
Ellagic Acid and Schisandrins: Natural Biaryl Polyphenols with Therapeutic Potential to Overcome Multidrug Resistance in Cancer


Abstract
:1. Introduction
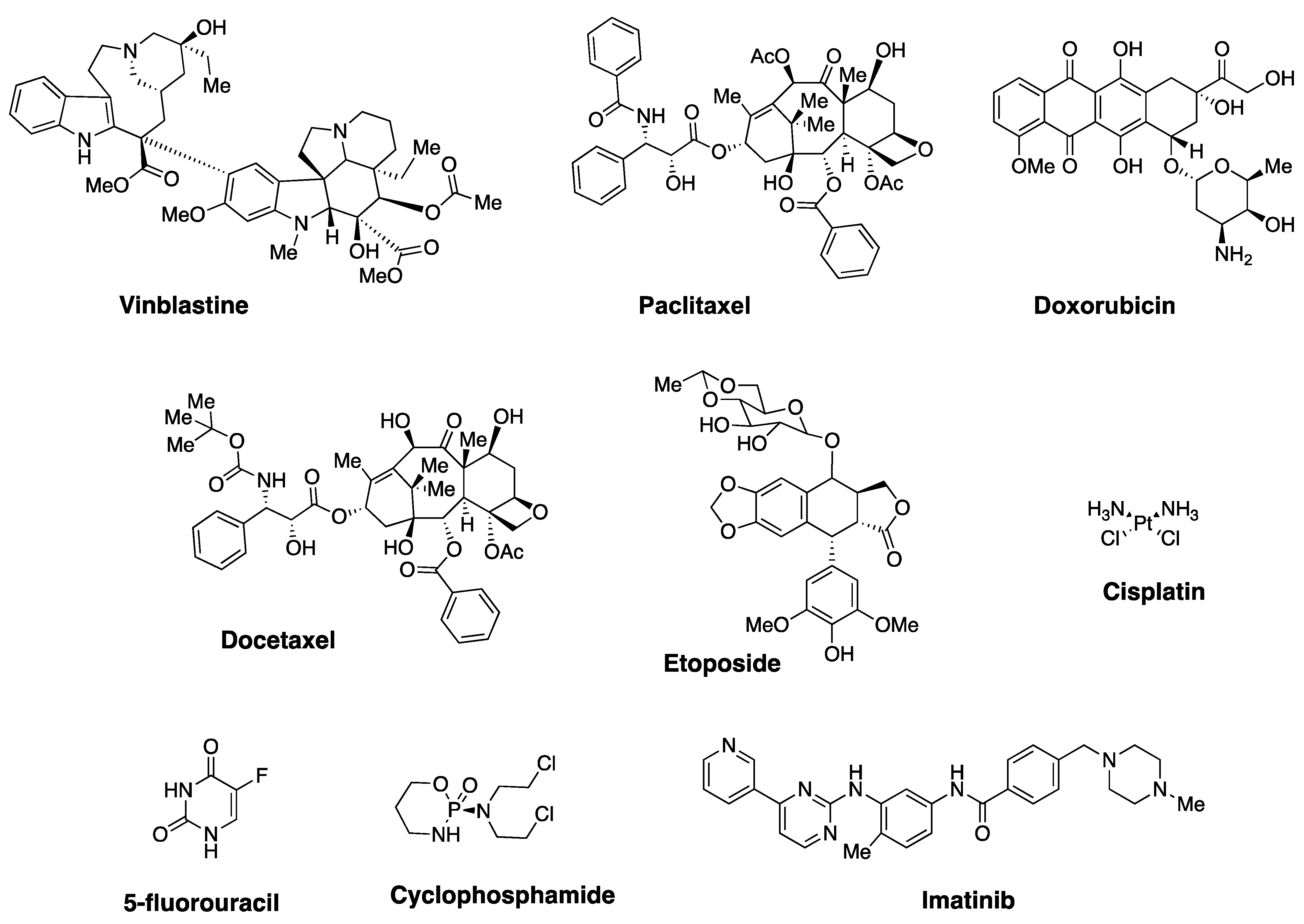
2. Natural Polyphenols and Their Anticancer Properties
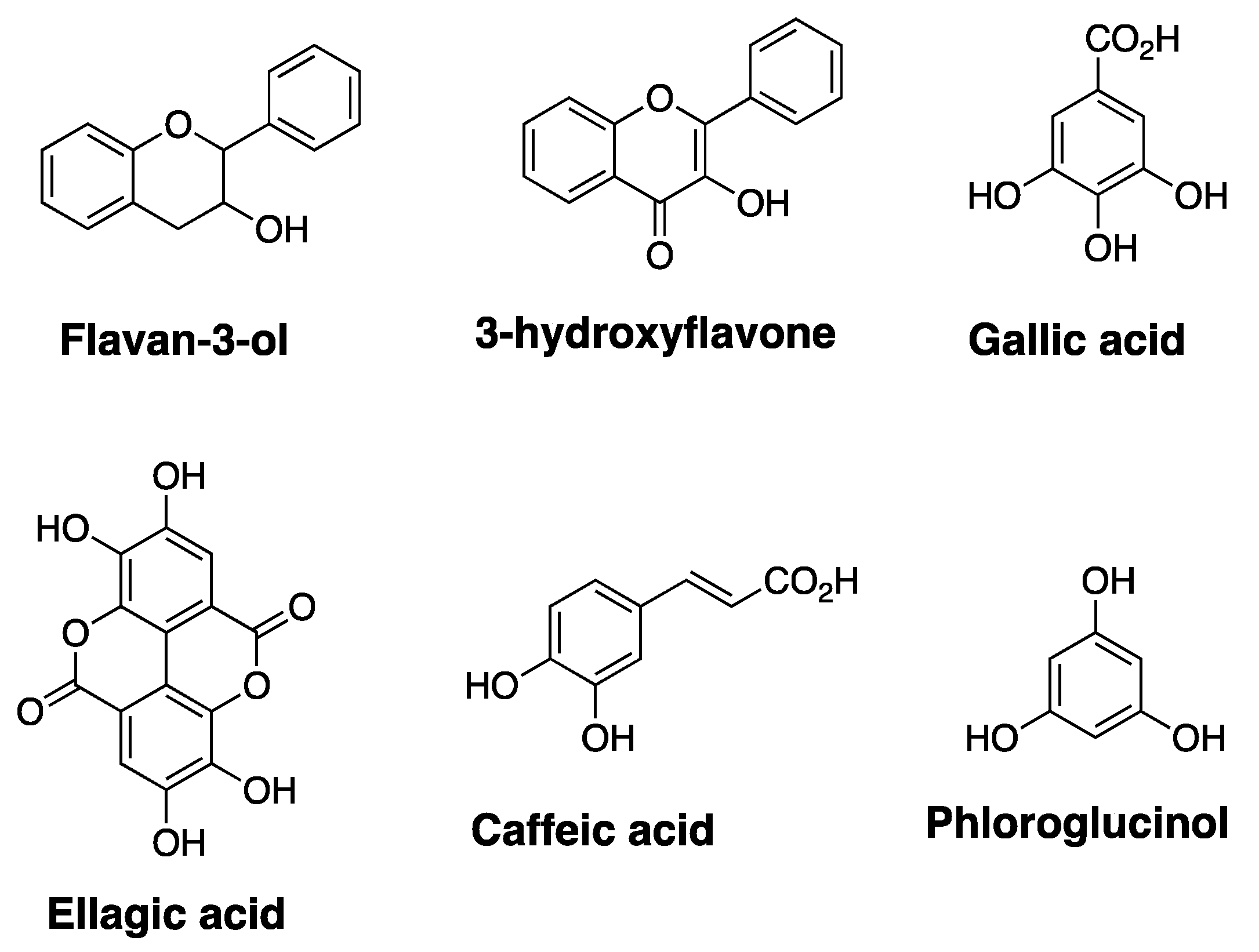
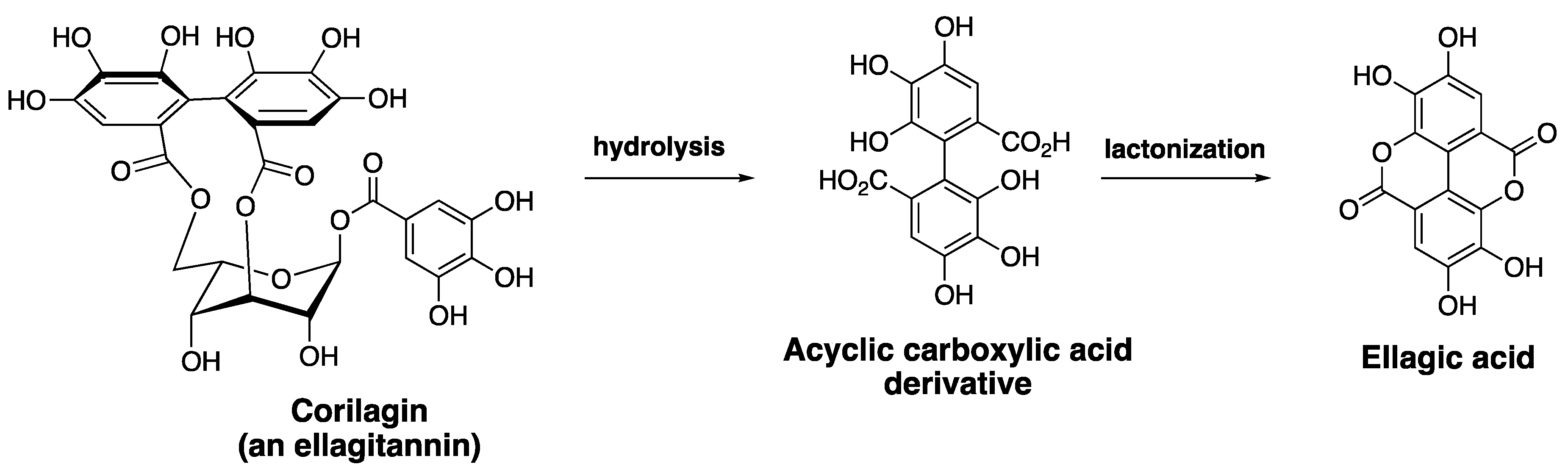
2.1. Ellagic Acid
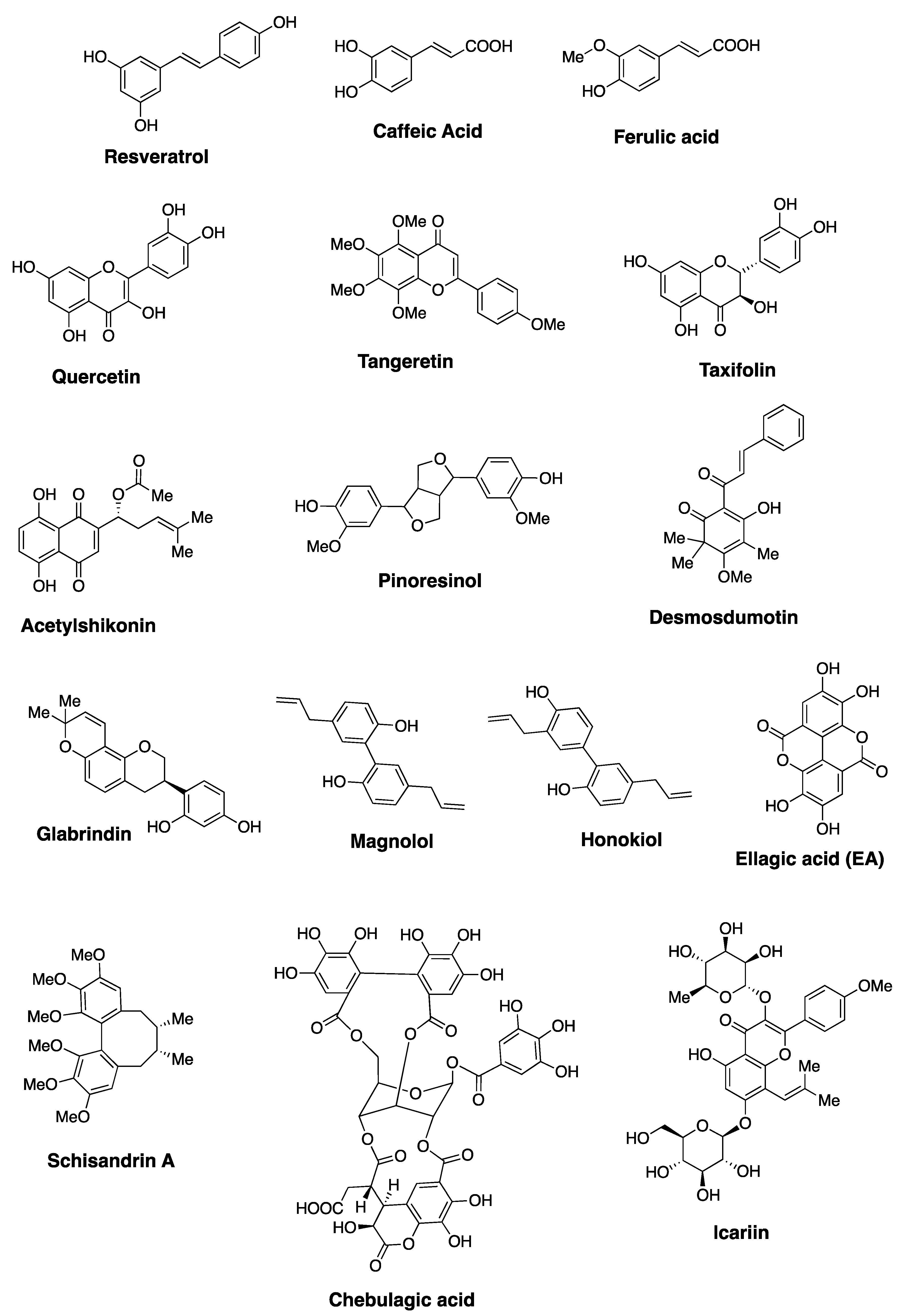
2.2. Schisandrins and Related Lignans
3. Atropisomerism in Drug Discovery
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, K.D.; Siegel, R.L.; Jemal, A. Cancer Statistics. CA Cancer J. Clin. 2018, 70, 7–30. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Lewis, L.D. Cancer pharmacotherapy: 21st century ’magic bullets’ and changing paradigms. Br. J. Clin. Pharmacol. 2006, 62, 1–4. [Google Scholar] [CrossRef]
- DeVita, V.T.; Chu, E. A History of Cancer Chemotherapy. Cancer Res. 2008, 68, 8643–8653. [Google Scholar] [CrossRef] [Green Version]
- Chabner, B.A.; Roberts, T.G., Jr. Timeline: Chemotherapy and the war on cancer. Nat. Rev. Cancer 2005, 5, 65–72. [Google Scholar] [CrossRef]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nat. Cell Biol. 2019, 575, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Sarah, C. Anticancer agents: Fighting resistance. Nat. Rev. Drug Discov. 2017, 16, 602. [Google Scholar] [CrossRef]
- Krishna, R.; Mayer, L.D. Multidrug resistance (MDR) in cancer. Mechanisms, reversal using modulators of MDR and the role of MDR modulators in influencing the pharmacokinetics of anticancer drugs. Eur. J. Pharm. Sci. 2000, 11, 265–283. [Google Scholar] [CrossRef]
- Yusuf, R.Z.; Duan, Z.; Lamendola, D.E.; Penson, R.T.; Seiden, M.V. Paclitaxel Resistance: Molecular Mechanisms and Pharmacologic Manipulation. Curr. Cancer Drug Targets 2003, 3, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Bachhuber, A.; Victor, A.; Steiner, E.; Mahlke, M.; Lehr, H.A.; Pilch, H.; Weikel, W.; Knapstein, P.G. p53 expression and resistance against paclitaxel in patients with metastatic breast cancer. J. Cancer Res. Clin. Oncol. 2003, 129, 295–302. [Google Scholar] [CrossRef]
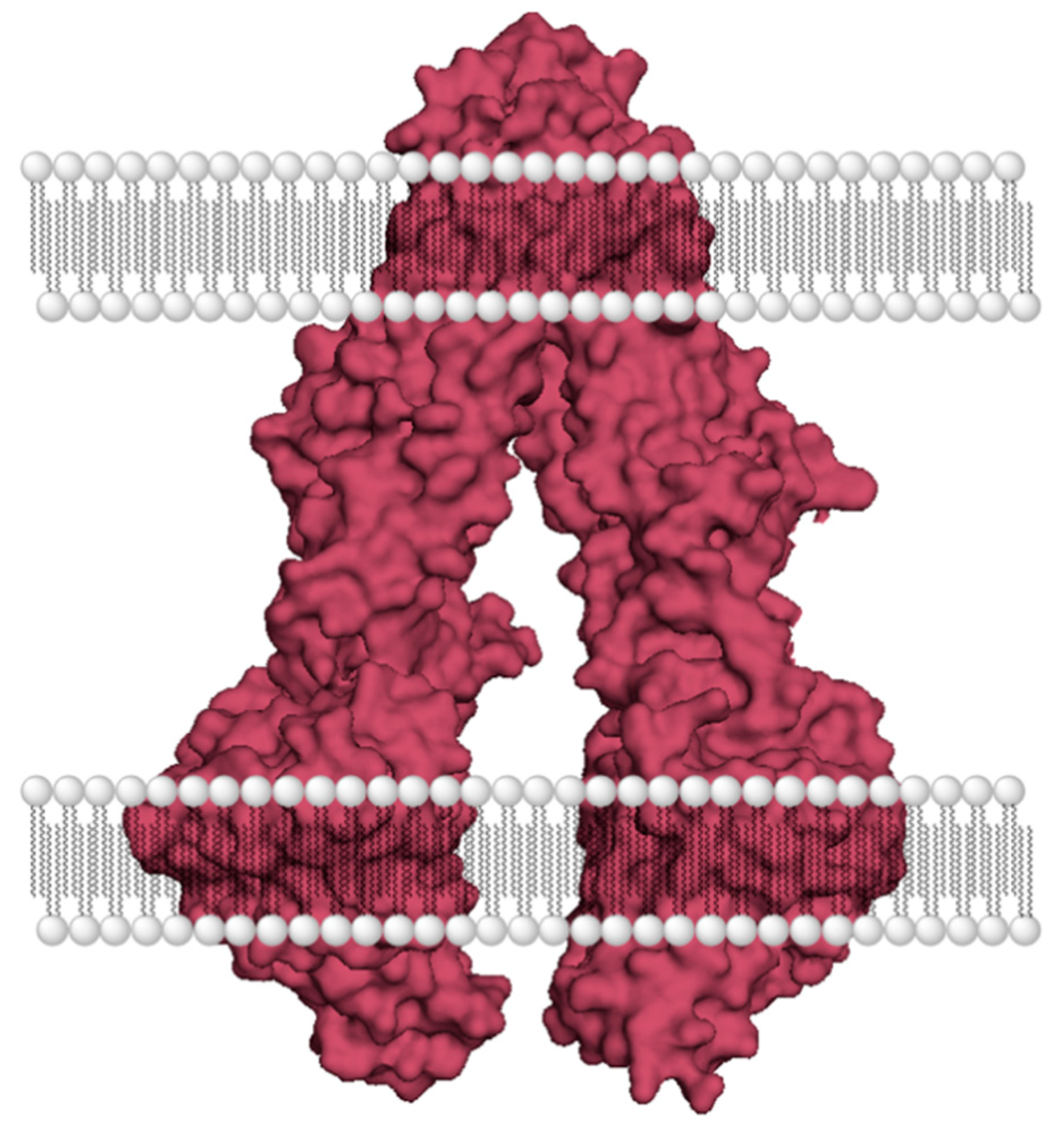
- Lepper, E.R.; Nooter, K.; Verweij, J.; Acharya, M.R.; Figg, W.D.; Sparreboom, A. Mechanisms of resistance to anticancer drugs: The role of the polymorphic ABC transporters ABCB1 and ABCG2. Pharmacogenomics 2005, 6, 115–138. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP–dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kathawala, R.J.; Gupta, P.; Ashby, C.R.; Chen, Z.-S. The modulation of ABC transporter-mediated multidrug resistance in cancer: A review of the past decade. Drug Resist. Updat. 2015, 18, 1–17. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Pastan, I. Biochemistry of multidrug resistance mediated by the multidrug transporter. Annu. Rev. Biochem. 1993, 62, 385–427. [Google Scholar] [CrossRef]
