
A Molecular Networking Strategy: High-Throughput Screening and Chemical Analysis of Brazilian Cerrado Plant Extracts against Cancer Cells

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
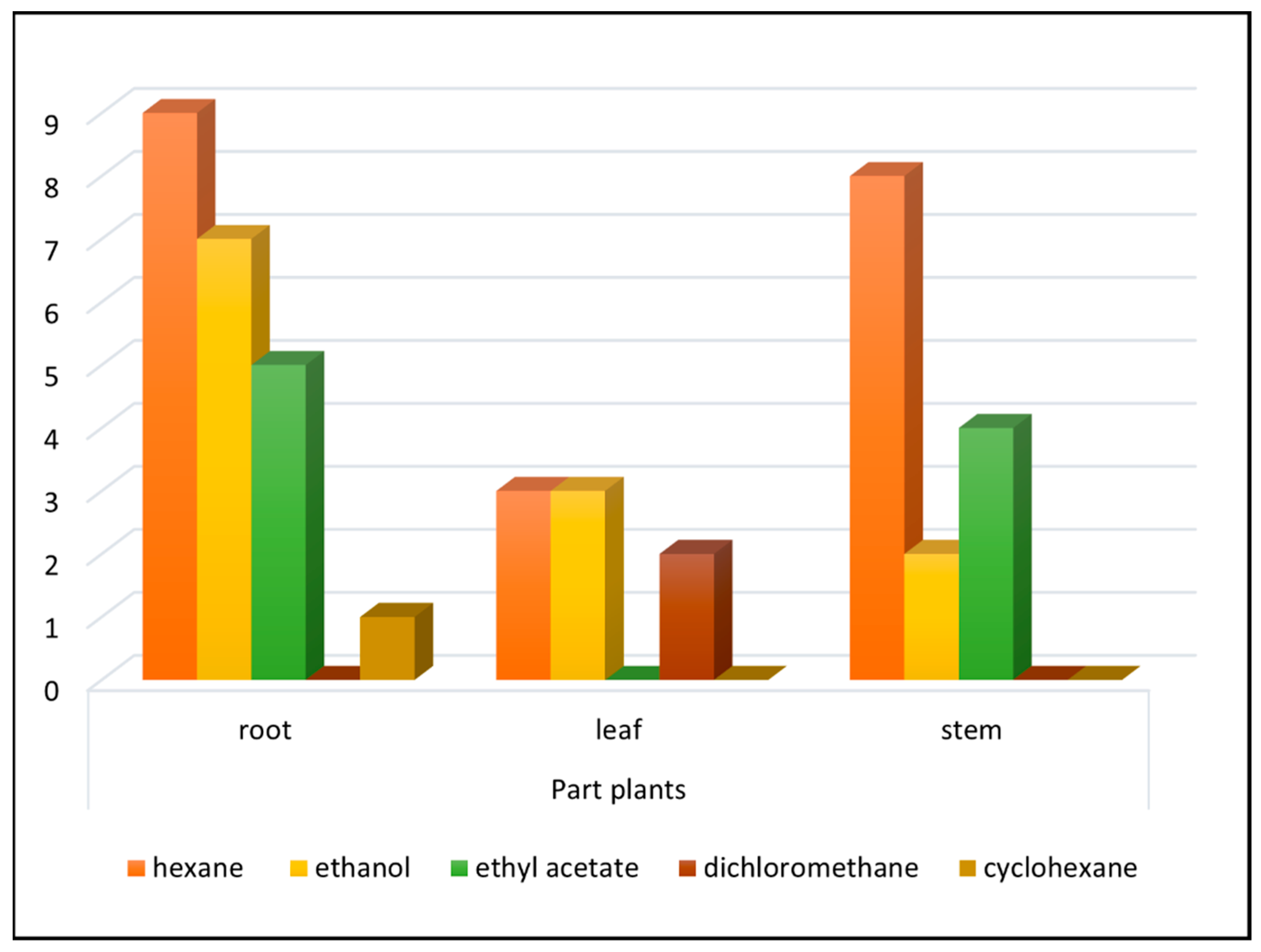
2.1. Plant Extracts
2.2. Antiproliferative Bioassays
2.3. HRESIMS Data Acquisition
2.4. Data Processing and Molecular Networking Construction
3. Results
3.1. High-Throughput Screening
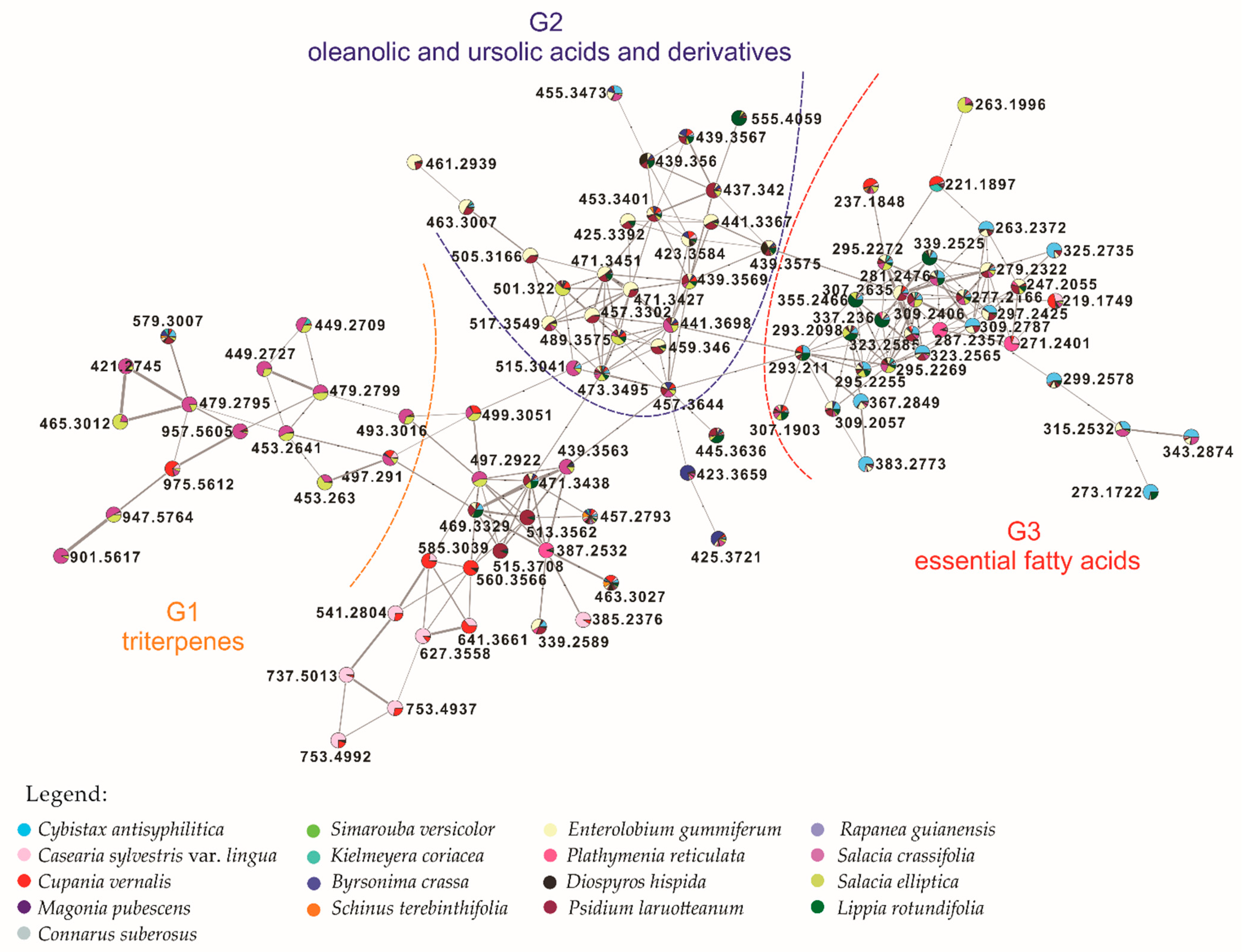
3.2. Molecular Networking
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Lahsen, M.; Mercedes, M.C.; Bustamante, M.M.C.; Dalla-Nora, E.L. Undervaluing and Overexploiting the Brazilian Cerrado at Our Peril. Environ. Sci. Policy Sustain. Dev. 2016, 58, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.X.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechn. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilon, A.C.; Selegato, D.M.; Fernandes, R.P.; Bueno, P.C.P.; Pinho, D.R.; Neto, F.C.; Freire, R.T.; Castro-Gamboa, I.; Bolzani, V.S.; Lopes, N.P. Metabolômica de Plantas: Métodos e Desafios. Quim. Nova 2020, 43, 329–354. [Google Scholar] [CrossRef]
- Demarque, D.P.; Dusi, R.G.; de Sousa, F.D.M.; Grossi, S.M.; Silverio, M.R.S.; Lopes, N.P.; Espindola, L.S. Mass Spectrometry-Based Metabolomics Approach in the Isolation of Bioactive Natural Products. Sci. Rep. 2020, 10, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosun, F.; Mihoglugil, F.; Beutler, J.A.A.; Eroglu Ozkan, E.; Miski, M. Neopapillarine, an Unusual Coumarino-Alkaloid from the Root Extract of Neocryptodiscus papillaris with Cytotoxic Activity on Renal Cancer Cells. Molecules 2020, 25, 3040. [Google Scholar] [CrossRef] [PubMed]
