Bennett, M.J.; Draye, X. Post-embryonic root organogenesis in cereals: Branching out from model plants. Trends Plant Sci. 2013, 18, 459–467. [Google Scholar] [CrossRef]
- Beatty, P.H.; Good, A.G. Future Prospects for Cereals That Fix Nitrogen. Science 2011, 333, 416–417. [Google Scholar] [CrossRef]
- Dent, D.; Cocking, E. Establishing symbiotic nitrogen fixation in cereals and other non-legume crops: The Greener Ni-trogen Revolution. Agric. Food Secur. 2017, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
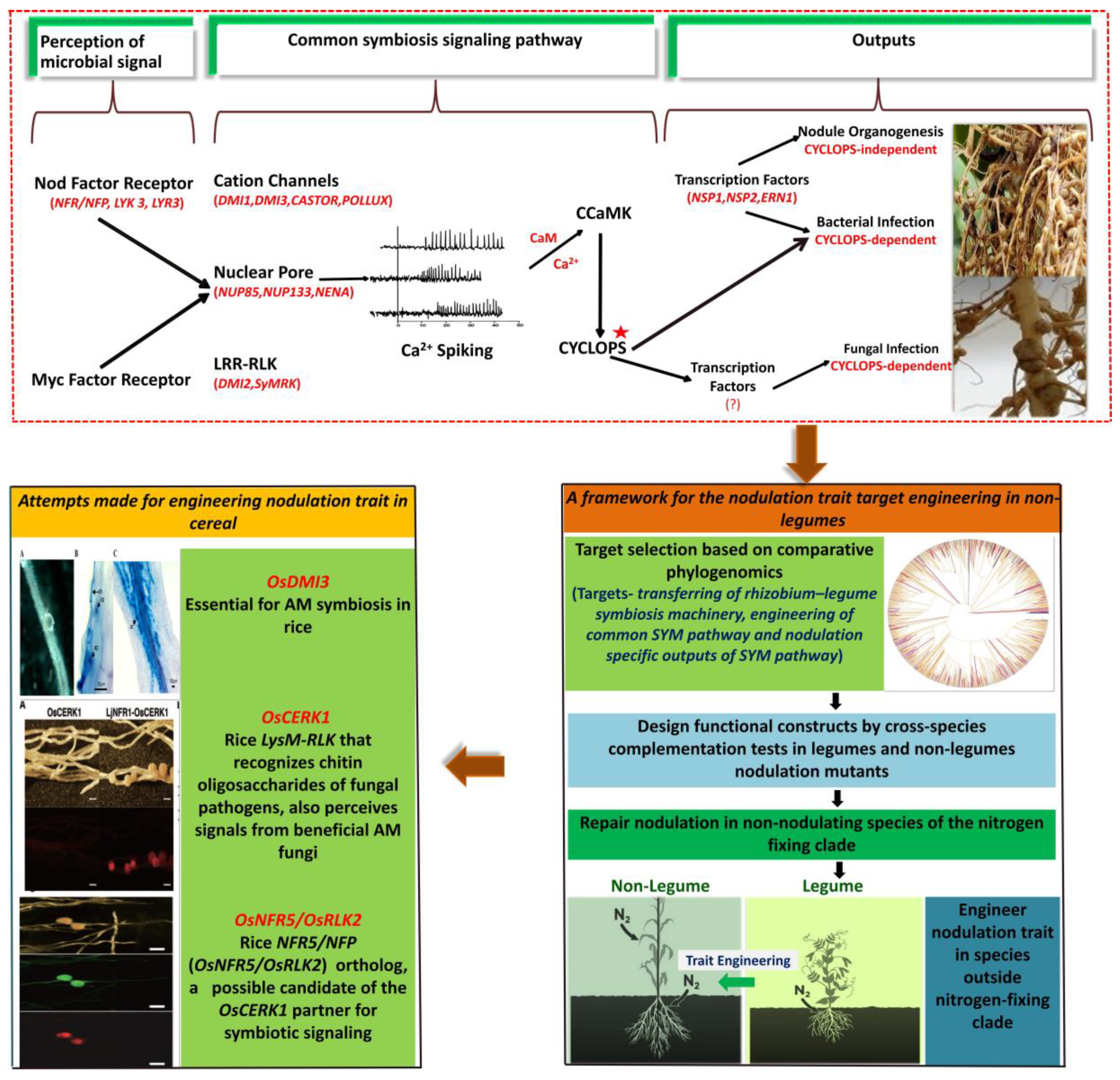
- Chen, C.; Gao, M.; Liu, J.; Zhu, H. Fungal Symbiosis in Rice Requires an Ortholog of a Legume Common Symbiosis Gene Encoding a Ca2+/Calmodulin-Dependent Protein Kinase. Plant Physiol. 2007, 145, 1619–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, K.; Kozaki, T.; Kouzai, Y.; Ozawa, K.; Ishii, K.; Asamizu, E.; Okabe, Y.; Umehara, Y.; Miyamoto, A.; Kobae, Y.; et al. The Bifunctional Plant Receptor, OsCERK1, Regulates Both Chitin-Triggered Immunity and Arbuscular Mycorrhizal Symbiosis in Rice. Plant Cell Physiol. 2014, 55, 1864–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyata, K.; Hayafune, M.; Kobae, Y.; Kaku, H.; Nishizawa, Y.; Masuda, Y.; Shibuya, N.; Nakagawa, T. Evaluation of the role of the LysM receptor-like kinase, OsNFR5/OsRLK2 for AM symbiosis in rice. Plant Cell Physiol. 2016, 57, 2283–2290. [Google Scholar] [CrossRef] [Green Version]
- Griesmann, M.; Chang, Y.; Liu, X.; Song, Y.; Haberer, G.; Crook, M.B.; Billault-Penneteau, B.; Lauressergues, D.; Keller, J.; Imanishi, L.; et al. Phylogenomics reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 2018, 361, eaat1743. [Google Scholar] [CrossRef] [Green Version]
- Van Velzen, R.; Doyle, J.J.; Geurts, R. A Resurrected Scenario: Single Gain and Massive Loss of Nitrogen-Fixing Nodulation. Trends Plant Sci. 2019, 24, 49–57. [Google Scholar] [CrossRef]
- Billault-Penneteau, B.; Sandré, A.; Folgmann, J.; Parniske, M.; Pawlowski, K. Dryas as a Model for Studying the Root Symbioses of the Rosaceae. Front. Plant Sci. 2019, 10, 661. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; York, T.; Pumplin, N.; Mueller, L.A.; Harrison, M.J. Genes conserved for arbuscular mycorrhizal symbiosis identified through phylogenomics. Nat. Plants 2016, 2, 15208. [Google Scholar] [CrossRef]
- Van Velzen, R.; Holmer, R.; Bu, F.; Rutten, L.; van Zeijl, A.; Liu, W.; Santuari, L.; Cao, Q.; Sharma, T.; Shen, D.; et al. Comparative genomics of the nonlegume Parasponia reveals insights into evolution of nitrogen-fixing rhizobium symbioses. Proc. Natl. Acad. Sci. USA 2018, 115, E4700–E4709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.E.D.; Schroeder, J.I. Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Wu, T.; Wang, M.; Shi, S.; Yuan, G.; Li, X.; Chong, H.; Wu, B.; Zheng, P. Enzymatic biosynthesis and immobili-zation of polyprotein verified at the single-molecule level. Nat. Commun. 2019, 10, 2775. [Google Scholar] [CrossRef]
- Rubio, L.M.; Ludden, P.W. Biosynthesis of the Iron-Molybdenum Cofactor of Nitrogenase. Annu. Rev. Microbiol. 2008, 62, 93–111. [Google Scholar] [CrossRef] [Green Version]
- Curatti, L.; Rubio, L.M. Challenges to develop nitrogen-fixing cereals by direct nif-gene transfer. Plant Sci. 2014, 225, 130–137. [Google Scholar] [CrossRef]
- Mus, F.; Crook, M.B.; Garcia, K.; Garcia Costas, A.; Geddes, B.A.; Kouri, E.D.; Paramasivan, P.; Ryu, M.H.; Oldroyd, G.E.D.; Poole, P.S.; et al. Symbiotic nitrogen fixation and the challenges to its extension to non-legumes. Appl. Environ. Microbiol. 2016, 82, 3698–3710. [Google Scholar] [CrossRef] [Green Version]
- Biswas, B.; Gressho, P.M. The role of symbiotic nitrogen fixation in sustainable production of biofuels. Int. J. Mol. Sci. 2014, 15, 7380–7397. [Google Scholar] [CrossRef] [Green Version]
