
Contemporary Mouse Models in Glioma Research

 ,
, 
 , ,
, ,
Abstract
:1. Introduction
2. Evolution of Cancer Mouse Models
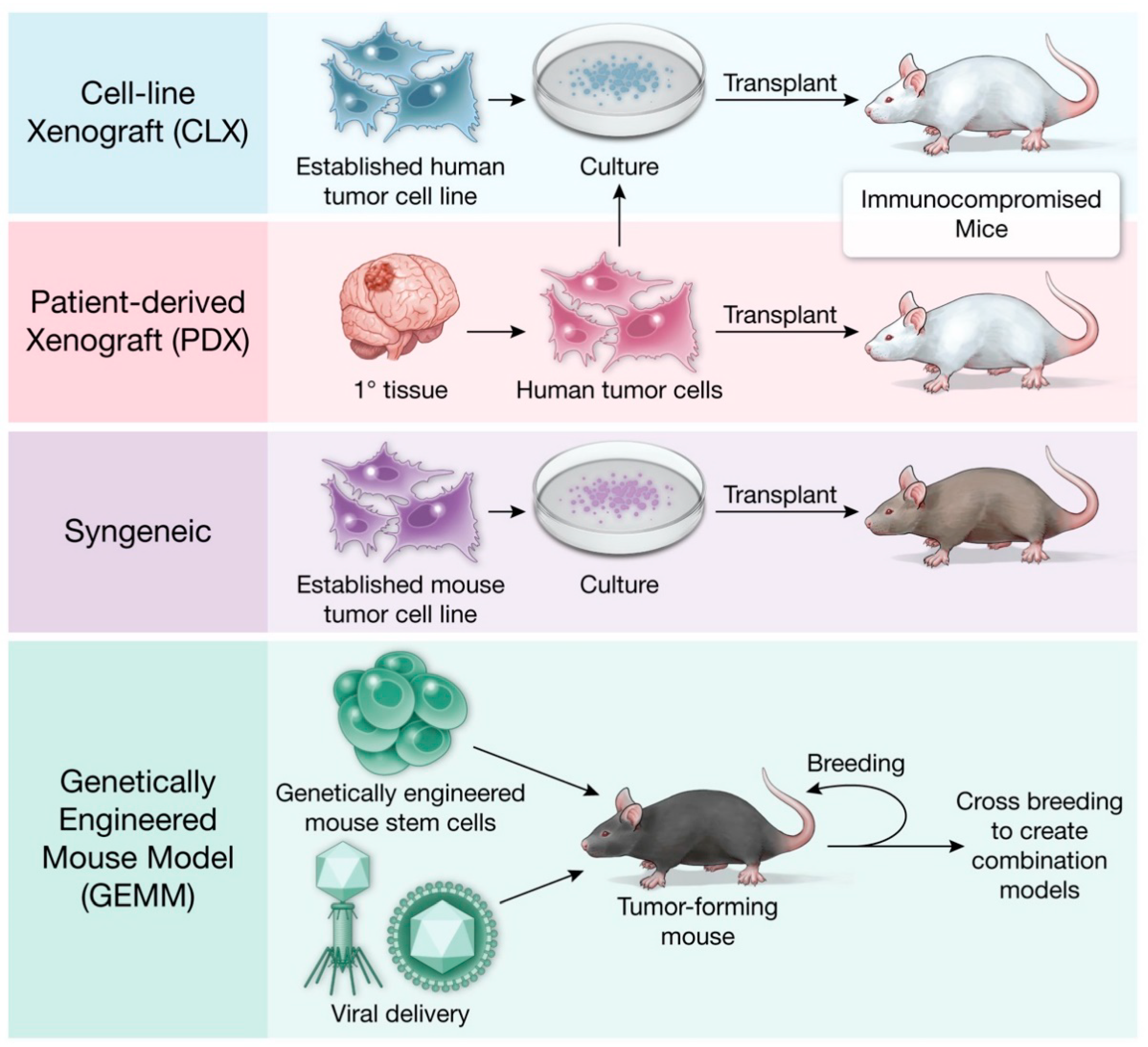
3. Xenograft Models
Immunology Research in Xenograft Models
4. Syngeneic Models
5. Genetically Engineered Mouse Models (GEMMs)
5.1. Somatic Gene Delivery Models
5.2. Conditional Allele-Specific Models
5.3. Transposon/Transposase Models
6. Special Consideration for IDH1/2 Mutations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Chinwalla, A.T.; Cook, L.L.; Delehaunty, K.D.; Fewell, G.A.; Fulton, L.A.; Fulton, R.S.; Graves, T.A.; Hillier, L.W.; Mardis, E.R.; McPherson, J.D.; et al. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- Noorani, I. Genetically Engineered Mouse Models of Gliomas: Technological Developments for Translational Discoveries. Cancers 2019, 11, 1335. [Google Scholar] [CrossRef] [Green Version]
- Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [CrossRef]
- Chen, R.; Smith-Cohn, M.; Cohen, A.L.; Colman, H. Glioma Subclassifications and Their Clinical Significance. Neurotherapeutics 2017, 14, 284–297. [Google Scholar] [CrossRef] [Green Version]
- Robertson, F.L.; Marques-Torrejon, M.A.; Morrison, G.M.; Pollard, S.M. Experimental models and tools to tackle glioblastoma. Dis. Model. Mech. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Stylli, S.S.; Luwor, R.B.; Ware, T.M.; Tan, F.; Kaye, A.H. Mouse models of glioma. J. Clin. Neurosci. 2015, 22, 619–626. [Google Scholar] [CrossRef]
- Imagi, S. Experimental study on heterotransplantation of Tanaka’s rabbit-hepatoma and intestinal cancer into guinea pigs. Gan 1950, 41, 231–233. [Google Scholar]
- Toolan, H.W. Growth of human tumors in cortisone-treated laboratory animals: The possibility of obtaining permanently transplantable human tumors. Cancer Res. 1953, 13, 389–394. [Google Scholar]
- Povlsen, C.O.; Spang-Thomsen, M.; Rygaard, J.; Visfeldt, J. Heterotransplantation of human malignant tumours to athymic nude mice. In Immunodeficient Animals for Cancer Research; Sparrow, S., Ed.; Palgrave Macmillan: London, UK, 1980; pp. 95–103. [Google Scholar] [CrossRef]
- Okada, S.; Vaeteewoottacharn, K.; Kariya, R. Application of Highly Immunocompromised Mice for the Establishment of Patient-Derived Xenograft (PDX) Models. Cells 2019, 8, 889. [Google Scholar] [CrossRef] [Green Version]
- Talmadge, J.E.; Singh, R.K.; Fidler, I.J.; Raz, A. Murine models to evaluate novel and conventional therapeutic strategies for cancer. Am. J. Pathol. 2007, 170, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Oh, T.; Fakurnejad, S.; Sayegh, E.T.; Clark, A.J.; Ivan, M.E.; Sun, M.Z.; Safaee, M.; Bloch, O.; James, C.D.; Parsa, A.T. Immunocompetent murine models for the study of glioblastoma immunotherapy. J. Transl. Med. 2014, 12, 107. [Google Scholar] [CrossRef] [Green Version]
- Chulpanova, D.S.; Kitaeva, K.V.; Rutland, C.S.; Rizvanov, A.A.; Solovyeva, V.V. Mouse Tumor Models for Advanced Cancer Immunotherapy. Int. J. Mol. Sci. 2020, 21, 4118. [Google Scholar] [CrossRef]
- Smith, H.W.; Muller, W.J. Transgenic mouse models—A seminal breakthrough in oncogene research. Cold Spring Harb. Protoc. 2013, 2013, 1099–1108. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Wagner, E.F.; Palmiter, R.D. The origins of oncomice: A history of the first transgenic mice genetically engineered to develop cancer. Genes Dev. 2007, 21, 2258–2270. [Google Scholar] [CrossRef] [Green Version]
- Adams, J.M.; Harris, A.W.; Pinkert, C.A.; Corcoran, L.M.; Alexander, W.S.; Cory, S.; Palmiter, R.D.; Brinster, R.L. The c-myc oncogene driven by immunoglobulin enhancers induces lymphoid malignancy in transgenic mice. Nature 1985, 318, 533–538. [Google Scholar] [CrossRef]
- Brinster, R.L.; Chen, H.Y.; Messing, A.; van Dyke, T.; Levine, A.J.; Palmiter, R.D. Transgenic mice harboring SV40 T-antigen genes develop characteristic brain tumors. Cell 1984, 37, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Stewart, T.A.; Pattengale, P.K.; Leder, P. Spontaneous mammary adenocarcinomas in transgenic mice that carry and express MTV/myc fusion genes. Cell 1984, 38, 627–637. [Google Scholar] [CrossRef]
- Gu, H.; Zou, Y.R.; Rajewsky, K. Independent control of immunoglobulin switch recombination at individual switch regions evidenced through Cre-loxP-mediated gene targeting. Cell 1993, 73, 1155–1164. [Google Scholar] [CrossRef]
- Jung, S.; Rajewsky, K.; Radbruch, A. Shutdown of class switch recombination by deletion of a switch region control element. Science 1993, 259, 984–987. [Google Scholar] [CrossRef]
- Akter, F.; Simon, B.; de Boer, N.L.; Redjal, N.; Wakimoto, H.; Shah, K. Pre-clinical tumor models of primary brain tumors: Challenges and opportunities. Biochim. Et Biophys. Acta. Rev. Cancer 2021, 1875, 188458. [Google Scholar] [CrossRef]
- Brighi, C.; Reid, L.; Genovesi, L.A.; Kojic, M.; Millar, A.; Bruce, Z.; White, A.L.; Day, B.W.; Rose, S.; Whittaker, A.K.; et al. Comparative study of preclinical mouse models of high-grade glioma for nanomedicine research: The importance of reproducing blood-brain barrier heterogeneity. Theranostics 2020, 10, 6361–6371. [Google Scholar] [CrossRef]
- Frese, K.K.; Tuveson, D.A. Maximizing mouse cancer models. Nat. Rev. Cancer 2007, 7, 645–658. [Google Scholar] [CrossRef]
- Yi, Y.; Hsieh, I.Y.; Huang, X.; Li, J.; Zhao, W. Glioblastoma Stem-Like Cells: Characteristics, Microenvironment, and Therapy. Front. Pharm. 2016, 7, 477. [Google Scholar] [CrossRef] [Green Version]
- Adhikari, B.; Li, J.; Brandel, M.G.; Futalan, D.; Akers, J.; Deming, T.; Chen, C.C.; Carter, B.S. The use of TMZ embedded hydrogels for the treatment of orthotopic human glioma xenografts. J. Clin. Neurosci. 2017, 45, 288–292. [Google Scholar] [CrossRef]
- Kaye, A.H.; Morstyn, G.; Gardner, I.; Pyke, K. Development of a xenograft glioma model in mouse brain. Cancer Res. 1986, 46, 1367–1373. [Google Scholar] [PubMed]
- Irtenkauf, S.M.; Sobiechowski, S.; Hasselbach, L.A.; Nelson, K.K.; Transou, A.D.; Carlton, E.T.; Mikkelsen, T.; deCarvalho, A.C. Optimization of Glioblastoma Mouse Orthotopic Xenograft Models for Translational Research. Comp. Med. 2017, 67, 300–314. [Google Scholar]
- Miner, M.W.; Liljenbäck, H.; Virta, J.; Merisaari, J.; Oikonen, V.; Westermarck, J.; Li, X.G.; Roivainen, A. (2S, 4R)-4-[(18)F]Fluoroglutamine for In vivo PET Imaging of Glioma Xenografts in Mice: An Evaluation of Multiple Pharmacokinetic Models. Mol. Imaging Biol. 2020, 22, 969–978. [Google Scholar] [CrossRef] [Green Version]
- Pellegatta, S.; Poliani, P.L.; Corno, D.; Menghi, F.; Ghielmetti, F.; Suarez-Merino, B.; Caldera, V.; Nava, S.; Ravanini, M.; Facchetti, F.; et al. Neurospheres enriched in cancer stem-like cells are highly effective in eliciting a dendritic cell-mediated immune response against malignant gliomas. Cancer Res. 2006, 66, 10247–10252. [Google Scholar] [CrossRef] [Green Version]
- Pellegatta, S.; Finocchiaro, G. Dendritic cell vaccines for cancer stem cells. Methods Mol. Biol. 2009, 568, 233–247. [Google Scholar] [CrossRef]