- Devkota, K.P.; Covell, D.; Ransom, T.; McMahon, J.B.; Beutler, J.A. Growth Inhibition of Human Colon Carcinoma Cells by Sesquiterpenoids and Tetralones of Zygogynum Calothyrsum. J. Nat. Prod. 2013, 76, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Long, S.A.; Huang, S.; Kambala, A.; Ren, L.; Wilson, J.; Goetz, M.; Hao, X.; Yang, X.; Goncharova, E.I.; Jia, L.; et al. Identification of Potential Modulators Of Osteosarcoma Metastasis by High-Throughput Cellular Screening of Natural Products. Chem. Biol. Drug Des. 2021, 97, 77–86. [Google Scholar] [CrossRef]
- Henrich, C.J.; Budhu, A.; Yu, Z.; Evans, J.R.; Goncharova, E.I.; Ransom, T.T.; Wang, X.W.; McMahon, J.B. High-Throughput Screening for Identification of Inhibitors of EpCAM-Dependent Growth of Hepatocellular Carcinoma Cells. Chem. Biol. Drug Des. 2013, 82, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoemaker, R.H. The NCI60 Human Tumour Cell Line Anticancer Drug Screen. Nat. Rev. Cancer 2006, 6, 813–823. [Google Scholar] [CrossRef]
- Su, G.; Burant, C.F.; Beecher, C.W.; Athey, B.D.; Meng, F. Integrated Metabolome and Transcriptome Analysis of the NCI60 Dataset. BMC Bioinform. 2011, 12, 36. [Google Scholar] [CrossRef] [Green Version]
- Holbeck, S.L.; Collins, J.M.; Doroshow, J.H. Analysis of Food and Drug Administration-Approved Anticancer Agents in the NCI60 Panel of Human Tumor Cell Lines. Mol. Cancer Ther 2010, 9, 1451–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EMBRAPA. Empresa Brasileira de Pesquisa Agropecuária. Mapa de Solos do Distrito Federal (1:100.000); EMBRAPA-PI. Serviço de Produção de Informação: Brasília, Brazil, 1978. [Google Scholar]