- Rosenblueth, M.; Ormeño-Orrillo, E.; López-López, A.; Rogel, M.A.; Reyes-Hernández, B.J.; Martínez-Romero, J.C.; Reddy, P.M.; Martínez-Romero, E. Nitrogen Fixation in Cereals. Front. Microbiol. 2018, 9, 1794. [Google Scholar] [CrossRef] [Green Version]
- Good, A.G.; Beatty, P.H. Biotechnological approaches to improving nitrogen use efficiency in plants: Alanine ami-notransferase as a case study. In The Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 165–191. [Google Scholar]
- McAllister, C.H.; Beatty, P.H.; Good, A.G. Engineering nitrogen use efficient crop plants: The current status. Plant Biotechnol. J. 2012, 10, 1011–1025. [Google Scholar] [CrossRef]
- Fischer, J.J.; Beatty, P.H.; Good, A.G.; Muench, D.G. Manipulation of microRNA expression to improve nitrogen use efficiency. Plant Sci. 2013, 210, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.C.; Eriksson, D.; Møller, I.S.; Schjoerring, J.K. Cytosolic glutamine synthetase: A target for improvement of crop nitrogen use efficiency? Trends Plant Sci. 2014, 19, 656–663. [Google Scholar] [CrossRef]
- Good, A.G.; Johnson, S.J.; De Pauw, M.; Carroll, R.T.; Savidov, N.; Vidmar, J.; Lu, Z.; Taylor, G.; Stroeher, V. Engineering nitrogen use efficiency with alanine aminotransferase. Can. J. Bot. 2007, 85, 252–262. [Google Scholar] [CrossRef]
- Shrawat, A.K.; Carroll, R.T.; DePauw, M.; Taylor, G.J.; Good, A.G. Genetic engineering of improved nitrogen use efficiency in rice by the tissue-specific expression of alanine aminotransferase. Plant Biotechnol. J. 2008, 6, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.H.; Shrawat, A.K.; Carroll, R.T.; Zhu, T.; Good, A.G. Transcriptome analysis of nitrogen-efficient rice over-expressing alanine aminotransferase. Plant Biotechnol. J. 2009, 7, 562–576. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.H.; Carroll, R.T.; Shrawat, A.K.; Guevara, D.; Good, A.G. Physiological analysis of nitrogen-efficient rice over-expressing alanine aminotransferase under different N regimes. Botany 2013, 91, 866–883. [Google Scholar] [CrossRef]
- Beatty, P.H.; Klein, M.S.; Fischer, J.J.; Lewis, I.A.; Muench, D.G.; Good, A.G. Understanding Plant Nitrogen Metabolism through Metabolomics and Computational Approaches. Plants 2016, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Gambhir, G.; Dass, A.; Tripathi, A.K.; Singh, A.; Jha, A.K.; Rakshit, S. Genetically modified crops: Current sta-tus and future prospects. Planta 2020, 251, 1–27. [Google Scholar] [CrossRef]
- Wolt, J.D.; Wang, K.; Yang, B. The Regulatory Status of Genome-edited Crops. Plant Biotechnol. J. 2016, 14, 510–518. [Google Scholar] [CrossRef] [Green Version]
- ISAAA Database. GM Approval Database Retrieved on 17 November 2019. Available online: https://www.isaaa.org/gmapprovaldatabase/default.asp (accessed on 26 December 2020).
- Kumar, A.; Kumar, R.; Singh, N.; Mansoori, A. Regulatory Framework and Policy Decisions for Genome-Edited Crops. In Concepts and Strategies in Plant Sciences; Springer: Berlin, Germany, 2020; pp. 193–201. [Google Scholar]
- Pramanik, D.; Shelake, R.M.; Kim, M.J.; Kim, J.Y. CRISPR-mediated engineering across the central dogma in plant biology for basic research and crop improvement. Mol. Plant 2020, 14, 127–150. [Google Scholar] [CrossRef]


{kind=link}
| Crop Name | Population | Target Traits | Sample Tissue | Profiling | Significant Outcome | Reference |
|---|---|---|---|---|---|---|
| Oryza sativa | Zhenshan 97 × Minghui 63 (RIL) | Metabolome | Flag leaf and seed | Liquid chromatography (LC)–electrospray ionization (ESI)–MS/MS system | Identified twenty-four candidate genes, underlying phenolics, and related pathways | [10] |
| Oryza sativa | Sasanishiki × Habatak (BIL) | Metabolome | Seed | Liquid chromatography-quadrupole-time-of-flight-mass spectrometry | Identified genomic region and genes potentially involved in the biogenesis of apigenin-6,8-di-C-a-L-arabinoside | [40] |
