
Spermatogenic Activity and Sperm Traits in Post-Pubertal and Adult Tomcats (Felis catus): Implication of Intra-Male Variation in Sperm Size

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Sample Collection and Sperm Recovery
2.3. Sperm Analyses
2.3.1. Motility
2.3.2. Plasma Membrane Integrity and Acrosomal Status
2.3.3. Mitochondrial Activity
2.3.4. DNA Integrity
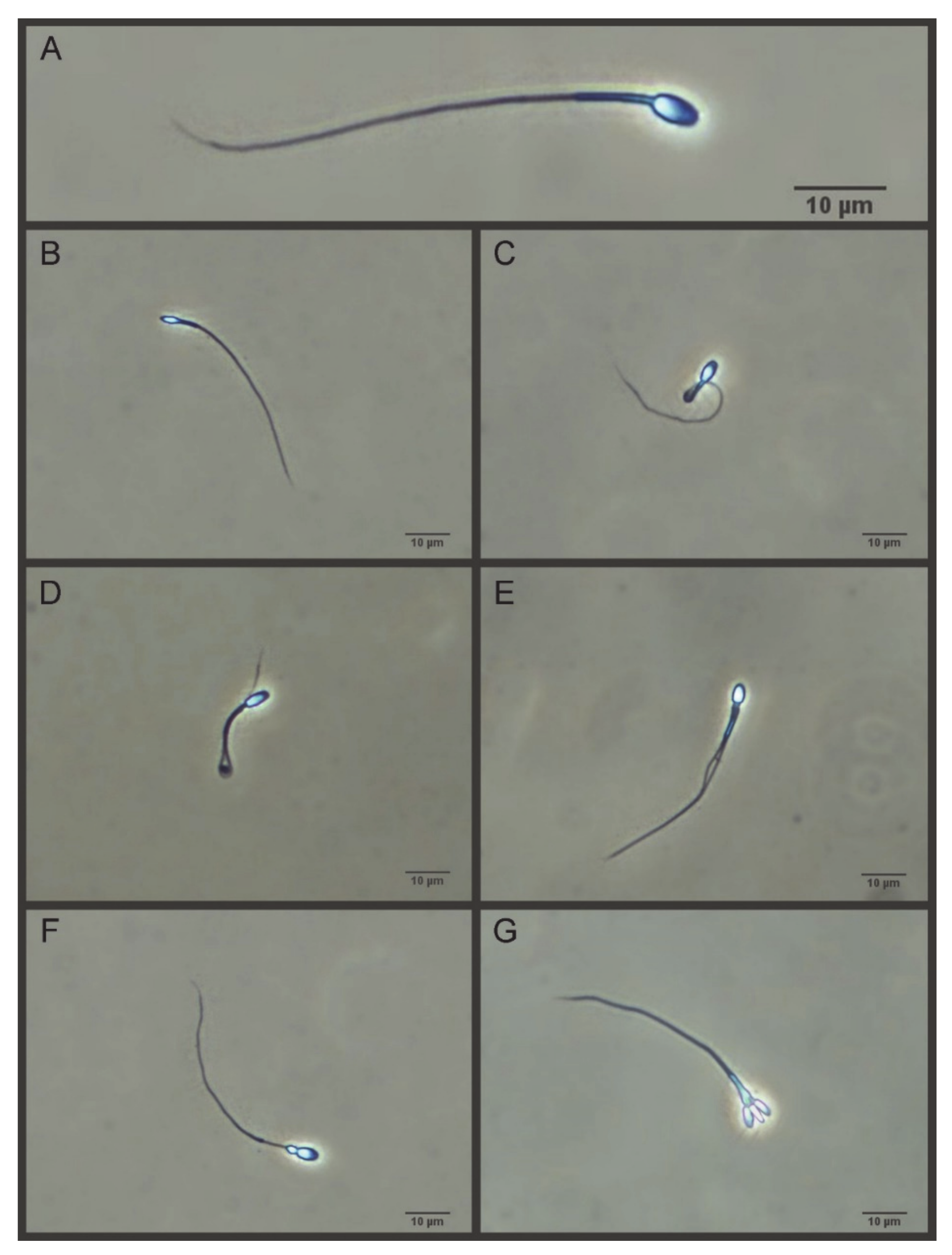
2.3.5. Morphometry and Morphology
