Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. CAM Assay
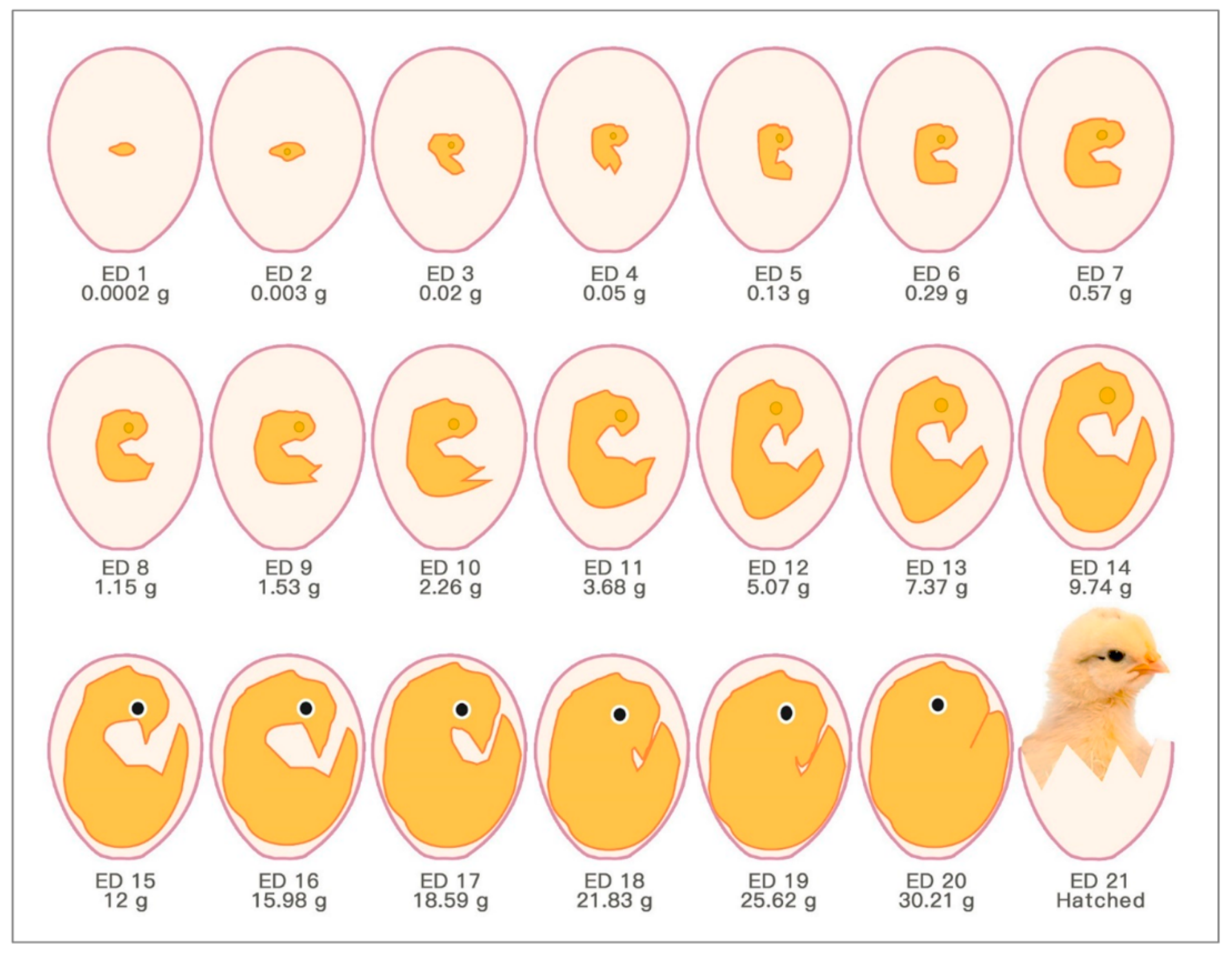
2.1. Development of Fertilised Chick Eggs in Incubation
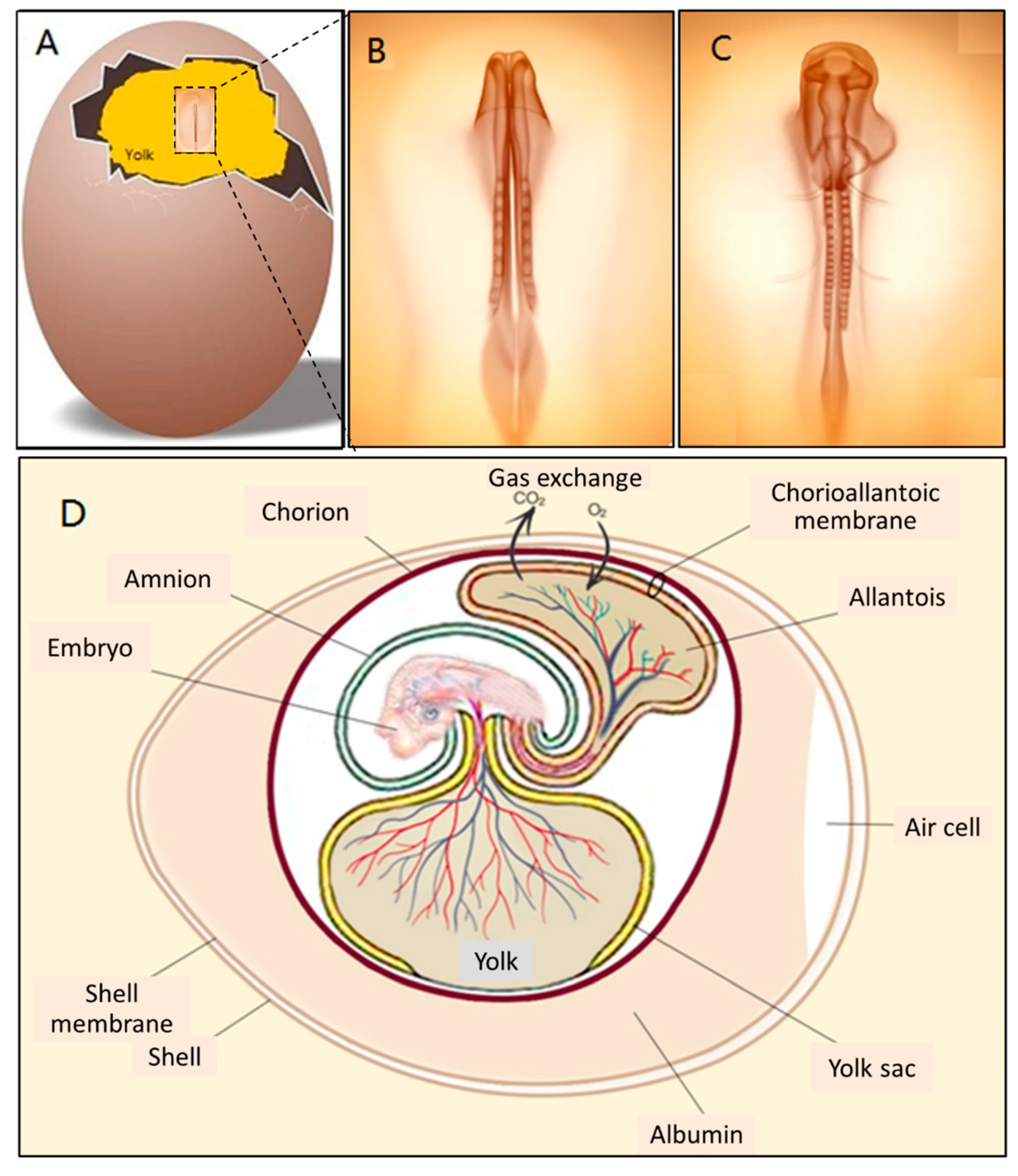
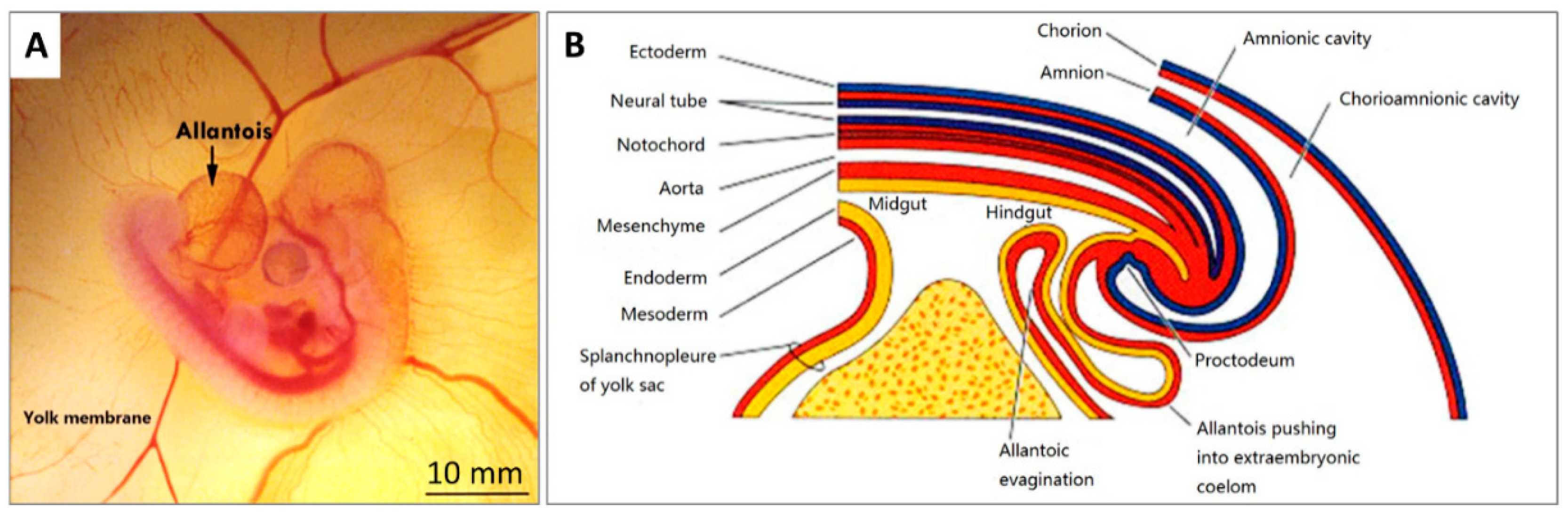
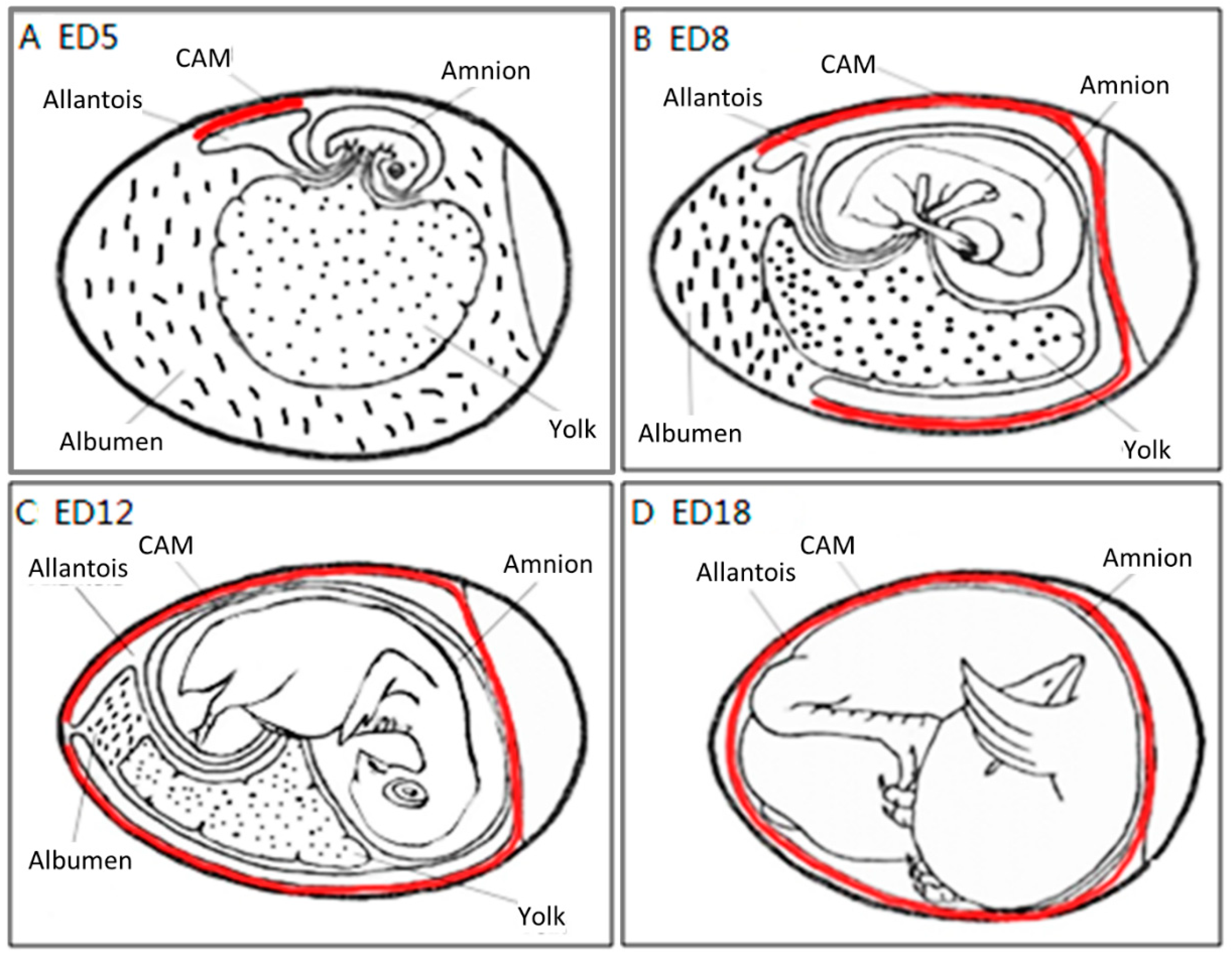
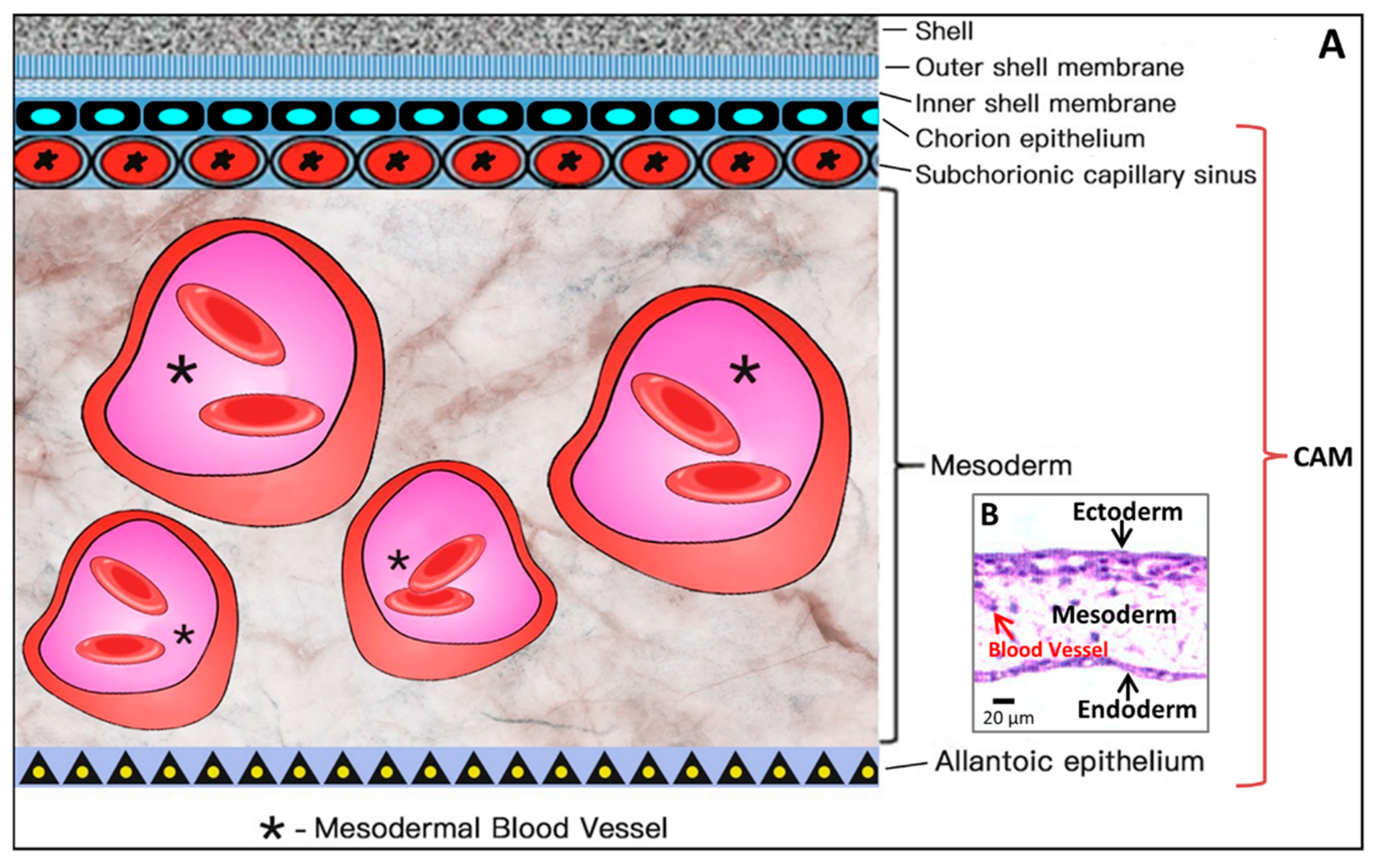
2.2. CAM Development and Physiological Function
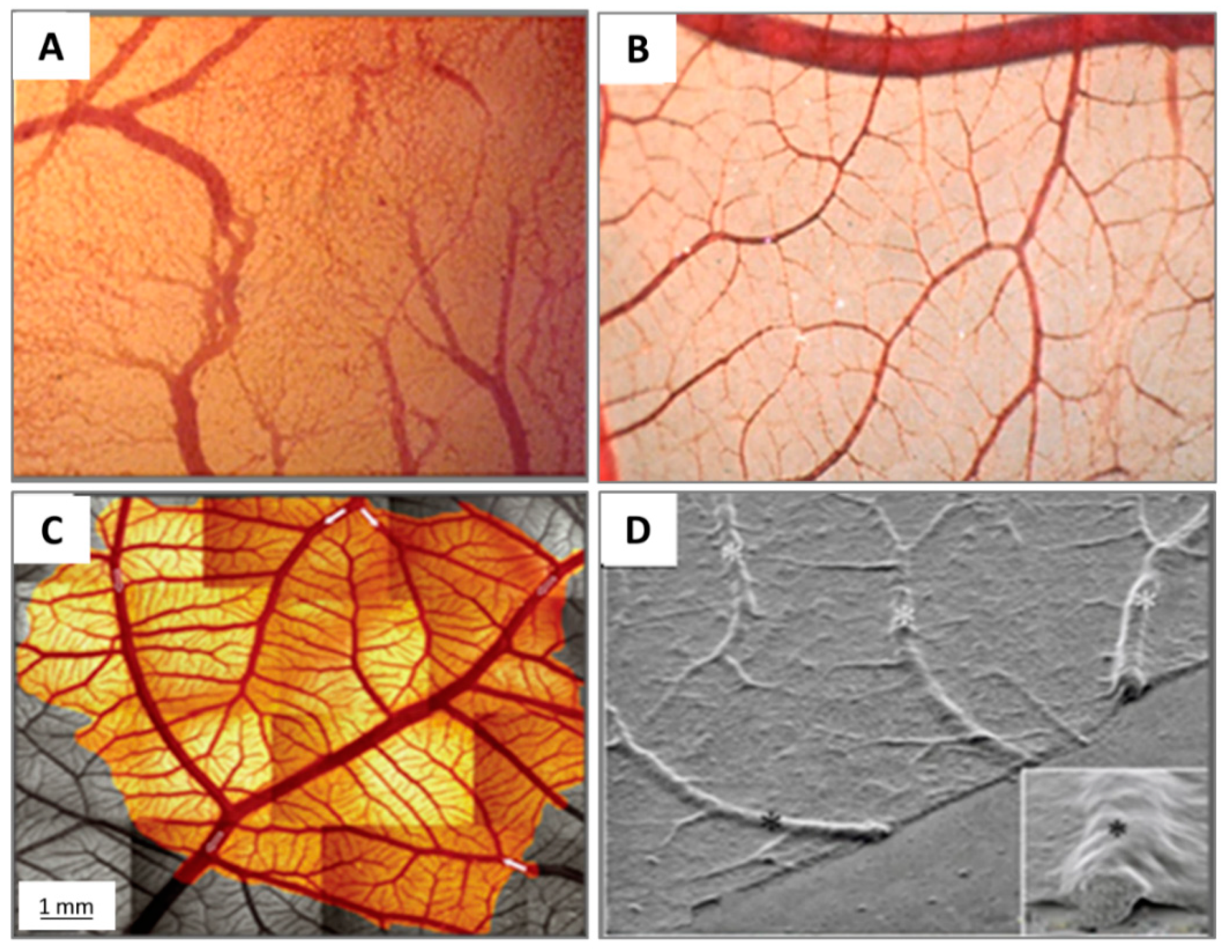
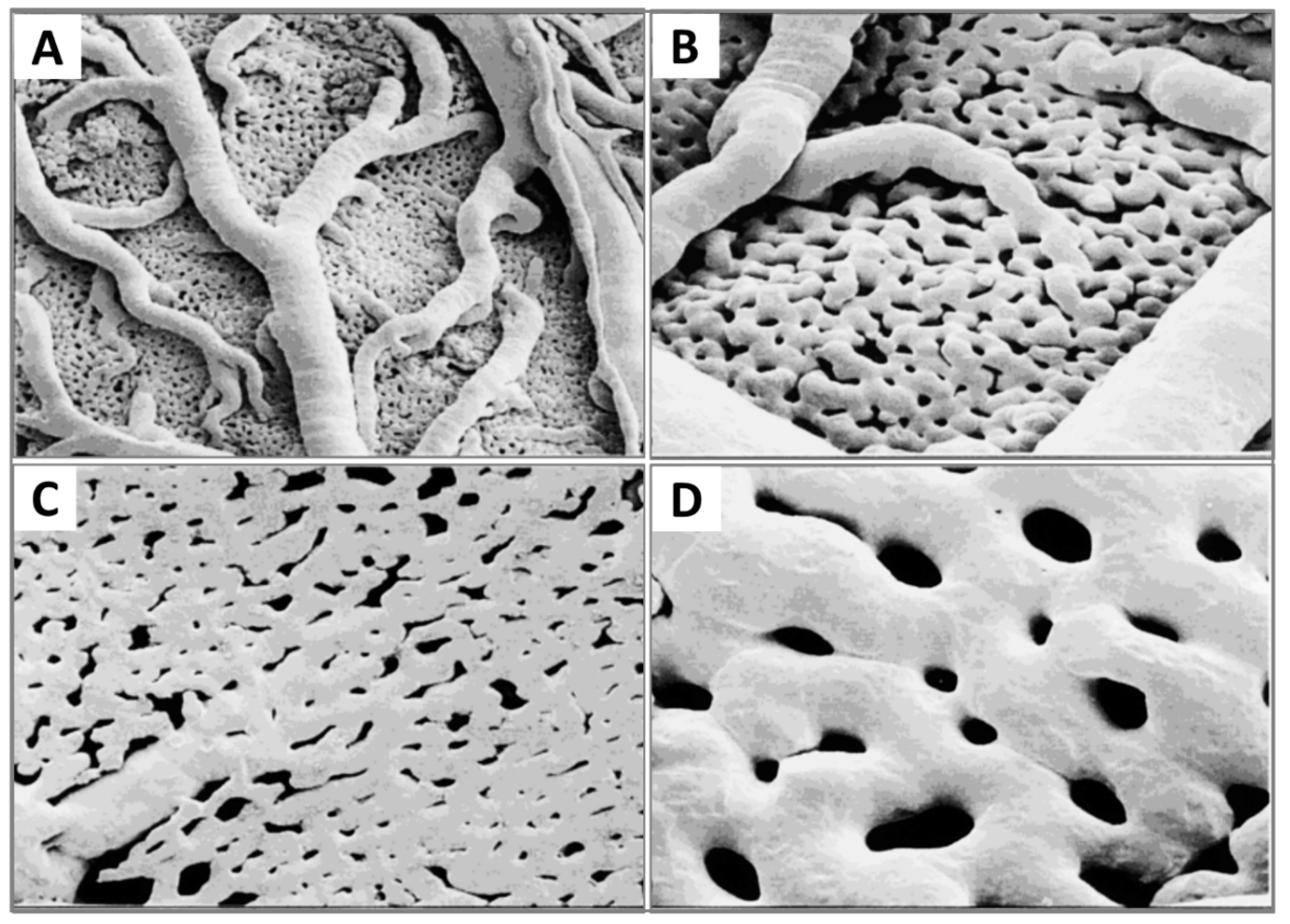
2.3. Microcirculation and Morphology in the CAM
2.4. Growth and Regulation Factors of Angiogenesis in CAM
3. CAM Assays in Preclinical Biomedical and Pharmacological Research
3.1. Drug Pharmacokinetics and Biodistribution
3.1.1. Drug Delivery System
3.1.2. Conventional Analysis Approach
3.1.3. Advanced Approaches
3.1.4. Vessel Permeability
3.2. Biocompatibility
3.3. The Efficacy of Drugs
3.3.1. Pro-Angiogenic Agents
3.3.2. Anti-Angiogenic Agents
3.3.3. Wound Healing
3.3.4. Tumour Growth and Metastasis
3.4. Screening of Drug Toxicity
3.4.1. Irritation
3.4.2. Toxicity
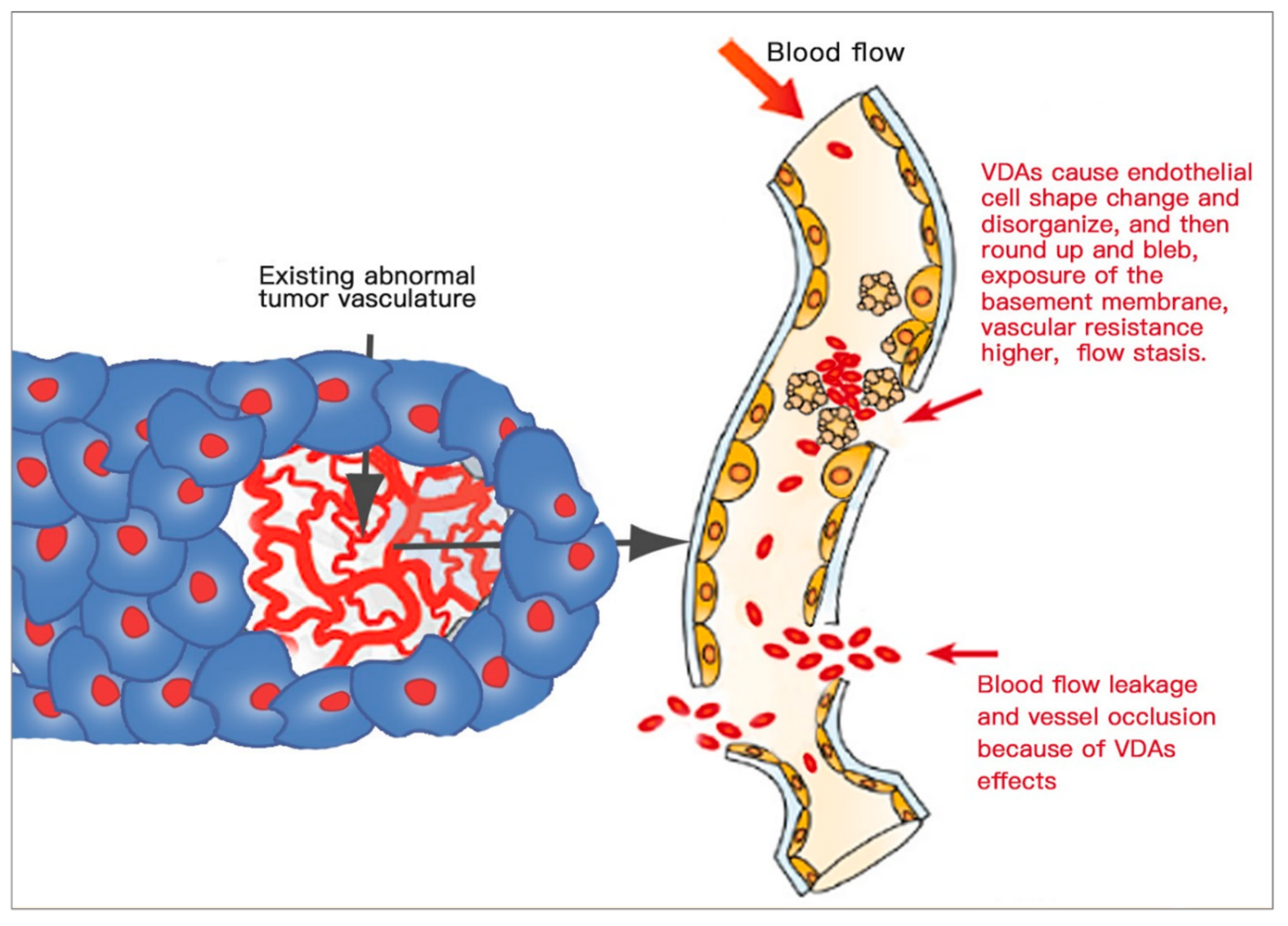
3.4.3. Anti-Vascular Selectivity
3.5. Time Course of CAM Applications for Different Assays
3.5.1. Drug Tests
3.5.2. Vascular Assays
3.5.3. Tumour Implantation
3.5.4. Cell Growth Factors and Antibody Inhibitors
3.6. Advantages and Drawbacks
4. Methods to Image and Evaluate the Changes in the Vasculature of the CAM
4.1. Light Microscopic Methods
4.2. X-ray Methods
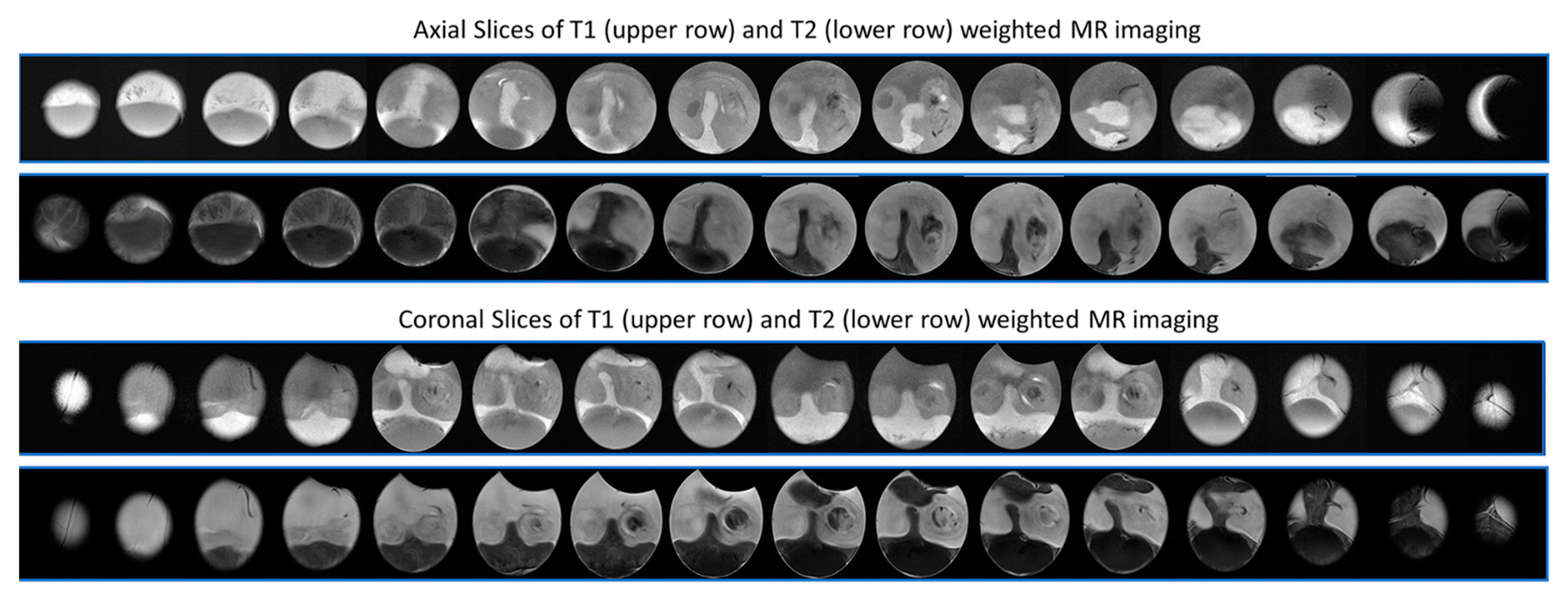
4.3. Magnetic Resonance Methods
4.4. Gamma-Ray Methods
4.5. Acoustic Methods
4.6. Optical Techniques and Methods
4.6.1. Optical Coherence Tomography (OCT) Principle
4.6.2. Laser Doppler Principle
4.6.3. Laser Speckle Principle
4.7. Artificial Intelligence (AI) Techniques and Algorithms
Author Contributions
Funding
Conflicts of Interest
References
- Staton, C.A.; Stribbling, S.M.; Tazzyman, S.; Hughes, R.; Brown, N.J.; Lewis, C.E. Current methods for assaying angiogenesis in vitro and in vivo. Int. J. Exp. Pathol. 2004, 85, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The chick embryo chorioallantoic membrane in the study of tumor angiogenesis. Rom. J. Morphol. Embryol. 2008, 49, 131–135. [Google Scholar]
- Chase, D.M.; Chaplin, D.J.; Monk, B.J. The development and use of vascular targeted therapy in ovarian cancer. Gynecol. Oncol. 2017, 145, 393–406. [Google Scholar] [CrossRef] [Green Version]
- Li, J. Research status mode Chicken chorioallantoic membrane applied for angiogenesis. Med. Front. 2015, 5, 4. [Google Scholar]
- Wang, S.; Liu, Y.; Feng, Y.; Zhang, J.; Swinnen, J.; Li, Y.; Ni, Y. A Review on Curability of Cancers: More Efforts for Novel Therapeutic Options Are Needed. Cancers (Basel) 2019, 11, 1782. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Nico, B.; Perra, M.T.; Longo, V.; Maxia, C.; Annese, T.; Piras, F.; Murtas, D.; Sirigu, P. Erythropoietin is involved in angiogenesis in human primary melanoma. Int. J. Exp. Pathol. 2010, 91, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Richards, L.M.; Kazmi, S.M.; Davis, J.L.; Olin, K.E.; Dunn, A.K. Low-cost laser speckle contrast imaging of blood flow using a webcam. Biomed. Opt. Express 2013, 4, 2269–2283. [Google Scholar] [CrossRef] [Green Version]
- Makanya, A.N.; Dimova, I.; Koller, T.; Styp-Rekowska, B.; Djonov, V. Dynamics of the Developing Chick Chorioallantoic Membrane Assessed by Stereology, Allometry, Immunohistochemistry and Molecular Analysis. PLoS ONE 2016, 11, e0152821. [Google Scholar] [CrossRef] [Green Version]
- Romanoff, A. The Avian Embryo: Structural and Functional Development; Macmillan: New York, NY, USA, 1960. [Google Scholar]
- Bellairs, R.; Osmond, M. Extra-Embryonic Membranes. In Atlas of Chick Development; Elsevier: Amsterdam, The Netherlands, 2014; pp. 127–129. [Google Scholar] [CrossRef]
- Ribatti, D.; Nico, B.; Vacca, A.; Roncali, L.; Burri, P.H.; Djonov, V. Chorioallantoic membrane capillary bed: A useful target for studying angiogenesis and anti-angiogenesis in vivo. Anat. Rec. 2001, 264, 317–324. [Google Scholar] [CrossRef]
- Kind, C. The development of the circulating blood volume of the chick embryo. Anat. Embryol. (Berl) 1975, 147, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Maina, J.N. Structure and Function of the Shell and the Chorioallantoic Membrane of the Avian Egg: Embryonic Respiration. In The Biology of the Avian Respiratory System; Springer: New York, NY, USA, 2017; pp. 219–247. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane (CAM). A multifaceted experimental model. Mech. Dev. 2016, 141, 70–77. [Google Scholar] [CrossRef]
- Nowak-Sliwinska, P.; Segura, T.; Iruela-Arispe, M.L. The chicken chorioallantoic membrane model in biology, medicine and bioengineering. Angiogenesis 2014, 17, 779–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, A.; Lindenbaum, E.S. The two- and three-dimensional structure of the microcirculation of the chick chorioallantoic membrane. Acta Anat. (Basel) 1988, 131, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Mueller, C.A.; Burggren, W.W.; Tazawa, H. The Physiology of the Avian Embryo. In Sturkie’s Avian Physiology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 739–766. [Google Scholar] [CrossRef]
- DeFouw, D.O.; Rizzo, V.J.; Steinfeld, R.; Feinberg, R.N. Mapping of the microcirculation in the chick chorioallantoic membrane during normal angiogenesis. Microvasc. Res. 1989, 38, 136–147. [Google Scholar] [CrossRef]
- Tazawa, H.; Whittow, G. Incubation Physiology: Sturkie’s Avian Physiology; Academic Press: New York NY, USA, 2000; pp. 617–637. [Google Scholar]
- Rahn, H.; Paganelli, C.V.; Ar, A. The avian egg: Air-cell gas tension, metabolism and incubation time. Respir. Physiol. 1974, 22, 297–309. [Google Scholar] [CrossRef]
- Kurz, H.; Ambrosy, S.; Wilting, J.; Marme, D.; Christ, B. Proliferation pattern of capillary endothelial cells in chorioallantoic membrane development indicates local growth control, which is counteracted by vascular endothelial growth factor application. Dev. Dyn. 1995, 203, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Piiper, J.; Tazawa, H.; Ar, A.; Rahn, H. Analysis of chorioallantoic gas exchange in the chick embryo. Respir. Physiol. 1980, 39, 273–284. [Google Scholar] [CrossRef]
- Rahn, H.; Ar, A. Gas exchange of the avian egg time structure and function. Am. Zool. 1980, 20, 477–487. [Google Scholar] [CrossRef]
- Vargas, A.; Zeisser-Labouebe, M.; Lange, N.; Gurny, R.; Delie, F. The chick embryo and its chorioallantoic membrane (CAM) for the in vivo evaluation of drug delivery systems. Adv. Drug Deliv. Rev. 2007, 59, 1162–1176. [Google Scholar] [CrossRef]
- Eichmann, A.; Yuan, L.; Moyon, D.; Lenoble, F.; Pardanaud, L.; Breant, C. Vascular development: From precursor cells to branched arterial and venous networks. Int. J. Dev. Biol. 2005, 49, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Ausprunk, D.H.; Knighton, D.R.; Folkman, J. Differentiation of vascular endothelium in the chick chorioallantois: A structural and autoradiographic study. Dev. Biol. 1974, 38, 237–248. [Google Scholar] [CrossRef]