| Triticum aestivum | Excalibur × Kukri (DH) | Metabolome | Flag leaf | Liquid chromatography electrospray ionization tandem mass spectrometric | Identified five major phenology-related loci | [47] |
| Triticum aestivum | KN9204 × J41 (RIL) | Metabolome | Kernel | Liquid chromatography-mass spectrometry | Identified 1005 mQTLs, linked with 24 candidate genes which modulating different metabolite levels, of which two genes are involved in flavonoids synthesis and modification. | [56] |
| Zea mays | BB RIL lines (197) and ZY RIL lines (197) | Metabolome | Mature Kernel | Liquid chromatography-mass spectrometry | Identified candidate genes for maize quality improvement | [37] |
| Zea mays | B73 × By804 (RIL) | Primary metabolism | Leaf at seedling stage, leaf at reproductive stage, and kernel | Gas chromatography time-of-flight mass spectrometry | Identified 297 mQTLs for 79 primary metabolites across three tissues | [35] |
| Hordeum vulgare | Maresi × CamB (RIL) | Metabolome | Flag leaf | Liquid chromatography–mass spectrometry | Reported mQTL in a genomic region of SNP 3011-111 and SSR Bmag0692 have linkages with metabolites | [42] |
| Hordeum vulgare | Landraces and elite genotypes | Metabolome | Flag leaf | Ion chromatography-mass spectrometry, High-performance liquid chromatography | Identified mQTLs for metabolites linked with antioxidant defense | [43] |
| Solanum lycopersicum | Introgression lines | Secondary metabolites | Fruit | Ultra performance liquid chromatography | Identified 679 mQTLs for secondary metabolites | [45] |
| Solanum lycopersicum | Introgression lines | Secondary metabolites | Fruit | Ultraperformance liquid chromatography-tandem mass spectrometry | Identified mQTLs which decrease the variability for primary and secondary metabolites called canalization metabolite quantitative trait loci (cmQTL) | [46] |
| Solanum lycopersicum | Introgression lines | Metabolome | Fruit | Gas chromatography–mass spectrometry | Identified putative 30 mQTLs for amino acids and organic acids | [27] |
| Solanum lycopersicum | RIL | Metabolome | Germinating seed | Gas chromatography-time-of-flight/mass spectrometry | Identified mQTLs for metabolites within several QTL hotspots | [48] |
| Brassica napus | Tapidor × Ningyou7 (DH) | Glucosinolates | Leaf and seed | High-performance liquid chromatography | Identified 105 mQTLs that affected glucosinolate concentration in either or both of the organs | [44] |
| Oryza sativa | Landraces and elite varieties | Metabolome | Grains | Liquid chromatography electrospray ionization tandem mass spectrometric | Identified new candidate genes which influence important metabolic and/or morphological traits | [16] |
| Oryza sativa | Landraces accessions | Secondary metabolites | Leaf | Liquid chromatography quadrupole time-of-flight mass spectrometry | Identified 323 associations among 143 SNPs and 89 metabolites | [54] |
| Oryza sativa | Landraces accessions | Phenolamides | Leaf | Liquid chromatography–mass spectrometry | Identified two spermidine hydroxyl cinnamoyl transferases (Os12g27220 and Os12g27254) that could underline the natural variation levels of spermidine conjugates in rice | [51] |
| Oryza sativa | Landraces accessions | Metabolome | Leaf | Liquid chromatography–mass spectrometry | Identified 36 candidate genes controlling metabolite levels which are of potential physiological and nutritional significance | [34] |
| Zea mays | Inbred lines | Metabolome | Leaf | Gas chromatography–mass spectrometry | Identified 26 distinct metabolites with potential associations with SNPs, explaining up to 32.0% of genetic variance | [41] |
| Zea mays | Inbred lines | Oil components | Kernel | Ultra-performance liquid chromatography | Reported 74 loci potentially associated with kernel oil concentration and fatty acid content | [57] |