- Chen, Z.-S.; Tiwari, A.K. Multidrug resistance proteins (MRPs/ABCCs) in cancer chemotherapy and genetic diseases. FEBS J. 2011, 278, 3226–3245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locher, K.P. Structure and mechanism of ATP-binding cassette transporters. Philos. Trans. R. Soc. B Biol. Sci. 2008, 364, 239–245. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.; Tiriveedhi, V. Perplexing Role of P-Glycoprotein in Tumor Microenvironment. Front. Oncol. 2020, 10, 265. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Ling, V. The molecular basis of multidrug resistance in cancer: The early years of P-glycoprotein research. FEBS Lett. 2005, 580, 998–1009. [Google Scholar] [CrossRef] [Green Version]
- Darby, A.J.R.; Callaghan, R.; McMahon, M.R. P-glycoprotein inhibition: The past, the present and the future. Curr. Drug Metab. 2011, 12, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Dei, S.; Braconi, L.; Romanelli, M.N.; Teodori, E. Recent advances in the search of BCRP- and dual P-gp/BCRP-based multidrug resistance modulators. Cancer Drug Resist. 2019, 2, 710–743. [Google Scholar] [CrossRef] [Green Version]
- Horsey, A.J.; Cox, M.H.; Sarwat, S.; Kerr, I.D. The multidrug transporter ABCG2: Still more questions than answers. Biochem. Soc. Trans. 2016, 44, 824–830. [Google Scholar] [CrossRef] [Green Version]
- Beis, K. Structural basis for the mechanism of ABC transporters. Biochem. Soc. Trans. 2015, 43, 889–893. [Google Scholar] [CrossRef] [Green Version]
- Aller, S.G.; Yu, J.; Ward, A.; Weng, Y.; Chittaboina, S.; Zhuo, R.; Harrell, P.M.; Trinh, Y.T.; Zhang, Q.; Urbatsch, I.L.; et al. Structure of P-Glycoprotein Reveals a Molecular Basis for Poly-Specific Drug Binding. Science 2009, 323, 1718–1722. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Jaimes, K.F.; Aller, S.G. Refined structures of mouse P-glycoprotein. Protein Sci. 2014, 23, 34–46. [Google Scholar] [CrossRef]
- Alam, A.; Kung, R.; Kowal, J.; McLeod, R.A.; Tremp, N.; Broude, E.V.; Roninson, I.B.; Stahlberg, H.; Locher, K.P. Structure of a zosuquidar and UIC2-bound human-mouse chimeric ABCB1. Proc. Natl. Acad. Sci. USA 2018, 115, E1973–E1982. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xu, H.; Ashby, C.R., Jr.; Assaraf, Y.G.; Chen, Z.S.; Liu, H.M. Chemical molecular-based approach to overcome multidrug resistance in cancer by targeting P-glycoprotein (P-gp). Med. Res. Rev. 2021, 41, 525–555. [Google Scholar] [CrossRef] [PubMed]
- Dewanjee, S.; Dua, T.K.; Bhattacharjee, N.; Das, A.; Gangopadhyay, M.; Khanra, R.; Joardar, S.; Riaz, M.; De Feo, V.; Zia-UI-Haq, M. Natural products as alternative choices for P-glycoprotein (P-gp) inhibition. Molecules 2017, 22, 871. [Google Scholar] [CrossRef]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Qin, Z.; Zhang, W.-D.; Cheng, G.; Yehuda, A.G.; Ashby, C.R.; Chen, Z.-S.; Cheng, X.-D.; Qin, J.-J. Medicinal chemistry strategies to discover P-glycoprotein inhibitors: An update. Drug Resist. Updat. 2020, 49, 100681. [Google Scholar] [CrossRef]
- Abdallah, H.M.; Al-Abd, A.M.; El-Dine, R.S.; El-Halawany, A.M. P-glycoprotein inhibitors of natural origin as potential tumor chemo-sensitizers: A review. J. Adv. Res. 2015, 6, 45–62. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, S.; Hoti, S.L. Development of Fourth Generation ABC Inhibitors from Natural Products: A Novel Approach to Overcome Cancer Multidrug Resistance. Anti-Cancer Agents Med. Chem. 2015, 15, 605–615. [Google Scholar] [CrossRef]
