
IL-1β Antibody Protects Brain from Neuropathology of Hypoperfusion

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
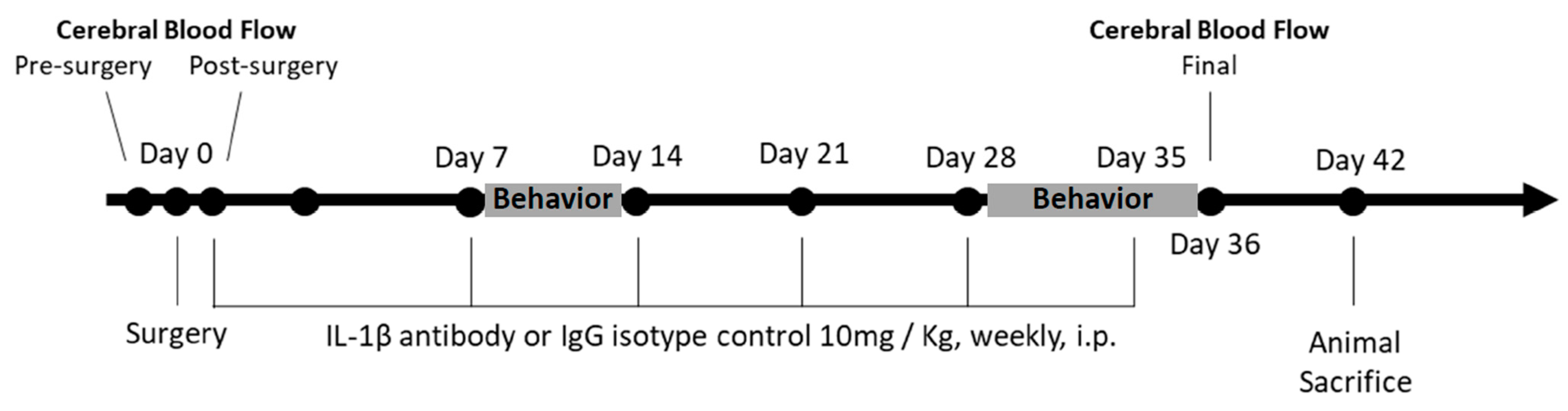
2.2. Implantation of Ameroid Constrictor Ring and Microcoil
2.3. Administration of IL-1β
2.4. Cerebral Blood Flow
2.5. Behavioral Assessment
2.6. Histochemistry
2.7. Statistical Analysis
3. Results
3.1. Body Weight and Mortality
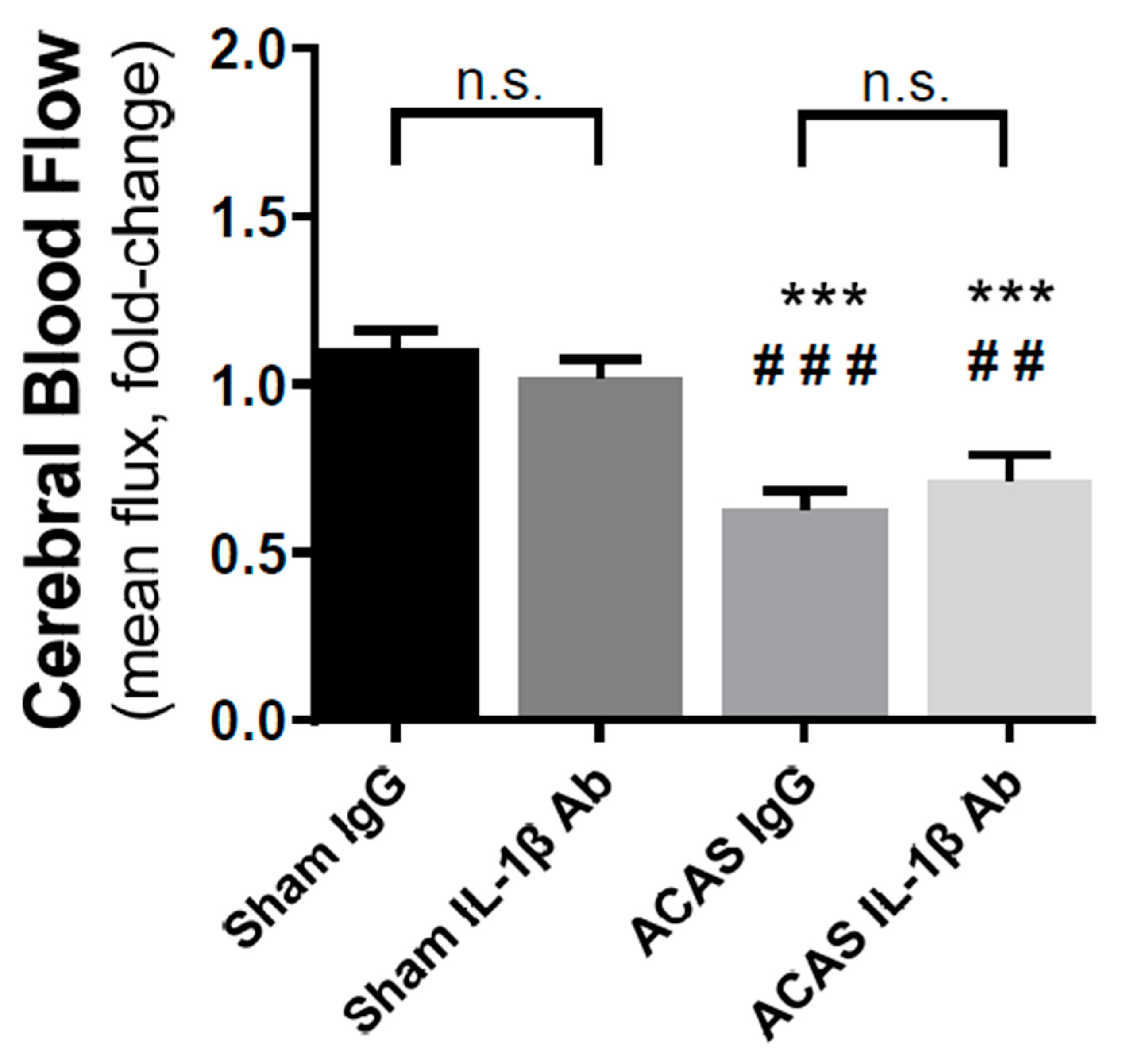
3.2. Cerebral Blood Flow
3.3. Grey Matter Damage
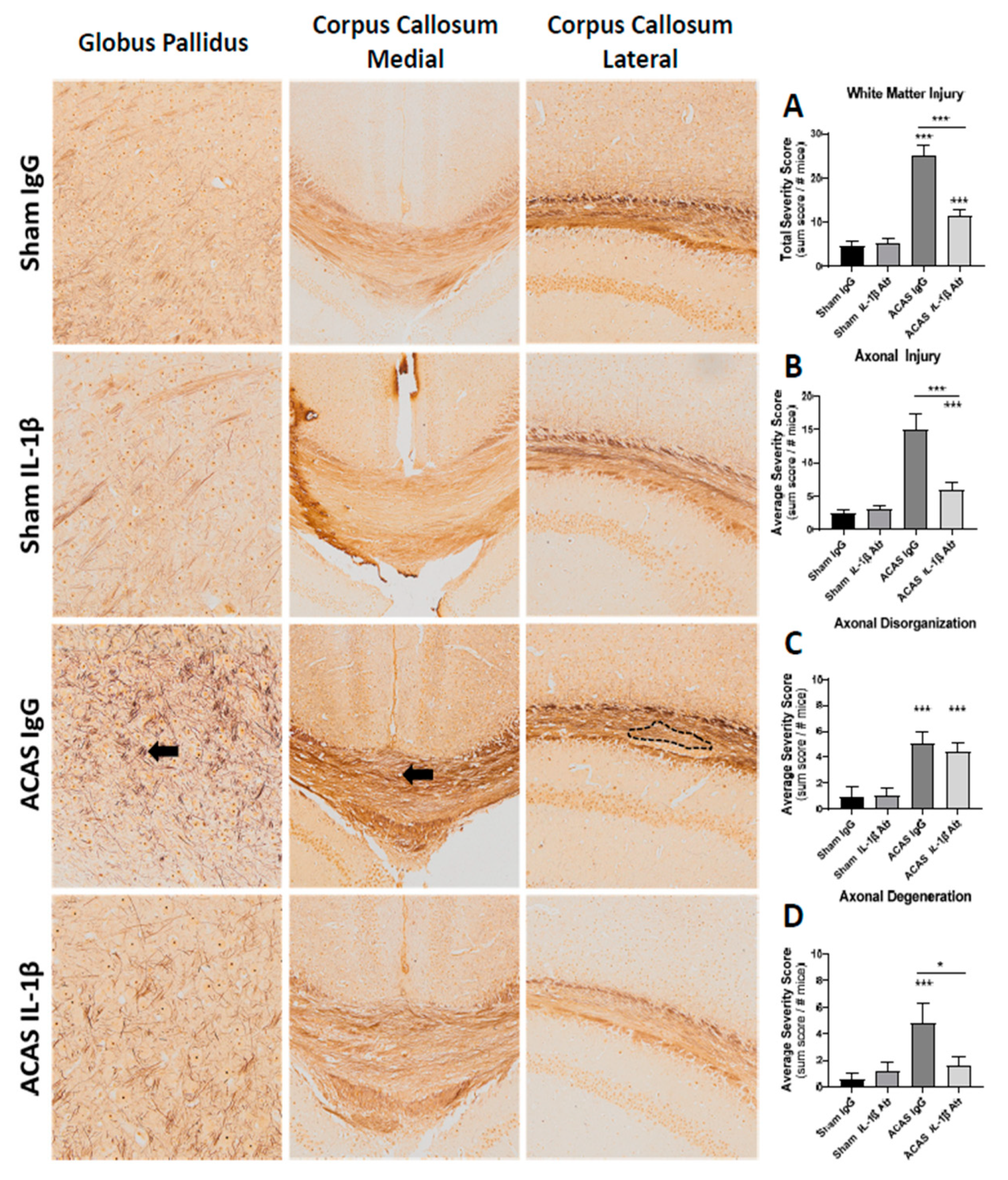
3.4. White Matter Damage
3.5. Behavioral Outcomes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kynast, J.; Lampe, L.; Luck, T.; Frisch, S.; Arelin, K.; Hoffmann, K.-T.; Loeffler, M.; Riedel-Heller, S.G.; Villringer, A.; Schroeter, M.L. White matter hyperintensities associated with small vessel disease impair social cognition beside attention and memory. Br. J. Pharmacol. 2018, 38, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Black, S.; Gao, F.; Bilbao, J. Understanding White Matter Disease: Imaging-Pathological Correlations in Vascular Cognitive Impairment. Stroke 2008, 40 (Suppl. S3), S48–S52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borroni, B.; Perani, D.; Broli, M.; Colciaghi, F.; Garibotto, V.; Paghera, B.; Agosti, C.; Giubbini, R.; Di Luca, M.; Padovani, A. Pre–clinical diagnosis of Alzheimer disease combining platelet amyloid precursor protein ratio and rCBF spect analysis. J. Neurol. 2005, 252, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Wyper, D.J.; Owens, J.; Patterson, J.; Kelly, R.C.; Hunter, R.; McCulloch, J. 123Iodo-MK-801: A spect agent for imaging the pattern and extent of glutamate (NMDA) receptor activation in Alzheimer’s disease. J. Psychiatr. Res. 1997, 31, 605–619. [Google Scholar] [CrossRef]
- Chao, L.L.; Buckley, S.T.; Kornak, J.; Schuff, N.; Madison, C.; Yaffe, K.; Miller, B.L.; Kramer, J.H.; Weiner, M.W. ASL Perfusion MRI Predicts Cognitive Decline and Conversion from MCI to Dementia. Alzheimer Dis. Assoc. Disord. 2010, 24, 19–27. [Google Scholar] [CrossRef]