- Wagemakers, M.; van der Wal, G.E.; Cuberes, R.; Alvarez, I.; Andrés, E.M.; Buxens, J.; Vela, J.M.; Moorlag, H.; Mooij, J.J.; Molema, G. COX-2 Inhibition Combined with Radiation Reduces Orthotopic Glioma Outgrowth by Targeting the Tumor Vasculature. Transl. Oncol. 2009, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markovic, D.S.; Vinnakota, K.; van Rooijen, N.; Kiwit, J.; Synowitz, M.; Glass, R.; Kettenmann, H. Minocycline reduces glioma expansion and invasion by attenuating microglial MT1-MMP expression. Brain Behav. Immun. 2011, 25, 624–628. [Google Scholar] [CrossRef]
- Daga, A.; Orengo, A.M.; Gangemi, R.M.; Marubbi, D.; Perera, M.; Comes, A.; Ferrini, S.; Corte, G. Glioma immunotherapy by IL-21 gene-modified cells or by recombinant IL-21 involves antibody responses. Int. J. Cancer 2007, 121, 1756–1763. [Google Scholar] [CrossRef]
- Cantini, G.; Pisati, F.; Mastropietro, A.; Frattini, V.; Iwakura, Y.; Finocchiaro, G.; Pellegatta, S. A critical role for regulatory T cells in driving cytokine profiles of Th17 cells and their modulation of glioma microenvironment. Cancer Immunol. Immunother. 2011, 60, 1739–1750. [Google Scholar] [CrossRef] [Green Version]
- Yamazoe, T.; Koizumi, S.; Yamasaki, T.; Amano, S.; Tokuyama, T.; Namba, H. Potent tumor tropism of induced pluripotent stem cells and induced pluripotent stem cell-derived neural stem cells in the mouse intracerebral glioma model. Int. J. Oncol. 2015, 46, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Morante, D.; Cortes-Canteli, M.; Sanz-Sancristobal, M.; Santos, A.; Perez-Castillo, A. Decreased CCAAT/enhancer binding protein β expression inhibits the growth of glioblastoma cells. Neuroscience 2011, 176, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Hülper, P.; Schulz-Schaeffer, W.; Dullin, C.; Hoffmann, P.; Harper, J.; Kurtzberg, L.; Lonning, S.; Kugler, W.; Lakomek, M.; Erdlenbruch, B. Tumor localization of an anti-TGF-β antibody and its effects on gliomas. Int. J. Oncol. 2011, 38, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Madala, H.R.; Punganuru, S.R.; Ali-Osman, F.; Zhang, R.; Srivenugopal, K.S. Brain- and brain tumor-penetrating disulfiram nanoparticles: Sequence of cytotoxic events and efficacy in human glioma cell lines and intracranial xenografts. Oncotarget 2018, 9, 3459–3482. [Google Scholar] [CrossRef] [Green Version]
- Larsson, S.; Wenger, A.; Dósa, S.; Sabel, M.; Kling, T.; Carén, H. Cell line-based xenograft mouse model of paediatric glioma stem cells mirrors the clinical course of the patient. Carcinogenesis 2018, 39, 1304–1309. [Google Scholar] [CrossRef]
- Weissenberger, J.; Priester, M.; Bernreuther, C.; Rakel, S.; Glatzel, M.; Seifert, V.; Kögel, D. Dietary curcumin attenuates glioma growth in a syngeneic mouse model by inhibition of the JAK1,2/STAT3 signaling pathway. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 5781–5795. [Google Scholar] [CrossRef] [Green Version]
- Kiviniemi, A.; Gardberg, M.; Autio, A.; Li, X.G.; Heuser, V.D.; Liljenback, H.; Kakela, M.; Sipila, H.; Kurkipuro, J.; Yla-Herttuala, S.; et al. Feasibility of experimental BT4C glioma models for somatostatin receptor 2-targeted therapies. Acta Oncol. 2014, 53, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhu, S.; Qian, L.; Pei, Y.; Qiu, Y.; Jiang, Y. RGD-modified PEG-PAMAM-DOX conjugates: In vitro and in vivo studies for glioma. Eur. J. Pharm. Biopharm. Off. J. Arb. Fur Pharm. Verfahr. EV 2011, 79, 232–240. [Google Scholar] [CrossRef]
- Luchman, H.A.; Stechishin, O.D.; Dang, N.H.; Blough, M.D.; Chesnelong, C.; Kelly, J.J.; Nguyen, S.A.; Chan, J.A.; Weljie, A.M.; Cairncross, J.G.; et al. An in vivo patient-derived model of endogenous IDH1-mutant glioma. Neuro Oncol. 2012, 14, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fareh, M.; Turchi, L.; Virolle, V.; Debruyne, D.; Almairac, F.; de-la-Forest Divonne, S.; Paquis, P.; Preynat-Seauve, O.; Krause, K.H.; Chneiweiss, H.; et al. The miR 302–367 cluster drastically affects self-renewal and infiltration properties of glioma-initiating cells through CXCR4 repression and consequent disruption of the SHH-GLI-NANOG network. Cell Death Differ. 2012, 19, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Jandial, R.; Neman, J.; Lim, P.P.; Tamae, D.; Kowolik, C.M.; Wuenschell, G.E.; Shuck, S.C.; Ciminera, A.K.; De Jesus, L.R.; Ouyang, C.; et al. Inhibition of GLO1 in Glioblastoma Multiforme Increases DNA-AGEs, Stimulates RAGE Expression, and Inhibits Brain Tumor Growth in Orthotopic Mouse Models. Int. J. Mol. Sci. 2018, 19, 406. [Google Scholar] [CrossRef] [Green Version]
- Hlavaty, J.; Jandl, G.; Liszt, M.; Petznek, H.; König-Schuster, M.; Sedlak, J.; Egerbacher, M.; Weissenberger, J.; Salmons, B.; Günzburg, W.H.; et al. Comparative evaluation of preclinical in vivo models for the assessment of replicating retroviral vectors for the treatment of glioblastoma. J. Neurooncol. 2011, 102, 59–69. [Google Scholar] [CrossRef]
- Ni, X.R.; Zhao, Y.Y.; Cai, H.P.; Yu, Z.H.; Wang, J.; Chen, F.R.; Yu, Y.J.; Feng, G.K.; Chen, Z.P. Transferrin receptor 1 targeted optical imaging for identifying glioma margin in mouse models. J. Neurooncol. 2020, 148, 245–258. [Google Scholar] [CrossRef]
- Beck, S.; Jin, X.; Yin, J.; Kim, S.H.; Lee, N.K.; Oh, S.Y.; Jin, X.; Kim, M.K.; Kim, E.B.; Son, J.S.; et al. Identification of a peptide that interacts with Nestin protein expressed in brain cancer stem cells. Biomaterials 2011, 32, 8518–8528. [Google Scholar] [CrossRef] [PubMed]
- Nicolaides, T.P.; Li, H.; Solomon, D.A.; Hariono, S.; Hashizume, R.; Barkovich, K.; Baker, S.J.; Paugh, B.S.; Jones, C.; Forshew, T.; et al. Targeted therapy for BRAFV600E malignant astrocytoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 7595–7604. [Google Scholar] [CrossRef] [Green Version]
- Santagata, S.; Xu, Y.M.; Wijeratne, E.M.; Kontnik, R.; Rooney, C.; Perley, C.C.; Kwon, H.; Clardy, J.; Kesari, S.; Whitesell, L.; et al. Using the heat-shock response to discover anticancer compounds that target protein homeostasis. ACS Chem. Biol. 2012, 7, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Moroz, M.A.; Huang, R.; Kochetkov, T.; Shi, W.; Thaler, H.; de Stanchina, E.; Gamez, I.; Ryan, R.P.; Blasberg, R.G. Comparison of corticotropin-releasing factor, dexamethasone, and temozolomide: Treatment efficacy and toxicity in U87 and C6 intracranial gliomas. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 3282–3292. [Google Scholar] [CrossRef] [Green Version]
- Burden-Gulley, S.M.; Qutaish, M.Q.; Sullivant, K.E.; Lu, H.; Wang, J.; Craig, S.E.; Basilion, J.P.; Wilson, D.L.; Brady-Kalnay, S.M. Novel cryo-imaging of the glioma tumor microenvironment reveals migration and dispersal pathways in vivid three-dimensional detail. Cancer Res. 2011, 71, 5932–5940. [Google Scholar] [CrossRef] [Green Version]