- Creemers, S.G.; Van Koetsveld, P.M.; De Herder, W.W.; Dogan, F.; Franssen, G.J.H.; A Feelders, R.; Hofland, L.J. MDR1 inhibition increases sensitivity to doxorubicin and etoposide in adrenocortical cancer. Endocr. Relat. Cancer 2019, 26, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Warner, E.; Tobe, S.W.; Andrulis, I.L.; Pei, Y.; Trachtenberg, J.; Skorecki, K.L. Phase I-II Study of Vinblastine and Oral Cyclosporin A in Metastatic Renal Cell Carcinoma. Am. J. Clin. Oncol. 1995, 18, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Pusztai, L.; Wagner, P.; Ibrahim, N.; Rivera, E.; Theriault, R.; Booser, D.; Symmans, F.W.; Wong, F.; Blumenschein, G.; Fleming, D.R.; et al. Phase II study of tariquidar, a selective P-glycoprotein inhibitor, in patients with chemotherapy-resistant, advanced breast carcinoma. Cancer 2005, 104, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.-I.; Tseng, Y.-J.; Chen, M.-H.; Huang, C.-Y.F.; Chang, P.M.-H. Clinical perspective of FDA approved drugs with P-glycoprotein inhibition activities for potential cancer therapeutics. Front. Oncol. 2020, 10, 561936. [Google Scholar] [CrossRef] [PubMed]
- Zu, Y.; Yang, Z.; Tang, S.; Han, Y.; Ma, J. Effects of P-Glycoprotein and Its Inhibitors on Apoptosis in K562 Cells. Molecules 2014, 19, 13061–13075. [Google Scholar] [CrossRef] [PubMed]
- Nanayakkara, A.K.; Vogel, P.D.; Wise, J.G. Prolonged inhibition of P-glycoprotein after exposure to chemotherapeutics increases cell mortality in multidrug resistant cultured cancer cells. PLoS ONE 2019, 14, e0217940. [Google Scholar] [CrossRef]
- Binkhathlan, Z.; Lavasanifar, A. P-glycoprotein inhibition as a therapeutic approach for overcoming multidrug resistance in cancer: Current status and future perspectives. Curr. Cancer Drug Targets 2013, 13, 326–346. [Google Scholar] [CrossRef]
- Gupta, P.; Gao, H.-L.; Ashar, Y.V.; Karadkhelkar, N.M.; Yoganathan, S.; Chen, Z.-S. Ciprofloxacin Enhances the Chemosensitivity of Cancer Cells to ABCB1 Substrates. Int. J. Mol. Sci. 2019, 20, 268. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Liu, X.; Chow, L.M.C. Flavonoids as P-gp Inhibitors: A Systematic Review of SARs. Curr. Med. Chem. 2019, 26, 4799–4831. [Google Scholar] [CrossRef]
- Mohana, S.; Ganesan, M.; Agilan, B.; Karthikeyan, R.; Srithar, G.; Mary, R.B.; Ananthakrishnan, D.; Velmurugan, D.; Prasad, N.R.; Ambudkar, S.V. Screening dietary flavonoids for the reversal of P-glycoprotein-mediated multidrug resistance in cancer. Mol. BioSyst. 2016, 12, 2458–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J. Predominately Uncultured Microbes as Sources of Bioactive Agents. Front. Microbiol. 2016, 7, 1832. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Marine-Sourced Anti-Cancer and Cancer Pain Control Agents in Clinical and Late Preclinical Development. Mar. Drugs 2014, 12, 255–278. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Li, J.W.-H.; Vederas, J.C. Drug Discovery and Natural Products: End of an Era or an Endless Frontier? Science 2009, 325, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Maier, M.E. Design and synthesis of analogues of natural products. Org. Biomol. Chem. 2015, 13, 5302–5343. [Google Scholar] [CrossRef] [Green Version]