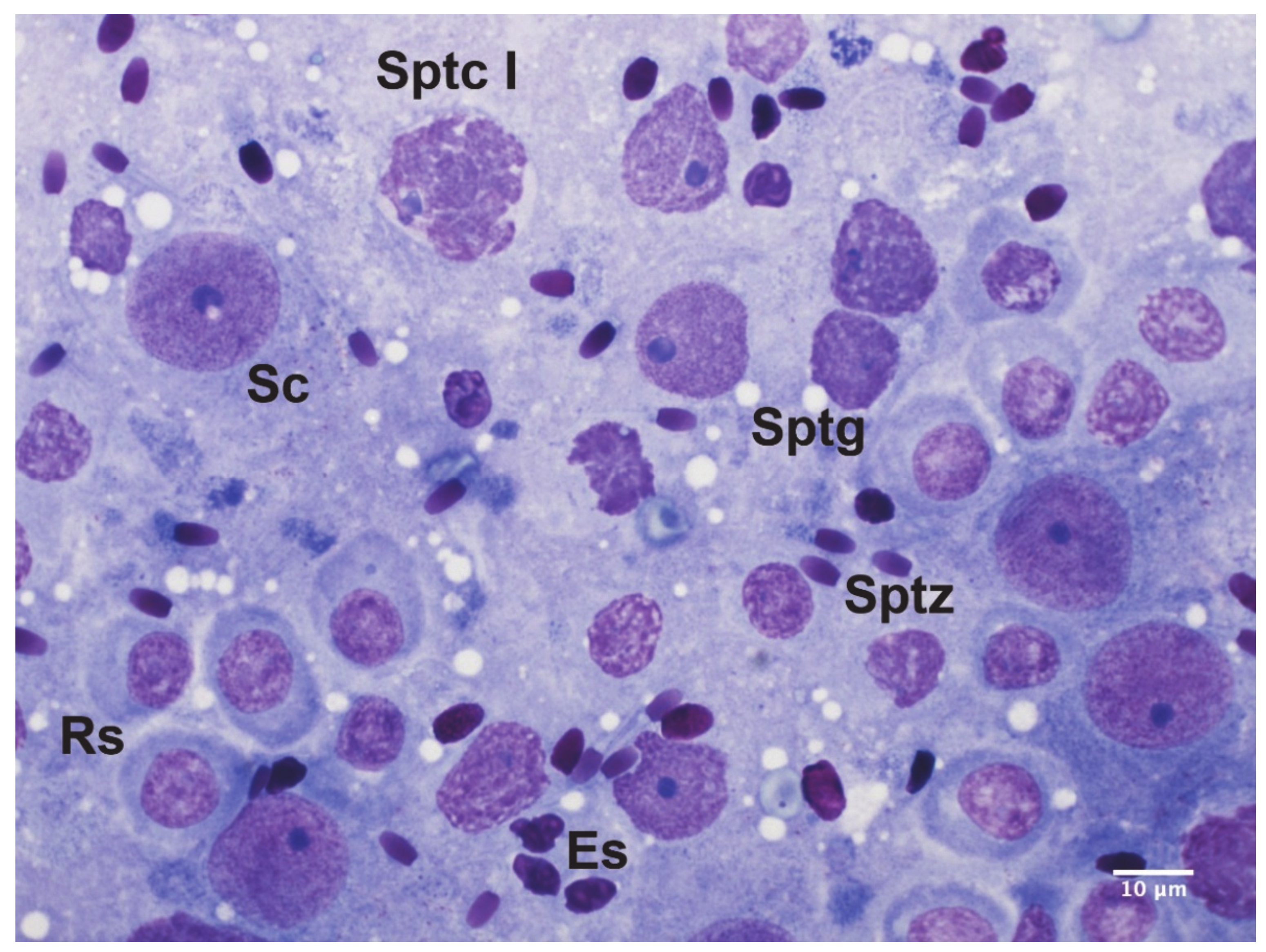
2.4. Spermatogenesis Assessment
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kitchener, A.C.; Breitenmoser-Würsten, C.H.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the Felidae. The final report of the Cat Classification Task Force of the IUCN/SSC Cat Specialist Group. Cat News 2017, 11, 1–80. [Google Scholar]
- The IUCN Red List of Threatened Species. Available online: www.iucnredlist.org (accessed on 17 December 2020).