| Zea mays | Inbred lines | Tocochromanol | Grain | High-performance liquid chromatography | Identified favorable ZmVTE4 haplotype and three novel gene targets for increasing the level of vitamin E and antioxidant | [58] |
| Zea mays | Inbred lines | Carotenoid | Grain | High-performance liquid chromatography | Identified 58 candidate genes involved in carotenoids biogenesis and retention in maize | [59] |
| Zea mays | Inbred lines | Metabolome | Kernel | Liquid chromatography–mass spectrometry | Identified significant causal variants for five candidate genes associated with metabolic traits | [50] |
| Triticum aestivum | Elite lines | Metabolome | Flag leaf | Gas chromatography–mass spectrometry | Reported potential associations for 6 metabolic characters, namely oxalic acid, ornithine, L-arginine, pentose alcohol III, L-tyrosine, and a sugar oligomer (oligo II), with between 1 and 17 associated SNPs | [55] |
| Solanum lycopersicum L. | Landrace accessions | Metabolome | Fruit | Gas chromatography–mass spectrometry | Identified 44 loci linked with 19 traits, including sucrose, ascorbate, malate, and citrate levels | [60] |
| Gene | Function of Gene | Phenotypes of Transgenics | Reference | |
|---|---|---|---|---|
| Phytohormones Engineering to Enhance Abiotic Stress Tolerance | ||||
| ABA | LOS5 | Key regulator of ABA biosynthesis | Enhanced ABA accumulation and drought tolerance in maize | [83] |
| AtLOS5 | Enhanced salinity tolerance attributed to enhanced Na+ efflux and H+ influx | [84] | ||
| MsZEP | Vital role in ABA biosynthesis | Heterologous expression of gene resulted in better salt and drought tolerance | [85] | |
| SnRK2.4 | Protein kinase involved in ABA signaling and root architecture maintenance | Exhibited enhanced tolerance to abiotic stress and improved photosynthesis in Arabidopsis | [86] | |
| Auxin | YUCCA6 | Auxin/IPA biosynthesis gene | Overexpression enhanced tolerance to drought and oxidative stress | [87] |
| OsIAA6 | Auxin/IAA gene family member | Enhanced drought tolerance via auxin biosynthesis regulation in transgenic rice | [88] | |
| IPT | Controls rate limiting step of cytokinin biosynthesis | Transgenic tomato showed enhanced growth and yield under salt stress | [89] | |
| Cytokinin | CKX | Cytokinin dehydrogenase | Overexpression led to enhanced drought tolerance in transgenic Arabidopsis | [90] |
| AtCKX1 | Overexpression led to enhanced drought tolerance through dehydration avoidance in transgenic barley | [91] | ||
| ERF-1 (JERF1) | Response factors of ethylene and jasmonates | Enhanced drought tolerance in rice | [92] | |
| Ethylene | ACC-Synthase | Catalyzes rate-limiting step in ethylene biosynthesis | Transgenic maize showed reduced ethylene levels with better drought tolerance (gene silencing) | [93] |
| ZmARGOS | Negative regulators of ethylene signal transduction | Enhanced drought tolerance in transgenic Arabidopsis and maize | [94] | |
| OsGSK1 | BR negative regulator | Improved tolerance of knockout mutants to cold, heat, salt, and drought stresses | [95] | |
| Brassinosteroids | AtHSD1 | Role in BR biosynthesis | Overproduction enhanced growth, yield, and salinity tolerance | [96] |
| BdBRI1 | BR-receptor gene | Down-regulation improved drought tolerance with dwarf phenotypes of purple false brome | [97] | |
| Metabolic Engineering of Secondary Metabolic Pathways Genes | ||||
| Flavonoid Biosynthetic Pathway | MYB12 | Transcription factor, regulate the biosynthesis of phenylpropanoid | Overexpression in Arabidopsis enhanced drought and salt tolerance | [98] |
| DFR-OX B | Catalyzes the reduction of dihydroflavonols to leucoanthocyanidins in anthocyanin biosynthesis | Overexpression in Brassica napus enhanced drought and salt tolerance | [99] | |
| PFG1/PAP1 | Overexpression in Arabidopsis enhanced oxidative and drought tolerance | [100] | ||