- Navis, A.C.; Hamans, B.C.; Claes, A.; Heerschap, A.; Jeuken, J.W.; Wesseling, P.; Leenders, W.P. Effects of targeting the VEGF and PDGF pathways in diffuse orthotopic glioma models. J. Pathol. 2011, 223, 626–634. [Google Scholar] [CrossRef]
- Bai, R.Y.; Staedtke, V.; Aprhys, C.M.; Gallia, G.L.; Riggins, G.J. Antiparasitic mebendazole shows survival benefit in 2 preclinical models of glioblastoma multiforme. Neuro Oncol. 2011, 13, 974–982. [Google Scholar] [CrossRef]
- Emdad, L.; Sarkar, D.; Lee, S.G.; Su, Z.Z.; Yoo, B.K.; Dash, R.; Yacoub, A.; Fuller, C.E.; Shah, K.; Dent, P.; et al. Astrocyte elevated gene-1: A novel target for human glioma therapy. Mol. Cancer Ther. 2010, 9, 79–88. [Google Scholar] [CrossRef] [Green Version]
- Jamal, M.; Rath, B.H.; Williams, E.S.; Camphausen, K.; Tofilon, P.J. Microenvironmental regulation of glioblastoma radioresponse. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 6049–6059. [Google Scholar] [CrossRef] [Green Version]
- Jamal, M.; Rath, B.H.; Tsang, P.S.; Camphausen, K.; Tofilon, P.J. The brain microenvironment preferentially enhances the radioresistance of CD133(+) glioblastoma stem-like cells. Neoplasia 2012, 14, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marian, C.O.; Cho, S.K.; McEllin, B.M.; Maher, E.A.; Hatanpaa, K.J.; Madden, C.J.; Mickey, B.E.; Wright, W.E.; Shay, J.W.; Bachoo, R.M. The telomerase antagonist, imetelstat, efficiently targets glioblastoma tumor-initiating cells leading to decreased proliferation and tumor growth. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Bruyère, C.; Abeloos, L.; Lamoral-Theys, D.; Senetta, R.; Mathieu, V.; Le Mercier, M.; Kast, R.E.; Cassoni, P.; Vandenbussche, G.; Kiss, R.; et al. Temozolomide modifies caveolin-1 expression in experimental malignant gliomas in vitro and in vivo. Transl. Oncol. 2011, 4, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Li, J.T.; Zheng, F.; Martin, E.; Kots, A.Y.; Krumenacker, J.S.; Choi, B.K.; McCutcheon, I.E.; Weisbrodt, N.; Bögler, O.; et al. Restoring soluble guanylyl cyclase expression and function blocks the aggressive course of glioma. Mol. Pharmacol. 2011, 80, 1076–1084. [Google Scholar] [CrossRef] [Green Version]
- Bruyère, C.; Madonna, S.; Van Goietsenoven, G.; Mathieu, V.; Dessolin, J.; Kraus, J.L.; Lefranc, F.; Kiss, R. JLK1486, a Bis 8-Hydroxyquinoline-Substituted Benzylamine, Displays Cytostatic Effects in Experimental Gliomas through MyT1 and STAT1 Activation and, to a Lesser Extent, PPARγ Activation. Transl. Oncol. 2011, 4, 126–137. [Google Scholar] [CrossRef]
- Qutaish, M.Q.; Sullivant, K.E.; Burden-Gulley, S.M.; Lu, H.; Roy, D.; Wang, J.; Basilion, J.P.; Brady-Kalnay, S.M.; Wilson, D.L. Cryo-image analysis of tumor cell migration, invasion, and dispersal in a mouse xenograft model of human glioblastoma multiforme. Mol. Imaging Biol. 2012, 14, 572–583. [Google Scholar] [CrossRef] [Green Version]
- Tabatabai, G.; Frank, B.; Wick, A.; Lemke, D.; von Kürthy, G.; Obermüller, U.; Heckl, S.; Christ, G.; Weller, M.; Wick, W. Synergistic antiglioma activity of radiotherapy and enzastaurin. Ann. Neurol. 2007, 61, 153–161. [Google Scholar] [CrossRef]
- Dong, J.; Zhou, G.; Tang, D.; Chen, Y.; Cui, B.; Dai, X.; Zhang, J.; Lan, Q.; Huang, Q. Local delivery of slow-releasing temozolomide microspheres inhibits intracranial xenograft glioma growth. J. Cancer Res. Clin. Oncol. 2012, 138, 2079–2084. [Google Scholar] [CrossRef]
- Le Calvé, B.; Rynkowski, M.; Le Mercier, M.; Bruyère, C.; Lonez, C.; Gras, T.; Haibe-Kains, B.; Bontempi, G.; Decaestecker, C.; Ruysschaert, J.M.; et al. Long-term in vitro treatment of human glioblastoma cells with temozolomide increases resistance in vivo through up-regulation of GLUT transporter and aldo-keto reductase enzyme AKR1C expression. Neoplasia 2010, 12, 727–739. [Google Scholar] [CrossRef]
- Najbauer, J.; Huszthy, P.C.; Barish, M.E.; Garcia, E.; Metz, M.Z.; Myers, S.M.; Gutova, M.; Frank, R.T.; Miletic, H.; Kendall, S.E.; et al. Cellular host responses to gliomas. PLoS ONE 2012, 7, e35150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staquicini, F.I.; Ozawa, M.G.; Moya, C.A.; Driessen, W.H.; Barbu, E.M.; Nishimori, H.; Soghomonyan, S.; Flores, L.G., 2nd; Liang, X.; Paolillo, V.; et al. Systemic combinatorial peptide selection yields a non-canonical iron-mimicry mechanism for targeting tumors in a mouse model of human glioblastoma. J. Clin. Investig. 2011, 121, 161–173. [Google Scholar] [CrossRef]
- Schauff, A.K.; Kim, E.L.; Leppert, J.; Nadrowitz, R.; Wuestenberg, R.; Brockmann, M.A.; Giese, A. Inhibition of invasion-associated thromboxane synthase sensitizes experimental gliomas to gamma-radiation. J. Neurooncol. 2009, 91, 241–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGee, M.C.; Hamner, J.B.; Williams, R.F.; Rosati, S.F.; Sims, T.L.; Ng, C.Y.; Gaber, M.W.; Calabrese, C.; Wu, J.; Nathwani, A.C.; et al. Improved intratumoral oxygenation through vascular normalization increases glioma sensitivity to ionizing radiation. Int. J. Radiat. Oncol. Biol. Phys. 2010, 76, 1537–1545. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Kim, K.H.; Lee, J.; Lee, Y.A.; Kim, M.; Lee, S.J.; Park, K.; Yang, H.; Jin, J.; Joo, K.M.; et al. Wnt activation is implicated in glioblastoma radioresistance. Lab. Investig. A J. Tech. Methods Pathol. 2012, 92, 466–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaponis, D.; Barnes, J.W.; Dellagatta, J.L.; Kesari, S.; Fast, E.; Sauvageot, C.; Panagrahy, D.; Greene, E.R.; Ramakrishna, N.; Wen, P.Y.; et al. Lonafarnib (SCH66336) improves the activity of temozolomide and radiation for orthotopic malignant gliomas. J. Neurooncol. 2011, 104, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.C.; Wang, W.; Golden, E.B.; Thomas, S.; Sivakumar, W.; Hofman, F.M.; Louie, S.G.; Schönthal, A.H. Green tea epigallocatechin gallate enhances therapeutic efficacy of temozolomide in orthotopic mouse glioblastoma models. Cancer Lett. 2011, 302, 100–108. [Google Scholar] [CrossRef]
- Matsumura, Y. Preclinical and clinical studies of NK012, an SN-38-incorporating polymeric micelles, which is designed based on EPR effect. Adv. Drug Deliv. Rev. 2011, 63, 184–192. [Google Scholar] [CrossRef]
- Redjal, N.; Chan, J.A.; Segal, R.A.; Kung, A.L. CXCR4 inhibition synergizes with cytotoxic chemotherapy in gliomas. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 6765–6771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuthalapati, S.; Zhou, Q.; Guo, P.; Lv, H.; Cosenza, S.; Reddy, M.V.; Reddy, E.P.; Gallo, J.M. Preclinical pharmacokinetic and pharmacodynamic evaluation of novel anticancer agents, ON01910.Na (Rigosertib, Estybon™) and ON013105, for brain tumor chemotherapy. Pharm. Res. 2012, 29, 2499–2511. [Google Scholar] [CrossRef]