- United States. Science, & Education Administration. Soil taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; US Department of Agriculture: Beltsville, MD, USA, 1975.
- Tang, H.F.; Yun, J.; Lin, H.W.; Chen, X.L.; Wang, X.J.; Cheng, G. Two New Triterpenoid Saponins Cytotoxic to Human Glioblastoma U251MG Cells from Ardisia Pusilla. Chem. Biodivers. 2009, 6, 1443–1452. [Google Scholar] [CrossRef]
- Dong, W.; Liu, X.; Li, X.; Yang, D.; Ding, L. A New Triterpene Saponin. Andr. Integr. Fitoter. 2011, 82, 782–785. [Google Scholar] [CrossRef]
- Katajamaa, M.; Miettinen, J.; Oresic, M. MZmine: Toolbox for Processing and Visualization of Mass Spectrometry Based Molecular Profile Data. Bioinformatics 2006, 22, 634–636. [Google Scholar] [CrossRef] [Green Version]
- Olivon, F.; Grelier, G.; Roussi, F.; Litaudon, M.; Touboul, D. MZmine 2 Data-Preprocessing to Enhance Molecular Networking Reliability. Anal. Chem. 2017, 89, 7836–7840. [Google Scholar] [CrossRef] [PubMed]
- Myers, O.D.; Sumner, S.J.; Li, S.; Barnes, S.; Du, X. One Step Forward for Reducing False Positive and False Negative Compound Identifications from Mass Spectrometry Metabolomics Data: New Algorithms for Constructing Extracted Ion Chromatograms and Detecting Chromatographic Peaks. Anal. Chem. 2017, 89, 8696–8703. [Google Scholar] [CrossRef]
- Nothias, L.F.; Petras, D.; Schmid, R.; Duhrkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-Based Molecular Networking in the GNPS Analysis Environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Dictionary of Natural Products 28.1. Available online: http://dnp.chemnetbase.com/ (accessed on 9 January 2020).