- Kurz, H. Physiology of angiogenesis. J. Neurooncol. 2000, 50, 17–35. [Google Scholar] [CrossRef] [PubMed]
- Melkonian, G.; Munoz, N.; Chung, J.; Tong, C.; Marr, R.; Talbot, P. Capillary plexus development in the day five to day six chick chorioallantoic membrane is inhibited by cytochalasin D and suramin. J. Exp. Zool. 2002, 292, 241–254. [Google Scholar] [CrossRef] [PubMed]
- Djonov, V.; Schmid, M.; Tschanz, S.A.; Burri, P.H. Intussusceptive angiogenesis: Its role in embryonic vascular network formation. Circ. Res. 2000, 86, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Palmer, M.E. Development of the chick chorioallantoic capillary plexus under normoxic and normobaric hypoxic and hyperoxic conditions: A morphometric study. J. Exp. Zool. 1992, 262, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Maibier, M.; Reglin, B.; Nitzsche, B.; Xiang, W.; Rong, W.W.; Hoffmann, B.; Djonov, V.; Secomb, T.W.; Pries, A.R. Structure and hemodynamics of vascular networks in the chorioallantoic membrane of the chicken. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H913–H926. [Google Scholar] [CrossRef] [Green Version]
- Wagner-Amos, K.; Seymour, R.S. Effect of local shell conductance on the vascularisation of the chicken chorioallantoic membrane. Respir. Physiol. Neurobiol. 2003, 134, 155–167. [Google Scholar] [CrossRef]
- Kirchner, L.M.; Schmidt, S.P.; Gruber, B.S. Quantitation of angiogenesis in the chick chorioallantoic membrane model using fractal analysis. Microvasc. Res. 1996, 51, 2–14. [Google Scholar] [CrossRef]
- Shumko, J.Z.; Defouw, D.O.; Feinberg, R.N. Vascular histodifferentiation in the chick chorioallantoic membrane: A morphometric study. Anat. Rec. 1988, 220, 179–189. [Google Scholar] [CrossRef]
- Smith, A.F.; Nitzsche, B.; Maibier, M.; Pries, A.R.; Secomb, T.W. Microvascular hemodynamics in the chick chorioallantoic membrane. Microcirculation 2016, 23, 512–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinaccio, C.; Nico, B.; Ribatti, D. Differential expression of angiogenic and anti-angiogenic molecules in the chick embryo chorioallantoic membrane and selected organs during embryonic development. Int. J. Dev. Biol. 2013, 57, 907–916. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Bai, L.; Zhou, J.; Chen, H.; Zhang, L. Sequential delivery of VEGF, FGF-2 and PDGF from the polymeric system enhance HUVECs angiogenesis in vitro and CAM angiogenesis. Cell Immunol. 2018, 323, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Endogenous Basic Fibroblast Growth Factor Is Implicated in the Vascularization of chick embryo chorioallantoic membrane. Dev. Biol. 1995, 170, 39–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forough, R.; Weylie, B.; Patel, C.; Ambrus, S.; Singh, U.S.; Zhu, J. Role of AKT/PKB signaling in fibroblast growth factor-1 (FGF-1)-induced angiogenesis in the chicken chorioallantoic membrane (CAM). J. Cell. Biochem. 2005, 94, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Baum, O.; Suter, F.; Gerber, B.; Tschanz, S.A.; Buergy, R.; Blank, F.; Hlushchuk, R.; Djonov, V. VEGF-A promotes intussusceptive angiogenesis in the developing chicken chorioallantoic membrane. Microcirculation 2010, 17, 447–457. [Google Scholar] [CrossRef]
- Asahara, T.; Chen, D.; Takahashi, T.; Fujikawa, K.; Kearney, M.; Magner, M.; Yancopoulos, G.D.; Isner, J.M. Tie2 receptor ligands, angiopoietin-1 and angiopoietin-2, modulate VEGF-induced postnatal neovascularization. Circ. Res. 1998, 83, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Drenkhahn, M.; Gescher, D.M.; Wolber, E.M.; Meyhoefer-Malik, A.; Malik, E. Expression of angiopoietin 1 and 2 in ectopic endometrium on the chicken chorioallantoic membrane. Fertil. Steril. 2004, 81 (Suppl. 1), 869–875. [Google Scholar] [CrossRef]
- Bussolino, E.D.R.M.; Ziche, M.; Bocchietto, E.; Olivero, M.; Naldini, L.; Gaudino, G.T.L.; Coffer, A.; Comoglio, P. Hepatocyte growth factor is a potent angiogenic factor which stimulates endothelial cell motility and growth. J. Cell Biol. 1992, 119, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Eva, V. Structural Analysis of the Angiogenesis in the Chicken Chorioallantoic Membrane. Doctoral Thesis, KU LEUVEN, Leuven, Belgium, 2011. [Google Scholar]
- Reuten, R.; Patel, T.R.; McDougall, M.; Rama, N.; Nikodemus, D.; Gibert, B.; Delcros, J.G.; Prein, C.; Meier, M.; Metzger, S.; et al. Structural decoding of netrin-4 reveals a regulatory function towards mature basement membranes. Nat. Commun. 2016, 7, 13515. [Google Scholar] [CrossRef] [Green Version]
- Rous, P.; Murppy, J. Tumor Implantations in the Developing Embryo. J. Am. Med. Assoc. 1911, 56, 741–742. [Google Scholar] [CrossRef]
- Nikiforidis, G.; Papazafiropoulos, D.; Siablis, D.; Karnabatidis, D.; Hatjikondi, O.; Dimopoulos, J. Quantitative assessment of angiogenesis in the chick embryo and its chorioallantoic membrane by computerised analysis of angiographic images. Eur. J. Radiol. 1999, 29, 168–179. [Google Scholar] [CrossRef]
- Lange, N.; Ballini, J.P.; Wagnieres, G.; van den Bergh, H. A new drug-screening procedure for photosensitizing agents used in photodynamic therapy for CNV. Investig. Ophthalmol. Vis. Sci. 2001, 42, 38–46. [Google Scholar]
- Li, J.; Sun, Z.; Zhang, J.; Shao, H.; Cona, M.M.; Wang, H.; Marysael, T.; Chen, F.; Prinsen, K.; Zhou, L.; et al. A dual-targeting anticancer approach: Soil and seed principle. Radiology 2011, 260, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Dikici, S.; Bullock, A.J.; Yar, M.; Claeyssens, F.; MacNeil, S. 2-deoxy-d-ribose (2dDR) upregulates vascular endothelial growth factor (VEGF) and stimulates angiogenesis. Microvasc. Res. 2020, 131, 104035. [Google Scholar] [CrossRef] [PubMed]
- Mangir, N.; Dikici, S.; Claeyssens, F.; MacNeil, S. Using ex Ovo Chick Chorioallantoic Membrane (CAM) Assay To Evaluate the Biocompatibility and Angiogenic Response to Biomaterials. ACS Biomater. Sci. Eng. 2019, 5, 3190–3200. [Google Scholar] [CrossRef]
- Nowak-Sliwinska, P.; Ballini, J.P.; Wagnieres, G.; van den Bergh, H. Processing of fluorescence angiograms for the quantification of vascular effects induced by anti-angiogenic agents in the CAM model. Microvasc. Res. 2010, 79, 21–28. [Google Scholar] [CrossRef]
- Vu, B.T.; Shahin, S.A.; Croissant, J.; Fatieiev, Y.; Matsumoto, K.; Le-Hoang Doan, T.; Yik, T.; Simargi, S.; Conteras, A.; Ratliff, L.; et al. Chick chorioallantoic membrane assay as an in vivo model to study the effect of nanoparticle-based anticancer drugs in ovarian cancer. Sci. Rep. 2018, 8, 8524. [Google Scholar] [CrossRef]
- Buchele, B.; Zugmaier, W.; Genze, F.; Simmet, T. High-performance liquid chromatographic determination of acetyl-11-keto-alpha-boswellic acid, a novel pentacyclic triterpenoid, in plasma using a fluorinated stationary phase and photodiode array detection: Application in pharmacokinetic studies. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2005, 829, 144–148. [Google Scholar] [CrossRef]