- Lamfers, M.L.; Idema, S.; Bosscher, L.; Heukelom, S.; Moeniralm, S.; van der Meulen-Muileman, I.H.; Overmeer, R.M.; van der Valk, P.; van Beusechem, V.W.; Gerritsen, W.R.; et al. Differential effects of combined Ad5- delta 24RGD and radiation therapy in in vitro versus in vivo models of malignant glioma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 7451–7458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertrand, Y.; Currie, J.C.; Poirier, J.; Demeule, M.; Abulrob, A.; Fatehi, D.; Stanimirovic, D.; Sartelet, H.; Castaigne, J.P.; Béliveau, R. Influence of glioma tumour microenvironment on the transport of ANG1005 via low-density lipoprotein receptor-related protein 1. Br. J. Cancer 2011, 105, 1697–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, S.S.; Kamoun, W.S.; Farrar, C.T.; Kirkpatrick, N.D.; Niemeyer, E.; de Graaf, A.M.; Sorensen, A.G.; Munn, L.L.; Jain, R.K.; Fukumura, D. Angiopoietin-2 interferes with anti-VEGFR2-induced vessel normalization and survival benefit in mice bearing gliomas. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 3618–3627. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, B.; McEllin, B.; Camacho, C.V.; Tomimatsu, N.; Sirasanagandala, S.; Nannepaga, S.; Hatanpaa, K.J.; Mickey, B.; Madden, C.; Maher, E.; et al. EGFRvIII and DNA double-strand break repair: A molecular mechanism for radioresistance in glioblastoma. Cancer Res. 2009, 69, 4252–4259. [Google Scholar] [CrossRef] [Green Version]
- Momiyama, M.; Zhao, M.; Kimura, H.; Tran, B.; Chishima, T.; Bouvet, M.; Endo, I.; Hoffman, R.M. Inhibition and eradication of human glioma with tumor-targeting Salmonella typhimurium in an orthotopic nude-mouse model. Cell Cycle 2012, 11, 628–632. [Google Scholar] [CrossRef] [Green Version]
- Menon, L.G.; Pratt, J.; Yang, H.W.; Black, P.M.; Sorensen, G.A.; Carroll, R.S. Imaging of human mesenchymal stromal cells: Homing to human brain tumors. J. Neurooncol. 2012, 107, 257–267. [Google Scholar] [CrossRef]
- Ahmed, A.U.; Thaci, B.; Alexiades, N.G.; Han, Y.; Qian, S.; Liu, F.; Balyasnikova, I.V.; Ulasov, I.Y.; Aboody, K.S.; Lesniak, M.S. Neural stem cell-based cell carriers enhance therapeutic efficacy of an oncolytic adenovirus in an orthotopic mouse model of human glioblastoma. Mol. Ther. J. Am. Soc. Gene Ther. 2011, 19, 1714–1726. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Ong, L.C.; Ranganath, S.H.; Zheng, L.; Kee, I.; Zhan, W.; Yu, S.; Chow, P.K.; Wang, C.H. A Dual Tracer 18F-FCH/18F-FDG PET Imaging of an Orthotopic Brain Tumor Xenograft Model. PLoS ONE 2016, 11, e0148123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschner, S.; Felix, M.C.; Hartmann, L.; Bierbaum, M.; Maros, M.E.; Kerl, H.U.; Wenz, F.; Glatting, G.; Kramer, M.; Giordano, F.A.; et al. In vivo micro-CT imaging of untreated and irradiated orthotopic glioblastoma xenografts in mice: Capabilities, limitations and a comparison with bioluminescence imaging. J. Neurooncol. 2015, 122, 245–254. [Google Scholar] [CrossRef]
- Yin, J.; Kim, J.K.; Moon, J.H.; Beck, S.; Piao, D.; Jin, X.; Kim, S.H.; Lim, Y.C.; Nam, D.H.; You, S.; et al. hMSC-mediated concurrent delivery of endostatin and carboxylesterase to mouse xenografts suppresses glioma initiation and recurrence. Mol. Ther. J. Am. Soc. Gene Ther. 2011, 19, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- von Baumgarten, L.; Brucker, D.; Tirniceru, A.; Kienast, Y.; Grau, S.; Burgold, S.; Herms, J.; Winkler, F. Bevacizumab has differential and dose-dependent effects on glioma blood vessels and tumor cells. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 17, 6192–6205. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Yamauchi, K.; Yamamoto, N.; Tsuchiya, H.; Tomita, K.; Bouvet, M.; Wessels, J.; Hoffman, R.M. A color-coded orthotopic nude-mouse treatment model of brain-metastatic paralyzing spinal cord cancer that induces angiogenesis and neurogenesis. Cell Prolif. 2009, 42, 75–82. [Google Scholar] [CrossRef]
- Khan, N.; Mupparaju, S.; Hou, H.; Williams, B.B.; Swartz, H. Repeated assessment of orthotopic glioma pO(2) by multi-site EPR oximetry: A technique with the potential to guide therapeutic optimization by repeated measurements of oxygen. J. Neurosci. Methods 2012, 204, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Kil, W.J.; Cerna, D.; Burgan, W.E.; Beam, K.; Carter, D.; Steeg, P.S.; Tofilon, P.J.; Camphausen, K. In vitro and in vivo radiosensitization induced by the DNA methylating agent temozolomide. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2008, 14, 931–938. [Google Scholar] [CrossRef] [Green Version]
- Gwak, H.S.; Shingu, T.; Chumbalkar, V.; Hwang, Y.H.; DeJournett, R.; Latha, K.; Koul, D.; Alfred Yung, W.K.; Powis, G.; Farrell, N.P.; et al. Combined action of the dinuclear platinum compound BBR3610 with the PI3-K inhibitor PX-866 in glioblastoma. Int. J. Cancer 2011, 128, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.U.; Tyler, M.A.; Thaci, B.; Alexiades, N.G.; Han, Y.; Ulasov, I.V.; Lesniak, M.S. A comparative study of neural and mesenchymal stem cell-based carriers for oncolytic adenovirus in a model of malignant glioma. Mol. Pharm. 2011, 8, 1559–1572. [Google Scholar] [CrossRef] [Green Version]
- Verhoeff, J.J.; Stalpers, L.J.; Claes, A.; Hovinga, K.E.; Musters, G.D.; Peter Vandertop, W.; Richel, D.J.; Leenders, W.P.; van Furth, W.R. Tumour control by whole brain irradiation of anti-VEGF-treated mice bearing intracerebral glioma. Eur. J. Cancer 2009, 45, 3074–3080. [Google Scholar] [CrossRef]
- Verhoeff, J.J.; Stalpers, L.J.; Coumou, A.W.; Koedooder, K.; Lavini, C.; Van Noorden, C.J.; Haveman, J.; Vandertop, W.P.; van Furth, W.R. Experimental iodine-125 seed irradiation of intracerebral brain tumors in nude mice. Radiat. Oncol. 2007, 2, 38. [Google Scholar] [CrossRef] [Green Version]
- Han, J.H.; Yoon, J.S.; Chang, D.Y.; Cho, K.G.; Lim, J.; Kim, S.S.; Suh-Kim, H. CXCR4-STAT3 Axis Plays a Role in Tumor Cell Infiltration in an Orthotopic Mouse Glioblastoma Model. Mol. Cells 2020, 43, 539–550. [Google Scholar] [CrossRef]
- Schötterl, S.; Miemietz, J.T.; Ilina, E.I.; Wirsik, N.M.; Ehrlich, I.; Gall, A.; Huber, S.M.; Lentzen, H.; Mittelbronn, M.; Naumann, U. Mistletoe-Based Drugs Work in Synergy with Radio-Chemotherapy in the Treatment of Glioma In Vitro and In Vivo in Glioblastoma Bearing Mice. Evid. Based Complementary Altern. Med. Ecam 2019, 2019, 1376140. [Google Scholar] [CrossRef] [Green Version]
- Verhoye, M.; van der Sanden, B.P.; Rijken, P.F.; Peters, H.P.; Van der Kogel, A.J.; Pée, G.; Vanhoutte, G.; Heerschap, A.; Van der Linden, A. Assessment of the neovascular permeability in glioma xenografts by dynamic T(1) MRI with Gadomer-17. Magn. Reson. Med. 2002, 47, 305–313. [Google Scholar] [CrossRef]
- Wang, S.; Khan, A.; Lang, F.F.; Schaefer, T.S. Conditional gene expression in human intracranial xenograft tumors. BioTechniques 2001, 31, 196–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemke, D.; Pfenning, P.N.; Sahm, F.; Klein, A.C.; Kempf, T.; Warnken, U.; Schnölzer, M.; Tudoran, R.; Weller, M.; Platten, M.; et al. Costimulatory protein 4IgB7H3 drives the malignant phenotype of glioblastoma by mediating immune escape and invasiveness. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Camphausen, K.; Purow, B.; Sproull, M.; Scott, T.; Ozawa, T.; Deen, D.F.; Tofilon, P.J. Orthotopic growth of human glioma cells quantitatively and qualitatively influences radiation-induced changes in gene expression. Cancer Res. 2005, 65, 10389–10393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.F.; Sun, X.K.; Lan, Y.; Han, C.; Zhang, Y.D.; Chen, G. Linarin sensitizes tumor necrosis factor-related apoptosis (TRAIL)-induced ligand-triggered apoptosis in human glioma cells and in xenograft nude mice. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 95, 1607–1618. [Google Scholar] [CrossRef]