- Tautenhahn, R.; Patti, G.J.; Rinehart, D.; Siuzdak, G. XCMS Online: A Web-Based Platform to Process Untargeted Metabolomic Data. Anal. Chem. 2012, 84, 5035–5039. [Google Scholar] [CrossRef] [Green Version]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Costa, R.C.; Santana, D.B.; Araujo, R.M.; de Paula, J.E.; do Nascimento, P.C.; Lopes, N.P.; Braz-Filho, R.; Espindola, L.S. Discovery of the Rapanone and Suberonone Mixture as a Motif for Leishmanicidal and Antifungal Applications. Bioorg. Med. Chem. 2014, 22, 135–140. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhao, X.E.; Zhu, S.; Dang, J.; Qiao, X.; Qiu, Z.; Tao, Y. Simultaneous Determination of Oleanolic Acid and Ursolic Acid by in Vivo Microdialysis via UHPLC-MS/MS Using Magnetic Dispersive Solid Phase Extraction Coupling with Microwave-Assisted Derivatization and Its Application to a Pharmacokinetic Study of Arctiumlappa, L. Root Extract in Rats. J. Agric. Food Chem. 2018, 66, 3975–3982. [Google Scholar] [CrossRef]
- Amara, S.; Zheng, M.; Tiriveedhi, V. Oleanolic Acid Inhibits High Salt-Induced Exaggeration of Warburg-like Metabolism in Breast Cancer Cells. Cell Biochem. Biophys 2016, 74, 427–434. [Google Scholar] [CrossRef] [Green Version]
- Ayeleso, T.B.; Matumba, M.G.; Mukwevho, E. Oleanolic Acid and Its Derivatives: Biological Activities and Therapeutic Potential in Chronic Diseases. Molecules 2017, 22, 1915. [Google Scholar] [CrossRef] [Green Version]
- Novotny, L.; Abdel-Hamid, M.E.; Hamza, H.; Masterova, I.; Grancai, D. Development of LC-MS Method for Determination of Ursolic Acid: Application to the Analysis of Ursolic Acid in Staphylea holocarpa Hemsl. J. Pharm. Biomed. Anal. 2003, 31, 961–968. [Google Scholar] [CrossRef]
- Kahnt, M.; Heller, L.; Grabandt, P.; Al-Harrasi, A.; Csuk, R. Platanic acid: A New Scaffold for the Synthesis of Cytotoxic Agents. Eur. J. Med. Chem. 2018, 143, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, I.; Peixoto, J.L.B.; Silva, C.C.D.; Shuquel, I.T.A.; Basso, E.A.; Vidotti, G.J. Triterpenic Acids from Eugenia Moraviana. J. Braz. Chem. Soc. 2001, 12, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, T.; Kashiwada, Y.; Kilkuskie, R.E.; Cosentino, L.M.; Ballas, L.M.; Jiang, J.B.; Janzen, W.P.; Chen, I.S.; Lee, K.H. Anti-AIDS Agents, 11. Betulinic Acid and Platanic Acid as Anti-HIV Principles from Syzigium Claviflorum, and the Anti-HIV Activity of Structurally Related Triterpenoids. J. Nat. Prod. 1994, 57, 243–247. [Google Scholar] [CrossRef]
- Dhar Dubey, K.K.; Sharma, G.; Kumar, A. Conjugated Linolenic Acids: Implication in Cancer. J. Agric. Food Chem. 2019, 67, 6091–6101. [Google Scholar] [CrossRef]
- Giordano, C.; Plastina, P.; Barone, I.; Catalano, S.; Bonofiglio, D. N-3 Polyunsaturated Fatty Acid Amides: New Avenues in the Prevention and Treatment of Breast Cancer. Int. J. Mol. Sci. 2020, 21, 2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okubo, R.; Noguchi, H.; Hamazaki, K.; Sekiguchi, M.; Kinoshita, T.; Katsumata, N.; Narisawa, T.; Uezono, Y.; Xiao, J.; Matsuoka, Y.J. Fear of Cancer Recurrence Among Breast Cancer Survivors Could be Controlled by Prudent Dietary Modification with Polyunsaturated Fatty Acids. J. Affect. Disord 2019, 245, 1114–1118. [Google Scholar] [CrossRef] [PubMed]