- Bradshaw, J. Normal feline behavior... and why problem behaviours develop. J. Feline Med. Surg. 2018, 20, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Andrews, C.G.; Thomas, D.G.; Yapura, J.; Potter, M.A. Reproductive biology of the 38 extant felid species: A review. Mammal Rev. 2019, 49, 16–30. [Google Scholar] [CrossRef] [Green Version]
- Thongphakdee, A.; Sukparangsi, W.; Comizzoli, P.; Chatdarong, K. Reproductive biology and biotechnology in wild felids. Theriogenology 2020, 150, 360–373. [Google Scholar] [CrossRef] [PubMed]
- França, L.R.; Godinho, C.L. Testis morphometry, seminiferous epithelium cycle length, and daily sperm production in domestic cats (Felis catus). Biol. Reprod. 2003, 68, 1554–1561. [Google Scholar] [CrossRef] [Green Version]
- Tsutsui, T.; Kuwabara, S.; Kuwabara, K.; Kugota, Y.; Kinjo, T.; Hori, T. Development of spermatogenic function in the sex maturation process in male cats. J. Vet. Med. Sci. 2004, 66, 1125–1127. [Google Scholar] [CrossRef] [Green Version]
- Siemieniuch, M.J.; Wocławek-Potocka, I. Morphological features of the seminiferous epithelium in cat (Felis catus, L. 1758) testes. J. Reprod. Dev. 2007, 53, 1125–1130. [Google Scholar] [CrossRef] [Green Version]
- Cocchia, N.; Ciani, F.; El-Rass, R.; Russo, M.; Borzacchiello, G.; Esposito, V.; Montagnaro, S.; Avallone, L.; Tortora, G.; Lorizio, R. Cryopreservation of feline epididymal spermatozoa from dead and alive animals and its use in assisted reproduction. Zygote 2010, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- De Sousa Barbosa, B.; Rodrigues Silva, H.V.; Evaristo de Almeida Tabosa, B.; Gothardo Pereira Nunes, T.; de Magalhães, F.F.; Daniel Machado da Silva, L. Morphological and morphometric characterization of domestic cat epididymal sperm. Reprod. Domest. Anim. 2019, 54, 1630–1636. [Google Scholar] [CrossRef] [PubMed]
- Siemieniuch, M.J. Apoptotic changes in the epithelium germinativum of the cat (Felis catus s. domestica, L. 1758) at different ages and breeding seasons. Reprod. Domest. Anim. 2008, 43, 473–476. [Google Scholar] [CrossRef]
- Schön, J.; Neumann, S.; Wildt, D.E.; Pukazhenthi, B.S.; Jewgenow, K. Localization of oestrogen receptors in the epididymis during sexual maturation of the domestic cat. Reprod. Domest. Anim. 2009, 44, 294–301. [Google Scholar] [CrossRef]
- Braun, B.C.; Okuyama, M.W.; Müller, K.; Dehnhard, M.; Jewgenow, K. Steroidogenic enzymes, their products and sex steroid receptors during testis development and spermatogenesis in the domestic cat (Felis catus). J. Steroid. Biochem. Mol. Biol. 2018, 178, 135–149. [Google Scholar] [CrossRef]
