
Cross-Reactivity and Sequence Homology Between Alpha-Synuclein and Food Products: A Step Further for Parkinson’s Disease Synucleinopathy




Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibodies and Antigens
2.2. Preparation of Food Antigens
2.3. Enzyme Linked Immunosorbent Assay (ELISA) for Demonstrating the Reaction of Various Antibodies With Food Antigens
2.4. Determination of Specificity of Antigen-Antibody Reaction
2.5. Amino Acid Sequence Similarity Between Alpha-Synuclein and Food Proteins
2.6. Statistical Analysis
3. Results
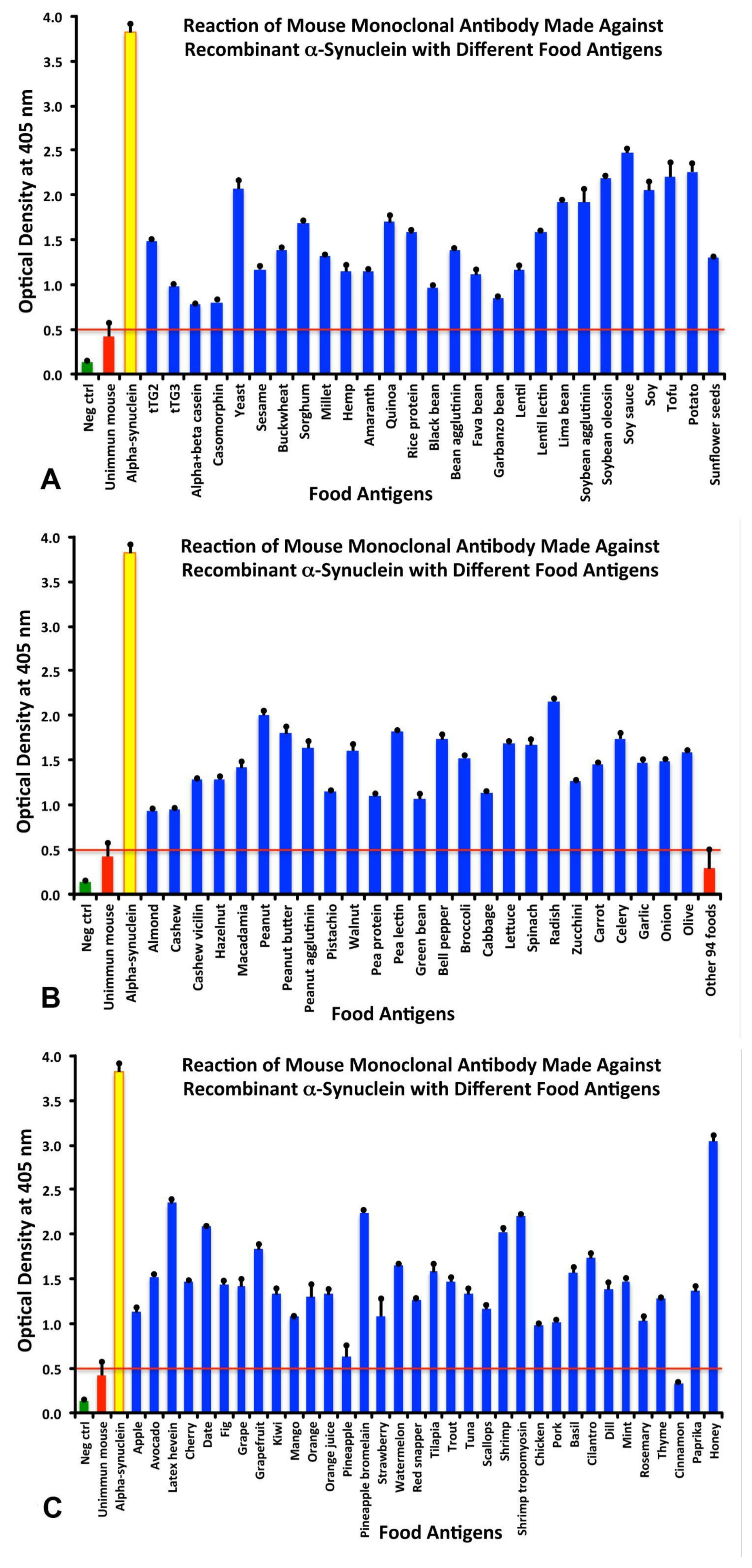
3.1. Cross Reactivity Between aSN and Different Food Antigens.
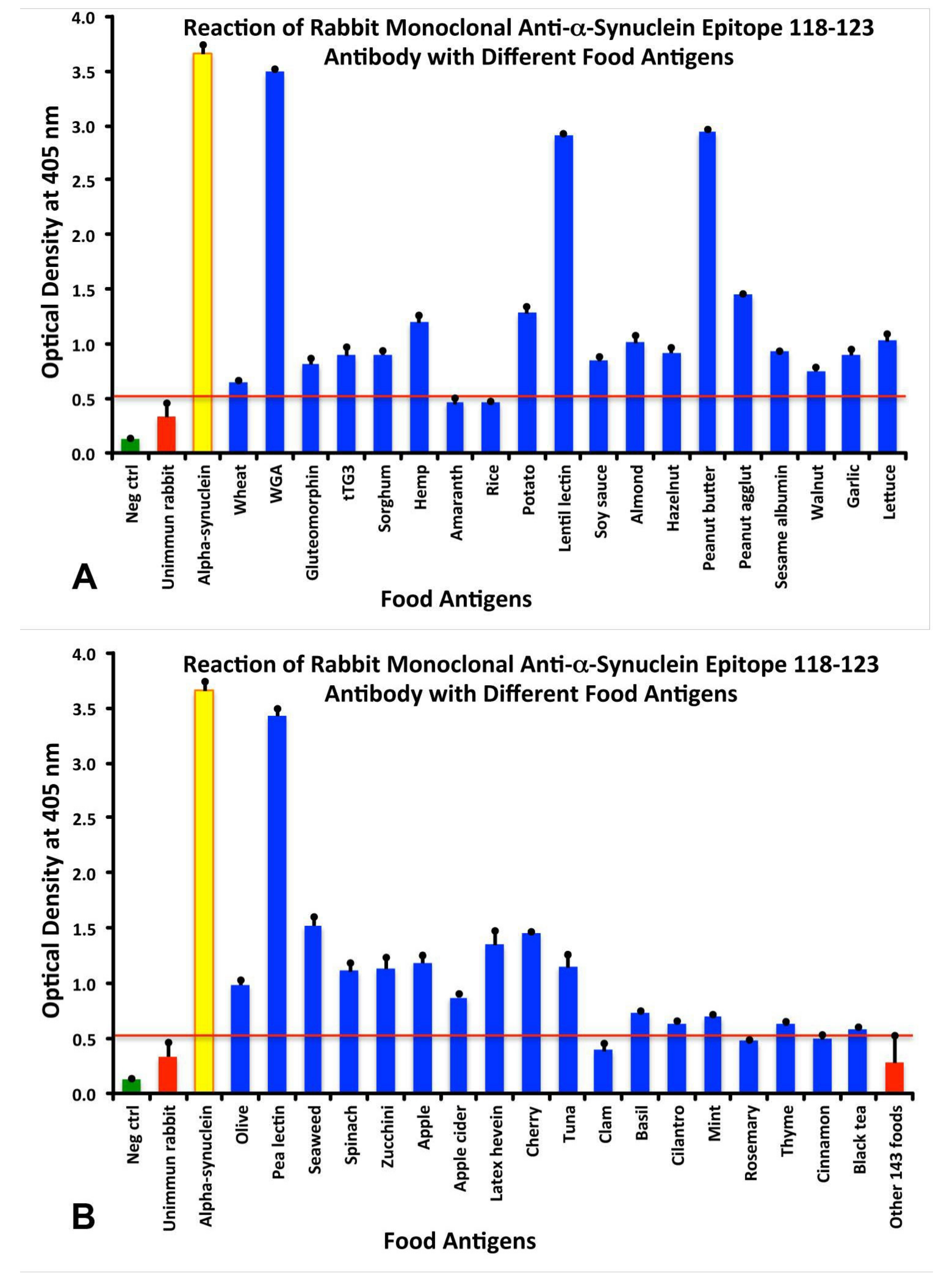
3.2. Cross Reactivity Between the Epitope 118–123 of aSN with Different Food Antigens
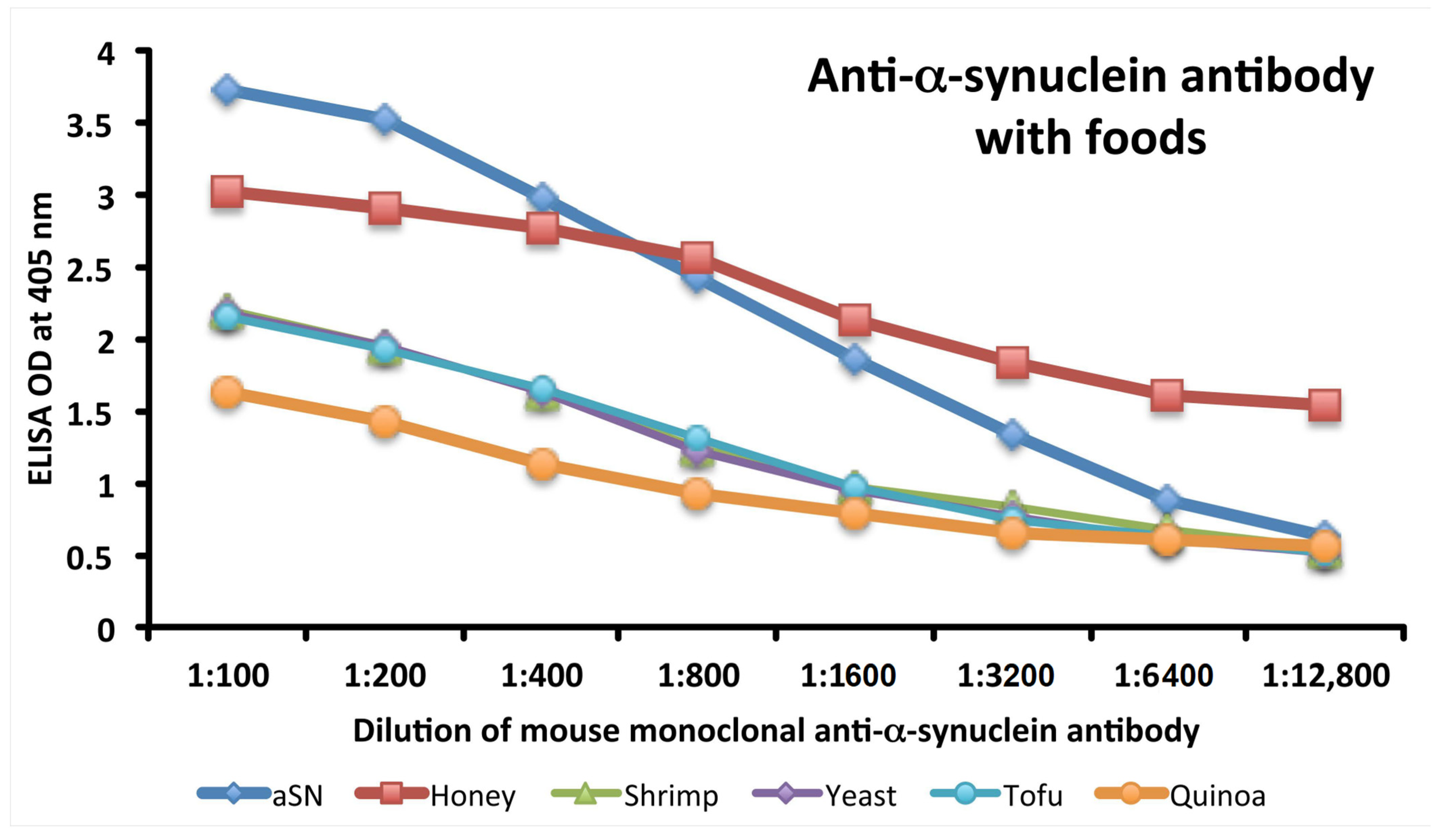
3.3. Analytical Specificity of Anti-aSN Antibody Binding to Selected Food Antigens Performed by Serial Dilutions
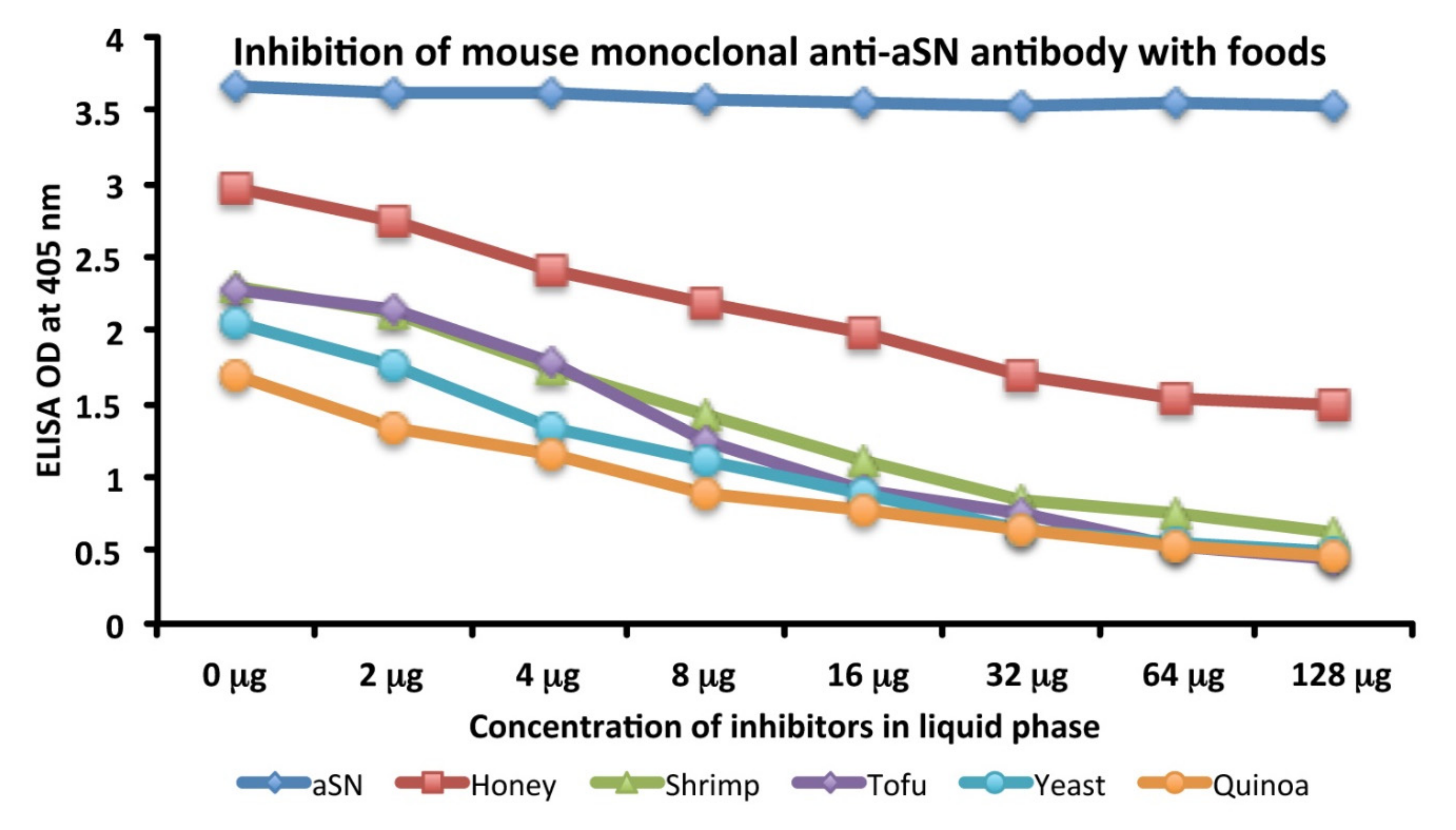
3.4. Analytical Specificity of Mouse Monoclonal Anti-aSN Antibody Binding to Selected Food Antigens Performed by Serial Inhibitions