- Espindola, L.S.; Dusi, R.G.; Demarque, D.P.; Braz, R.; Yan, P.C.; Bokesch, H.R.; Gustafson, K.R.; Beutler, J.A. Cytotoxic Triterpenes from Salacia Crassifolia and Metabolite Profiling of Celastraceae Species. Molecules 2018, 23, 1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Liu, Y.; Zhong, J.; Bi, Y.; Liu, Y.; Ren, Z.; Li, X.; Jia, J.; Yu, M.; Yu, X. Pristimerin Induces Apoptosis and Autophagy via Activation of ROS/ASK1/JNK Pathway in Human Breast Cancer in Vitro and in Vivo. Cell Death Discov. 2019, 5, 125. [Google Scholar] [CrossRef] [Green Version]
- Kucera, O.; Mezera, V.; Moravcova, A.; Endlicher, R.; Lotkova, H.; Drahota, Z.; Cervinkova, Z. In Vitro Toxicity of Epigallocatechin Gallate in Rat Liver Mitochondria and Hepatocytes. Oxid Med. Cell Longev. 2015, 2015, 476180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurbitz, C.; Heise, D.; Redmer, T.; Goumas, F.; Arlt, A.; Lemke, J.; Rimbach, G.; Kalthoff, H.; Trauzold, A. Epicatechin Gallate and Catechin Gallate are Superior to Epigallocatechin Gallate in Growth Suppression and Anti-Inflammatory Activities in Pancreatic Tumor Cells. Cancer Sci. 2011, 102, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Bader, A.; Braca, A.; De Tommasi, N.; Morelli, I. Further Constituents from Caralluma Negevensis. Phytochemistry 2003, 62, 1277–1281. [Google Scholar] [CrossRef]
- Wishart, D.S. Is Cancer a Genetic Disease or a Metabolic Disease? EBioMedicine 2015, 2, 478–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albernaz, L.C.; de Paula, J.E.; Romero, G.A.; Silva Mdo, R.; Grellier, P.; Mambu, L.; Espindola, L.S. Investigation of Plant Extracts in Traditional Medicine of the Brazilian Cerrado Against Protozoans and Yeasts. J. Ethnopharmacol 2010, 131, 116–121. [Google Scholar] [CrossRef]
- De Mesquita, M.L.; Leao, W.F.; Ferreira, M.R.; de Paula, J.E.; Espindola, L.S.; Soares, L.A. Reversed-Phase-Liquid Chromatography Method for Separation and Quantification of Gallic Acid from Hydroalcoholic Extracts of Qualea Grandiflora and Qualea Parviflora. Pharmacogn. Mag. 2015, 11, 316–321. [Google Scholar] [CrossRef] [Green Version]
- De Mesquita, M.L.; de Paula, J.E.; Pessoa, C.; de Moraes, M.O.; Costa-Lotufo, L.V.; Grougnet, R.; Michel, S.; Tillequin, F.; Espindola, L.S. Cytotoxic Activity of Brazilian Cerrado Plants Used in Traditional Medicine Against Cancer Cell Lines. J. Ethnopharmaco. 2009, 123, 439–445. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Plant Species/Plant Organ (Solvent)* | Codes BR/NCI60 | IC50 (μg/mL) | NCI-60 Sensitivity Potency (μg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Colon | Renal | Osteosarcoma | ||||||||