- Fotinos, N.; Campo, M.A.; Popowycz, F.; Gurny, R.; Lange, N. 5-Aminolevulinic acid derivatives in photomedicine: Characteristics, application and perspectives. Photochem. Photobiol. 2006, 82, 994–1015. [Google Scholar] [CrossRef] [PubMed]
- Mangir, N.; Raza, A.; Haycock, J.W.; Chapple, C.; Macneil, S. An Improved In Vivo Methodology to Visualise Tumour Induced Changes in Vasculature Using the Chick Chorionic Allantoic Membrane Assay. In Vivo 2018, 32, 461–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dikici, S.; Mangir, N.; Claeyssens, F.; Yar, M.; MacNeil, S. Exploration of 2-deoxy-D-ribose and 17beta-Estradiol as alternatives to exogenous VEGF to promote angiogenesis in tissue-engineered constructs. Regen. Med. 2019, 14, 179–197. [Google Scholar] [CrossRef] [PubMed]
- Meadows, D. Recent developments with biosensing technology and applications in the pharmaceutical industry. Adv. Drug Deliv. Rev. 1996, 21, 179–189. [Google Scholar] [CrossRef]
- Valdes, T.I.; Klueh, U.; Kreutzer, D.; Moussy, F. Ex ova chick chorioallantoic membrane as a novel in vivo model for testing biosensors. J. Biomed. Mater. Res. A 2003, 67, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Klueh, U.; Dorsky, D.I.; Kreutzer, D.L. Enhancement of implantable glucose sensor function in vivo using gene transfer-induced neovascularization. Biomaterials 2005, 26, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Murphy. The effect of adult chicken organ grafts on CAM. J. Exp. Med. 1916, 24, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Chiba, A.; Yui, C.; Hirano, S. Liver reconstruction on the chorioallantoic membrane of the chick embryo. Arch. Histol. Cytol. 2010, 73, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, M.; DeRuiter, M.C.; Carretero, A.; Ruberte, J. Microvascular assembly and cell invasion in chick mesonephros grafted onto chorioallantoic membrane. J. Anat. 2003, 202, 213–225. [Google Scholar] [CrossRef]
- Bertossi, M.; Virgintino, D.; Coltey, P.; Errede, M.; Mancini, L.; Roncali, L. Angiogenesis and endothelium phenotype expression in embryonic adrenal gland and cerebellum grafted onto chorioallantoic membrane. Angiogenesis 1999, 3, 305–315. [Google Scholar] [CrossRef]
- Moreno-Jimenez, I.; Hulsart-Billstrom, G.; Lanham, S.A.; Janeczek, A.A.; Kontouli, N.; Kanczler, J.M.; Evans, N.D.; Oreffo, R.O. The chorioallantoic membrane (CAM) assay for the study of human bone regeneration: A refinement animal model for tissue engineering. Sci. Rep. 2016, 6, 32168. [Google Scholar] [CrossRef]
- Noiman, T.; Buzhor, E.; Metsuyanim, S.; Harari-Steinberg, O.; Morgenshtern, C.; Dekel, B.; Goldstein, R.S. A rapid in vivo assay system for analyzing the organogenetic capacity of human kidney cells. Organogenesis 2011, 7, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Maas, J.W.; Le Noble, F.A.; Dunselman, G.A.; de Goeij, A.F.; Boudier, H.A.S.; Evers, J.L. The Chick Embryo Chorioallantoic Membrane as a Model to Investigate the Angiogenic Properties of Human Endometrium. Gynecol. Obs. Investig. 1999, 48, 5. [Google Scholar] [CrossRef]
- Kunzi-Rapp, K.; Rück, A.; Kaufmann, R. Characterization of the chick chorioallantoic membrane model as a short-term in vivo system for human skin. Arch. Derm. Res. 1999, 6, 290–295. [Google Scholar] [CrossRef]
- Isachenko, V.; Isachenko, E.; Mallmann, P.; Rahimi, G. Increasing Follicular and Stromal Cell Proliferation in Cryopreserved Human Ovarian Tissue after Long-Term Precooling Prior to Freezing: In Vitro versus Chorioallantoic Membrane (CAM) Xenotransplantation. Cell Transplant. 2013, 22, 2053–2061. [Google Scholar] [CrossRef]
- Ausprunk, D.H.; Knighton, D.R.; Folkman, J. Vascularization of normal and neoplastic tissues grafted to the chick chorioallantois. Role of host and preexisting graft blood vessels. Am. J. Pathol 1975, 79, 597–618. [Google Scholar] [PubMed]
- Wong, C.; Inman, E.; Spaethe, R.; Helgerson, S. Fibrin-based biomaterials to deliver human growth factors. Thromb. Haemost. 2003, 89, 573–582. [Google Scholar] [CrossRef]
- Steffens, G.C.; Yao, C.; Prevel, P.; Markowicz, M.; Schenck, P.; Noah, E.M.; Pallua, N. Modulation of angiogenic potential of collagen matrices by covalent incorporation of heparin and loading with vascular endothelial growth factor. Tissue Eng. 2004, 10, 1502–1509. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane in the Study of Angiogenesis and Metastasis; Springer: Dordrecht, The Netherlands, 2010. [Google Scholar] [CrossRef]
- Beckers, M.; Gladis-Villanueva, M.; Hamann, W.; Schmutzler, W.; Zwadlo-Klarwasser, G. The use of the chorio-allantoic membrane of the chick embryo as test for anti-inflammatory activity. Inflamm Res. 1997, 46, 29–30. [Google Scholar] [CrossRef] [PubMed]
- Wilting, J.; Christ, B.; Bokeloh, M. A modified chorioallantoic membrane (CAM) assay for qualitative and quantitative study of growth factors. Studies on the effects of carriers, PBS, angiogenin, and bFGF. Anat. Embryol. (Berl) 1991, 183, 259–271. [Google Scholar] [CrossRef]
- Langer, R.; Folkman, J. Polymers for the sustained release of proteins and other macromolecules. Nature 1976, 263, 797–800. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.Y.; Moses, H.L. Transforming growth factor beta 1-induced changes in cell migration, proliferation, and angiogenesis in the chicken chorioallantoic membrane. J. Cell Biol. 1990, 111, 731–741. [Google Scholar] [CrossRef]
- Riboldi, E.; Musso, T.; Moroni, E.; Urbinati, C.; Bernasconi, S.; Rusnati, M.; Adorini, L.; Presta, M.; Sozzani, S. Cutting edge: Proangiogenic properties of alternatively activated dendritic cells. J. Immunol. 2005, 175, 2788–2792. [Google Scholar] [CrossRef]
- Ribatti, D.; Nico, B.; Vacca, A.; Presta, M. The gelatin sponge-chorioallantoic membrane assay. Nat. Protoc. 2006, 1, 85–91. [Google Scholar] [CrossRef]
- Dreesmann, L.; Ahlers, M.; Schlosshauer, B. The pro-angiogenic characteristics of a cross-linked gelatin matrix. Biomaterials 2007, 28, 5536–5543. [Google Scholar] [CrossRef]
- Yao, C.; Markowicz, M.; Pallua, N.; Noah, E.M.; Steffens, G. The effect of cross-linking of collagen matrices on their angiogenic capability. Biomaterials 2008, 29, 66–74. [Google Scholar] [CrossRef]
- Kanczler, J.M.; Barry, J.; Ginty, P.; Howdle, S.M.; Shakesheff, K.M.; Oreffo, R.O. Supercritical carbon dioxide generated vascular endothelial growth factor encapsulated poly(DL-lactic acid) scaffolds induce angiogenesis in vitro. Biochem. Biophys. Res. Commun. 2007, 352, 135–141. [Google Scholar] [CrossRef]
- Wacker, B.K.; Scott, E.A.; Kaneda, M.M.; Alford, S.K.; Elbert, D.L. Delivery of sphingosine 1-phosphate from poly(ethylene glycol) hydrogels. Biomacromolecules 2006, 7, 1335–1343. [Google Scholar] [CrossRef] [Green Version]
- Auerbach, R.; Lewis, R.; Shinners, B.; Kubai, L.; Akhtar, N. Angiogenesis assays: A critical overview. Clin. Chem. 2003, 49, 32–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane. In Vivo Models to Study Angiogenesis; Elsevier: Amsterdam, The Netherlands, 2018; pp. 1–23. [Google Scholar] [CrossRef]