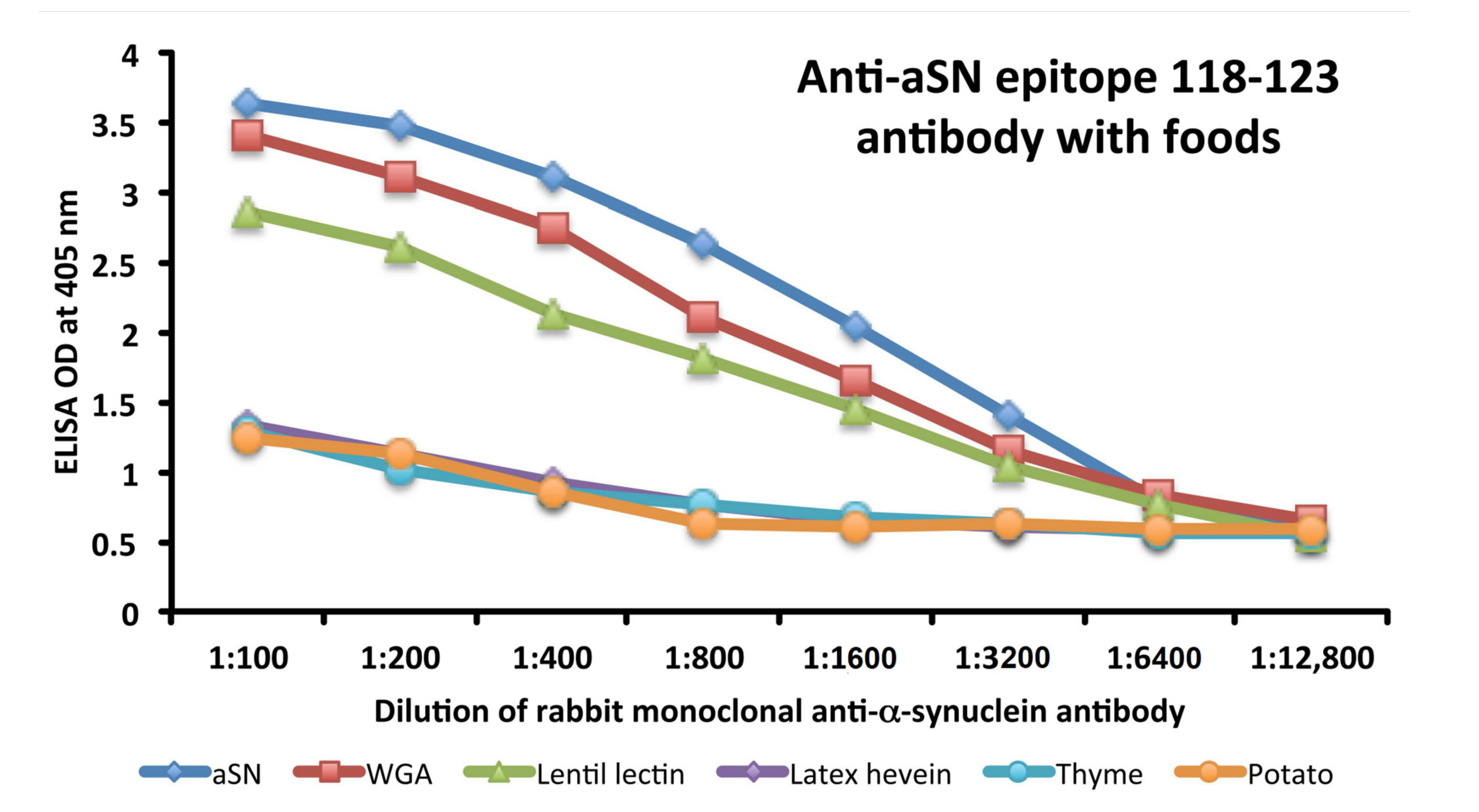
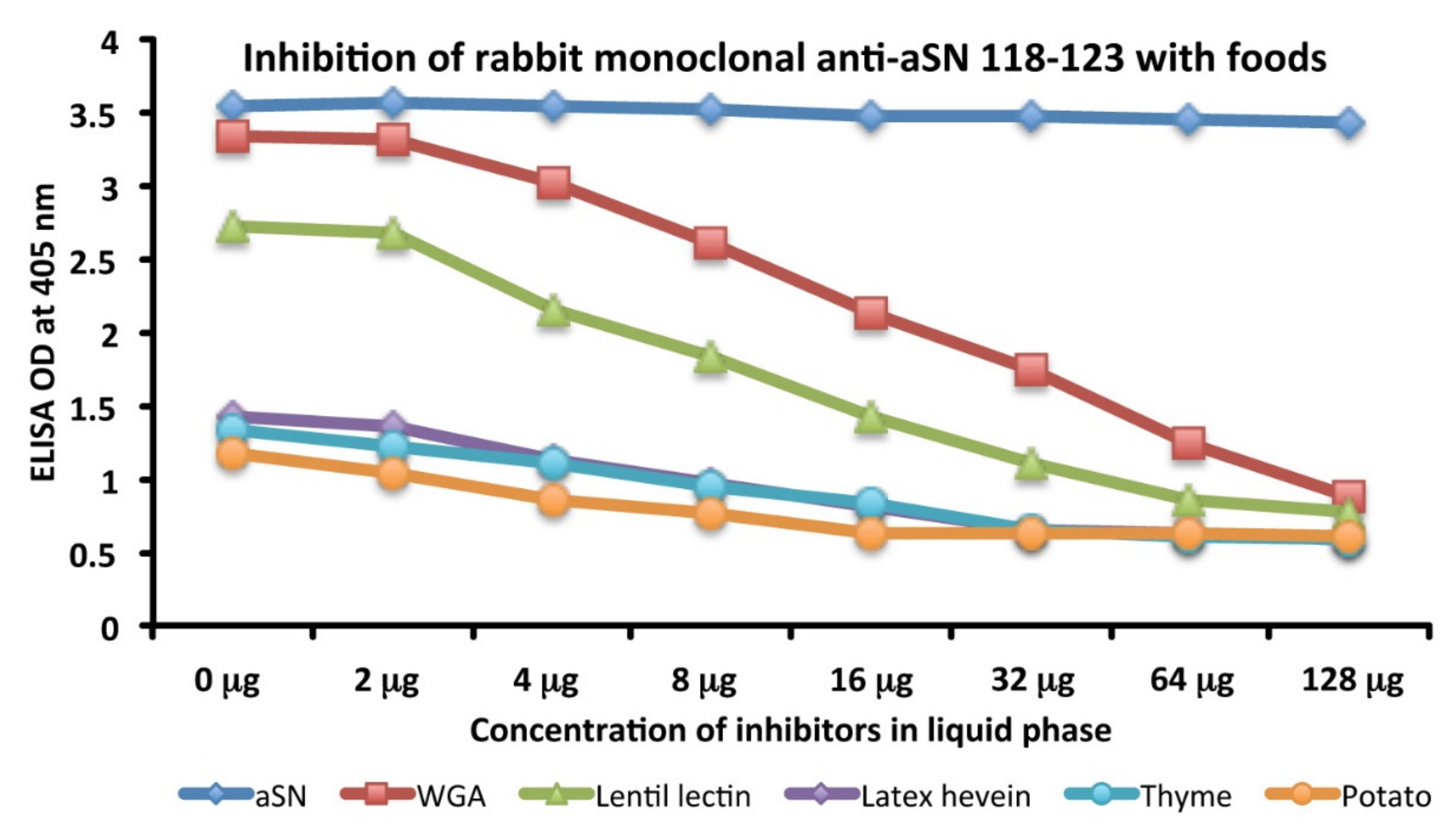
3.5. Analytical Specificity of the Rabbit Monoclonal Anti-aSN Epitope 118–123 Antibody Binding to Selected Food Antigens Performed by Serial Inhibitions
3.6. Sequence Similarity Between Alpha-Synuclein and Food Antigens
4. Discussion
- The presence of antibodies in the blood that cross-react with aSN may interfere with the accurate measurement of aSN antibodies used in support of PD diagnosis and progression assessment of the disease.