| Colo205 | KM12 | A498 | U031 | MG63 | MG 63.3 | Mean GI50/Most Sensitive Cell Line | Mean TGI | Mean LC50 | ||
| Peschiera affinis SB (h) | BR 075 | 17.8 | 19.1 | >20 | >20 | >20 | >20 | - | - | - |
| Cybistax antisyphilitica SB (h) | BR 125 N192795 | - | - | - | - | - | - | 85 | >100 | >100 |
| Tabebuia caraiba L (h) | BR 139 | >20 | 18.9 | >20 | >20 | >20 | >20 | - | - | - |
| Casearia sylvestris var. lingua SW (h) | BR 177 N192825 | 4.6 | 4.5 | 3.6 | 2.7 | 3.9 | 5.7 | 11/2.5 NCI-H522 | 28 | 76 |
| Cupania vernalis L (h) | BR 193 N192827 | 12.3 | 10 | 18.7 | 7.6 | 13.8 | 15 | 43 | 140 | 490 |
| Magonia pubescens R (e) | BR 197 | 4.2 | 7.2 | 3.6 | 3.9 | 14.5 | 14.5 | - | - | - |
| Magonia pubescens RW (e) | BR 204 N192797 | 3.5 | 5.5 | 2.9 | 3.8 | 15.4 | 15.6 | 15/0.3SR | 34 | 81 |
| Simarouba versicolor RB (e) | BR 254 N192829 | 6.8 | >20 | 3.7 | 5 | <1.3 | <1.3 | 4.9/0.7 NCI-H522 | 29 | 620 |
| Simaba suffruticosa L (a) | BR 261 | 16.8 | 1.6 | >20 | 17.6 | 2.2 | 4.9 | - | - | - |
| Kielmeyera coriacea SW (h) | BR 331 N192831 | 11.9 | 4.9 | 15.9 | 14.9 | >20 | >20 | 56 | 210 | 600 |
| Byrsonima crassa RB (h) | BR 411 N192833 | 15 | 15 | >20 | >20 | >20 | >20 | 220 | 530 | 910 |
| Schinus terebinthifolia L (d) | BR 436 N192835 | 12.4 | 6.9 | 16.2 | 7.8 | 3.4 | 14.8 | 100/0.9 NCI-H522 | 87 | 360 |
| Enterolobium gummiferum RW (e) | BR 467 | 10.5 | 10.9 | <1.3 | 4.6 | 12.6 | 14.9 | - | - | - |
| Enterolobium gummiferum SB (h) | BR 469 N192837 | 2.2 | 4.9 | >20 | >20 | >20 | >20 | 180 | 710 | 980 |
| Plathymenia reticulata RW (h) | BR 489 N192839 | 8.9 | 6.7 | 12.4 | 4.9 | >20 | >20 | 60 | 330 | 680 |
| Diospyros hispida R (a) | BR 501 N192799 | - | - | - | - | - | - | 7/0.7 NCI-H522 | 30 | 81 |
| Maprounea guianensis RB (a) | BR 536 | 8.1 | 9.9 | 6.7 | >20 | 9.6 | 8.5 | - | - | - |
| Psidium laruotteanum SB (h) | BR 549 N192841 | - | - | - | - | - | - | 87 | 340 | 830 |
| Andira humilis SB (e) | BR 587 | >20 | 18.6 | >20 | >20 | >20 | >20 | - | - | - |
| Rapanea guianensis SW (h) | BR 624 | 5.3 | 3.2 | 4.3 | 4.4 | 17.8 | 14.9 | - | - | - |
| Rapanea guianensis RW (e) | BR 627 N192801 | 3.5 | 4.0 | 2.7 | 3.7 | 14.6 | 14 | 16/2.4 SR | 361 | 81 |
| Salacia crassifolia RW (h) | BR 640 N192803 | >20 | 1.7 | 1.6 | >20 | 3.7 | 5.7 | 0.3/0.1 HCT-15 | 1.0 | 5.0 |
| Salacia elliptica RW (a) | BR 652 N192805 | 3.6 | 3.4 | 3.5 | 2.0 | 5.1 | 7.1 | 2.0/0.7 MOLT-4 | 5.1 | 20 |
| Lippia rotundifolia SW (a) | BR 660 N192843 | 12.3 | 9.9 | >20 | >20 | >20 | >20 | - | - | - |
| Connarus suberosus RW (a) | BR 693 N192845 | 5.7 | 4.5 | 4.7 | 7.7 | 17.6 | 19 | 44 | 200 | 830 |
| Compound Name | Molecular Formula | m/z | Rt (min) |
|---|---|---|---|
| oleanolic acid* | C30H48O3 | 439.3567 | 9.7 |
| ursolic acid* | C30H48O3 | 439.3567 | 9.7 |
| 3α-cyclopenta[α]chrysene-3α-carboxylic acid | C29H46O4 | 441.3367 | 7.5 |
| platanic acid | C29H46O4 | 459.346 | 7.6 |
| linoleic acid | C18H32O2 | 281.2476 | 8.4 |
| linoleic acid ethyl ester | C20H36O3 | 309.2787 | 10.0 |