- Vasquez, B.P.; Zakzanis, K.K. The neuropsychological profile of vascular cognitive impairment not demented: A meta-analysis. J. Neuropsychol. 2015, 9, 109–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y.; Zheng, Y.; Luo, Y.; Du, Y.; Zhao, Y.; Guan, J.; Zhang, X.; Fu, J. TREM-2-p38 MAPK signaling regulates neuroinflammation during chronic cerebral hypoperfusion combined with diabetes mellitus. J. Neuroinflamm. 2020, 17, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farkas, E.; Luiten, P.G.; Bari, F. Permanent, bilateral common carotid artery occlusion in the rat: A model for chronic cerebral hypoperfusion-related neurodegenerative diseases. Brain Res. Rev. 2007, 54, 162–180. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhang, B.; Lu, K.; Deng, J.; Zhao, F.; Zhao, B.-Q.; Zhao, Y. Prevention of Hippocampal Neuronal Damage and Cognitive Function Deficits in Vascular Dementia by Dextromethorphan. Mol. Neurobiol. 2016, 53, 3494–3502. [Google Scholar] [CrossRef]
- Fu, X.; Zhang, J.; Guo, L.; Xu, Y.; Sun, L.; Wang, S.; Feng, Y.; Gou, L.; Zhang, L.; Liu, Y. Protective role of luteolin against cognitive dysfunction induced by chronic cerebral hypoperfusion in rats. Pharmacol. Biochem. Behav. 2014, 126, 122–130. [Google Scholar] [CrossRef]
- Hou, X.; Liang, X.; Chen, J.-F.; Zheng, J. Ecto-5′-nucleotidase (CD73) is involved in chronic cerebral hypoperfusion-induced white matter lesions and cognitive impairment by regulating glial cell activation and pro-inflammatory cytokines. Neuroscience 2015, 297, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Lambertsen, K.L.; Biber, K.; Finsen, B. Inflammatory Cytokines in Experimental and Human Stroke. Br. J. Pharmacol. 2012, 32, 1677–1698. [Google Scholar] [CrossRef] [Green Version]
- Lambertsen, K.L.; Finsen, B.; Clausen, B.H. Post-stroke inflammation—Target or tool for therapy? Acta Neuropathol. 2019, 137, 693–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaventura, A.; Liberale, L.; Vecchié, A.; Casula, M.; Carbone, F.; Dallegri, F.; Montecucco, F. Update on Inflammatory Biomarkers and Treatments in Ischemic Stroke. Int. J. Mol. Sci. 2016, 17, 1967. [Google Scholar] [CrossRef]
- Emsley, H.C.A.; Smith, C.J.; Georgiou, R.F.; Vail, A.; Hopkins, S.J.; Rothwell, N.J.; Tyrrell, P.J. A randomised phase II study of interleukin-1 receptor antagonist in acute stroke patients. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1366–1372. [Google Scholar] [CrossRef] [Green Version]