- Park, C.R.; Kim, H.Y.; Song, M.G.; Lee, Y.S.; Youn, H.; Chung, J.K.; Cheon, G.J.; Kang, K.W. Efficacy and Safety of Human Serum Albumin-Cisplatin Complex in U87MG Xenograft Mouse Models. Int. J. Mol. Sci. 2020, 21, 7932. [Google Scholar] [CrossRef]
- Palma, C.; Bigioni, M.; Irrissuto, C.; Nardelli, F.; Maggi, C.A.; Manzini, S. Anti-tumour activity of tachykinin NK1 receptor antagonists on human glioma U373 MG xenograft. Br. J. Cancer 2000, 82, 480–487. [Google Scholar] [CrossRef] [Green Version]
- Bagci-Onder, T.; Wakimoto, H.; Anderegg, M.; Cameron, C.; Shah, K. A dual PI3K/mTOR inhibitor, PI-103, cooperates with stem cell-delivered TRAIL in experimental glioma models. Cancer Res. 2011, 71, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Yuan, A.L.; Ricks, C.B.; Bohm, A.K.; Lun, X.; Maxwell, L.; Safdar, S.; Bukhari, S.; Gerber, A.; Sayeed, W.; Bering, E.A.; et al. ABT-888 restores sensitivity in temozolomide resistant glioma cells and xenografts. PLoS ONE 2018, 13, e0202860. [Google Scholar] [CrossRef] [Green Version]
- Wakimoto, H.; Mohapatra, G.; Kanai, R.; Curry, W.T., Jr.; Yip, S.; Nitta, M.; Patel, A.P.; Barnard, Z.R.; Stemmer-Rachamimov, A.O.; Louis, D.N.; et al. Maintenance of primary tumor phenotype and genotype in glioblastoma stem cells. Neuro Oncol. 2012, 14, 132–144. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Luk, S.; Kiyokawa, J.; Onozato, M.L.; Iafrate, A.J.; Shah, K.; Martuza, R.L.; Rabkin, S.D.; Batchelor, T.T.; Cahill, D.P.; et al. Genetically distinct glioma stem-like cell xenografts established from paired glioblastoma samples harvested before and after molecularly targeted therapy. Sci. Rep. 2019, 9, 139. [Google Scholar] [CrossRef]
- Klink, B.; Miletic, H.; Stieber, D.; Huszthy, P.C.; Valenzuela, J.A.C.; Balss, J.; Wang, J.; Schubert, M.; Sakariassen, P.Ø.; Sundstrøm, T.; et al. A Novel, Diffusely Infiltrative Xenograft Model of Human Anaplastic Oligodendroglioma with Mutations in FUBP1, CIC, and IDH1. PLoS ONE 2013, 8, e59773. [Google Scholar] [CrossRef]
- Zeng, W.; Tang, Z.; Li, Y.; Yin, G.; Liu, Z.; Gao, J.; Chen, Y.; Chen, F. Patient-derived xenografts of different grade gliomas retain the heterogeneous histological and genetic features of human gliomas. Cancer Cell Int. 2020, 20, 1. [Google Scholar] [CrossRef]
- Ding, H.; Roncari, L.; Shannon, P.; Wu, X.; Lau, N.; Karaskova, J.; Gutmann, D.H.; Squire, J.A.; Nagy, A.; Guha, A. Astrocyte-specific expression of activated p21-ras results in malignant astrocytoma formation in a transgenic mouse model of human gliomas. Cancer Res. 2001, 61, 3826–3836. [Google Scholar] [PubMed]
- Xue, W.; Ton, H.; Zhang, J.; Xie, T.; Chen, X.; Zhou, B.; Guo, Y.; Fang, J.; Wang, S.; Zhang, W. Patient-derived orthotopic xenograft glioma models fail to replicate the magnetic resonance imaging features of the original patient tumor. Oncol. Rep. 2020, 43, 1619–1629. [Google Scholar] [CrossRef]
- Joo, K.M.; Kim, J.; Jin, J.; Kim, M.; Seol, H.J.; Muradov, J.; Yang, H.; Choi, Y.L.; Park, W.Y.; Kong, D.S.; et al. Patient-specific orthotopic glioblastoma xenograft models recapitulate the histopathology and biology of human glioblastomas in situ. Cell Rep. 2013, 3, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Ishiuchi, S.; Kanda, T.; Hasegawa, M.; Nakazato, Y. Analysis of interleukin-6 gene expression in primary human gliomas, glioblastoma xenografts, and glioblastoma cell lines. Brain Tumor Pathol. 2001, 18, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter-Fogle, A.E.; Harris, P.L.R.; Brady-Kalnay, S.M.; Sloan, A.E. Generation of Glioblastoma Patient-Derived Intracranial Xenografts for Preclinical Studies. Int. J. Mol. Sci. 2020, 21, 5113. [Google Scholar] [CrossRef]
- Yamashita, A.S.; da Costa Rosa, M.; Borodovsky, A.; Festuccia, W.T.; Chan, T.; Riggins, G.J. Demethylation and epigenetic modification with 5-azacytidine reduces IDH1 mutant glioma growth in combination with temozolomide. Neuro Oncol. 2019, 21, 189–200. [Google Scholar] [CrossRef]
- Sarkaria, J.N.; Carlson, B.L.; Schroeder, M.A.; Grogan, P.; Brown, P.D.; Giannini, C.; Ballman, K.V.; Kitange, G.J.; Guha, A.; Pandita, A.; et al. Use of an orthotopic xenograft model for assessing the effect of epidermal growth factor receptor amplification on glioblastoma radiation response. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2006, 12, 2264–2271. [Google Scholar] [CrossRef] [Green Version]
- Kitange, G.J.; Carlson, B.L.; Mladek, A.C.; Decker, P.A.; Schroeder, M.A.; Wu, W.; Grogan, P.T.; Giannini, C.; Ballman, K.V.; Buckner, J.C.; et al. Evaluation of MGMT promoter methylation status and correlation with temozolomide response in orthotopic glioblastoma xenograft model. J. Neurooncol. 2009, 92, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Scigliano, S.; Pinel, S.; Poussier, S.; Fouyssac, F.; Plenat, F.; Karcher, G.; Chastagner, P. Measurement of hypoxia using invasive oxygen-sensitive electrode, pimonidazole binding and 18F-FDG uptake in anaemic or erythropoietin-treated mice bearing human glioma xenografts. Int. J. Oncol. 2008, 32, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Wakimoto, H.; Tanaka, S.; Curry, W.T.; Loebel, F.; Zhao, D.; Tateishi, K.; Chen, J.; Klofas, L.K.; Lelic, N.; Kim, J.C.; et al. Targetable signaling pathway mutations are associated with malignant phenotype in IDH-mutant gliomas. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2014, 20, 2898–2909. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, V.; Mashimo, T.; An, Z.; Vemireddy, V.; Piccirillo, S.; Askari, P.; Hulsey, K.M.; Zhang, S.; de Graaf, R.A.; Patel, T.R.; et al. In vivo MRS measurement of 2-hydroxyglutarate in patient-derived IDH-mutant xenograft mouse models versus glioma patients. Magn. Reson. Med. 2020, 84, 1152–1160. [Google Scholar] [CrossRef]
- Linninger, A.; Hartung, G.A.; Liu, B.P.; Mirkov, S.; Tangen, K.; Lukas, R.V.; Unruh, D.; James, C.D.; Sarkaria, J.N.; Horbinski, C. Modeling the diffusion of D-2-hydroxyglutarate from IDH1 mutant gliomas in the central nervous system. Neuro Oncol. 2018, 20, 1197–1206. [Google Scholar] [CrossRef]