- Gualandris, A.; Rusnati, M.; Bastaki, M.; Ziche, M.; Presta, M. Basic Fribroblast Growth Factor Overexpression in Endothelial Cells: An Autocrine Mechanism for Angiogenesis and Angioproliferative Diseases; Springer: Boston, MA, USA, 1998. [Google Scholar] [CrossRef]
- Ehrbar, M.; Djonov, V.G.; Schnell, C.; Tschanz, S.A.; Martiny-Baron, G.; Schenk, U.; Wood, J.; Burri, P.H.; Hubbell, J.A.; Zisch, A.H. Cell-demanded liberation of VEGF121 from fibrin implants induces local and controlled blood vessel growth. Circ. Res. 2004, 94, 1124–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, W.J.; Kayton, M.L.; Patton, A.; O’Connor, S.; He, M.; Vu, H.; Baibakov, G.; Lorang, D.; Knezevic, V.; Kohn, E.; et al. A novel technique for quantifying changes in vascular density, endothelial cell proliferation and protein expression in response to modulators of angiogenesis using the chick chorioallantoic membrane (CAM) assay. J. Transl. Med. 2004, 2, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribatti, D. The Chick Embryo Chorioallantoic Membrane as an In Vivo Assay to Study Antiangiogenesis. Pharmaceuticals 2010, 3, 482–513. [Google Scholar] [CrossRef]
- As, M.N.; Deshpande, R.; Kale, V.P.; Bhonde, R.R.; Datar, S.P. Establishment of an in ovo chick embryo yolk sac membrane (YSM) assay for pilot screening of potential angiogenic and anti-angiogenic agents. Cell Biol. Int. 2018, 42, 1474–1483. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, S.; Zhang, Z.; Luo, Q. Laser speckle contrast imaging: Monitoring blood flow dynamics and vascular structure of photodynamic therapy. In Proceedings of the Optics in Health Care and Biomedical Optics: Diagnostics and Treatment II, Beijing, China, 12 November 2004. [Google Scholar]
- Ismail, M.S.; Torsten, U.; Dressler, C.; Diederichs, J.E.; Huske, S.; Weitzel, H.; Berlien, H.P. Photodynamic Therapy of Malignant Ovarian Tumours Cultivated on CAM. Lasers Med. Sci. 1999, 14, 91–96. [Google Scholar] [CrossRef]
- FDA, W.H.C.F.G. Guidance for industry: Chronic cutaneous ulcer and burn wounds-developing products for treatment. Wound Repair. Regen. 2001, 9, 258–268. [Google Scholar] [CrossRef]
- Ribatti, D.; Vacca, A.; Ranieri, G.; Sorino, S.; Roncali, L. The Chick Embryo Chorioallantoic Membrane as an in vivo Wound Healing Model. Pathol. Res. Pract. 1996, 192, 1068–1076. [Google Scholar] [CrossRef]
- Ribatti, D.; Nico, B.; Vacca, A.; Roncali, L.; Presta, M. Endogenous and exogenous fibroblast growth factor-2 modulate wound healing in the chick embryo chorioallantoic membrane. Angiogenesis 1999, 3, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.J.; Xue, M.; Thompson, P.; Davey, R.A.; Whitmont, K.; Smith, S.; Buisson-Legendre, N.; Sztynda, T.; Furphy, L.J.; Cooper, A.; et al. Activated protein C prevents inflammation yet stimulates angiogenesis to promote cutaneous wound healing. Wound Repair. Regen. 2005, 13, 284–294. [Google Scholar] [CrossRef]
- Guerra, A.; Belinha, J.; Mangir, N.; MacNeil, S.; Natal Jorge, R. Sprouting Angiogenesis: A Numerical Approach with Experimental Validation. Ann. Biomed. Eng. 2021, 49, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.B. Transplantability of Tissues to the Embryo of Foreign Species : Its Bearing on Questions of Tissue Specificity and Tumor Immunity. J. Exp. Med. 1913, 17, 482–493. [Google Scholar] [CrossRef] [Green Version]
- Deryugina, E.I.; Quigley, J.P. Chick embryo chorioallantoic membrane model systems to study and visualize human tumor cell metastasis. Histochem. Cell Biol. 2008, 130, 1119–1130. [Google Scholar] [CrossRef] [Green Version]
- Karnofsky, D.A.; Ridgway, L.P.; Patterson, P.A. Tumor transplantation to the chick embryo. Ann. N. Y. Acad. Sci. 1952, 55, 313–329. [Google Scholar] [CrossRef]
- Kunzi-Rapp, K.; Westphal-Frosch, C.; Schneckenburger, H. Test system for human tumor cell sensitivity to drugs on chicken chorioallantoic membranes. Vitr. Cell. Dev. Biol. 1992, 28, 565–566. [Google Scholar] [CrossRef] [PubMed]
- Knighton, D.; Ausprunk, D.; Tapper, D.; Folkman, J. Avascular and vascular phases of tumour growth in the chick embryo. Br. J. Cancer 1977, 35, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Bobek, V.; Plachy, J.; Pinterova, D.; Kolostova, K.; Boubelik, M.; Jiang, P.; Yang, M.; Hoffman, R.M. Development of a green fluorescent protein metastatic-cancer chick-embryo drug-screen model. Clin. Exp. Metastasis 2004, 21, 347–352. [Google Scholar] [CrossRef]
- Pinto, M.T.; Ribeiro, A.S.; Conde, I.; Carvalho, R.; Paredes, J. The Chick Chorioallantoic Membrane Model: A New In Vivo Tool to Evaluate Breast Cancer Stem Cell Activity. Int. J. Mol. Sci. 2020, 22, 334. [Google Scholar] [CrossRef]
- Armstrong, P.B.; Quigley, J.P.; Sidebottom, E. Transepithelial invasion and intramesenchymal infiltration of the chick embryo chorioallantois by tumor cell lines. Cancer Res. 1982, 42, 1826–1837. [Google Scholar]
- Scher, C.D.; Haudenschild, C.; Klagsbrun, M. The chick chorioallantoic membrane as a model system for the study of tissue invasion by viral transformed cells. Cell 1976, 8, 373–382. [Google Scholar] [CrossRef]
- Wilson, S.M.; Chambers, A.F. Experimental metastasis assays in the chick embryo. Curr. Protoc. Cell Biol. 2004, 21, 19.6.1–19.6.24. [Google Scholar] [CrossRef]
- Quigley, J.P.; Armstrong, P.B. Tumor cell intravasation alu-cidated: The chick embryo opens the window. Cell 1998, 94, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.J. The human tumor grown in the egg. Ann. N. Y. Acad. Sci. 1958, 76, 764–769. [Google Scholar] [CrossRef]
- Dunker, N.; Jendrossek, V. Implementation of the Chick Chorioallantoic Membrane (CAM) Model in Radiation Biology and Experimental Radiation Oncology Research. Cancers (Basel) 2019, 11, 1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syrovets, T.; Gschwend, J.E.; Buchele, B.; Laumonnier, Y.; Zugmaier, W.; Genze, F.; Simmet, T. Inhibition of IkappaB kinase activity by acetyl-boswellic acids promotes apoptosis in androgen-independent PC-3 prostate cancer cells in vitro and in vivo. J. Biol. Chem. 2005, 280, 6170–6180. [Google Scholar] [CrossRef] [Green Version]
- Pastorino, F.; Brignole, C.; Di Paolo, D.; Nico, B.; Pezzolo, A.; Marimpietri, D.; Pagnan, G.; Piccardi, F.; Cilli, M.; Longhi, R.; et al. Targeting liposomal chemotherapy via both tumor cell-specific and tumor vasculature-specific ligands potentiates therapeutic efficacy. Cancer Res. 2006, 66, 10073–10082. [Google Scholar] [CrossRef] [Green Version]
- Chin, W.W.; Lau, W.K.; Bhuvaneswari, R.; Heng, P.W.; Olivo, M. Chlorin e6-polyvinylpyrrolidone as a fluorescent marker for fluorescence diagnosis of human bladder cancer implanted on the chick chorioallantoic membrane model. Cancer Lett. 2007, 245, 127–133. [Google Scholar] [CrossRef]
- Gottfried, V.; Davidi, R.; Averguj, C.; Kimel, S. In vivo damage to chorioallantoic membrane blood vessels by porphycene-induced photodynamic therapy. J. Photochem. Photobiol. B 1995, 30, 115–121. [Google Scholar] [CrossRef]
- Valiulyte, I.; Curkunaviciute, R.; Ribokaite, L.; Kazlauskas, A.; Vaitkeviciute, M.; Skauminas, K.; Valanciute, A. The Anti-Tumorigenic Activity of Sema3C in the Chick Embryo Chorioallantoic Membrane Model. Int. J. Mol. Sci. 2019, 20, 5672. [Google Scholar] [CrossRef] [Green Version]