- Filliers, M.; Rijsselaere, T.; Bossaert, P.; De Causmaecker, V.; Dewulf, J.; Pope, C.E.; Van Soom, A. Computer-assisted sperm analysis of fresh epididymal cat spermatozoa and the impact of cool storage (4 °C) on sperm quality. Theriogenology 2008, 70, 1550–1559. [Google Scholar] [CrossRef]
- Ros-Santaella, J.L.; Pintus, E. Rooibos (Aspalathus linearis) extract enhances boar sperm velocity up to 96 hours of semen storage. PLoS ONE 2017, 12, e0183682. [Google Scholar] [CrossRef] [Green Version]
- Pintus, E.; Jovičić, M.; Kadlec, M.; Ros-Santaella, J.L. Divergent effect of fast- and slow-releasing H2S donors onboar spermatozoa under oxidative stress. Sci. Rep. 2020, 10, 6508. [Google Scholar] [CrossRef]
- Ros-Santaella, J.L.; Kadlec, M.; Pintus, E. Pharmacological activity of honeybush (Cyclopia intermedia) in boar spermatozoa during semen storage and under oxidative stress. Animals 2020, 10, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, R.A.P.; Vickers, S.E. Use of fluorescent probes to assess membrane integrity in mammalian spermatozoa. J. Reprod. Fertil. 1990, 88, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Cheuquemán, C.; Faúndez, R.; Sánchez, R.; Risopatrón, J. Changes in sperm function and structure after freezing in domestic cat spermatozoa. Andrologia 2018, 50, e13080. [Google Scholar] [CrossRef]
- Thuwanut, P.; Chatdarong, K.; Techakumphu, M.; Axnér, E. The effect of antioxidants on motility, viability, acrosome integrity and DNA integrity of frozen-thawed epididymal cat spermatozoa. Theriogenology 2008, 70, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Ros-Santaella, J.L.; Domínguez-Rebolledo, A.E.; Garde, J.J. Sperm flagellum volume determines freezability in red deer spermatozoa. PLoS ONE 2014, 9, e112382. [Google Scholar] [CrossRef] [Green Version]
- Ros-Santaella, J.L.; Pintus, E.; Garde, J.J. Intramale variation in sperm size: Functional significance in a polygynous mammal. PeerJ 2015, 3, e1478. [Google Scholar] [CrossRef] [Green Version]
- Leme, D.P.; Visacre, E.; Castro, V.B.; Lopes, M.D. Testicular cytology by fine needle aspiration in domestic cats. Theriogenology 2018, 106, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Pintus, E.; Ros-Santaella, J.L.; Garde, J.J. Beyond Testis Size: Links between Spermatogenesis and Sperm Traits in a Seasonal Breeding Mammal. PLoS ONE 2015, 10, e0139240. [Google Scholar] [CrossRef] [PubMed]
- Bodet, L.; Albaric, O.; Topie, E.; Dagher, E.; Chocteau, F.; Gogny, A. Cytological description of testicular cell populations in sexually mature cats with normal spermatogenesis. Reprod. Domest. Anim. 2020, 55, 1–20. [Google Scholar] [CrossRef]
- Ros-Santaella, J.L.; Kotrba, R.; Pintus, E. High-energy diet enhances spermatogenic function and increases sperm midpiece length in fallow deer (Dama dama) yearlings. R. Soc. Open Sci. 2019, 6, 181972. [Google Scholar] [CrossRef] [Green Version]