- Hattori, Y.; Enmi, J.-I.; Kitamura, A.; Yamamoto, Y.; Saito, S.; Takahashi, Y.; Iguchi, S.; Tsuji, M.; Yamahara, K.; Nagatsuka, K.; et al. A Novel Mouse Model of Subcortical Infarcts with Dementia. J. Neurosci. 2015, 35, 3915–3928. [Google Scholar] [CrossRef]
- Quintana, D.D.; Ren, X.; Hu, H.; Engler-Chiurazzi, E.B.; Rellick, S.L.; Lewis, S.E.; Povroznik, J.M.; Simpkins, J.W.; Alvi, M. Gradual common carotid artery occlusion as a novel model for cerebrovascular Hypoperfusion. Metab. Brain Dis. 2018, 33, 2039–2044. [Google Scholar] [CrossRef]
- Lachmann, H.J.; Kone-Paut, I.; Kuemmerle-Deschner, J.B.; Leslie, K.S.; Hachulla, E.; Quartier, P.; Gitton, X.; Widmer, A.; Patel, N.; Hawkins, P.N. Use of Canakinumab in the Cryopyrin-Associated Periodic Syndrome. N. Engl. J. Med. 2009, 360, 2416–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aday, A.W.; Ridker, P.M. Antiinflammatory Therapy in Clinical Care: The CANTOS Trial and Beyond. Front. Cardiovasc. Med. 2018, 5, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhimolea, E. Canakinumab. mAbs 2010, 2, 3–13. [Google Scholar] [CrossRef]
- Osborn, O.; Brownell, S.E.; Sanchez-Alavez, M.; Salomon, D.; Gram, H.; Bartfai, T. Treatment with an Interleukin 1 beta antibody improves glycemic control in diet-induced obesity. Cytokine 2008, 44, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Gram, H. The long and winding road in pharmaceutical development of canakinumab from rare genetic autoinflammatory syndromes to myocardial infarction and cancer. Pharmacol. Res. 2020, 154, 104139. [Google Scholar] [CrossRef] [PubMed]
- Alamed, J.; Wilcock, D.M.; Diamond, D.M.; Gordon, M.N.; Morgan, D. Two-day radial-arm water maze learning and memory task; robust resolution of amyloid-related memory deficits in transgenic mice. Nat. Protoc. 2006, 1, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Doll, D.N.; Engler-Chiurazzi, E.B.; Lewis, S.E.; Hu, H.; Kerr, A.E.; Ren, X.; Simpkins, J.W. Lipopolysaccharide exacerbates infarct size and results in worsened post-stroke behavioral outcomes. Behav. Brain Funct. 2015, 11, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Mahmood, A.; Lu, D.; Qu, C.; Kazmi, H.; Goussev, A.; Zhang, Z.G.; Noguchi, C.T.; Schallert, T.; Chopp, M. Histological and functional outcomes after traumatic brain injury in mice null for the erythropoietin receptor in the central nervous system. Brain Res. 2008, 1230, 247–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchihara, T. Silver diagnosis in neuropathology: Principles, practice and revised interpretation. Acta Neuropathol. 