- Lokman, N.A.; Elder, A.S.; Ricciardelli, C.; Oehler, M.K. Chick chorioallantoic membrane (CAM) assay as an in vivo model to study the effect of newly identified molecules on ovarian cancer invasion and metastasis. Int. J. Mol. Sci. 2012, 13, 9959–9970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Piterski, C.; Nitka, S. Evaluation of the hen’s egg test-chorioallantonic membrane (CAM) method in prediction of the eye irritation potential formulated personal wash products. Cutan. Ocul. Toxicol. 2007, 26, 25–36. [Google Scholar] [CrossRef]
- Xie, X.; Tan, X. Application of Hen’s egg-chorioallantoic membrane assay in safety evaluation of cosmetics. China Trop. Med. 2009, 9, 2. [Google Scholar]
- Abdelkader, H.; Ismail, S.; Hussein, A.; Wu, Z.; Al-Kassas, R.; Alany, R.G. Conjunctival and corneal tolerability assessment of ocular naltrexone niosomes and their ingredients on the hen’s egg chorioallantoic membrane and excised bovine cornea models. Int. J. Pharm. 2012, 432, 1–10. [Google Scholar] [CrossRef] [PubMed]
- de Avila, R.I.; de Sousa Vieira, M.; Gaeti, M.P.; Moreira, L.C.; de Brito Rodrigues, L.; de Oliveira, G.A.; Batista, A.C.; Vinhal, D.C.; Menegatti, R.; Valadares, M.C. Toxicity evaluation of the photoprotective compound LQFM048: Eye irritation, skin toxicity and genotoxic endpoints. Toxicology 2017, 376, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Hamamichi, S.; Nishigori, H. Establishment of a chick embryo shell-less culture system and its use to observe change in behavior caused by nicotine and substances from cigarette smoke. Toxicol. Lett. 2001, 119, 95–102. [Google Scholar] [CrossRef]
- Datar, S.; Bhonde, R.R. Shell-less chick embryo culture as an alternative in vitro model to investigate glucose-induced malformations in mammalian embryos. Rev. Diabet Stud. 2005, 2, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Dou, W.; Yan, C.; Liu, T.; Wei, D. Study on Toxicity of Angelica Sinensis Injection and Astragalus Membranaceus Injection in Chorioallantoic Membrane Model. Chin. J. Exp. Tradit. Med. Formulae 2009, 15. [Google Scholar]
- Siemann, D.W.; Bibby, M.C.; Dark, G.G.; Dicker, A.P.; Eskens, F.A.; Horsman, M.R.; Marme, D.; Lorusso, P.M. Differentiation and definition of vascular-targeted therapies. Clin. Cancer Res. 2005, 11, 416–420. [Google Scholar]
- Hinnen, P.; Eskens, F.A. Vascular disrupting agents in clinical development. Br. J. Cancer 2007, 96, 1159–1165. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Wakelee, H. Vascular disrupting agents. J. Thorac. Oncol. 2010, 5, S482–S483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siemann, D.W. The unique characteristics of tumor vasculature and preclinical evidence for its selective disruption by Tumor-Vascular Disrupting Agents. Cancer Treat. Rev. 2011, 37, 63–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhang, X.; Wang, G.; Peng, Y.; Zhang, X.; Wu, H.; Yu, B.; Sun, J. Preclinical Pharmacokinetics of C118P, a Novel Prodrug of Microtubules Inhibitor and Its Metabolite C118 in Mice, Rats, and Dogs. Molecules 2018, 23, 2883. [Google Scholar] [CrossRef] [Green Version]
- Muenzner, J.K.; Biersack, B.; Kalie, H.; Andronache, I.C.; Kaps, L.; Schuppan, D.; Sasse, F.; Schobert, R. Gold(I) biscarbene complexes derived from vascular-disrupting combretastatin A-4 address different targets and show antimetastatic potential. ChemMedChem 2014, 9, 1195–1204. [Google Scholar] [CrossRef]
- Jiang, Z.; Wu, M.; Miao, J.; Duan, H.; Zhang, S.; Chen, M.; Sun, L.; Wang, Y.; Zhang, X.; Zhu, X.; et al. Deoxypodophyllotoxin exerts both anti-angiogenic and vascular disrupting effects. Int. J. Biochem. Cell Biol. 2013, 45, 1710–1719. [Google Scholar] [CrossRef]
- Kim, K.W.; Lee, J.M.; Jeon, Y.S.; Lee, I.J.; Choi, Y.; Park, J.; Kiefer, B.; Kim, C.; Han, J.K.; Choi, B.I. Vascular disrupting effect of CKD-516: Preclinical study using DCE-MRI. Investig. New Drugs 2013, 31, 1097–1106. [Google Scholar] [CrossRef]
- Pasquier, E.; Sinnappan, S.; Munoz, M.A.; Kavallaris, M. ENMD-1198, a new analogue of 2-methoxyestradiol, displays both antiangiogenic and vascular-disrupting properties. Mol. Cancer 2010, 9, 1408–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Dai, M.; Lin, L.P.; Li, P.K.; Ding, J. Anti-angiogenic and vascular disrupting effects of C9, a new microtubule-depolymerizing agent. Br. J. Pharm. 2009, 156, 1228–1238. [Google Scholar] [CrossRef] [Green Version]
- Petitclerc, E.; Deschesnes, R.G.; Cote, M.F.; Marquis, C.; Janvier, R.; Lacroix, J.; Miot-Noirault, E.; Legault, J.; Mounetou, E.; Madelmont, J.C.; et al. Antiangiogenic and antitumoral activity of phenyl-3-(2-chloroethyl)ureas: A class of soft alkylating agents disrupting microtubules that are unaffected by cell adhesion-mediated drug resistance. Cancer Res. 2004, 64, 4654–4663. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.B.; Chen, C.P.; Yeh, T.K.; Song, J.S.; Chang, C.Y.; Chuu, J.J.; Tung, F.F.; Ho, P.Y.; Chen, T.W.; Lin, C.H.; et al. BPR0C261 is a novel orally active antitumor agent with antimitotic and anti-angiogenic activities. Cancer Sci. 2011, 102, 182–191. [Google Scholar] [CrossRef]
- Chen, C.P.; Hu, C.B.; Yeh, K.C.; Song, J.S.; Yeh, T.K.; Tung, F.F.; Hwang, L.L.; Tseng, H.Y.; Huang, Y.C.; Shy, H.S.; et al. Antiangiogenic activities and cisplatin-combined antitumor activities of BPR0L075. Anticancer Res. 2010, 30, 2813–2822. [Google Scholar] [PubMed]
- Tuma, M.C.; Malikzay, A.; Ouyang, X.; Surguladze, D.; Fleming, J.; Mitelman, S.; Camara, M.; Finnerty, B.; Doody, J.; Chekler, E.L.; et al. Antitumor Activity of IMC-038525, a Novel Oral Tubulin Polymerization Inhibitor. Transl. Oncol. 2010, 3, 318–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tozer, G.M. Measuring tumour vascular response to antivascular and antiangiogenic drugs. Br. J. Radiol. 2003, 76, S23–S35. [Google Scholar] [CrossRef]
- Sosa, J.A.; Elisei, R.; Jarzab, B.; Balkissoon, J.; Lu, S.P.; Bal, C.; Marur, S.; Gramza, A.; Yosef, R.B.; Gitlitz, B.; et al. Randomized safety and efficacy study of fosbretabulin with paclitaxel/carboplatin against anaplastic thyroid carcinoma. Thyroid 2014, 24, 232–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zweifel, M.; Jayson, G.C.; Reed, N.S.; Osborne, R.; Hassan, B.; Ledermann, J.; Shreeves, G.; Poupard, L.; Lu, S.P.; Balkissoon, J.; et al. Phase II trial of combretastatin A4 phosphate, carboplatin, and paclitaxel in patients with platinum-resistant ovarian cancer. Ann. Oncol. 2011, 22, 2036–2041. [Google Scholar] [CrossRef] [PubMed]
- Siemann, D.W.; Shi, W. Dual targeting of tumor vasculature: Combining Avastin and vascular disrupting agents (CA4P or OXi4503). Anticancer Res. 2008, 28, 2027–2031. [Google Scholar] [PubMed]
- Garon, E.B.; Neidhart, J.D.; Gabrail, N.Y.; de Oliveira, M.R.; Balkissoon, J.; Kabbinavar, F. A randomized Phase II trial of the tumor vascular disrupting agent CA4P (fosbretabulin tromethamine) with carboplatin, paclitaxel, and bevacizumab in advanced nonsquamous non-small-cell lung cancer. Onco Targets Ther. 2016, 9, 7275–7283. [Google Scholar] [CrossRef] [Green Version]