| Carotenoid Biosynthetic Pathway | β-LCY1 | Involved in beta-carotene biosynthesis pathway | Overexpression in Nicotiana tabacum enhanced drought and salt tolerance | [101] |
| Inhibition in Arabidopsis and Nicotiana enhanced salinity tolerance | [102] | |||
| IPP biosynthetic pathway | GGPS | Involved in the synthesis of an osmolyte glucosyl glycerol | Overexpression in Arabidopsis thaliana enhanced osmotic stress tolerance | [103] |
| Metabolic Engineering for Enhancing Photosynthetic Efficiency | ||||
| Light Harvesting Enzyme | PsbS | Plays a crucial role in xanthophyll-dependent nonphotochemical quenching | Overexpression increases leaf CO2 uptake and plant dry matter productivity in tobacco | [104] |
| Overexpression reduces water loss per CO2 assimilated in tobacco | [105] | |||
| Calvin–Benson cycle | SBPase | Key regulator of carbon flux | Overexpression enhances photosynthesis against high temperature stress in transgenic rice | [106] |
| Overexpression increases photosynthetic carbon assimilation, leaf area, and biomass yield in tobacco | [107] | |||
| Overexpression increases photosynthesis and grain yield in wheat | [108] | |||
| Photorespiration | GCS H-protein | Catalyzes the degradation of glycine | Overexpressing increases biomass yield in transgenic tobacco plants | [109] |
| GDC-L protein | Catalyzes the tetrahydrofolate-dependent catabolism of glycine | Overexpression increased rates of CO2 assimilation, photorespiration, and dry weight in Arabidopsis | [110] | |
| GDC-T protein | Tetrahydrofolate dependent protein, catalyzes glycine | Overexpression neither altered photosynthetic CO2 uptake nor plant growth in Arabidopsis | [111] | |
| Electron Transport | Algal Cyt c6 | Participates in algal photosynthetic electron transport chain | Overexpression increase CO2 assimilation rates and plant growth in Arabidopsis | [112] |
| Constitutive expression enhanced water use efficiency, chlorophyll and carotenoid content in tobacco | [113] | |||
| Rieske FeS | Regulates electron transfer | Constitutive expression enhanced photosynthetic electron transport rates, chlorophyll and carotenoid content | [114] | |
| Carbon transport | Cyanobacterial inorganic carbon transporter B | Regulates CO2 concentration mechanism | Significantly higher photosynthetic rates and biomass was observed in overexpressed Arabidopsis lines | [115,116] |
| Overexpression enhanced CO2 assimilation rates in rice and tobacco | [117] | |||
| Genome Editing Mediated Metabolic Engineering | ||||
| CRISPR/Cas9 multiplex gene editing | IFS (isoflavone synthase) | Plays significant role in biosynthesis of isoflavonoids | Mutation enhanced isoflavone content and resistance to soya bean mosaic virus (SMV) | [118] |
| GmSPL9 genes | Regulate plant architecture | Targeted mutagenesis altered plant architecture and yield in soybean | [119] | |
| SGR (Stay green) | Regulates plant chlorophyll degradation and senescence | Significantly improved lycopene content in tomato fruit | [120] | |
| SAPK2 | Primary mediator of ABA signaling | Enhanced sensitivity to drought stress and ROS in rice | [121] | |
| ARGOS8 | Negative regulator of ethylene responses | Enhanced drought tolerance and yield in maize | [122] | |
| SIMAPK3 | Participates in SA or JA defense-signaling pathways | Enhanced drought tolerance in tomato | [123] | |
| Metabolic Engineering for Biofortification of Vitamin A, Fe and Zn | ||||
| Vitamin A | Phytoene synthase (PSY) and phytoene desaturase(CrtI) gene | Participate in carotenoid biosynthetic pathway | Enhanced nutritional value of golden rice by increasing provitamin A content | [124] |
| Increase Increase total carotenoid content in transgenic wheat | [125] | |||
| Iron (Fe) | Soyfer H-1 | Soybean ferritin gene involved in storage of iron | Overexpression enhanced iron content in rice seed | [126] |