2007, 113, 483–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esenwa, C.C.; Elkind, M.S. Inflammatory risk factors, biomarkers and associated therapy in ischaemic stroke. Nat. Rev. Neurol. 2016, 12, 594–604. [Google Scholar] [CrossRef]
- Rayasam, A.; Hsu, M.; Kijak, J.A.; Kissel, L.; Hernandez, G.; Sandor, M.; Fabry, Z. Immune responses in stroke: How the immune system contributes to damage and healing after stroke and how this knowledge could be translated to better cures? Immunology 2018, 154, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Hu, H.; Farooqi, I.; Simpkins, J.W. Blood substitution therapy rescues the brain of mice from ischemic damage. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Voet, S.; Srinivasan, S.; Lamkanfi, M.; Van Loo, G. Inflammasomes in neuroinflammatory and neurodegenerative diseases. EMBO Mol. Med. 2019, 11, e10248. [Google Scholar] [CrossRef]
- Patterson, S.L. Immune dysregulation and cognitive vulnerability in the aging brain: Interactions of microglia, IL-1β, BDNF and synaptic plasticity. Neuropharmacology 2015, 96, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Gram, H. Preclinical characterization and clinical development of ILARIS® (canakinumab) for the treatment of autoinflammatory diseases. Curr. Opin. Chem. Biol. 2016, 32, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rothwell, N. Interleukin-1 and neuronal injury: Mechanisms, modification, and therapeutic potential. Brain Behav. Immun. 2003, 17, 152–157. [Google Scholar] [CrossRef]
- Mulcahy, N.J.; Ross, J.; Rothwell, N.J.; Loddick, S.A. Delayed administration of interleukin-1 receptor antagonist protects against transient cerebral ischaemia in the rat. Br. J. Pharmacol. 2003, 140, 471–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touzani, O.; Boutin, H.; Chuquet, J.; Rothwell, N. Potential mechanisms of interleukin-1 involvement in cerebral ischaemia. J. Neuroimmunol. 1999, 100, 203–215. [Google Scholar] [CrossRef]