- Monk, B.; Herzog, T.; Alvarez, R.; Chan, J.; Chase, D.; Couchenour, R.; Cram, C.; Bookman, M. FOCUS study: Physician’s choice chemotherapy (PCC) plus bevacizumab and CA4P versus PCC plus bevacizumab and placebo in platinum-resistant ovarian cancer. In Proceedings of the International Gynecologic Cancer Societ, Lisbon, Portugal, 29–31 October 2016. [Google Scholar]
- Su, M.; Huang, J.; Liu, S.; Xiao, Y.; Qin, X.; Liu, J.; Pi, C.; Luo, T.; Li, J.; Chen, X.; et al. The anti-angiogenic effect and novel mechanisms of action of Combretastatin A-4. Sci. Rep. 2016, 6, 28139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahal, K.; Resch, M.; Ficner, R.; Schobert, R.; Biersack, B.; Mueller, T. Effects of the tumor-vasculature-disrupting agent verubulin and two heteroaryl analogues on cancer cells, endothelial cells, and blood vessels. ChemMedChem 2014, 9, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Rovithi, M.; Avan, A.; Funel, N.; Leon, L.G.; Gomez, V.E.; Wurdinger, T.; Griffioen, A.W.; Verheul, H.M.W.; Giovannetti, E. Development of bioluminescent chick chorioallantoic membrane (CAM) models for primary pancreatic cancer cells: A platform for drug testing. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Kue, C.S.; Tan, K.Y.; Lam, M.L.; Lee, H.B. Chick embryo chorioallantoic membrane (CAM): An alternative predictive model in acute toxicological studies for anti-cancer drugs. Exp. Anim. 2015, 64, 129–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, S.L.; Heng, P.W.; Chan, L.W. The chick chorioallantoic membrane imaging method as a platform to evaluate vasoactivity and assess irritancy of compounds. J. Pharm. Pharm. 2012, 64, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Larger, E.; Marre, M.; Corvol, P.; Gasc, J.M. Hyperglycemia-induced defects in angiogenesis in the chicken chorioallantoic membrane model. Diabetes 2004, 53, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Ferician, O.; Cimpean, A.M.; Avram, S.; Raica, M. Endostatin Effects on Tumor Cells and Vascular Network of Human Renal Cell Carcinoma Implanted on Chick Embryo Chorioallantoic Membrane. Anticancer Res. 2015, 35, 6521–6528. [Google Scholar]
- Schlatter, P.; Konig, M.F.; Karlsson, L.M.; Burri, P.H. Quantitative study of intussusceptive capillary growth in the chorioallantoic membrane (CAM) of the chicken embryo. Microvasc. Res. 1997, 54, 65–73. [Google Scholar] [CrossRef]
- Ribatti, D. The chick embryo chorioallantoic membrane as a model for tumor biology. Exp. Cell Res. 2014, 328, 314–324. [Google Scholar] [CrossRef]
- Kunz, P.; Schenker, A.; Sahr, H.; Lehner, B.; Fellenberg, J. Optimization of the chicken chorioallantoic membrane assay as reliable in vivo model for the analysis of osteosarcoma. PLoS ONE 2019, 14, e0215312. [Google Scholar] [CrossRef]
- Hu, J.; Ishihara, M.; Chin, A.I.; Wu, L. Establishment of xenografts of urological cancers on chicken chorioallantoic membrane (CAM) to study metastasis. Precis. Clin. Med. 2019, 2, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Eckrich, J.; Kugler, P.; Buhr, C.R.; Ernst, B.P.; Mendler, S.; Baumgart, J.; Brieger, J.; Wiesmann, N. Monitoring of tumor growth and vascularization with repetitive ultrasonography in the chicken chorioallantoic-membrane-assay. Sci. Rep. 2020, 10, 18585. [Google Scholar] [CrossRef]
- Merckx, G.; Tay, H.; Lo Monaco, M.; van Zandvoort, M.; De Spiegelaere, W.; Lambrichts, I.; Bronckaers, A. Chorioallantoic Membrane Assay as Model for Angiogenesis in Tissue Engineering: Focus on Stem Cells. Tissue Eng. Part. B Rev. 2020. [Google Scholar] [CrossRef]
- McDonald, D.M.; Choyke, P.L. Imaging of angiogenesis: From microscope to clinic. Nat. Med. 2003, 9, 713–725. [Google Scholar] [CrossRef]
- Weibel, E.R. Stereological Methods; Academic Press: London, UK, 1979. [Google Scholar]
- Ribatti, D. Chicken chorioallantoic membrane angiogenesis model. Methods Mol. Biol. 2012, 843, 47–57. [Google Scholar] [CrossRef]
- Palmer, T.D.; Lewis, J.; Zijlstra, A. Quantitative analysis of cancer metastasis using an avian embryo model. J. Vis. Exp. 2011. [Google Scholar] [CrossRef] [Green Version]
- Zijlstra, A.; Lewis, J.D. Visualization and Quantification of De Novo Angiogenesis in Ex Ovo Chicken Embryos. In The Textbook of Angiogenesis and Lymphangiogenesis: Methods and Applications; Springer: Dordrecht, The Netherlands, 2012; pp. 217–240. [Google Scholar] [CrossRef]
- Kellermeier, M.; Bert, C.; Muller, R.G. A novel concept for CT with fixed anodes (FACT): Medical imaging based on the feasibility of thermal load capacity. Phys. Med. 2015, 31, 425–434. [Google Scholar] [CrossRef]
- Padhani, A.R.; Neeman, M. Challenges for imaging angiogenesis. Br. J. Radiol. 2001, 74, 886–890. [Google Scholar] [CrossRef]
- Agostini, A.; Borgheresi, A.; Mari, A.; Floridi, C.; Bruno, F.; Carotti, M.; Schicchi, N.; Barile, A.; Maggi, S.; Giovagnoni, A. Dual-energy CT: Theoretical principles and clinical applications. Radiol. Med. 2019, 124, 1281–1295. [Google Scholar] [CrossRef]
- Henning, A.L.; Jiang, M.X.; Yalcin, H.C.; Butcher, J.T. Quantitative three-dimensional imaging of live avian embryonic morphogenesis via micro-computed tomography. Dev. Dyn. 2011, 240, 1949–1957. [Google Scholar] [CrossRef] [Green Version]
- Siamblis, D.; Karnabatidis, D.; Hatjikondi, O.; Kalogeropoulou, C.; Kardamakis, D.; Dimopoulos, J. A novel radiological approach for the experimental study of angiogenesis: Angiography of the chick embryo and its chorioallantoic membrane. Eur. J. Radiol. 1996, 21, 220–224. [Google Scholar] [CrossRef]
- Karnabatidis, D.; Dimopoulos, J.C.A.; Siablis, D.; Papazafiropoulos, D.; Kalogeropoulou, C.P.; Nikiforidis, G. Quantification of the ionising radiation effect over angiogenesis in the chick embryo and its chorioallantoic membrane by computerised analysis of angiographic images. Acta Radiol. 2016, 42, 333–338. [Google Scholar] [CrossRef]
- Siablis, D.; Liatsikos, E.N.; Karnabatidis, D.; Kagadis, G.C.; Sakelaropoulos, G.C.; Maroulis, J.; Kardamakis, D.; Athanassopoulos, A.; Perimenis, P.; Nikiforidis, G.; et al. Digital subtraction angiography and computer assisted image analysis for the evaluation of the antiangiogenetic effect of ionizing radiation on tumor angiogenesis. Int. Urol. Nephrol. 2006, 38, 407–411. [Google Scholar] [CrossRef]
- Zhang, X.; Yelbuz, T.M.; Cofer, G.P.; Choma, M.A.; Kirby, M.L.; Johnson, G.A. Improved preparation of chick embryonic samples for magnetic resonance microscopy. Magn. Reson. Med. 2003, 49, 1192–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnick, I.E.; Fowler, C.B.; Mason, J.T.; Potter, K. Novel mineral contrast agent for magnetic resonance studies of bone implants grown on a chick chorioallantoic membrane. Magn. Reson. Imaging 2011, 29, 1244–1254. [Google Scholar] [CrossRef]