- Day, C.P.; Merlino, G.; Van Dyke, T. Preclinical mouse cancer models: A maze of opportunities and challenges. Cell 2015, 163, 39–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huszthy, P.C.; Daphu, I.; Niclou, S.P.; Stieber, D.; Nigro, J.M.; Sakariassen, P.; Miletic, H.; Thorsen, F.; Bjerkvig, R. In vivo models of primary brain tumors: Pitfalls and perspectives. Neuro Oncol. 2012, 14, 979–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richmond, A.; Su, Y. Mouse xenograft models vs GEM models for human cancer therapeutics. Dis. Model. Mech. 2008, 1, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosma, G.C.; Custer, R.P.; Bosma, M.J. A severe combined immunodeficiency mutation in the mouse. Nature 1983, 301, 527–530. [Google Scholar] [CrossRef]
- Shultz, L.D.; Lyons, B.L.; Burzenski, L.M.; Gott, B.; Chen, X.; Chaleff, S.; Kotb, M.; Gillies, S.D.; King, M.; Mangada, J.; et al. Human lymphoid and myeloid cell development in NOD/LtSz-scid IL2R gamma null mice engrafted with mobilized human hemopoietic stem cells. J. Immunol. 2005, 174, 6477–6489. [Google Scholar] [CrossRef] [Green Version]
- da Hora, C.C.; Schweiger, M.W.; Wurdinger, T.; Tannous, B.A. Patient-Derived Glioma Models: From Patients to Dish to Animals. Cells 2019, 8, 1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyai, M.; Tomita, H.; Soeda, A.; Yano, H.; Iwama, T.; Hara, A. Current trends in mouse models of glioblastoma. J. Neurooncol. 2017, 135, 423–432. [Google Scholar] [CrossRef]
- Choi, Y.; Lee, S.; Kim, K.; Kim, S.-H.; Chung, Y.-J.; Lee, C. Studying cancer immunotherapy using patient-derived xenografts (PDXs) in humanized mice. Exp. Mol. Med. 2018, 50, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morton, J.J.; Bird, G.; Refaeli, Y.; Jimeno, A. Humanized Mouse Xenograft Models: Narrowing the Tumor-Microenvironment Gap. Cancer Res. 2016, 76, 6153–6158. [Google Scholar] [CrossRef] [Green Version]
- Semenkow, S.; Li, S.; Kahlert, U.D.; Raabe, E.H.; Xu, J.; Arnold, A.; Janowski, M.; Chol Oh, B.; Brandacher, G.; Bulte, J.W.M.; et al. An immunocompetent mouse model of human glioblastoma. Oncotarget 2017, 8, 61072. [Google Scholar] [CrossRef] [Green Version]
- House, C.D.; Hernandez, L.; Annunziata, C.M. Recent technological advances in using mouse models to study ovarian cancer. Front. Oncol. 2014, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, B.; Mathew, R.K.; Polson, E.S.; Williams, J.; Wurdak, H. Spontaneous Glioblastoma Spheroid Infiltration of Early-Stage Cerebral Organoids Models Brain Tumor Invasion. Slas Discov. Adv. Life Sci. R D 2018, 23, 862–868. [Google Scholar] [CrossRef] [Green Version]
- Song, H.K.; Hwang, D.Y. Use of C57BL/6N mice on the variety of immunological researches. Lab. Anim. Res. 2017, 33, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Trunova, G.V.; Makarova, O.V.; Diatroptov, M.E.; Bogdanova, I.M.; Mikchailova, L.P.; Abdulaeva, S.O. Morphofunctional characteristic of the immune system in BALB/c and C57BL/6 mice. Bull. Exp. Biol. Med. 2011, 151, 99–102. [Google Scholar] [CrossRef]
- Taketo, M.; Schroeder, A.C.; Mobraaten, L.E.; Gunning, K.B.; Hanten, G.; Fox, R.R.; Roderick, T.H.; Stewart, C.L.; Lilly, F.; Hansen, C.T.; et al. FVB/N: An inbred mouse strain preferable for transgenic analyses. Proc. Natl. Acad. Sci. USA 1991, 88, 2065–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.M.; Bae, Y.M.; Choi, M.H.; Hong, S.T. Cyst formation, increased anti-inflammatory cytokines and expression of chemokines support for Clonorchis sinensis infection in FVB mice. Parasitol. Int. 2012, 61, 124–129. [Google Scholar] [CrossRef]
- Hede, S.M.; Hansson, I.; Afink, G.B.; Eriksson, A.; Nazarenko, I.; Andrae, J.; Genove, G.; Westermark, B.; Nistér, M. GFAP promoter driven transgenic expression of PDGFB in the mouse brain leads to glioblastoma in a Trp53 null background. Glia 2009, 57, 1143–1153. [Google Scholar] [CrossRef]
- Vitucci, M.; Irvin, D.M.; McNeill, R.S.; Schmid, R.S.; Simon, J.M.; Dhruv, H.D.; Siegel, M.B.; Werneke, A.M.; Bash, R.E.; Kim, S.; et al. Genomic profiles of low-grade murine gliomas evolve during progression to glioblastoma. Neuro Oncol. 2017, 19, 1237–1247. [Google Scholar] [CrossRef]
- Zhu, Y.; Guignard, F.; Zhao, D.; Liu, L.; Burns, D.K.; Mason, R.P.; Messing, A.; Parada, L.F. Early inactivation of p53 tumor suppressor gene cooperating with NF1 loss induces malignant astrocytoma. Cancer Cell 2005, 8, 119–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, A.; Wu, H.; Pandolfi, P.P.; Louis, D.N.; Van Dyke, T. Astrocyte inactivation of the pRb pathway predisposes mice to malignant astrocytoma development that is accelerated by PTEN mutation. Cancer Cell 2002, 1, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Xiao, A.; Yin, C.; Yang, C.; Di Cristofano, A.; Pandolfi, P.P.; Van Dyke, T. Somatic induction of Pten loss in a preclinical astrocytoma model reveals major roles in disease progression and avenues for target discovery and validation. Cancer Res. 2005, 65, 5172–5180. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Knobbe, C.B.; Itsumi, M.; Elia, A.J.; Harris, I.S.; Chio, I.I.; Cairns, R.A.; McCracken, S.; Wakeham, A.; Haight, J.; et al. D-2-hydroxyglutarate produced by mutant IDH1 perturbs collagen maturation and basement membrane function. Genes Dev. 2012, 26, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Lindberg, N.; Kastemar, M.; Olofsson, T.; Smits, A.; Uhrbom, L. Oligodendrocyte progenitor cells can act as cell of origin for experimental glioma. Oncogene 2009, 28, 2266–2275. [Google Scholar] [CrossRef] [Green Version]
- Uhrbom, L.; Kastemar, M.; Johansson, F.K.; Westermark, B.; Holland, E.C. Cell type-specific tumor suppression by Ink4a and Arf in Kras-induced mouse gliomagenesis. Cancer Res. 2005, 65, 2065–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhrbom, L.; Dai, C.; Celestino, J.C.; Rosenblum, M.K.; Fuller, G.N.; Holland, E.C. Ink4a-Arf loss cooperates with KRas activation in astrocytes and neural progenitors to generate glioblastomas of various morphologies depending on activated Akt. Cancer Res. 2002, 62, 5551–5558. [Google Scholar] [PubMed]
- Hambardzumyan, D.; Amankulor, N.M.; Helmy, K.Y.; Becher, O.J.; Holland, E.C. Modeling Adult Gliomas Using RCAS/t-va Technology. Transl. Oncol. 2009, 2, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Boije, M.; Westermark, B.; Uhrbom, L. PDGF-B Can sustain self-renewal and tumorigenicity of experimental glioma-derived cancer-initiating cells by preventing oligodendrocyte differentiation. Neoplasia 2011, 13, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Uhrbom, L.; Hesselager, G.; Ostman, A.; Nistér, M.; Westermark, B. Dependence of autocrine growth factor stimulation in platelet-derived growth factor-B-induced mouse brain tumor cells. Int. J. Cancer 2000, 85, 398–406. [Google Scholar] [CrossRef]
- Amankulor, N.M.; Kim, Y.; Arora, S.; Kargl, J.; Szulzewsky, F.; Hanke, M.; Margineantu, D.H.; Rao, A.; Bolouri, H.; Delrow, J.; et al. Mutant IDH1 regulates the tumor-associated immune system in gliomas. Genes Dev. 2017, 31, 774–786. [Google Scholar] [CrossRef] [Green Version]
- Kwon, C.H.; Zhao, D.; Chen, J.; Alcantara, S.; Li, Y.; Burns, D.K.; Mason, R.P.; Lee, E.Y.; Wu, H.; Parada, L.F. Pten haploinsufficiency accelerates formation of high-grade astrocytomas. Cancer Res. 2008, 68, 3286–3294. [Google Scholar] [CrossRef] [Green Version]