| OsNAS2 | Participates in iron-acquisition | Overexpression enhanced Fe and Zn content in rice endosperm | [127] | |
| Zinc (Zn) | HvNAS1 (Nicotianamine Synthas) | Metal chelator, involved in accumulation of Fe and Zn | Overexpressing enhanced Fe and Zn contents in the leaves, flowers, and seeds in rice | [128] |
| Metabolic Engineering for Abiotic Stress Tolerance | ||||
| Transcription Factor | TTG2 | WRKY TF regulates diverse biological processes | Regulate trichome development and enhance salinity tolerance in Brassica | [129] |
| ERF-2 (like) | Ethylene response TF, regulates various stress responses | Overexpression enhanced submergence tolerance in Arabidopsis | [130] | |
| NAC 19, 82 | TF plays important roles in development, abiotic, biotic stress responses, and biosynthesis | Overexpression led to regulate ROS and cell death in tobacco leaves | [131] | |
| HSFA4A | Heat shock transcription factor | Enhanced desiccation tolerance in seeds and activate antioxidant system in Arabidopsis | [132] | |
| CDF1 | Regulates expression of floral activator genes | Regulate flowering time and freezing tolerance in Arabidopsis | [133] | |
| Kinases | MAPKKK 4 | Regulates growth, development, and immune responses | Regulation of ROS induced cell death in tobacco leaves, lipid peroxidation, and DNA degradation | [134] |
| MAPKKK 18, 19 | Regulates plant immunity and hormone responses | Regulates ROS formation and cell death in tobacco | [135] | |
| CPK2 | Regulates cellular responses to various stimuli | Regulates ROS and cell death control through interaction with RbohD in tobacco | [136] | |
| MKK1 | Regulates stresses, growth, and development | Enhanced response of plants to pathogenic bacteria and drought stress in tobacco | [137] | |
| Transporters | SWEET | Plays important role in sucrose translocation and crop yields | Regulates plant growth and development and also participates in biotic and abiotic stress response | [138] |
| HMA | Heavy metal ATPase, response to Cd stress | Played an important role in Cd translocation in the leaves of Brassica napus | [139] | |
| ABC | Regulates uptake and allocation of metabolites and xenobiotics | Significantly induced under Cd stress and regulate ion channels | [140] | |
| AQPs (Aquaporins) | Facilitates molecule movement across the membranes | Overexpression enhances salt stress tolerance in transgenic tobacco | [141] | |
| Metabolic Engineering for Terpenoids/Volatile Compounds | ||||
| Monoterpenoids | Linalool synthase (LIS) | Catalyzes the formation of acyclic monoterpene linalool | Transgenic petunia plants result in the accumulation of S-linalyl-beta-D-glucopyranoside | [142] |
| Engineering of terpenoid pathway led enhanced aroma and flavor in tomato | [143] | |||
| Limonene Synthase | Catalyzes the cyclization of geranyl pyrophosphate to (4S)-limonene | Modified essential oil content in transgenic lines in transgenic mint | [144] | |
| β-Glucosidase | Catalyzes the hydrolysis of the glycosidic bonds and release glucose | Affects the emission of plant volatiles, plant-environment communication and aroma | [145] | |
| Sesquiterpenoids | Trichodiene synthase | Catalyzes the formation of trichodiene | Transgenic tobacco enhanced the expression of active enzyme and low-level accumulation of its sesquiterpenoid product | [146] |
| zingiberene synthase (ZIS) | Catalyzes the reaction forming zingiberene and other mono- and sesquiterpenes | Overexpression led to enhanced both mono-and sesquiterpene content in tomato fruit | [147] | |
| Germacrene A synthase | Key cytosolic enzyme of sesquiterpene lactone biosynthesis pathway | Transgenic lines with strong transgene expression showed growth retardation and FaNES1-expressing lines enhanced the resistance against the aphids | [148] | |