- Yoganathan, S.; Yin, N.; He, Y.; Mesleh, M.F.; Gu, Y.G.; Miller, S.J. An efficient chemical synthesis of carboxylate-isostere analogs of daptomycin. Org. Biomol. Chem. 2013, 11, 4680–4685. [Google Scholar] [CrossRef] [Green Version]
- Yoganathan, S.; Miller, S.J. Structure Diversification of Vancomycin through Peptide-Catalyzed, Site-Selective Lipidation: A Catalysis-Based Approach to Combat Glycopeptide-Resistant Pathogens. J. Med. Chem. 2015, 58, 2367–2377. [Google Scholar] [CrossRef] [Green Version]
- Herrera, S. Synthetic biology offers alternative pathways to natural products. Nat. Biotechnol. 2005, 23, 270–271. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant Polyphenols: Chemical Properties, Biological Activities, and Synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-J.; Kim, O.H.; Kim, Y.H.; Lim, J.H.; Kim, S.; Park, J.-W.; Kwon, T.K. Quercetin arrests G2/M phase and induces caspase-dependent cell death in U937 cells. Cancer Lett. 2006, 240, 234–242. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Khan, I.A.; Ur-Rehman, M.-; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phytotherapy Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef]
- Brito, A.F.; Ribeiro, M.; Abrantes, A.M.; Pires, A.S.; Teixo, R.J.; Tralhao, J.G.; Botelho, M.F. Quercetin in Cancer Treatment, Alone or in Combination with Conventional Therapeutics? Curr. Med. Chem. 2015, 22, 3025–3039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.-Z.; Qiao, S.-F.; Zhang, J.-H.; Li, K. Quercetin Increase the Chemosensitivity of Breast Cancer Cells to Doxorubicin Via PTEN/Akt Pathway. Anti-Cancer Agents Med. Chem. 2015, 15, 1185–1189. [Google Scholar] [CrossRef] [PubMed]
- Hadjeri, M.; Barbier, M.; Ronot, X.; Mariotte, A.-M.; Boumendjel, A.; Boutonnat, J. Modulation of P-Glycoprotein-Mediated Multidrug Resistance by Flavonoid Derivatives and Analogues. J. Med. Chem. 2003, 46, 2125–2131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, X.; Morris, M.E. Flavonoids Are Inhibitors of Breast Cancer Resistance Protein (ABCG2)-Mediated Transport. Mol. Pharmacol. 2004, 65, 1208–1216. [Google Scholar] [CrossRef] [Green Version]
- Quideau, S.; Feldman, K.S. Ellagitannin Chemistry. Chem. Rev. 1996, 96, 475–504. [Google Scholar] [CrossRef]
- Feldman, K.S. Recent progress in ellagitannin chemistry. Phytochemistry 2005, 66, 1984–2000. [Google Scholar] [CrossRef] [PubMed]
- Smyth, J.E.; Butler, N.M.; Keller, P.A. A twist of nature—The significance of atropisomers in biological systems. Nat. Prod. Rep. 2015, 32, 1562–1583. [Google Scholar] [CrossRef] [Green Version]
- Toenjes, S.T.; Gustafson, J.L. Atropisomerism in medicinal chemistry: Challenges and opportunities. Future Med. Chem. 2018, 10, 409–422. [Google Scholar] [CrossRef]
- Ríos, J.-L.; Giner, R.M.; Marín, M.; Recio, M.C. A Pharmacological Update of Ellagic Acid. Planta Medica 2018, 84, 1068–1093. [Google Scholar] [CrossRef] [Green Version]