- Stelletta, C.; Juyena, N.S.; Ponce Salazar, D.; Ruiz, J.; Gutierrez, G. Testicular cytology of alpaca: Comparison between impressed and smeared slides. Anim. Reprod. Sci. 2011, 125, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Pintus, E.; Ros-Santaella, J.L. Assessment of reproductive traits in male gemsbok (Oryx gazella) (Bovidae, Hippotraginae). Spixiana 2018, 41, 205–209. [Google Scholar]
- Da Paz, R.C.R.; Leme, D.P.; Zůge, R.M.; Pessuti, C.; Santos, E.F.; Barnabe, R.C. Testicular fine needle aspiration cytology as a diagnostic tool in jaguar (Panthera onca) infertility. Braz. J. Vet. Res. Anim. Sci. 2003, 40, 100–107. [Google Scholar]
- Neubauer, K.; Jewgenow, K.; Blottner, S.; Wildt, D.E.; Pukazhenthi, B.S. Quantity rather than quality in teratospermic males: A histomorphometric and flow cytometric evaluation of spermatogenesis in the domestic cat (Felis catus). Biol. Reprod. 2004, 71, 1517–1524. [Google Scholar] [CrossRef]
- De Barros, J.B.; de Paula, T.A.; da Matta, S.L.; Fonseca, C.C.; Leite, F.L.; Rossi, J.L., Jr.; de Oliveira, P.C.; da Costa, E.P. Sertoli cell index and spermatic reserves in adult captive African lions (Panthera leo, Linnaeus, 1758). Anim. Reprod. Sci. 2007, 102, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.M.; Chiarini-Garcia, H.; Morato, R.G.; Alvarenga, R.L.; França, L.R. Duration of spermatogenesis and daily sperm production in the jaguar (Panthera onca). Theriogenology 2008, 70, 1136–1146. [Google Scholar] [CrossRef]
- De Azevedo, M.H.; de Paula, T.A.; Matta, S.L.; Fonseca, C.C.; da Costa, E.P.; Costa, D.S.; Peixoto, J.V. Cell population indexes of spermatogenic yield and testicular sperm reserves in adult jaguars (Panthera onca). Anim. Reprod. Sci. 2010, 118, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Guião-Leite, F.L. Análise Morfológica do Testículo e do Processo Espermatogênico da Onça Parda (Puma concolor, Wozencraft, 1993) adulta. Master’s Thesis, Universidade Federal de Viçosa, Minas Gerais, Brazil, 2002. [Google Scholar]
- Silva, R.C.; Costa, G.M.; Andrade, L.M.; França, L.R. Testis stereology, seminiferous epithelium cycle length, and daily sperm production in the ocelot (Leopardus pardalis). Theriogenology 2010, 73, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Rodríguez, J. DNA replication and germ cell apoptosis during spermatogenesis in the cat. J. Androl. 2002, 23, 484–490. [Google Scholar] [PubMed]