- Kivrak Pfiffner, F.; Waschkies, C.; Tian, Y.; Woloszyk, A.; Calcagni, M.; Giovanoli, P.; Rudin, M.; Buschmann, J. A new in vivo magnetic resonance imaging method to noninvasively monitor and quantify the perfusion capacity of three-dimensional biomaterials grown on the chorioallantoic membrane of chick embryos. Tissue Eng. Part. C Methods 2015, 21, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnock, G.; Turtoi, A.; Blomme, A.; Bretin, F.; Bahri, M.A.; Lemaire, C.; Libert, L.C.; Seret, A.E.; Luxen, A.; Castronovo, V.; et al. In vivo PET/CT in a human glioblastoma chicken chorioallantoic membrane model: A new tool for oncology and radiotracer development. J. Nucl. Med. 2013, 54, 1782–1788. [Google Scholar] [CrossRef] [Green Version]
- Kolkman, R.G.M.; Wang, L.V.; Huisjes, A.; Siphanto, R.I.; Steenbergen, W.; van Leeuwen, T.G. Photoacoustic imaging of blood vessels in the chorioallantoic membrane of a chicken embryo. In Proceedings of the Photons Plus Ultrasound: Imaging and Sensing, San Jose, CA, USA, 12 July 2004. [Google Scholar]
- Zeng, L.; Da, X.; Gu, H.; Yang, D.; Yang, S.; Xiang, L. High antinoise photoacoustic tomography based on a modified filtered backprojection algorithm with combination wavelet. Med. Phys. 2007, 34, 556–563. [Google Scholar] [CrossRef]
- Eom, J.; Park, S.J.; Lee, B.H. Noncontact photoacoustic tomography of in vivo chicken chorioallantoic membrane based on all-fiber heterodyne interferometry. J. Biomed. Opt. 2015, 20, 106007. [Google Scholar] [CrossRef] [Green Version]
- Telenko, S.A.; Vargas, G.; Nelson, J.S.; Milner, T.E. Coherent thermal wave imaging of subsurface chromophores in biological materials. Phys. Med. Biol. 2002, 47, 657–671. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Liu, G.; Chen, Z.; De Magalhães, N. In-vivo three-dimensional Doppler variance imaging for tumor angiogenesis on chorioallantoic membrane. In Proceedings of the Dynamics and Fluctuations in Biomedical Photonics VIII, San Francisco, CA, USA, 10 February 2011. [Google Scholar]
- Huang, D.; Swanson, E.A.; Lin, C.P.; Schuman, J.S.; Stinson, W.G.; Chang, W.; Hee, M.R.; Flotte, T.; Gregory, K.; Puliafito, C.A.; et al. Optical coherence tomography. Science 1991, 254, 1178–1181. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, J.G.; Pitris, C.; Boppart, S.A.; Brezinski, M.E. Optical coherence tomography: An emerging technology for biomedical imaging and optical biopsy. Neoplasia 2000, 2, 9–25. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.; Mao, Y.; Flueraru, C.; Sherif, S. Optical coherence tomography: Technology and applications. In Proceedings of the International Conference on Optical Instruments and Technology: Optical Systems and Optoelectronic Instruments, Beijing, China, 26 January 2009. [Google Scholar]
- Chen, Z.; Milner, T.E.; Srinivas, S.; Wang, X.; Malekafzali, A.; van Gemert, M.J.; Nelson, J.S. Noninvasive imaging of in vivo blood flow velocity using optical Doppler tomography. Opt. Lett. 1997, 22, 1119–1121. [Google Scholar] [CrossRef] [PubMed]
- Bonner, R.; Nossal, R. Model for laser Doppler measurements of blood flow in tissue. Appl. Opt. 1981, 20, 2097–2107. [Google Scholar] [CrossRef]
- O’Doherty, J.; McNamara, P.; Clancy, N.T.; Enfield, J.G.; Leahy, M.J. Comparison of instruments for investigation of microcirculatory blood flow and red blood cell concentration. J. Biomed. Opt. 2009, 14, 034025. [Google Scholar] [CrossRef]
- Forrester, K.R.; Tulip, J.; Leonard, C.; Stewart, C.; Bray, R.C. A laser speckle imaging technique for measuring tissue perfusion. IEEE Trans. Biomed. Eng. 2004, 51, 2074–2084. [Google Scholar] [CrossRef]
- Tay, S.L.; Heng, P.W.; Chan, L.W. The CAM-LDPI method: A novel platform for the assessment of drug absorption. J. Pharm. Pharm. 2012, 64, 517–529. [Google Scholar] [CrossRef]
- Leahy, M.J.; de Mul, F.F.; Nilsson, G.E.; Maniewski, R. Principles and practice of the laser-Doppler perfusion technique. Technol. Health Care 1999, 7, 143–162. [Google Scholar] [CrossRef] [PubMed]
- Borozdova, M.A.; Fedosov, I.V.; Tuchin, V.V. Laser Doppler anemometer signal processing for blood flow velocity measurements. Quantum Electron. 2015, 45, 275–282. [Google Scholar] [CrossRef]
- Fercher, A.F.; Briers, J.D. Flow visualization by means of single-exposure speckle photography. Opt. Commun. 1981, 37, 326–330. [Google Scholar] [CrossRef]
- van As, K.; Boterman, J.; Kleijn, C.R.; Kenjeres, S.; Bhattacharya, N. Laser speckle imaging of flowing blood: A numerical study. Phys. Rev. E 2019, 100, 033317. [Google Scholar] [CrossRef]
- Dunn, A.K. Laser speckle contrast imaging of cerebral blood flow. Ann. Biomed. Eng. 2012, 40, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wen, D.; Huang, S.; Gui, S.; Zhang, Z.; Lu, J.; Li, P. Laser speckle contrast imaging of blood flow in the deep brain using microendoscopy. Opt. Lett. 2018, 43, 5627–5630. [Google Scholar] [CrossRef]
- Du, Z.; Zan, T.; Li, H.; Li, Q. A study of blood flow dynamics in flap delay using the full-field laser perfusion imager. Microvasc. Res. 2011, 82, 284–290. [Google Scholar] [CrossRef]
- Forrester, K.R.; Stewart, C.; Tulip, J.; Leonard, C.; Bray, R.C. Comparison of laser speckle and laser Doppler perfusion imaging: Measurement in human skin and rabbit articular tissue. Med. Biol. Eng. Comput. 2002, 40, 687–697. [Google Scholar] [CrossRef]
- Lin, M.-C.; Lai, W.-H.; Tien, C.-H.; Matin, M.A.; Iftekharuddin, K.M.; Awwal, A.A.S.; Márquez, A.; García Vázquez, M. Characterization of retinal blood vessel by laser speckle correlation time constant (Conference Presentation). Proceedings of Optics and Photonics for Information Processing X, San Diego, CA, USA, 2 November 2016. [Google Scholar]
- Bezemer, R.; Klijn, E.; Khalilzada, M.; Lima, A.; Heger, M.; van Bommel, J.; Ince, C. Validation of near-infrared laser speckle imaging for assessing microvascular (re)perfusion. Microvasc. Res. 2010, 79, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Nadort, A.; Kalkman, K.; van Leeuwen, T.G.; Faber, D.J. Quantitative blood flow velocity imaging using laser speckle flowmetry. Sci. Rep. 2016, 6, 25258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, L.; Yan, T.; Xiao, Z.; Xi, J.; Li, Y. Hatching eggs classification based on deep learning. Multimed. Tools Appl. 2017, 77, 22071–22082. [Google Scholar] [CrossRef]
- Lecun, Y.; Bottou, L.; Bengio, Y.; Haffner, P. Gradient-based learning applied to document recognition. Proc. IEEE 1998, 86, 2278–2324. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Geng, L.; Gao, Z.; Li, S.; Fu, X.; Wang, W.; Feng, Y.; Wang, S.; Li, Y.; Fang, Y.; et al. An Innovative Algorithm to Process Imaging Data for Detection of Weak Chick Embryos in Vaccine Production. In Proceedings of the IEEE 5th International Conference on Universal Village UV2020, Bonston, MA, USA, 25 October 2020. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ED | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 12 | 14 | 16 | 18 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Blood Volume (mL) | 0.04 | 0.10 | 0.17 | 0.26 | 0.37 | 0.51 | 0.68 | 1.15 | 2.15 | 3.13 | 2.58 |
| Growth Factors | Description | Functions | Ref. |
|---|---|---|---|