- Compromised BBB, which is often observed in PD patients, may result in the entry of food cross-reactive antibodies into the central nervous system, where their binding to aSN can further contribute to Lewy body pathology.
- The presence in the blood of food antigens that cross-react to aSN may interfere with treatment modalities in which anti-aSN antibodies are used to target misfolded proteins in PD.
- High cross-reactivity of a food-specific antibody with aSN may affect some physiological functions of aSN, thus putting individuals at risk for synucleinopathy.
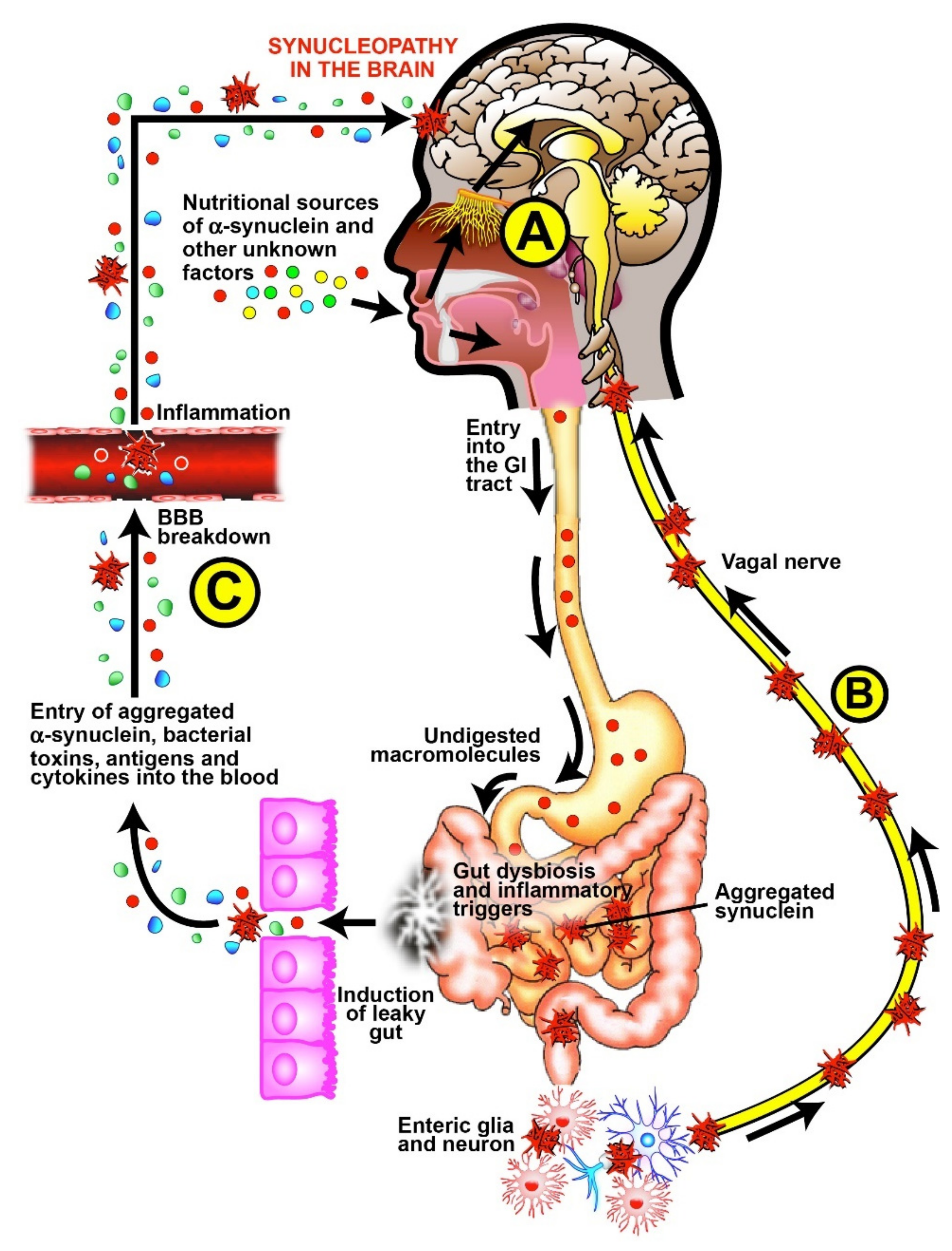
- Glutamate neurotransmission and glutamate receptors play a pivotal role in brain physiology and are altered in PD [96,97]. On the other hand, the 33-mer peptide (p57–89) from the a2-gliadin subtype is considered the supramolecule that induces celiac disease [98]. Intriguingly, the component of the NMDA glutamate receptor ion channel—the human GRINA protein has a significant sequence homology with the 33-mer gliadin peptide [99]. The authors suggested that 33-mer gliadin molecule is a natural antagonist interfering with the normal interactions of GRINA, thus impacting the extraintestinal manifestations of celiac disease. This is a pathophysiological example of how molecular similarity between a luminal gluten-originated nutrient might affect PD behavior through the human glutamate receptor GRINA protein. In addition, celiac disease has been associated with “leaky gut” syndrome, an increase in the permeability of the intestinal mucosa [100]. It has also been shown that dietary imbalance impairs BBB properties, potentially favoring transmigration and leading to neuroinflammation [101]. Figure 7 summarizes schematically the triple-hit Hypothesis of alpha-synucleopathy in the gut-brain axes of patients with Parkinson’s disease.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lerner, A.; Matthias, T. Mosaic of Autoimmunity, the Novel Factors of Autoimmune Diseases Revisited, 2nd ed.; Gluten and Autoimmunogenesis; Shoenfield, Y., Perricone, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 315–321. [Google Scholar]
- Lerner, A.; Matthias, T. Autoimmunity in celiac disease: Extra-intestinal manifestations. Autoimm. Rev. 2019, 18, 241–246. [Google Scholar]
- Lerner, A.; Matthias, T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun Rev. 2015, 14, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T. Don’t forget the exogenous microbial transglutaminases: It is immunogenic and potentially pathogenic. AIMS Biophysics 2016, 3, 546–552. [Google Scholar] [CrossRef]
- Lerner, A.; Shoenfeld, Y.; Matthias, T. A Review: Gluten ingestion side effects and withdrawal advantages in non-celiac autoimmune diseases. Nutritional Rev. 2017, 75, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Matthias, T. Gut—the Trojan horse in remote organs’ autoimmunity. J. Clin. Cell Immunol. 2016, 7, 401. [Google Scholar]
- Vojdani, A.; Gushgari, L.R.; Vojdani, E. Interaction between food antigens and the immune system: Association with autoimmune disorders. Autoimmun Rev. 2020, 19, 102459. [Google Scholar] [CrossRef]
- Gershteyn, I.M.; Ferreira, L.M.R. Immunodietica: A data-driven approach to investigate interactions between diet and autoimmune disorders. J. Transl Autoimmun. 2019, 1, 100003. [Google Scholar] [CrossRef]
- Hvatum, M.; Kanerud, L.; Hällgren, R.; Brandtzaeg, P. The gut-joint axis: Cross reactive food antibodies in rheumatoid arthritis. Gut 2006, 55, 1240–1247. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Matthias, T. Rheumatoid arthritis-celiac disease relationship: Joints get that gut feeling. Autoimm Rev. 2015, 14, 1038–1047. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. The gut feeling of the joints: Celiac disease and rheumatoid arthritis are related. Internat J. Celiac Dis. 2019, 7, 21–25. [Google Scholar]
- Borba, V.V.; Lerner, A.; Matthias, T.; Shoenfeld, Y. Bovine milk proteins as a trigger for autoimmune diseases: Myth or reality? Internat J. of Celiac Dis. 2020, 8, 4–15. [Google Scholar]
- Esposito, S.; Bonavita, S.; Sparaco, M.; Gallo, A.; Tedeschi, G. The role of diet in multiple sclerosis: A review. Rev. Nutr Neurosci. 2018, 21, 377–390. [Google Scholar] [CrossRef]
- Lerner, A.; Ramesh, A.; Matthias, T. Are non-celiac autoimmune diseases responsive to gluten-free diet? Internat J. Celiac Dis. 2017, 5, 164–167. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Going gluten free in non-celiac autoimmune diseases: The missing ingredient. Expert Rev. Clin. Immunol. 2018, 14, 873–875. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Berthelot, L.; Jeremias, P.; Abbad, L.; Matthias, T.; Monteiro, R.C. Gut-kidney axis: Gluten, transglutaminase, celiac disease and IgA nephropathy. J. Clin. Cell Immunol. 2017, 8, 499–503. [Google Scholar] [CrossRef] [Green Version]
- Choi, I.Y.; Lee, C.; Longo, V.D. Nutrition and fasting mimicking diets in the prevention and treatment of autoimmune diseases and immunosenescence. Mol. Cell Endocrinol. 2017, 455, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Holzbauer, S.M.; DeVries, A.S.; Sejvar, J.J.; Lees, C.H.; Adjemian, J.; McQuiston, J.H.; Medus, C.; Lexau, C.A.; Harris, J.R.; Sergio E Recuenco, S.E.; et al. Epidemiologic investigation of immune-mediated polyradiculoneuropathy among abattoir workers exposed to porcine brain. PLoS ONE 2010, 5, e9782. [Google Scholar] [CrossRef] [PubMed]
- Mohan, M.; Okeoma, C.M.; Sestak, K. Dietary Gluten and Neurodegeneration: A Case for Preclinical Studies. Int. J. Mol. Sci. 2020, 21, 5407. [Google Scholar] [CrossRef]