- Achari, C.; Reddy, V.G.; Reddy, T.C.M.; Reddanna, P. Chebulagic acid synergizes the cytotoxicity of doxorubicin in human hepatocellular carcinoma through COX–2 dependant modulation of MDR-1. Med. Chem. 2011, 7, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Evtyugin, D.D.; Magina, S.; Evtuguin, D.V. Recent Advances in the Production and Applications of Ellagic Acid and Its Derivatives. A Review. Molecules 2020, 25, 2745. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A. Ellagic acid in suppressing in vivo and in vitro oxidative stresses. Mol. Cell. Biochem. 2018, 448, 27–41. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.A.; Selma, M.V.; García-Conesa, M.T.; Espín, J.C. Urolithins, the rescue of “old” metabolites to understand a “new” concept: Metabotypes as a nexus among phenolic metabolism, microbiota dysbiosis, and host health status. Mol. Nutr. Food Res. 2017, 61, 1500901. [Google Scholar] [CrossRef]
- Gupta, P.; Mohammad, T.; Khan, P.; Alajmi, M.F.; Hussain, A.; Rehman, M.T.; Hassan, M.I. Evaluation of ellagic acid as an inhibitor of sphingosine kinase 1: A targeted approach towards anticancer therapy. Biomed. Pharmacother. 2019, 118, 109245. [Google Scholar] [CrossRef]
- Ceci, C.; Tentori, L.; Atzori, M.G.; Lacal, P.M.; Bonanno, E.; Scimeca, M.; Cicconi, R.; Mattei, M.; de Martino, M.G.; Vespasiani, G.; et al. Ellagic Acid Inhibits Bladder Cancer Invasiveness and In Vivo Tumor Growth. Nutrients 2016, 8, 744. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liang, X.; Niu, C.; Wang, X. Ellagic acid promotes A549 cell apoptosis via regulating the phosphoinositide 3-kinase/protein kinase B pathway. Exp. Ther. Med. 2018, 16, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, R.; Mohammad, T.; Gupta, P.; Haque, A.; Alajmi, M.F.; Hussain, A.; Hassan, M.I. Molecular interaction studies on ellagic acid for its anticancer potential targeting pyruvate dehydrogenase kinase 3. RSC Adv. 2019, 9, 23302–23315. [Google Scholar] [CrossRef] [Green Version]
- Gulzar, M.; Syed, S.B.; Khan, F.I.; Khan, P.; Ali, S.; Hasan, G.M.; Taneja, P.; Hassan, I. Elucidation of interaction mechanism of ellagic acid to the integrin linked kinase. Int. J. Biol. Macromol. 2019, 122, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Sachs, J.; Döhl, K.; Weber, A.; Bonus, M.; Ehlers, F.; Fleischer, E.; Klinger, A.; Gohlke, H.; Pietruszka, J.; Schmitt, L.; et al. Novel 3,4-Dihydroisocoumarins Inhibit Human P-gp and BCRP in Multidrug Resistant Tumors and Demonstrate Substrate Inhibition of Yeast Pdr5. Front. Pharmacol. 2019, 10, 400. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Sheng, D.; Tan, Q. Determination of Schisandrin A and Schisandrin B in Traditional Chinese Medicine Preparation Huganpian Tablet by RP-HPLC. Chem. Pharm. Bull. 2019, 67, 713–716. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Reiner, J.; Xie, J. Progress on the Chemistry of Dibenzocyclooctadiene Lignans. Chem. Rev. 2005, 105, 4581–4609. [Google Scholar] [CrossRef]
- Wei, M.; Liu, Y.; Pi, Z.; Li, S.; Hu, M.; He, Y.; Yue, K.; Liu, T.; Liu, Z.; Song, F.; et al. Systematically Characterize the Anti-Alzheimer’s Disease Mechanism of Lignans from S. chinensis based on In-Vivo Ingredient Analysis and Target-Network Pharmacology Strategy by UHPLC⁻Q-TOF-MS. Molecules 2019, 24, 1203. [Google Scholar] [CrossRef] [Green Version]