- Silva, H.V.R.; Nunes, T.G.P.; Ribeiro, L.R.; Freitas, L.A.; de Oliveira, M.F.; Assis Neto, A.C.; Silva, A.R.; Silva, L.D.M.D. Morphology, morphometry, ultrastructure, and mitochondrial activity of jaguar (Panthera onca) sperm. Anim. Reprod. Sci. 2019, 203, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Das Neves, J.L.; Wagenaar, I. 2018. Morphology and ultrastructure of the testes and spermatozoa of the South African leopard (Panthera pardus). Acta Zoologica 2019, 100, 109–118. [Google Scholar] [CrossRef]
- Schmehl, M.L.; Graham, E.F. Ultrastructure of the domestic tom cat (Felis domestica) and tiger (Panthera tigris altaica) spermatozoa. Theriogenology 1989, 31, 861–874. [Google Scholar] [CrossRef]
- Oliveira, P.F.; Alves, M.G. The Sertoli cell at a glance. In Sertoli Cell Metabolism and Spermatogenesis, 1st ed.; Oliveira, P.F., Alves, M.G., Eds.; Springer Publisher: Heidelberg, Germany, 2015; pp. 3–12. ISBN 978-3-319-19790-6. [Google Scholar] [CrossRef]
- Gutiérrez-Reinoso, M.A.; García-Herreros, M. Normozoospermic versus teratozoospermic domestic cats: Differential testicular volume, sperm morphometry, and subpopulation structure during epididymal maturation. Asian J. Androl. 2016, 18, 871–878. [Google Scholar] [CrossRef]
- Alves, I.P.; Cancelli, C.H.B.; Grassi, T.L.M.; Oliveira, P.R.H.; Franciscato, D.A.; Carreira, J.T.; Koivisto, M.B. Evaluation of sperm head dimensions and chromatin integrity of epididymal sperm from domestic cats using the toluidine blue technique. Anim. Reprod. Sci. 2018, 197, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Hingst, O.; Blottner, S.; Franz, C. Chromatin condensation in cat spermatozoa during epididymal transit as studied by aniline blue and acridine orange staining. Andrologia 1995, 27, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, A.I.; Fioratti, E.G.; Ramos, R.S.; Neves, R.C.; Cardoso, G.S.; Landim-Alvarenga, F.C.; Lopes, M.D. High incidence of “Dag-like” sperm defect in the domestic cat. J. Feline Med. Surg. 2013, 15, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Vernocchi, V.; Morselli, M.G.; Lange Consiglio, A.; Faustini, M.; Luvoni, G.C. DNA fragmentation and sperm head morphometry in cat epididymal spermatozoa. Theriogenology 2014, 82, 982–987. [Google Scholar] [CrossRef] [PubMed]
- Gravance, C.G.; Champion, Z.J.; Casey, P.J. Computer-assisted sperm head morphometry analysis (ASMA) of cryopreserved ram spermatozoa. Theriogenology 1998, 49, 1219–1230. [Google Scholar] [CrossRef]
- Malo, A.F.; Garde, J.J.; Soler, A.J.; García, A.J.; Gomendio, M.; Roldan, E.R. Male fertility in natural populations of red deer is determined by sperm velocity and the proportion of normal spermatozoa. Biol. Reprod. 2005, 72, 822–829. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Post-Pubertal | Adult | p Value |
|---|---|---|---|
| Body mass (kg) | 4.28 ± 0.62 | 4.29 ± 0.56 | 0.980 |
| Testis mass (g) | 1.24 ± 0.19 | 1.49 ± 0.52 | 0.275 |