| VEGF | Vascular endothelial growth factor | To exert vascular permeability and endothelial cell recruitment | [37] |
| FGF | Fibroblast growth factor | To elicit fibrocyte proliferation | [11,39,40] |
| PDGF | Platelet-derived growth factor | To stimulate vascular stability | [38] |
| ANG | Angiopoietin | To act on endothelial sprouting, vessel wall remodelling and mural cell recruitment | [37] |
| HGF | Hepatocyte growth factor | As a cytokine to stimulate proliferation and morphogenesis of epithelia | [37] |
| HIF | Hypoxia-inducible factor | To induce expression of VEGF and its receptors | [14,37] |
| Endostain | A proteolytic fragment of collagen XVIII (a component of the basement membrane) | To act as an endogenous anti-angiogenic molecule | [37] |
| Model in Technical Materials | Ref. |
|---|---|
| Filter disks | [74] |
| Culture coverslide glasses | [75] |
| Inert synthetic polymers: Elvax 40 (ethylene-vinyl acetate copolymer) | [76] |
| Hydron (a poly-2-hydroxyethylmethacrylate polymer) | [76] |
| Methylcellulose disks | [77] |
| Alginate pellet | [78] |
| Gelatin sponges | [79] |
| Cross-linked collagen hydrolysate | [80] |
| Cross-linked collagen matrices | [81] |
| Cross-linked and heparinised collagen matrices | [81] |
| Fibrin matrices | [71] |
| Poly(D,L-lactic acid) (PLA) | [82] |
| Poly(ethylene glycol) (PEG) | [83] |
| Modified Matrigel mixtures | [84] |
| Paraffin and plastic embedding | [85] |
| Direct | Indirect |
|---|---|
| Angiostatin, bevacizumab (Avastin) Arresten, canstatin, combrestatin Endostatin, thrombospondin Tumstatin, methoxyestradiol, vitaxin | Targeting EGF-receptor tyrosine kinase Sunitinib Targeting the VEGF receptor Targeting the PDGF receptor Targeting Erb-B2 Receptor Tyrosine Kinase 2 (ERBB-2) Targeting the interferon alpha receptor * |
| Drugs | Type of Tumour | Ref. |
|---|---|---|
| Hydrophobic derivatives of boswellic acid | PC-3 human prostate cancer | [111] |
| Monoclonal antibodies (anti-GD2) or Asn-Gly-Arg (NGR) peptides incorporated in liposomes | Neuroblastoma | [112] |
| Photosensitiser Ce6 | Bladder cancer | [113] |
| Bis(methoxyethyl)-di-n-propylporphycene (BEPPn) | RR 1022, epithelial cells from Rous sarcoma virus (RSV) | [114] |
| Photosensitising drug: methylene blue (MB) | Human malignant ovarian adenocarcinoma | [92] |
| Sema3C | Glioblastoma (U87 MG cells) | [115] |
| Streptokinase and gemcitabine | Lewis lung carcinoma | [103] |
| Neutralising antibody against protein A | Ovarian cancer | [116] |
| Doxorubicin in nanoparticles | Ovarian cancer | [53] |
| VDAs Name | Company | Stage of Clinical Development | Ref. |
|---|---|---|---|
| Tubulin Binding | |||
| CA4P (fosbretabulin) | Mateon Therapeutics | Phase 2/3 | [3,49] |
| C118P | Sanhome Pharmaceutical | Phase 2 | [128] |
| Combretastatin A4 gold derivative | University Bayreuth | New formulation under research | [129] |
| Deoxypodophyllotoxin (DPT) | China Pharmaceutical University | New formulation under research | [130] |
| NPI-2358 (plinabulin) | BeyondSpring | Phase 3 | [3] |
| BNC105P | Bionomics | Phase 2 | [3] |
| EPC2407 (crolibulin) | Immune Pharmaceuticals | Phase 1/2 | [127] |
| OXi4503 (CA1P) | Mateon Therapeutics | Phase 1/2 | [3] |
| CKD-516 | Chong Kun Dang Pharmaceutical | Phase 1 | [3,131] |
| MN-029 | Medicinova | Phase 1 | [3] |
| ENMD-1198 | EntreMed | Phase 1 | [132] |
| C9 | Shanghai Institute of Materia Medica, Chinese Academy of Sciences | New formulation under research | [133] |
| Phenyl-3-(2-chloroethyl) urea (CEU) | IMOTEP Inc. | New formulation under research | [134] |
| BPR0C261 | National Health Research Institutes, Taiwan | New formulation under research | [135] |
| BPR0L075 | National Health Research Institutes, Taiwan | New formulation under research | [136] |
| IMC-038525 | ImClone Systems | New formulation under research | [137] |
| ABT-751 | Abbott | Phase 2 | [3] |
| AVE8062 (ombrabulin) | Sanofi-Aventis | Development terminated | [3] |
| CYT997 (lexibulin) | Gilead | Development terminated | [3] |
| Dolastatin-10 | Marine Biotech | Phase 2 | [3] |
| MPC-6827 (verubulin, Azixa) | Myrexis | Phase 2 | [3] |
| TZT-1027 (soblidotin) | Daiichi-Sankyo | Phase 2 | [3] |
| ZD6126 (ANG453) | AstraZeneca | Phase 2 | [3] |
| Flavonoids | |||
| ASA404 or DMXAA | Antisoma | Phase 3 | [3] |
| VDAs | Test Methods | Ref. |
|---|---|---|
| Combretastatin A4-phosphate (CA4P) | Topical paper | [144] |
| CA4 gold derivative | Topical thin silicon foil | [129] |
| Deoxypodophyllotoxin (DPT) | Topical filter paper disks | [130] |
| C9 | Topical | [133] |
| Verubulin (MPC-6827) | Topical silicon foil ring | [145] |
| Phenyl-3-(2 chloroethyl) urea (CEU) | Intravenous injection | [134] |
| CAM Features | Research Methodology | |
|---|---|---|
| Advantages | Small size and easy to handle | Reproducibility and reliability |
| Contains rich nutrients and intensive angiogenesis capacity | Great accessibility | |
| Rapid vascular growth | Rapid screening platform | |
| Necessary organs isolated within the CAM, in vivo | Broad imaging modalities available | |
| High embryo survival rate | No mobility of animal | |
| Complete circulatory system (intravascular delivery of drugs) | Cost-effectiveness | |
| Naturally immune deficient, different tissues and species transplantations without immune responses | Topical and intravascular administration of drugs | |
| No requirement for animal protocol approval, less animal welfare burden | ||
| Limitations | Chicken origin of the assay limiting availability of reagents | Very thin and fragile, careful manipulation required |
| CAM under rapid change | Complex choice of protocols available | |
| Developed vascular network difficult to be distinguished from new capillaries | Difficult real-time monitoring | |
| Sensitive to environmental factors (temperature, oxygen tension) | Short observation time after treatment | |
| Differences in drug metabolism with mammals | Post-grafting non-specific inflammatory reactions after ED 15 | |
| False angiogenesis due to dust and other irritants | Oral drug administration not feasible |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Wang, S.; Feng, Y.; Zhang, J.; Du, Y.; Zhang, J.; Ongeval, C.V.; Ni, Y.; Li, Y. Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research. Cells 2021, 10, 463. https://doi.org/10.3390/cells10020463
Chen L, Wang S, Feng Y, Zhang J, Du Y, Zhang J, Ongeval CV, Ni Y, Li Y. Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research. Cells. 2021; 10(2):463. https://doi.org/10.3390/cells10020463
Chicago/Turabian StyleChen, Lei, Shuncong Wang, Yuanbo Feng, Jinyong Zhang, Yuqing Du, Jiang Zhang, Chantal Van Ongeval, Yicheng Ni, and Yue Li. 2021. "Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research" Cells 10, no. 2: 463. https://doi.org/10.3390/cells10020463
APA StyleChen, L., Wang, S., Feng, Y., Zhang, J., Du, Y., Zhang, J., Ongeval, C. V., Ni, Y., & Li, Y. (2021). Utilisation of Chick Embryo Chorioallantoic Membrane as a Model Platform for Imaging-Navigated Biomedical Research. Cells, 10(2), 463. https://doi.org/10.3390/cells10020463