| Diterpenoids | Taxadiene synthase | Catalyzes the chemical reaction geranylgeranyl diphosphate | Enhanced level of toxoids was found in genetically engineering plant | [149] |
| Metabolic Engineering for Biotic Stress Tolerance | ||||
| Pathogen Perception | EFR (EF-Tu receptor) | Pattern recognition receptor (PRR), binds to prokaryotic protein EF-TU | Expression in susceptible genotypes reduced bacterial wilt incidence and enhanced yield | [150] |
| Bs2 | Bs2 gene is a member of the NBS-LRR class of R genes | Transgenic tomato conferred resistance to bacterial spot disease | [151] | |
| Pathogen Effector Binding | Os11N3/OsSWEET14 | Encode sucrose transporters | Transgenic wheat provided effective resistance to Fusarium graminearum | [152] |
| Xa27 | Important R-genes, effective against Xoo | Provided resistance to different strains of Xoo and bacterial leaf streak | [153] | |
| Defence Signaling Pathways | NPR1 | Master immune regulatory gene | Mediate broad-spectrum disease resistance without compromising plant fitness in Arabidopsis thaliana and rice | [154] |
| IPA1/OsSPL14 | Regulate rice plant architecture | Enhanced yield and disease resistance in rice | [155] | |
| Recessive Resistance Alleles | Mlo (Mildew Locus O) | Knockdown resulted in powdery mildew resistance | Loss of function mutation confer resistance to powdery mildew fungi | [156] |
| bs5 | Recessive genes resistant to bacterial spot | Confers disease resistance against Xanthomonas euvesicatoria in pepper and tomato | [157] | |
| Dominant Resistance Proteins | PFLP | Ferrodoxin like protein, involved in redox reactions | Overexpression induced hypersensitive reaction and resistance in tobacco | [158] |
| Lr34 | Wheat multipathogen resistant gene | Confer resistance to anthracnose and rust in sorghum | [159] | |
| Oxalate oxidase | Participates in degradation of oxalic acid | Enhanced resistance to Sclerotinia sclerotium in oilseed rape | [160] | |
| Antimicrobial Compound Production | Rs-AFP defensin (Raphanus sativus antifungal protein) | Antifungal plant defensins | Transgenic wheat conferred resistance to Fusarium graminearum and Rhizoctonia cerealis | [161] |
| Virus KP4 | Fungal killer toxin encoded by RNA virus | Transgenic wheat showed resistance to loose smut | [162] | |
| MsrA1 | Involved in mannan biosynthesis | Transgenic Brassica Juncea exhibited resistance to fungal phytopathogens | [163] | |
| RNAi Mediated | AC1 from bean golden mosaic virus | Modulates virus induced gene silencing | Transgenic common bean (Phaseolus vulgaris) conferred resistance to ban golden mosaic virus | [164] |
| Coat protein gene from potato virus Y | Protects RNA genome | Exhibited resistance to mixed virus infection in potato | [165] | |
| Crops | Genes with Donor Organism | Carotenoid Content | References |
|---|---|---|---|
| Rice | Narcissus pseudonarcissus (crtB) | Combination of transgenes enabled biosynthesis of provitamin A in the rice endosperm (Golden Rice 1) | [171] |
| Erwinia uredovora (crtI) | |||
| Zea mays (PSY) | Increase in total carotenoids up to 23-fold (Golden Rice II) | [124] | |
| Erwinia uredovora (crtI) | |||
| Wheat | Zea mays (PSY) | The total carotenoids content was increased up to 10-fold | [125] |
| Erwinia uredovora (crtI) | |||
| Erwinia uredovora (crtB, crtI) | Total carotenoid content increased by 8-fold and beta-carotene content increased by 65-fold | [181] | |
| Erwinia uredovora (crtB) | Increase in the beta-carotene content by 31-fold | [182] | |
| Triticum aestivum (HYD) | |||
| Potato | Pantoea ananatis (crtB) | Total carotenoid increased by 4-fold with major increase in beta-carotene and lutein content | [183] |
| Pantoea ananatis (crtE) | Total carotenoid up by 2.5-fold and beta-carotene content by 14-fold | [184] | |
| Pantoea ananatis (crtB, crtI, crtY) | Total carotenoid increased by 20-fold and that of beta-carotene by 3600-fold | [185] | |