| linolenic acid ethyl ester | C20H34O2 | 307.2635 | 9.6 |
| 13-keto-9Z,11E-octadecadienoic acid | C18H30O3 | 295.2272 | 8.4 |
| 9-oxo-10E,12Z-octadecadienoic acid | C18H30O3 | 295.2269 | 8.5 |
| stearidonic acid | C18H28O2 | 277.2166 | 8.3 |
| 9(10)-epoxy-12Z-octadecenoic acid | C18H32O3 | 279.2322 | 8.1 |
| 9S,13R-12-oxophytodienoic acid | C18H28O3 | 293.2098 | 5.3 |
| pristimerin | C30H40O4 | 465.3012 | 10.3 |
| tingenone | C28H36O3 | 421.2745 | 8.6 |
| 20-oxo-20,21-seco-tingen-21-oic acid | C28H36O5 | 453.263 | 5.9 |
| (-)-catechin gallate | C22H18O10 | 443.0972 | 3.7 |
| epigallocatechin gallate | C22H17O11 | 459.0944 | 3.2 |
| luteolin 3′,4′-di-O-beta-D-glucopyranoside | C27H29O16 | 628.1959 | 10.7 |
| 13-docosenamide | C22H43NO3 | 338.342 | 11.7 |
| 9-octadecenamide | C18H35NO | 282.2793 | 10.1 |
| N-phenyl-1-naphthylamine | C16H13N | 220.1124 | 8.2 |
| (2R,3S,4S,5R,6S)-2-[[(2S,3R,4R)-3,4-dihydroxy-4-(hydroxymetil)oxolan-2-yl]oxymetil]-6-(3,4,5-trimethoxyphenoxy)oxane-3,4,5-triol | C20H30O13 | 496.1988 | 3.0 |
| (3R,5R,6R,7S,9S,10R,13R,17R)-17-((R)-5-ethoxy-5-oxypentan-2-yl)-10,13-dimethylhexadecahydro-1H-cyclopenta[a]phenanthreno-3,6,7-triyl triacetate | C32H50O | 585.3039 | 9.0 |
| 1-linoleoilglycerol | C16H36O3 | 355.282 | 7.9 |
| hesperidine | C28H34O15 | 611.498 | 4.1 |
| palmitamide | C20H41NO | 256.2639 | 10.0 |
| 4-(2,6,6-trimethyl-4-oxo-2-ciclohexen-1-yl)-2-butanyl beta-D-glucopyranoside | C19H32O7 | 373.2192 | 4.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortelo, P.C.; Demarque, D.P.; Dusi, R.G.; Albernaz, L.C.; Braz-Filho, R.; Goncharova, E.I.; Bokesch, H.R.; Gustafson, K.R.; Beutler, J.A.; Espindola, L.S. A Molecular Networking Strategy: High-Throughput Screening and Chemical Analysis of Brazilian Cerrado Plant Extracts against Cancer Cells. Cells 2021, 10, 691. https://doi.org/10.3390/cells10030691
Cortelo PC, Demarque DP, Dusi RG, Albernaz LC, Braz-Filho R, Goncharova EI, Bokesch HR, Gustafson KR, Beutler JA, Espindola LS. A Molecular Networking Strategy: High-Throughput Screening and Chemical Analysis of Brazilian Cerrado Plant Extracts against Cancer Cells. Cells. 2021; 10(3):691. https://doi.org/10.3390/cells10030691
Chicago/Turabian StyleCortelo, Patrícia C., Daniel P. Demarque, Renata G. Dusi, Lorena C. Albernaz, Raimundo Braz-Filho, Ekaterina I. Goncharova, Heidi R. Bokesch, Kirk R. Gustafson, John A. Beutler, and Laila S. Espindola. 2021. "A Molecular Networking Strategy: High-Throughput Screening and Chemical Analysis of Brazilian Cerrado Plant Extracts against Cancer Cells" Cells 10, no. 3: 691. https://doi.org/10.3390/cells10030691
APA StyleCortelo, P. C., Demarque, D. P., Dusi, R. G., Albernaz, L. C., Braz-Filho, R., Goncharova, E. I., Bokesch, H. R., Gustafson, K. R., Beutler, J. A., & Espindola, L. S. (2021). A Molecular Networking Strategy: High-Throughput Screening and Chemical Analysis of Brazilian Cerrado Plant Extracts against Cancer Cells. Cells, 10(3), 691. https://doi.org/10.3390/cells10030691