- de Vries, N.A.; Bruggeman, S.W.; Hulsman, D.; de Vries, H.I.; Zevenhoven, J.; Buckle, T.; Hamans, B.C.; Leenders, W.P.; Beijnen, J.H.; van Lohuizen, M.; et al. Rapid and robust transgenic high-grade glioma mouse models for therapy intervention studies. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 3431–3441. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Acquaviva, J.; Ramachandran, P.; Boskovitz, A.; Woolfenden, S.; Pfannl, R.; Bronson, R.T.; Chen, J.W.; Weissleder, R.; Housman, D.E.; et al. Oncogenic EGFR signaling cooperates with loss of tumor suppressor gene functions in gliomagenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 2712–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhrbom, L.; Hesselager, G.; Nistér, M.; Westermark, B. Induction of brain tumors in mice using a recombinant platelet-derived growth factor B-chain retrovirus. Cancer Res. 1998, 58, 5275–5279. [Google Scholar]
- Nunez, F.J.; Mendez, F.M.; Kadiyala, P.; Alghamri, M.S.; Savelieff, M.G.; Garcia-Fabiani, M.B.; Haase, S.; Koschmann, C.; Calinescu, A.A.; Kamran, N.; et al. IDH1-R132H acts as a tumor suppressor in glioma via epigenetic up-regulation of the DNA damage response. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef]
- Wei, Q.; Clarke, L.; Scheidenhelm, D.K.; Qian, B.; Tong, A.; Sabha, N.; Karim, Z.; Bock, N.A.; Reti, R.; Swoboda, R.; et al. High-grade glioma formation results from postnatal pten loss or mutant epidermal growth factor receptor expression in a transgenic mouse glioma model. Cancer Res. 2006, 66, 7429–7437. [Google Scholar] [CrossRef] [Green Version]
- Bender, A.M.; Collier, L.S.; Rodriguez, F.J.; Tieu, C.; Larson, J.D.; Halder, C.; Mahlum, E.; Kollmeyer, T.M.; Akagi, K.; Sarkar, G.; et al. Sleeping beauty-mediated somatic mutagenesis implicates CSF1 in the formation of high-grade astrocytomas. Cancer Res. 2010, 70, 3557–3565. [Google Scholar] [CrossRef] [Green Version]
- Marumoto, T.; Tashiro, A.; Friedmann-Morvinski, D.; Scadeng, M.; Soda, Y.; Gage, F.H.; Verma, I.M. Development of a novel mouse glioma model using lentiviral vectors. Nat. Med. 2009, 15, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Rankin, S.L.; Zhu, G.; Baker, S.J. Review: Insights gained from modelling high-grade glioma in the mouse. Neuropathol. Appl. Neurobiol. 2012, 38, 254–270. [Google Scholar] [CrossRef] [Green Version]
- Ahronian, L.G.; Lewis, B.C. Using the RCAS-TVA system to model human cancer in mice. Cold Spring Harb. Protoc. 2014, 2014, 1128–1135. [Google Scholar] [CrossRef] [PubMed]
- Holland, E.C.; Varmus, H.E. Basic fibroblast growth factor induces cell migration and proliferation after glia-specific gene transfer in mice. Proc. Natl. Acad. Sci. USA 1998, 95, 1218–1223. [Google Scholar] [CrossRef] [Green Version]
- Holland, E.C.; Hively, W.P.; DePinho, R.A.; Varmus, H.E. A constitutively active epidermal growth factor receptor cooperates with disruption of G1 cell-cycle arrest pathways to induce glioma-like lesions in mice. Genes Dev. 1998, 12, 3675–3685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouabe, H.; Okkenhaug, K. Gene targeting in mice: A review. Methods Mol. Biol. 2013, 1064, 315–336. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.H.; Kim, S.P. Surviving travel or travelling to survive: The association of travel distance with survival in muscle invasive bladder cancer. Transl. Androl. Urol. 2018, 7, S83–S85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mignone, J.L.; Kukekov, V.; Chiang, A.S.; Steindler, D.; Enikolopov, G. Neural stem and progenitor cells in nestin-GFP transgenic mice. J. Comp. Neurol. 2004, 469, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Weissenberger, J.; Steinbach, J.; Malin, G.; Spada, S.; Rülicke, T.; Aguzzi, A. Development and malignant progression of astrocytomas in GFAP-v. Oncogene 1997, 14, 2005–2013. [Google Scholar] [CrossRef] [Green Version]
- Reilly, K.M.; Loisel, D.A.; Bronson, R.T.; McLaughlin, M.E.; Jacks, T. Nf1;Trp53 mutant mice develop glioblastoma with evidence of strain-specific effects. Nat. Genet. 2000, 26, 109–113. [Google Scholar] [CrossRef]
- Borodovsky, A.; Meeker, A.K.; Kirkness, E.F.; Zhao, Q.; Eberhart, C.G.; Gallia, G.L.; Riggins, G.J. A model of a patient-derived IDH1 mutant anaplastic astrocytoma with alternative lengthening of telomeres. J. Neurooncol. 2015, 121, 479–487. [Google Scholar] [CrossRef]
- Yang, H.; Ye, D.; Guan, K.L.; Xiong, Y. IDH1 and IDH2 mutations in tumorigenesis: Mechanistic insights and clinical perspectives. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 5562–5571. [Google Scholar] [CrossRef] [Green Version]
- Philip, B.; Yu, D.X.; Silvis, M.R.; Shin, C.H.; Robinson, J.P.; Robinson, G.L.; Welker, A.E.; Angel, S.N.; Tripp, S.R.; Sonnen, J.A.; et al. Mutant IDH1 Promotes Glioma Formation In Vivo. Cell Rep. 2018, 23, 1553–1564. [Google Scholar] [CrossRef]
- Tateishi, K.; Wakimoto, H.; Iafrate, A.J.; Tanaka, S.; Loebel, F.; Lelic, N.; Wiederschain, D.; Bedel, O.; Deng, G.; Zhang, B.; et al. Extreme Vulnerability of IDH1 Mutant Cancers to NAD+ Depletion. Cancer Cell 2015, 28, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Rohle, D.; Popovici-Muller, J.; Palaskas, N.; Turcan, S.; Grommes, C.; Campos, C.; Tsoi, J.; Clark, O.; Oldrini, B.; Komisopoulou, E.; et al. An Inhibitor of Mutant IDH1 Delays Growth and Promotes Differentiation of Glioma Cells. Science 2013, 340, 626–630. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, T.; Bunse, L.; Pusch, S.; Sahm, F.; Wiestler, B.; Quandt, J.; Menn, O.; Osswald, M.; Oezen, I.; Ott, M.; et al. A vaccine targeting mutant IDH1 induces antitumour immunity. Nature 2014, 512, 324–327. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yu, J.; Tu, L.; Huang, N.; Li, H.; Luo, Y. Isocitrate Dehydrogenase Mutations in Glioma: From Basic Discovery to Therapeutics Development. Front. Oncol. 2019, 9, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, M.S.; Mellinghoff, I.K. Histone-Mutant Glioma: Molecular Mechanisms, Preclinical Models, and Implications for Therapy. Int. J. Mol. Sci. 2020, 21, 7193. [Google Scholar] [CrossRef] [PubMed]
- Lowe, B.R.; Maxham, L.A.; Hamey, J.J.; Wilkins, M.R.; Partridge, J.F. Histone H3 Mutations: An Updated View of Their Role in Chromatin Deregulation and Cancer. Cancers 2019, 11, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosaab, A.; El-Ayadi, M.; Khorshed, E.N.; Amer, N.; Refaat, A.; El-Beltagy, M.; Hassan, Z.; Soror, S.H.; Zaghloul, M.S.; El-Naggar, S. Histone H3K27M Mutation Overrides Histological Grading in Pediatric Gliomas. Sci. Rep. 2020, 10, 8368. [Google Scholar] [CrossRef]


{kind=link}
| Model | Tumor Source | Immune Status | Cost | Labor/Time |
|---|---|---|---|---|
| CLX | Human | (−) | $ | + |
| PDX | Human | (−) | $$ | ++ |
| Syngeneic | Mouse | (+) | $ | ++ |
| GEMM | Mouse | (+) | $$$ | +++ |
| Mouse Species | Brain Tumor Modeled | Tumor Cell Line | Reference |
|---|---|---|---|