- Grant, W.B. Using Multicountry Ecological and Observational Studies to Determine Dietary Risk Factors for Alzheimer’s Disease. J. Am. Coll Nutr. 2016, 35, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Seidl, S.E.; Santiago, J.A.; Bilyk, H.; Potashkin, J.A. The emerging role of nutrition in Parkinson’s disease. Front. Aging Neurosci. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Maraki, M.I.; Yannakoulia, M.; Stamelou, M.; Stefanis, L.; Xiromerisiou, G.; Kosmidis, M.H.; Dardiotis, E.; Hadjigeorgiou, G.M.; Sakka, P.; Costas A Anastasiou, C.A.; et al. Mediterranean diet adherence is related to reduced probability of prodromal Parkinson’s disease. Mov. Disord. 2019, 34, 48–57. [Google Scholar] [CrossRef] [PubMed]
- McCarty, F.M.; Lerner, A. Low risk of Parkinson’s disease in quasi-vegan cultures may reflect GCN2-mediated upregulation of parkin. Adv. Nutr. 2020, 12, 355–362. [Google Scholar] [CrossRef]
- Riccio, P.; Rossano, R. Undigested food and gut microbiota may cooperate in the pathogenesis of neuroinflammatory diseases: A matter of barriers and a proposal on the origin of organ specificity. Nutrients 2019, 11, 2714. [Google Scholar] [CrossRef] [Green Version]
- Artur, M.A.S.; Zhao, T.; Ligterink, W.; Schranz, E.; Hilhorst, H.W.M. Dissecting the genomic diversification of late embryogenesis abundant (LEA) protein gene families in plants. Genome Biol. Evol. 2019, 11, 459–471. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, M.; Cheng, H.; Sun, N.; Simu Liu, S.; Li, S.; Wang, Y.; Zheng, Y.; Uversky, V.N. The effect of phosphorylation on the salt-tolerance-related functions of the soybean protein PM18, a member of the group-3 LEA protein family. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 1291–1303. [Google Scholar] [CrossRef]
- Liu, H.; Xing, M.; Yang, W.; XMu, X.; Wang, X.; Lu, F.; Wang, Y.; Zhang, L. Genome-wide Identification of and functional tnsights tnto the late embryogenesis abundant (LEA) gene family in bread wheat (Triticum aestivum). Sci. Rep. 2019, 9, 13375. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Kumar, D.; Kumar, S.; Rampuria, S.; Reddy, A.R.; Kirti, P.B. Ectopic expression of an atypical hydrophobic group 5 LEA protein from wild peanut Arachis diogoi confers abiotic stress tolerance in tobacco. PLoS ONE 2016, 11, e0150609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Li, X. Identification and phylogenetic analysis of late embryogenesis abundant proteins family in tomato (Solanum lycopersicum). Planta 2015, 241, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Meng, B.; Chen, W.; Ge, X.; Liu, S.; Yu, J. A proteomic study on post diapaused embryonic development of brine shrimp (Artemia francis-cana). Proteomics 2007, 7, 3580–3591. [Google Scholar] [CrossRef]
- George, J.M. The Synucleins. Genome Biol. 2002, 3. [Google Scholar] [CrossRef]
- Vaccaro, R.; Toni, M.; Casini, A.; Vivacqua, G.; Yu, S.; D’este, L.; Cioni, C. Localization of α- synuclein in teleost central nervous system: Immunohistochemical and Western blot evidence by 3D5 monoclonal antibody in the common carp, Cyprinus carpio. J. Comp. Neurol. 2015, 523, 1095–1124. [Google Scholar] [CrossRef] [Green Version]
- Tiunova, A.A.; Anokhin, K.V.; Saha, A.R.; Schmidt, O.; Hanger, D.P.; Anderton, B.H.; Davies, A.M.; Ninkina, N.N.; Buchman, V.L. Chicken synucleins: Cloning and expression in the developing embryo. Mech Dev. 2000, 99, 195–198. [Google Scholar] [CrossRef]
- Killinger, B.A.; Labrie, V. Vertebrate food products as a potential source of prion-like α-synuclein. NPJ Parkinsons Dis. 2017, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Zhao, Y. Evolutionary aspects of the synuclein super-family and sub-families based on large-scale phylogenetic and group-discrimination analysis. Biochem. Biophys. Res. Commun. 2013, 441, 308–317. [Google Scholar] [CrossRef]
- Barbour, R.; Kling, K.; Anderson, J.P.; Banducci, K.; Cole, T.; Diep, L.; Fox, M.; Goldstein, J.M.; Soriano, F.; Seubert, P.; et al. Red blood cells are the major source of alpha-synuclein in blood. Neurodegener. Dis. 2008, 5, 55–59. [Google Scholar] [CrossRef]
- Tian, C.; Liu, G.; Gao, L.; Soltys, D.; Pan, C.; Stewart, T.; Shi, M.; Xie, Z.; Liu, N.; Feng, T.; et al. Erythrocytic α-synuclein as a potential biomarker for Parkinson’s disease. Transl Neurodegener. 2019, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Freire, M.; Semba, R.D.; Ubaida-Mohien, C.; Fabbri, E.; Scalzo, P.; Højlund, K.; Dufresne, C.; Lyashkov, A.; Ferrucci, L. The human skeletal muscle proteome project: A reappraisal of the current literature. J. Cachexia Sarcopenia Muscle 2017, 8, 5–18. [Google Scholar] [CrossRef]
- Lerner, A.; Neidhöfer, S.; Matthias, T. The gut microbiome feelings of the brain: Perspective for Non-Microbiologists. Microorganisms 2017, 5, 66. [Google Scholar] [CrossRef]
- Vacchi, E.; Kaelin-Lang, A.; Melli, G. Tau and alpha synuclein synergistic effect in neurodegenerative diseases: When the periphery is the core. Int. J. Mol. Sci. 2020, 21, 5030. [Google Scholar] [CrossRef]
- Kujawska, M.; Jodynis-Liebert, J. What is the evidence that Parkinson’s disease is a prion disorder, which originates in the gut? Int. J. Mol. Sci. 2018, 19, 3573. [Google Scholar] [CrossRef] [Green Version]
- Boulos, C.; Yaghi, N.; Hayeck, R.E.; Heraoui, G.N.; Fakhoury-Sayegh, N. Nutritional risk factors, microbiota and Parkinson’s disease: What Is the current evidence? Nutrients 2019, 11, 1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A. The intestinal luminal sources of a-synuclein: A gastroenterologist perspective. Nutr. Rev. 2021. [Google Scholar] [CrossRef]