| Solanum tuberosum (β-CHX) | Beta-carotene content was increased from trace level to 3.31 μg/g FW | [186] | |
| Brassica oleracea (Or) | Carotenoid content was increased by 10-fold | [177] | |
| Corn | Zea mays (PSY) | Increased level of beta-carotene content including hydroxy- and keto-carotenoids | [187] |
| Gentiana lutea (LCYE, β-CHX) | |||
| Paracoccus (crtW) | |||
| Pantoea ananatis (crtI) | |||
| Pantoea ananatis (crtB, crtI, zds) | Total carotenoids up by 34-fold with preferential accumulation of beta-carotene | [188] | |
| Zea mays (PSY) | The transgenic kernels contained 169-fold the normal amount of β-carotene | [189] | |
| Pantoea ananatis (crtI) | |||
| Tomato | Erwinia uredovora (crtI) | The β-carotene content increased about threefold, up to 45% of the total carotenoid content | [190] |
| Solanum lycopersicum (LCYB) | 7-fold increase in fruit beta-carotene content | [172] | |
| Arabidopsis thaliana (LCYB) | 12-fold increase in beta-carotene content along with beta-cryptoxanthin and zeaxanthin accumulation | [191] | |
| Capsicum annuum (β-CHX) | |||
| Erwinia uredovora (crtB) | Total fruit carotenoids upby 2–4-fold in fruits | [192] | |
| Solanum lycopersicum (LCYB) | Carotenoid content was increased by 2-fold while beta-carotene is up by 27-fold | [173] | |
| Arabidopsis thaliana (HMGR) | Total carotenoid content increased by 1.6-fold and beta-carotene by 2.2-fold | [193] | |
| Escherichia coli (dxs) | |||
| Capsicum annuum (FIB) | Total carotenoid content was up by 2-fold | [194] | |
| Narcissus pseudonarcissus (crtY) | 4.5-fold increase in beta-carotene and >50% increase in total carotenoid accumulation | [195] | |
| Citrus (LCYB1) | Beta-carotene level was increased by 4.1-fold, and the total carotenoid content increased by 30% in the fruits | [196] | |
| Cassava | Erwinia uredovora (crtB) | Total carotenoidcontent increase by 15-fold and that of beta-carotene by 37-fold | [197] |
| Arabidopsis thaliana (DXS) | |||
| Phytoene synthase | Total carotenoid content increased by 33-fold and beta-carotene by 15-fold | [198] | |
| Bacterial (crtB) | Total carotenoid content increased by 30-fold with beta-carotene accounting for 80–90% of total carotenoid content | [199] | |
| Arabidopsis thaliana (DXS) | |||
| Sorghum | Zea mays (PSY) | 24-fold increase in beta-carotene content | [200] |
| Pantoea ananatis (crtI) | |||
| Arabidopsis thaliana (DXS) | |||
| Hordeum vulgare (HGGT) | |||
| Melon | Or | Total carotenoid content increased by 11-fold | [176] |
| Cauliflower | Or | Beta-carotene content increased by 7-fold | [201] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, V.; Gupta, P.; Priscilla, K.; SharanKumar; Hangargi, B.; Veershetty, A.; Ramrao, D.P.; Suresh, S.; Narasanna, R.; Naik, G.R.; et al. Metabolomics Intervention Towards Better Understanding of Plant Traits. Cells 2021, 10, 346. https://doi.org/10.3390/cells10020346
Sharma V, Gupta P, Priscilla K, SharanKumar, Hangargi B, Veershetty A, Ramrao DP, Suresh S, Narasanna R, Naik GR, et al. Metabolomics Intervention Towards Better Understanding of Plant Traits. Cells. 2021; 10(2):346. https://doi.org/10.3390/cells10020346
Chicago/Turabian StyleSharma, Vinay, Prateek Gupta, Kagolla Priscilla, SharanKumar, Bhagyashree Hangargi, Akash Veershetty, Devade Pandurang Ramrao, Srinivas Suresh, Rahul Narasanna, Gajanana R. Naik, and et al. 2021. "Metabolomics Intervention Towards Better Understanding of Plant Traits" Cells 10, no. 2: 346. https://doi.org/10.3390/cells10020346
APA StyleSharma, V., Gupta, P., Priscilla, K., SharanKumar, Hangargi, B., Veershetty, A., Ramrao, D. P., Suresh, S., Narasanna, R., Naik, G. R., Kumar, A., Guo, B., Zhuang, W., Varshney, R. K., Pandey, M. K., & Kumar, R. (2021). Metabolomics Intervention Towards Better Understanding of Plant Traits. Cells, 10(2), 346. https://doi.org/10.3390/cells10020346