| BALB/c OlaHsd-Foxn1nu | Glioma | BT3 cells | [29] |
| C57BL6/N | High Grade Glioma | GL261 cells | [30,31,32,33,34,35,36,37] |
| High Grade Glioma | U87 and GL261 cells | [38] | |
| CBA, BALB/c, AKR, C57 black, and RIII | Glioma | Rat C6 cells | [27] |
| CD-1, Nude, and NOD CRISPR Prkdc IL2Rγnull | Glioma | DAOY and T98G cells | [39] |
| CIEA-NOG | Glioblastoma (GBM) | Patient derived glioma cell lines | [40] |
| C6B3F1 | High Grade Glioma | Mouse Tu2449, Tu9648 and Tu251 mouse glioma cell lines | [41] |
| Foxn1nu/nu | Glioma | BT4C cells | [29,42] |
| ICR | High Grade Glioma | C6 rat glioma cells | [43] |
| Non-obese/diabetic (NOD)/Severe combined immunodeficient (SCID) | Isocitrate dehydrogenase 1 (IDH1) Mutated Glioma | Patient derived IDH1 mutant oligoastrocytoma | [44] |
| GBM | TG1 human GBM cell line | [45] | |
| GBM | T98 and U87 glioma cell lines | [46] | |
| Nude/NOD/SCID | High Grade Glioma | U87, U118, LN18, LN229 cell lines | [47] |
| Not reported | GBM | U87 and U373 glioma cell lines | [48] |
| Nude | GBM | Ink4a/ARF−/− Id4 astrocyte cells | [49] |
| Malignant Astrocytoma | Commercial malignant cell lines | [50] | |
| High Grade Glioma | BT70 malignant glioma cell line | [51] | |
| High Grade Glioma | U87 human glioma cell line and C6 rat glioma cell line | [52] | |
| GBM | LN229 and U87 human glioma cells | [53] | |
| High Grade Glioma | E98 and E473 glioma cell lines | [54] | |
| GBM | Mouse GL261 cell line | [55] | |
| GBM | Human U87 glioma and rat 9L glio-sarcoma cell lines | [56] | |
| GBM | Patient derived GBM cell lines | [57,58,59] | |
| High Grade Glioma | Human astroglioma U373 and T98G and oligodendroglioma Hs683 cell lines | [60] | |
| High Grade Glioma | Human glioma U87, U251, U373, A172, LN18, LN229, and D54 cell lines | [61] | |
| High Grade Glioma | Hs683 cells | [62] | |
| High Grade Glioma | LN229 cells | [63,64] | |
| High Grade Glioma | SHG44 cells | [65] | |
| High Grade Glioma | T98G and U373 cells | [66] | |
| High Grade Glioma | U87, U251 and D566 cells | [67] | |
| High Grade Glioma | U87 cells | [68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90] | |
| GBM | U87. LNZ308, LN229 cells | [91] | |
| High Grade Glioma | U87, U118, N10, U251, A172, and U373 cell lines | [92] | |
| High Grade Glioma | U251 cell line | [93,94] | |
| GBM | U87 and LN229 cell lines | [95] | |
| GBM | LN229 cell line | [96] | |
| Glioma | E102 and E106 glioma cell lines | [97] | |
| Glioma | SNB-19 U87 glioma cell lines with co-transfecting COS-7 cells with pTet-On and treated with doxycycline | [98] | |
| High Grade Glioma | Human T269 4IgB7H3 knockdown or control cells (orthotopic); LN-229 4IgB7H3 knockdown or control cells (subcutaneous) | [99] | |
| High Grade Glioma | U87 and U251 glioma cell lines | [100] | |
| Glioma | A-172, U343, U87 and T98G glioma cells | [101] | |
| Glioma | U87 glioma cell line | [102] | |
| Glioma | U373 human glioma cell line | [103] | |
| SCID | High Grade Glioma | GLI36-EGFRvIII engineered cells | [104] |
| Glioma | Patient-derived GSC lines | [105,106,107] |
| Mouse Species | Brain Tumor Modeled | Source of Tumor Cells | Reference |
|---|---|---|---|
| eGFP NOD/SCID mice | Oligodendroglioma | Patient-derived tumor cells | [108] |
| NOD-PrkdcSCID IL2Rγnull | Grade II-IV Glioma | Patient-derived glioma tissue | [109] |
| NOD-SCID | Malignant Astrocytoma | Embryonic stem cells | [110] |
| Glioma | Patient-derived high grade glioma tissue | [111] | |
| NOD/SCID Il2rg−/− (NOG) | GBM | Patient-derived GBM cells | [112] |
| Not reported | GBM | Patient-derived human GBM cells | [113] |
| NSG | GBM | Patient-derived GBM neuro-spheres | [114] |
| Nude | Glioma | Patient-derived IDH mutant glioma tissue | [115] |
| GBM | Patient-derived GBM tissue | [116,117] | |
| GBM | Patient-derived GBM tissue | [28,118] | |
| SCID | IDH Mutant Glioma | Patient-derived glioma neuro-spheres | [119] |
| IDH1 Mutated Glioma | GBM164, GBM196, and TB09 IDH1 mutant glioma PDX models | [120,121] |
| Mouse Species | Brain Tumor Modeled | Genetic Engineering | Reference |
|---|---|---|---|
| C57BL/6 and Tp53−/− | GBM | PDGFβ, p53 mutations | [138] |
| C57BL/6 | Glioma | Heterozygous TgGZT121, KRASG12D, GFAP-CreER, PP-CreER, NG2-CreER, and Rosa26-tdTomato mice crossed with mice with conditional PTEN, p53, Rb1, or NF1 loss | [139] |
| Glioma | Crossing of NF1flox+ mice with p53+/− mice and then crossed with wild type F1 C57BL/6 mice | [140] | |
| High Grade Astrocytoma | RB, phosphate and tensin homolog deleted on chromosome 10 (PTEN) mutations | [141,142] | |
| Crossed IDH1 and Nestin-Cre transgenic mice | IDH1 R132 Mutated Glioma | Nestin-Cre remodeling system | [143] |
| FVB/N mice | Oligodendrocyte | Ctv-a plasmid was transfected into an immortalized oligodendroglia cell line OLI-neu | [144] |
| FVB/N, C57BL/6, BALB/C, and 129 | GBM | K-Ras, Akt, Ink4a/Arf mutations | [145,146] |
| GBM | Platelet derived growh factor receptor (PDGF)β, Ink4a/Arf, PTEN mutations | [147] | |
| Gtv-a Arf−/− | High Grade Glioma | Induction with RCAS-PDGF-B | [148] |
| INK4a+/+ and INK4a−/− | GBM | PDGFβ | [149] |
| IDH1 mutant and wildtype gliomas | PDGF, Tp53, and IDH1 mutations | [150] | |
| MUT3 (Mice with mixed genetic background of C57BL/6, Sv129 and B6/CBA) | De novo GBM | Introduced PTEN and p53 flexed alleles into MUT3 mice | [151] |
| Not Reported | High Grade Glioma | K-Ras, p53, Ink4a/Arf mutations | [152] |
| GBM | EGFRvIII, Ink4a/Arf, PTEN mutations | [153] | |
| GBM | PDGFβ mutation | [154] | |
| Lower- and Higher-Grade Gliomas | Neuroblastoma RAS (NRAS) G12V and shp53 Sleeping beauty plasmids with or without shATRX and IDH1R132H | [155] | |
| RasB8 | High Grade Glioma | EGFRvIII and V12 Ras mutation | [156] |
| Rosa26-SB11 | High Grade Glioma | T2/onc mutagenic transposon | [157] |
| GFAP-Cre transgenic mice maintained with C57BL/6 and crossed with Tp53−/− mice | High Grade Glioma | H-Ras, AKT and Tp53 mutations | [158] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hicks, W.H.; Bird, C.E.; Traylor, J.I.; Shi, D.D.; El Ahmadieh, T.Y.; Richardson, T.E.; McBrayer, S.K.; Abdullah, K.G. Contemporary Mouse Models in Glioma Research. Cells 2021, 10, 712. https://doi.org/10.3390/cells10030712
Hicks WH, Bird CE, Traylor JI, Shi DD, El Ahmadieh TY, Richardson TE, McBrayer SK, Abdullah KG. Contemporary Mouse Models in Glioma Research. Cells. 2021; 10(3):712. https://doi.org/10.3390/cells10030712
Chicago/Turabian StyleHicks, William H., Cylaina E. Bird, Jeffrey I. Traylor, Diana D. Shi, Tarek Y. El Ahmadieh, Timothy E. Richardson, Samuel K. McBrayer, and Kalil G. Abdullah. 2021. "Contemporary Mouse Models in Glioma Research" Cells 10, no. 3: 712. https://doi.org/10.3390/cells10030712
APA StyleHicks, W. H., Bird, C. E., Traylor, J. I., Shi, D. D., El Ahmadieh, T. Y., Richardson, T. E., McBrayer, S. K., & Abdullah, K. G. (2021). Contemporary Mouse Models in Glioma Research. Cells, 10(3), 712. https://doi.org/10.3390/cells10030712