- Kharrazian, D.; Herbert, M.; Vojdani, A. Detection of islet cell immune reactivity with low glycemic index foods: Is this a concern for type 1 diabetes? J. Diabetes Res. 2017, 2017, 4124967. [Google Scholar] [CrossRef] [Green Version]
- Vojdani, A.; Turnpaugh, C.C. Antibodies against Group A Streptococcus, dopamine receptors, and ganglioside GM1 cross-react with a variety of food antigens, potentially interfering with biomarkers for PANS and PANDAS. Biomark. Neuropsychiatry 2020, 3, 100023. [Google Scholar] [CrossRef]
- Vojdani, A.; O’Bryan, T.; Green, J.A.; McCandless, J.; Woeller, K.N.; Vojdani, E.; Nourian, A.A.; Cooper, E.L. Immune response to dietary proteins, gliadin and cerebellar peptide in children with autism. Nutr. Neurosci. 2004, 7, 151–161. [Google Scholar] [CrossRef] [Green Version]
- Alaedini, A.; Okamoto, H.; Briani, C.; Wollenberg, K.; Shill, H.A.; Bushara, K.O.; Sander, H.W.; Green, P.H.R.; Hallett, M.; Latov, M. Immune cross-reactivity in celiac disease: Anti-gliadin antibodies bind to neuronal synapsin I. J. Immunol. 2007, 178, 6590–6595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vojdani, A.; Tarash, I. Cross-reaction between gliadin and different food and tissue antigens. Food Nutr. Sci. 2013, 4, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Vojdani, A.; Vojdani, E. Immunoreactivity of anti-AbP-42 specific antibody with toxic chemical and food antigens. J. Alzheimers Dis. Parkinsonism 2018, 8, 441. [Google Scholar] [CrossRef]
- McGlinchey, R.P.; Lacy, S.M.; Huffer, K.E.; Tayebi, N.; Sidransky, E.; Lee, J.C. C-terminal α-synuclein truncations are linked to cysteine cathepsin activity in Parkinson’s disease. J. Biol. Chem. 2019, 294, 9973–9984. [Google Scholar] [CrossRef] [Green Version]
- Ulusoy, A.; Febbraro, F.; Jensen, P.H.; Kirik, D.; Romero-Ramos, M. Co-expression of C-terminal truncated α-synuclein enhances full-length α-synuclein-induced pathology. Eur. J. Neurosci. 2010, 32, 409–422. [Google Scholar] [CrossRef]
- van der Wateren, I.M.; Knowles, T.P.J.; Buell, A.K.; Dobson, C.M.; Galvagnion, C. C-terminal truncation of α-synuclein promotes amyloid fibril amplification at physiological pH. Chem. Sci. 2018, 9, 5506–5516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levitan, K.; Chereau, D.; Cohen, S.I.A.; Knowles, T.P.J.; Dobson, C.M.; Fink, A.L.; Anderson, J.P.; Goldstein, J.M.; Millhauser, G.L. Conserved C-terminal charge exerts a profound influence on the aggregation rate of α-synuclein. J. Mol. Biol. 2011, 411, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Braak, H.; Rub, U.; Gai, P.; Del Tredici, K. Idiopathic Parkinson’s disease: Possible routes by which invulnerable neuronal types may be subject to neuroinvasion by an unknown pathogen. J. Neural Transm. 2003, 110, 517–536. [Google Scholar] [CrossRef]
- Hawkes, C.H.; Del Tredici, K.; Braak, H. Parkinson’s disease: A dual-hit hypothesis. Neuropathol. Appl. Neurobiol. 2007, 33, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Del Tredici, K.; Braak, H. Review: Sporadic Parkinson’s disease: Development and distribution of α-synuclein pathology. Neuropathol. Appl. Neurobiol. 2016, 42, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Spiegel, J.; Dillmann, U.; Grundmann, D.; Bürmann, J.; Faßbender, K.; Schäfer, K.H.; Unger, M.M. Fecal markers of intestinal inflammation and intestinal permeability are elevated in Parkinson’s disease. Parkinsonism Relat. Disord. 2018, 50, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Vojdani, A. Lectins, agglutinins, and their role in autoimmune reactivities. Altern. Ther. Health Med. 2015, 21, 46–51. [Google Scholar]
- Vojdani, A. Reaction of food-specific antibodies with different tissue antigens. Int. J. Food Sci Tech. 2020, 55, 1800–1815. [Google Scholar] [CrossRef] [Green Version]
- De Virgilio, A.; Greco, A.; Fabbrini, G.; Inghilleri, M.; Rizzo, M.I.; Gallo, A.; Conte, M.; Rosato, C.; Ciniglio Appiani, M.; de Vincentiis, M. Parkinson’s disease: Autoimmunity and neuroinflammation. Autoimmun. Rev. 2016, 15, 1005–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garretti, F.; Agalliu, D.; Lindestam Arlehamn, C.S.; Sette, A.; Sulzer, D. Autoimmunity in Parkinson’s disease: The role of α-synuclein-specific T cells. Front. Immunol. 2019, 10, 303. [Google Scholar] [CrossRef] [Green Version]
- Lundmark, K.; Westermark, G.T.; Olsén, A.; Westermark, P. Protein fibrils in nature can enhance amyloid protein A amyloidosis in mice: Cross-seeding as a disease mechanism. Proc. Natl. Acad. Sci. USA. 2005, 102, 6098–6102. [Google Scholar] [CrossRef] [Green Version]
- Miraglia, F.; Colla, E. Microbiome, Parkinson’s Disease and molecular mimicry. Cells 2019, 8, 222. [Google Scholar] [CrossRef] [Green Version]
- Okubo, H.; Miyake, Y.; Sasaki, S.; Murakami, K.; Tanaka, K.; Fukushima, W.; Kiyohara, C.; Tsuboi, Y.; Yamada, T.; Oeda, T.; et al. Dietary patterns and risk of Parkinson’s disease: A case-control study in Japan. Eur. J. Neurol. 2012, 19, 681–688. [Google Scholar] [CrossRef]
- Coimbra, C.G.; Junqueira, V.B.C. High doses of riboflavin and the elimination of dietary red meat promote the recovery of some motor functions in Parkinson’s disease patients. Braz. J. Med. Biol. Res. 2003, 36, 1409–1417. [Google Scholar] [CrossRef] [Green Version]
- Benvenga, S.; Guarneri, F. Molecular mimicry and autoimmune thyroid disease. Rev. Endocr. Metab. Disord. 2016, 17, 485–498. [Google Scholar] [CrossRef]
- Lyons-Weiler, J. Pathogenic priming likely contributes to serious and critical illness and mortality in COVID-19 via autoimmunity. J. Transl. Autoimm. 2020, 3, 100051. [Google Scholar] [CrossRef]