| Epididymis mass (g) | 0.27 ± 0.05 | 0.31 ± 0.06 | 0.071 |
| Parameter | Post-Pubertal | Adult | p Value |
|---|---|---|---|
| Spermatogenic subtypes (%) | |||
| Spermatogonia | 1.61 ± 0.88 | 1.53 ± 0.85 | 0.870 |
| Primary spermatocytes | 15.34 ± 5.62 | 14.57 ± 4.46 | 0.918 |
| Secondary spermatocytes | 0.60 ± 0.52 | 0.60 ± 0.40 | 0.918 |
| Round spermatids | 35.21 ± 3.18 | 34.87 ± 4.89 | 0.870 |
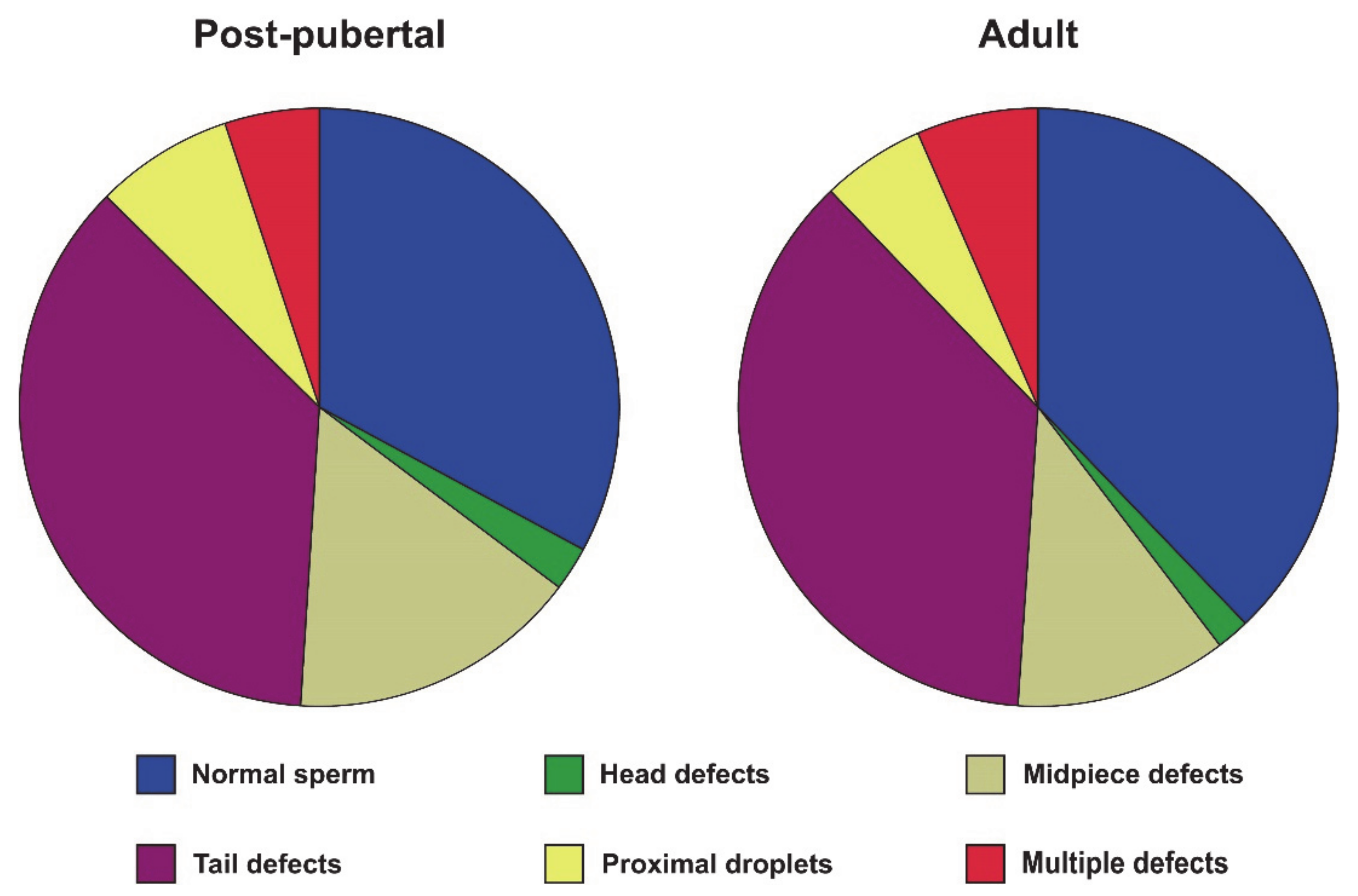
| Elongated spermatids | 26.03 ± 4.75 | 27.67 ± 5.63 | 0.536 |
| Spermatozoa | 21.22 ± 5.61 | 20.75 ± 7.66 | 0.888 |
| Spermatogenic indices | |||
| SEI (%) | 7.00 ± 4.65 | 5.92 ± 3.45 | 0.758 |
| SSEI | 4.46 ± 3.16 | 4.89 ± 3.38 | 0.799 |
| MI | 2.58 ± 0.94 | 2.53 ± 0.66 | 0.905 |
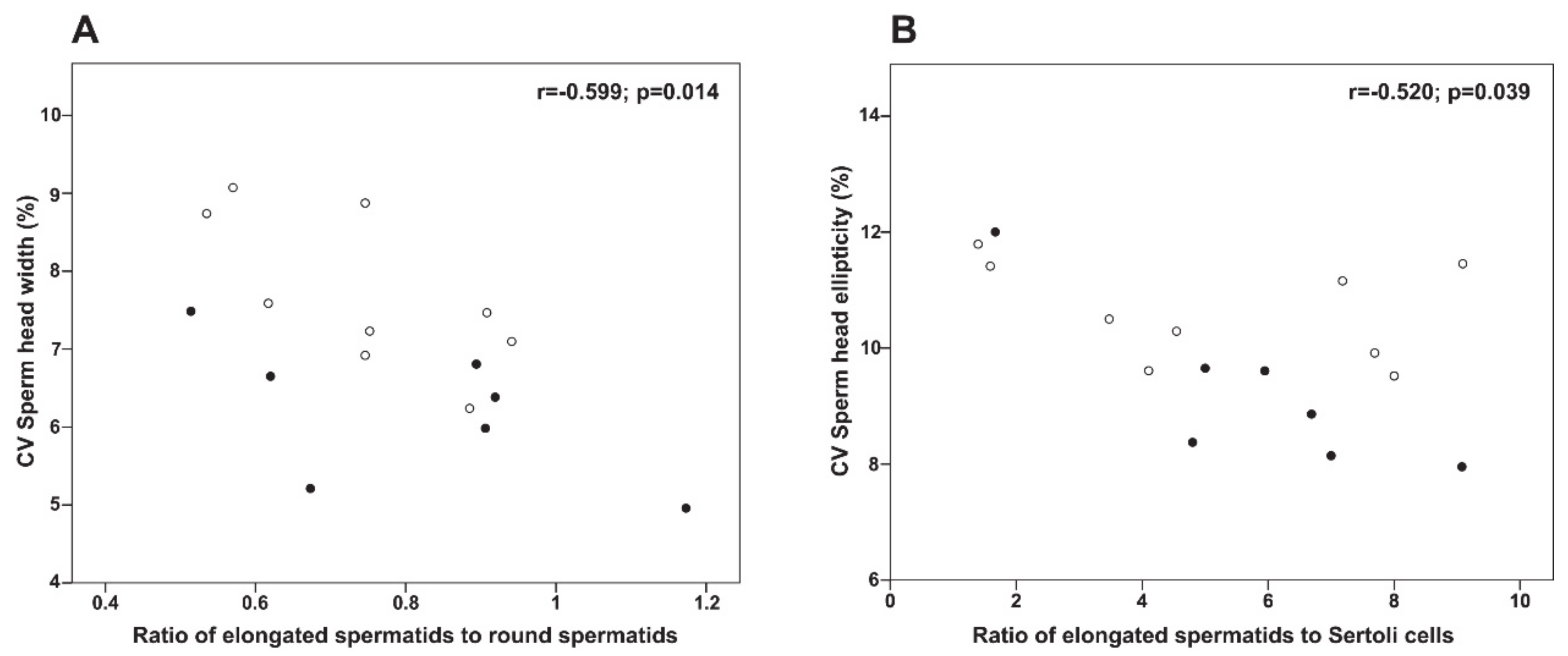
| ES/RS | 0.74 ± 0.15 | 0.81 ± 0.22 | 0.467 |
| ES/GC | 0.26 ± 0.05 | 0.28 ± 0.06 | 0.532 |
| RS/SC | 7.07 ± 3.79 | 7.24 ± 3.20 | 0.928 |
| ES/SC | 5.23 ± 2.86 | 5.74 ± 2.30 | 0.706 |
| GC/SC | 19.78 ± 10.41 | 21.07 ± 9.30 | 0.801 |
| Parameter | Post-Pubertal | Adult | p Value |
|---|---|---|---|
| Concentration (106/mL) | 40.60 ± 26.37 | 92.48 ± 36.85 | 0.016 |
| Intact plasma membrane (%) | 83.94 ± 4.97 | 80.79 ± 7.96 | 0.346 |
| Intact acrosome (%) | 87.50 ± 4.65 | 88.14 ± 6.24 | 0.816 |
| Active mitochondria (%) | 68.33 ± 9.87 | 62.86 ± 7.77 | 0.249 |
| Intact DNA (%) | 88.50 ± 12.14 | 92.00 ± 4.80 | 0.837 |
| Total motility (%) | 63.15 ± 18.74 | 58.19 ± 18.25 | 0.604 |
| Progressive motility (%) | 50.14 ± 15.01 | 47.33 ± 11.57 | 0.688 |
| VAP (µm/s) | 97.81 ± 32.65 | 99.04 ± 19.46 | 0.932 |
| VCL (µm/s) | 151.93 ± 34.14 | 158.74 ± 20.05 | 0.648 |
| VSL (µm/s) | 84.82 ± 25.22 | 87.39 ± 14.77 | 0.814 |
| ALH (µm) | 5.61 ± 1.43 | 5.69 ± 0.70 | 0.899 |
| BCF (Hz) | 15.96 ± 3.05 | 16.00 ± 1.98 | 0.978 |
| LIN (%) | 52.03 ± 8.67 | 52.43 ± 6.39 | 0.536 |
| STR (%) | 85.12 ± 6.17 | 86.76 ± 6.64 | 0.918 |
| Parameter | Sp4 | Sp3 | Sp2 | Sp1 |
|---|---|---|---|---|
| VAP (µm/s) | 138.07 ± 18.16 b | 76.11 ± 23.87 c | 177.10 ± 27.62 a | 6.09 ± 7.17 d |
| VCL (µm/s) | 201.23 ± 33.37 b | 136.34 ± 37.73 c | 235.22 ± 38.52 a | 20.54 ± 21.96 d |
| VSL (µm/s) | 126.64 ± 21.83 a | 70.25 ± 23.57 c | 107.71 ± 59.21 b | 3.48 ± 5.32 d |
| ALH (µm) | 7.28 ± 1.48 b | 4.51 ± 1.67 c | 9.37 ± 1.95 a | 1.02 ± 1.35 d |
| BCF (Hz) | 19.48 ± 7.67 b | 14.90 ± 7.86 c | 20.87 ± 4.96 a | 6.79 ± 3.36 d |
| LIN (%) | 64.01 ± 12.20 a | 51.87 ± 12.79 b | 45.31 ± 23.62 c | 14.15 ± 11.64 d |
| STR (%) | 91.63 ± 9.43 a | 92.42 ± 10.27 a | 58.45 ± 27.70 b | 44.11 ± 25.22 c |
| Post-pubertal (%) | 25.64 ± 15.45 | 21.38 ± 10.37 | 7.36 ± 11.48 | 45.61 ± 21.60 |
| Adult (%) | 24.40 ± 11.17 | 21.31 ± 10.37 | 4.48 ± 8.89 | 49.80 ± 16.84 |
| Parameter | Post-Pubertal | Adult | p Value |
|---|---|---|---|