- Ding, Q.; Li, X.; Sun, Y.; Zhang, X. Schizandrin A inhibits proliferation, migration and invasion of thyroid cancer cell line TPC-1 by down regulation of microRNA-429. Cancer Biomarkers 2019, 24, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Guo, M. Schizandrin A inhibits cellular phenotypes of breast cancer cells by repressing miR-155. IUBMB Life 2020, 72, 1640–1648. [Google Scholar] [CrossRef]
- Chen, B.-C.; Tu, S.-L.; Zheng, B.-A.; Dong, Q.-J.; Wan, Z.-A.; Dai, Q.-Q. Schizandrin A exhibits potent anticancer activity in colorectal cancer cells by inhibiting heat shock factor 1. Biosci. Rep. 2020, 40, 20200203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Rajamanicham, V.; Xu, S.; Liu, Z.; Yan, T.; Liang, G.; Guo, G.; Zhou, H.; Wang, Y. Schisandrin A inhibits triple negative breast cancer cells by regulating Wnt/ER stress signaling pathway. Biomed. Pharmacother. 2019, 115, 108922. [Google Scholar] [CrossRef] [PubMed]
- Nasser, M.I.; Zhu, S.; Chen, C.; Zhao, M.; Huang, H.; Zhu, P. A Comprehensive Review on Schisandrin B and Its Biological Properties. Oxidative Med. Cell. Longev. 2020, 2020, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Xian, H.; Feng, W.; Zhang, J. Schizandrin A enhances the efficacy of gefitinib by suppressing IKKbeta/NF-kappaB signaling in non-small cell lung cancer. Eur. J. Pharmacol. 2019, 855, 10–19. [Google Scholar] [CrossRef]
- Zhang, Z.-L.; Jiang, Q.-C.; Wang, S.-R. Schisandrin A reverses doxorubicin-resistant human breast cancer cell line by the inhibition of P65 and Stat3 phosphorylation. Breast Cancer 2018, 25, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Zhang, D.; Chu, X.; Wang, J. Schizandrin A enhances chemosensitivity of colon carcinoma cells to 5-fluorouracil through up-regulation of miR-195. Biomed. Pharmacother. 2018, 99, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Nasser, M.I.; Han, T.; Adlat, S.; Tian, Y.; Jiang, N. Inhibitory effects of Schisandrin B on human prostate cancer cells. Oncol. Rep. 2018, 41, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-N.; Zhang, C.-Y.; Jin, X.-D.; Li, Y.-Z.; Zheng, X.-Z.; Li, L. Inhibitory effect of schisandrin B on gastric cancer cells in vitro. World J. Gastroenterol. 2007, 13, 6506–6511. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Guo, G.; Yin, C.; Zhang, Y.; Zhao, C.; Qian, J.; Wang, O.; Zhang, X.; Liang, G. Schisandrin B exhibits potent anticancer activity in triple negative breast cancer by inhibiting STAT3. Toxicol. Appl. Pharmacol. 2018, 358, 110–119. [Google Scholar] [CrossRef]
- Xiang, S.-S.; Wang, X.-A.; Li, H.-F.; Shu, Y.-J.; Bao, R.-F.; Zhang, F.; Cao, Y.; Ye, Y.-Y.; Weng, H.; Wu, W.-G.; et al. Schisandrin B Induces Apoptosis and Cell Cycle Arrest of Gallbladder Cancer Cells. Molecules 2014, 19, 13235–13250. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhang, Q.; Bao, J.; Du, C.; Wang, J.; Tong, Q.; Liu, C. Schisandrin B inhibits the proliferation and invasion of glioma cells by regulating the HOTAIR-micoRNA-125a-mTOR pathway. Neuroreport 2017, 28, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lu, Q.; Shen, Y.; Hu, X. Schisandrin B enhances doxorubicin-induced apoptosis of cancer cells but not normal cells. Biochem. Pharmacol. 2006, 71, 584–595. [Google Scholar] [CrossRef]