- Chandra, R.; Hiniker, A.; Kuo, Y.M.; Nussbaum, R.L.; Liddle, R.A. α-Synuclein in gut endocrine cells and its implications for Parkinson’s disease. JCI Insight. 2017, 2, e92295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seguella, L.; Capuano, R.; Sarnelli, G.; Esposito, G. Play in advance against neurodegeneration: Exploring enteric glial cells in gut-brain axis during neurodegenerative diseases. Expert Rev. Clin. Pharmacol. 2019, 12, 555–564. [Google Scholar] [CrossRef]
- Vergnolle, N.; Cirillo, C. Neurons and glia in the enteric nervous system and epithelial barrier function. Physiology (Bethesda) 2018, 33, 269–280. [Google Scholar] [CrossRef]
- Cirillo, C.; Sarnelli, G.; Esposito, G.; Turco, F.; Steardo, L.; Cuomo, R. S100B protein in the gut: The evidence for enteroglial-sustained intestinal inflammation. World J. Gastroenterol. 2011, 17, 1261–1266. [Google Scholar] [CrossRef]
- Fitzgerald, E.; Murphy, S.; Martinson, H.A. Alpha-synuclein pathology and the role of microbiota in Parkinson’s disease. Front. Neurosci. 2019, 13, 369. [Google Scholar] [CrossRef]
- Khalil, M.; Zhang, Z.; Engel, M.A. Neuro-immune networks in gastrointestinal disorders. Visc. Med. 2019, 35, 52–60. [Google Scholar] [CrossRef]
- Barrett, P.J.; Greenamyre, J.T. Post-translational Modification of α-synuclein in Parkinson’s disease. Brain Res. 2015, 1628, 247–253. [Google Scholar] [CrossRef]
- Oueslati, A.; Fournier, M.; Lashuel, H.A. Role of post-translational modifications in modulating the structure, function and toxicity of alpha-synuclein: Implications for Parkinson’s disease pathogenesis and therapies. Prog. Brain Res. 2010, 183, 115–145. [Google Scholar] [CrossRef]
- González, N.; Arcos-López, T.; König, A.; Quintanar, L.; Menacho Márquez, M.; Outeiro, T.F.; Fernández, C.O. Effects of alpha-synuclein post-translational modifications on metal binding. J. Neurochem. 2019, 150, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Aminov, R.; Matthias, T. Dysbiosis may trigger autoimmune diseases via inappropriate posttranslational modification of host proteins. Front. Microbiol. 2016, 7, 84. [Google Scholar] [CrossRef]
- Anselmi, L.; Bove, C.; Coleman, H.; Le, K.; Subramanian, M.P.; Venkiteswaran, K.; Subramanian, T.; Travagli, R.A. Ingestion of subthreshold doses of environmental toxins induces ascending Parkinsonism in the rats. NPJ Parkinson’s Dis. 2018, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Sampson, T.R.; Challis, C.; Jain, N.; Moiseyenko, A.; Ladinsky, M.S.; Shastri, G.G.; Thron, T.; Needham, B.D.; Horvath, I.; Debelius, J.W.; et al. A gut bacterial amyloid promotes α-synuclein aggregation and motor impairment in mice. Elife 2020, 9, e53111. [Google Scholar] [CrossRef]
- Batista, C.R.A.; Gomes, G.F.; Candelario-Jalil, E.; Fiebich, B.L.; de Oliveira, A.C.P. Lipopolysaccharide-induced neuroinflammation as a bridge to understand neurodegeneration. Int. J. Mol. Sci. 2019, 20, 2293. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Lv, G.; Lee, J.S.; Jung, B.C.; Masuda-Suzukake, M.; Hong, C.-S.; Elvira Valera, E.; Lee, H.-J.; Paik, S.R.; Hasegawa, M.; et al. Exposure to bacterial endotoxin generates a distinct strain of α-synuclein fibril. Sci. Rep. 2016, 6, 30891. [Google Scholar] [CrossRef] [Green Version]
- Barbut, D.; Stolzenberg, E.; Zasloff, M. Gastrointestinal immunity and alpha-synuclein. J. Parkinson’s Dis. 2019, 9, S313–S322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kwon, S.H.; Kam, T.-I.; Panicker, N.; Karuppgounder, S.S.; Lee, S.; Lee, J.H.; Kim, W.R.; Kook, M.; Foss, C.A.; et al. Transneuronal propagation of pathologic α-synuclein from the gut to the brain models Parkinson’s disease. Neuron 2019, 103, 627–641. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased intestinal permeability correlates with sigmoid mucosa alpha-synuclein staining and endotoxin exposure markers in early Parkinson’s disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef] [Green Version]
- Mulak, A.; Koszewicz, M.; Panek-Jeziorna, M.; Koziorowska-Gawron, E.; Budrewicz, S. Fecal calprotectin as a marker of the gut immune system activation is elevated in Parkinson’s disease. Front. Neurosci. 2019, 13, 992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challis, C.; Hori, A.; Sampson, T.R.; Yoo, B.B.; Challis, R.C.; Hamilton, A.M.; Mazmanian, K.M.; Volpicelli-Daley, L.A.; Gradinaru, V. Gut-seeded α-synuclein fibrils promote gut dysfunction and brain pathology specifically in aged mice. Nat. Neurosci. 2020, 23, 327–336. [Google Scholar] [CrossRef] [PubMed]
- van IJzendoorn, S.C.D.; Derkinderen, P. The intestinal barrier in Parkinson’s disease: Current state of knowledge. J. Parkinson’s Dis. 2019, 9, S323–S329. [Google Scholar] [CrossRef] [Green Version]
- Kelly, L.P.; Carvey, P.M.; Keshavarzian, A.; Shannon, K.M.; Shaikh, M.; Bakay, R.A.E.; Kordower, J.H. Progression of intestinal permeability changes and alpha-synuclein expression in a mouse model of Parkinson’s disease. Mov. Disord. 2014, 29, 999–1009. [Google Scholar] [CrossRef]
- Maes, M.; Sirivichayakul, S.; Kanchanatawan, B.; Vojdani, A. Upregulation of the intestinal paracellular pathway with breakdown of tight and adherens junctions in deficit schizophrenia. Mol. Neurobiol. 2019, 56, 7056–7073. [Google Scholar] [CrossRef]
- Maes, M.; Sirivichayakul, S.; Kanchanatawan, B.; Vojdani, A. Breakdown of the paracellular tight and adherens junctions in the gut and blood brain barrier and damage to the vascular barrier in patients with deficit schizophrenia. Neurotox. Res. 2019, 36, 306–322. [Google Scholar] [CrossRef]
- Nagele, R.G.; Clifford, P.M.; Siu, G.; Levin, E.C.; Acharya, N.K.; Han, M.; Kosciuk, M.C.; Venkataraman, V.; Zavareh, S.; Zarrabi, S.; et al. Brain reactive autoantibodies prevalent in human sera increase intraneuronal amyloid-β (1–42) deposition. J. Alzheimers Dis. 2011, 25, 605–622. [Google Scholar] [CrossRef] [Green Version]