| Head length (µm) | 4.86 ± 0.26 | 4.97 ± 0.29 | 0.436 |
| Head width (µm) | 2.47 ± 0.10 | 2.42 ± 0.17 | 0.483 |
| Head perimeter (µm) | 11.84 ± 0.49 | 11.97 ± 0.53 | 0.598 |
| Head area (µm2) | 9.44 ± 0.65 | 9.46 ± 0.80 | 0.952 |
| Head ellipticity | 1.98 ± 0.13 | 2.07 ± 0.20 | 0.307 |
| §Midpiece width (µm) | 0.69 ± 0.04 | 0.69 ± 0.03 | 0.939 |
| Midpiece length (µm) | 7.67 ± 0.18 | 7.79 ± 0.24 | 0.268 |
| Principal piece length (µm) | 40.99 ± 1.59 | 40.06 ± 1.48 | 0.253 |
| Terminal piece length (µm) | 4.20 ± 0.57 | 4.16 ± 0.74 | 0.919 |
| Flagellum length (µm) | 52.85 ± 1.66 | 52.01 ± 1.98 | 0.369 |
| Sperm length (µm) | 57.72 ± 1.64 | 56.98 ± 2.12 | 0.447 |
| Flagellum volume (µm3) | 6.64 ± 0.75 | 6.53 ± 0.52 | 0.749 |
| Parameter | Post-Pubertal | Adult | p Value |
|---|---|---|---|
| Head length (%) | 7.00 ± 1.17 | 6.64 ± 1.05 | 0.535 |
| Head width (%) | 7.69 ± 0.98 | 6.21 ± 0.90 | 0.008 |
| Head perimeter (%) | 5.47 ± 0.92 | 5.24 ± 0.75 | 0.610 |
| Head area (%) | 10.21 ± 1.53 | 9.18 ± 0.91 | 0.139 |
| Head ellipticity (%) | 10.63 ± 0.85 | 9.23 ± 1.39 | 0.026 |
| §Midpiece width (%) | 10.58 ± 2.22 | 10.45 ± 1.61 | 0.901 |
| Midpiece length (%) | 3.89 ± 1.46 | 4.65 ± 1.83 | 0.369 |
| Principal piece length (%) | 3.56 ± 1.42 | 3.21 ± 0.51 | 1.000 |
| Terminal piece length (%) | 15.06 ± 3.78 | 16.23 ± 2.64 | 0.496 |
| Flagellum length (%) | 3.12 ± 1.36 | 2.55 ± 0.42 | 0.536 |
| Sperm length (%) | 2.99 ± 1.15 | 2.43 ± 0.34 | 0.408 |
| Flagellum volume (%) | 21.43 ± 4.93 | 21.21 ± 3.52 | 0.922 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintus, E.; Kadlec, M.; Karlasová, B.; Popelka, M.; Ros-Santaella, J.L. Spermatogenic Activity and Sperm Traits in Post-Pubertal and Adult Tomcats (Felis catus): Implication of Intra-Male Variation in Sperm Size. Cells 2021, 10, 624. https://doi.org/10.3390/cells10030624
Pintus E, Kadlec M, Karlasová B, Popelka M, Ros-Santaella JL. Spermatogenic Activity and Sperm Traits in Post-Pubertal and Adult Tomcats (Felis catus): Implication of Intra-Male Variation in Sperm Size. Cells. 2021; 10(3):624. https://doi.org/10.3390/cells10030624
Chicago/Turabian StylePintus, Eliana, Martin Kadlec, Barbora Karlasová, Marek Popelka, and José Luis Ros-Santaella. 2021. "Spermatogenic Activity and Sperm Traits in Post-Pubertal and Adult Tomcats (Felis catus): Implication of Intra-Male Variation in Sperm Size" Cells 10, no. 3: 624. https://doi.org/10.3390/cells10030624
APA StylePintus, E., Kadlec, M., Karlasová, B., Popelka, M., & Ros-Santaella, J. L. (2021). Spermatogenic Activity and Sperm Traits in Post-Pubertal and Adult Tomcats (Felis catus): Implication of Intra-Male Variation in Sperm Size. Cells, 10(3), 624. https://doi.org/10.3390/cells10030624