- Wang, S.; Wang, A.; Shao, M.; Lin, L.; Li, P.; Wang, Y. Schisandrin B reverses doxorubicin resistance through inhibiting P-glycoprotein and promoting proteasome-mediated degradation of survivin. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Xu, X.; Lu, Q.; Pan, Q.; Hu, X. Schisandrin B: A dual inhibitor of P-glycoprotein and multidrug resistance-associated protein 1. Cancer Lett. 2007, 246, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Qiangrong, P.; Wang, T.; Lu, Q.; Hu, X. Schisandrin B—A novel inhibitor of P-glycoprotein. Biochem. Biophys. Res. Commun. 2005, 335, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Kiso, Y.; Tohkin, M.; Hikino, H.; Ikeya, Y.; Taguchi, H. Mechanism of Antihepatotoxic Activity of Wuweizisu C and Gomisin A. Planta Medica 1985, 51, 331–334. [Google Scholar] [CrossRef]
- Lu, Y.H.; Lian, X.H.; Wei, D.Z.; Wang, Z.T. Activity of Schisandrin C Isolated from Schisandra chinensis against Human Cancer Cell Lines. Pharm. Biol. 2008, 46, 906–913. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Choi, Y.W.; Hyun, S.K.; Kwon, H.J.; Hwang, H.J.; Kim, G.Y.; Choi, B.T.; Kim, B.W.; Choi, I.W.; Moon, S.K.; et al. Induction of G1 arrest and apoptosis by schisandrin C isolated from Schizandra chinensis Baill in human leukemia U937 cells. Int. J. Mol. Med. 2009, 24, 495–502. [Google Scholar] [CrossRef] [Green Version]
- Amujuri, D.; Siva, B.; Poornima, B.; Sirisha, K.; Sarma, A.V.S.; Nayak, V.L.; Tiwari, A.K.; Purushotham, U.; Babu, K.S. Synthesis and biological evaluation of Schizandrin derivatives as potential anti-cancer agents. Eur. J. Med. Chem. 2018, 149, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.D.; Siva, B.; Bharathi, K.; Devi, A.; Kumar, P.P.; Anusha, K.; Lambhate, S.; Karunakar, T.; Tiwari, A.K.; Babu, K.S. Synthesis and biological evaluation of Schizandrin derivatives as tubulin polymerization inhibitors. Bioorg. Med. Chem. Lett. 2020, 30, 127354. [Google Scholar] [CrossRef] [PubMed]
- LaPlante, S.R.; Edwards, P.J.; Fader, L.D.; Jakalian, A.; Hucke, O. Revealing atropisomer axial chirality in drug discovery. ChemMedChem 2011, 6, 505–513. [Google Scholar] [CrossRef]
- Clayden, J.; Moran, W.J.; Edwards, P.J.; Laplante, S.R. The Challenge of Atropisomerism in Drug Discovery. Angew. Chem. Int. Ed. 2009, 48, 6398–6401. [Google Scholar] [CrossRef]
- Glunz, P.W. Recent encounters with atropisomerism in drug discovery. Bioorg. Med. Chem. Lett. 2018, 28, 53–60. [Google Scholar] [CrossRef]
- Zask, A.; Murphy, J.; Ellestad, G.A. Biological Stereoselectivity of Atropisomeric Natural Products and Drugs. Chirality 2013, 25, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Pietra, F. Why colchicine does not show mutarotation. With M05-2X density functional in the realm of tricky natural products. J. Phys. Org. Chem. 2007, 20, 1102–1107. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenol | Cancer Cell Line | Tumor Type | Anticancer Mechanism | Ref. |