- Li, X.; Blizzard, K.K.; Zeng, Z.; DeVries, A.; Hurn, P.D.; McCullough, L.D. Chronic behavioral testing after focal ischemia in the mouse: Functional recovery and the effects of gender. Exp. Neurol. 2004, 187, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Schönfeld, L.-M.; Jahanshahi, A.; Lemmens, E.; Bauwens, M.; Hescham, S.-A.; Schipper, S.; Lagiere, M.; Hendrix, S.; Temel, Y. Motor cortex stimulation does not lead to functional recovery after experimental cortical injury in rats. Restor. Neurol. Neurosci. 2017, 35, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Huang, R.; Shetty, R.A.; Thangthaeng, N.; Liu, R.; Chen, Z.; Sumien, N.; Rutledge, M.; Dillon, G.H.; Yuan, F.; et al. Transient focal cerebral ischemia induces long-term cognitive function deficit in an experimental ischemic stroke model. Neurobiol. Dis. 2013, 59, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Hattori, Y.; Enmi, J.I.; Iguchi, S.; Saito, S.; Yamamoto, Y.; Tsuji, M.; Nagatsuka, K.; Kalaria, R.N.; Iida, H.; Ihara, M. Gradual Carotid Artery Stenosis in Mice Closely Replicates Hypoperfusive Vascular Dementia in Humans. J. Am. Heart Assoc. 2016, 5, e002022. [Google Scholar] [CrossRef] [Green Version]
- Nishio, K.; Ihara, M.; Yamasaki, N.; Kalaria, R.N.; Maki, T.; Fujita, Y.; Ito, H.; Oishi, N.; Fukuyama, H.; Miyakawa, T.; et al. A Mouse Model Characterizing Features of Vascular Dementia with Hippocampal Atrophy. Stroke 2010, 41, 1278–1284. [Google Scholar] [CrossRef] [Green Version]
- Shibata, M.; Yamasaki, N.; Miyakawa, T.; Kalaria, R.N.; Fujita, Y.; Ohtani, R.; Ihara, M.; Takahashi, R.; Tomimoto, H. Selective Impairment of Working Memory in a Mouse Model of Chronic Cerebral Hypoperfusion. Stroke 2007, 38, 2826–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, A.; Saito, S.; Maki, T.; Oishi, N.; Ayaki, T.; Hattori, Y.; Yamamoto, Y.; Urushitani, M.; Kalaria, R.N.; Fukuyama, H.; et al. Gradual cerebral hypoperfusion in spontaneously hypertensive rats induces slowly evolving white matter abnormalities and impairs working memory. Br. J. Pharmacol. 2016, 36, 1592–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuloaga, K.L.; Johnson, L.A.; Roese, N.E.; Marzulla, T.; Zhang, W.; Nie, X.; Alkayed, F.N.; Hong, C.; Grafe, M.R.; Pike, M.M.; et al. High fat diet-induced diabetes in mice exacerbates cognitive deficit due to chronic hypoperfusion. Br. J. Pharmacol. 2015, 36, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Maher, C.O.; Anderson, R.E.; Martin, H.S.; McClelland, R.L.; Meyer, F.B. Interleukin-1β and adverse effects on cerebral blood flow during long-term global hypoperfusion. J. Neurosurg. 