|---|---|---|---|---|
| Ellagic acid | A549 | non-small cell lung cancer | SphK1 inhibition | [67] |
| PI3K/Akt pathway and PDK3 activity | [69,70] | |||
| T24, UM-UC-3, 5637, HT-1376 | bladder cancer | VEGF-mediated pathway | [68] | |
| HepG2 | liver cancer | PI3K/Akt pathway and PDK3 activity | [69,70] | |
| MCF-7 | breast cancer | PI3K/Akt pathway and PDK3 activity | [69,70] | |
| integrin-linked kinase (ILK) inhibition | [71] | |||
| Sch A | MDA-MB-231, BT-549 | breast cancer | cell cycle arrest/apoptosis via Wnt/ER pathway | [79] |
| RKO, DLD-1, SW620, SW480, HCT-116 | colorectal cancer | cell cycle arrest/apoptosis; cytotoxicity is via heat shock protein function | [78] | |
| increased sensitivity to 5-FU via PI3K/Akt and NF-kB signaling | [83] | |||
| HCC827/GR | Gefitinib-resistant lung cancer | inhibition of IKKb/NF-kB signaling | [81] | |
| MCF-7/Dox | Dox-resistant breast cancer | reversal of P-gp mediated MDR via Stat3 and NF-kB pathway | [82] | |
| Sch B | MDA-MB-231, BT-549, MDA-MB-468 | breast cancer | cell cycle arrest/apoptosis via Stat3 pathway | [86] |
| SMMC7721 | liver cancer | enhanced the anticancer activity of Dox | [89] | |
| MCF-7 | breast cancer | enhanced the anticancer activity of Dox | [89] | |
| MCF-7/ADR | Dox-resistant breast cancer | decreased P-gp expression and P-gp mediated efflux | [90,92] | |
| A2780/Dox | Dox-resistant ovarian cancer | decreased P-gp expression and P-gp mediated efflux | [90] | |
| HL60/ADR, HL60/MRP, K562/ADR | drug-resistant leukemia | sensitized the cells to daunorubicin; reversed P-gp mediated drug resistance | [91,92] | |
| Bcap37/ADR | drug-resistant breast cancer | reversed P-gp mediated drug resistance | [92] | |
| KBv200 | drug-resistant epidermoid carcinoma | reversed P-gp mediated drug resistance | [92] | |
| Sch C | Bel-7402 | liver cancer | undisclosed mechanism | [94] |
| Bcap37 | breast cancer | undisclosed mechanism | [94] | |
| KB-3-1 | nasopharyngeal cancer | undisclosed mechanism | [94] | |
| U937 | leukemia | cell cycle arrest and apoptosis | [95] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoganathan, S.; Alagaratnam, A.; Acharekar, N.; Kong, J. Ellagic Acid and Schisandrins: Natural Biaryl Polyphenols with Therapeutic Potential to Overcome Multidrug Resistance in Cancer. Cells 2021, 10, 458. https://doi.org/10.3390/cells10020458
Yoganathan S, Alagaratnam A, Acharekar N, Kong J. Ellagic Acid and Schisandrins: Natural Biaryl Polyphenols with Therapeutic Potential to Overcome Multidrug Resistance in Cancer. Cells. 2021; 10(2):458. https://doi.org/10.3390/cells10020458
Chicago/Turabian StyleYoganathan, Sabesan, Anushan Alagaratnam, Nikita Acharekar, and Jing Kong. 2021. "Ellagic Acid and Schisandrins: Natural Biaryl Polyphenols with Therapeutic Potential to Overcome Multidrug Resistance in Cancer" Cells 10, no. 2: 458. https://doi.org/10.3390/cells10020458
APA StyleYoganathan, S., Alagaratnam, A., Acharekar, N., & Kong, J. (2021). Ellagic Acid and Schisandrins: Natural Biaryl Polyphenols with Therapeutic Potential to Overcome Multidrug Resistance in Cancer. Cells, 10(2), 458. https://doi.org/10.3390/cells10020458