2003, 99, 907–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, A.; Tannenbaum, S.; Rordorf, C.; Lowe, P.J.; Floch, D.; Gram, H.; Roy, S. Pharmacokinetic and Pharmacodynamic Properties of Canakinumab, a Human Anti-Interleukin-1β Monoclonal Antibody. Clin. Pharmacokinet. 2012, 51, e1–e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grey Matter Damage | |
| Score | Degenerative Lesion |
| 1 | Anomolous structure |
| 2 | Necrotic tissue |
| 3 | Hole or tissue atrophy |
| Score | Pyramidal Cells Loss |
| 1 | Irregular cell layer |
| 2 | Thinning of cell layer |
| 3 | Gap in cell layer |
| Score | Granular Cells Loss |
| 1 | Apoptotic cells |
| 2 | Thinning of stratum granulosum |
| 3 | Gap in stratum granulosum |
| Score | Vacuolization |
| 1 | Sparse vacuoles |
| 2 | Moderate vacuoles |
| 3 | Dense vacuoles |
| White Matter Damage | |
| Score | Axonal Injury |
| 1 | Sparse axon damage |
| 2 | Moderate axon damage |
| 3 | Dense axon damage |
| Score | Axonal Disorganization |
| 1 | Majority of axon bundles retained |
| 2 | Moderate retention of axon bundles |
| 3 | Majority of axon bundles lost |
| Score | Axonal Degeneration |
| 1 | Sparse tortuous axons |
| 2 | Moderate tortuous axons |
| 3 | Dense tortuous axons |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quintana, D.; Ren, X.; Hu, H.; Corbin, D.; Engler-Chiurazzi, E.; Alvi, M.; Simpkins, J. IL-1β Antibody Protects Brain from Neuropathology of Hypoperfusion. Cells 2021, 10, 855. https://doi.org/10.3390/cells10040855
Quintana D, Ren X, Hu H, Corbin D, Engler-Chiurazzi E, Alvi M, Simpkins J. IL-1β Antibody Protects Brain from Neuropathology of Hypoperfusion. Cells. 2021; 10(4):855. https://doi.org/10.3390/cells10040855
Chicago/Turabian StyleQuintana, Dominic, Xuefang Ren, Heng Hu, Deborah Corbin, Elizabeth Engler-Chiurazzi, Muhammad Alvi, and James Simpkins. 2021. "IL-1β Antibody Protects Brain from Neuropathology of Hypoperfusion" Cells 10, no. 4: 855. https://doi.org/10.3390/cells10040855
APA StyleQuintana, D., Ren, X., Hu, H., Corbin, D., Engler-Chiurazzi, E., Alvi, M., & Simpkins, J. (2021). IL-1β Antibody Protects Brain from Neuropathology of Hypoperfusion. Cells, 10(4), 855. https://doi.org/10.3390/cells10040855