- Houser, M.C.; Tansey, M.G. The gut-brain axis: Is intestinal inflammation a silent driver of Parkinson’s disease pathogenesis? NPJ Parkinson’s Dis. 2017, 3, 3. [Google Scholar] [CrossRef]
- Klingelhoefer, L.; Reichmann, H. Pathogenesis of Parkinson disease—the gut-brain axis and environmental factors. Nat. Rev. Neurol. 2015, 11, 625–636. [Google Scholar] [CrossRef]
- Fields, C.R.; Bengoa-Vergniory, N.; Wade-Martins, R. Targeting alpha-synuclein as a therapy for Parkinson’s disease. Front. Mol. Neurosci. 2019, 12, 299. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.T.; Chung, C.H.-Y.; Iba, M.; Zhang, B.; Trojanowski, J.Q.; Luk, K.C.; Lee, V.M.Y. α-Synuclein immunotherapy blocks uptake and template propagation of misfolded α-synuclein and neurodegeneration. Cell Rep. 2014, 7, 2054–2065. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, S.; Fu, P.; Zhang, Z.; Lin, K.; Ko, J.K.-S.; Yung, K.K.-L. Roles of Glutamate Receptors in Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 4391. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-González, V.; Ogalla-García, E.; García-Quintanilla, M.; García-Quintanilla, A. Deciphering GRINA/Lifeguard1: Nuclear Location, Ca2+ Homeostasis and Vesicle Transport. Int. J. Mol. Sci. 2019, 20, 4005. [Google Scholar] [CrossRef] [Green Version]
- Baron, M.V.; Troncone, R.; Auricchio, S. Gliadin peptides as triggers of the proliferative and stress/innate immune response of the celiac small intestinal mucosa. Int. J. Mol. Sci. 2014, 15, 20518–20537. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Quintanilla, A.; Miranzo-Navarro, D. Extraintestinal manifestations of celiac disease: 33-mer gliadin binding to glutamate receptor GRINA as a new explanation. Bioessays 2016, 38, 427–439. [Google Scholar] [CrossRef]
- Obrenovich, M.E.M. Leaky gut, leaky brain? Microorganisms 2018, 6, 107. [Google Scholar] [CrossRef] [Green Version]
- Costa de Aquino, C.; Leitão, R.A.; Oliveira Alves, L.A.; Coelho-Santos, V.; Guerrant, R.L.; Ribeiro, C.F.; Malva, J.O.; Silva, A.P.; Oriá, R.B. Effect of hypoproteic and high-fat diets on hippocampal blood-brain barrier permeability and oxidative stress. Front. Nutr. 2018, 5, 131. [Google Scholar] [CrossRef]
- Lerner, A.; Benzvi, C. “Let Food Be Thy Medicine”: Gluten and Potential Role in Neurodegeneration. Cells 2021, 10, 756. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yeast Protein | Yeast Sequence | Mapped Start To End | Alpha-Synuclein Sequence | ID (%) |
|---|---|---|---|---|
| Structure of the S. cerevisiae Srs2 C-terminal domain in complex with PCNA conjugated to SUMO | MDIFSQ-LSRAK | 45–55 | MDVFMKGLSKAK | 58 |
| Proteasome Activator Complex | KATNGVVIATE | 19–29 | KAKEGVVAAAE * | 64 |
| Structure of the ribosomal 80S-eEF2-sordarin complex | FMKKLRAAK---LAAPE | 10–23 | FMKGLSKAKEGVVAAAE * | 53 |
| Crystal Structure of the S. cerevisiae glucokinase, Glk1 | MEKGLAPPKEG | 43–53 | M-KGLSKAKEG | 64 |
| Solution structure of the oxidized form of the Yap1 redox domain | MAKAKCSERGVVINAE | 60–75 | LSKAK--E-GVVAAAE | 56 |
| Cryo-EM Structure of the RIX1-REA1 Pre-60S Particle | KG-SKAK-GVNMAA | 471–482 | KGLSKAKEGV-VAA | 71 |
| Crystal structure of yeast elongation factor 2 in complex with sordarin | SKAGEIVLAA | 813–822 | SKAKEGVVAA * | 70 |
| Solution structure of the SBDbeta domain of yeast Ssa1 | KGRLSKEDIEKMVAEAE | 123–139 | KG-LSKAK-EGVVAAAE | 59 |
| Crystal structure of the RNA recognition motif of yeast eIF3b residues 76–161 | EATGKTK-GFLFV | 48–59 | EAAGKTKEGVLYV * | 69 |
| Structure of the ribosomal 80S-eEF2-sordarin complex from yeast | TKEAAAVAEGKSKQ | 9–22 | TKQGVAEAAGKTKE * | 50 |
| Crystal structure of a Prp8 C-terminal fragment | KQNDEEAAGASTVMKTK | 276–292 | KQGVAEAAG-----KTK * | 53 |
| Crystal Structures of Yeast Transketolase In Complex with Analogs of Thiamin Diphosphate | TPEGVAERAQKT | 650–661 | TKQGVAEAAGKT * | 67 |
| Crystal Structure of Yeast Aspartyl-Trna Synthetase Complexed with Trna Asp | KEGI-EMLRAAGK | 336–347 | KQGVAE---AAGK | 54 |
| A Gated Channel into The Proteasome Core Particle | TKEGVVLGV----EK | 42–52 | TKEGVVHGVATVAEK * | 67 |
| Structure of The Yeast Cytochrome Bc1 Complex Co-Crystallized with An Antibody Fv-Fragment | GSRYATKDGVAH | 22–33 | GSK—TKEGVVH * | 58 |
| Electron cryomicroscopy structure of S. cerevisiae FAS in the Apo state | SKTIKD-LVGGKSTV | 169–182 | SKT-KEGVVHGVATV * | 53 |
| Structure of the Ndi1 protein in complex with the competitive inhibitor, stigmatellin | VHLRTAVAKVEEK | 271–283 | VH---GVATVAEK * | 54 |
| Cryo-EM Structure of the Exocyst Complex | QVNSIGGVVVT | 781–791 | QVTNVGGAVVT | 64 |
| Arx1 pre-60S particle | NVGGALRVPG--AISEK | 44–58 | NVGGAV-VTGVTAVAQK | 53 |
| Crystallographic snapshots of eukaryotic dimethylallyltransferase acting on tRNA | IAGTTG-VGKSQL | 7–18 | IAAATGFVKKDQL * | 62 |
| Crystal structure of yeast aminopeptidase 1 (Ape1) | TAEGYGRIAVA | 104–114 | TVEGAGSIAAA * | 64 |
| Yeast RNA polymerase III initial transcribing complex | DNEDNE-GSEE-----EPE | 579–591 | DNEAYEMPSEEGYQDYEPE * | 53 |
| Soybean Agglutinin Protein | Soybean Agglutinin Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
|---|---|---|---|---|
| Uncharacterized protein LOC100790514 [Glycine max] | MNGLSK---GVVATA | 178–189 | MKGLSKAKEGVVAAA | 67 |
| Uncharacterized protein LOC100799919 [Glycine max] | GL-KTKEGVVLAVE | 39–51 | GLSKAKEGVVAAAE | 71 |
| Uncharacterized protein LOC100793479 isoform X1 [Glycine max] | FSTGLSKRTSKVAEKKEGTVAGA | 79–101 | FMKGLSK-----A--KEGVVAAA | 52 |
| Coleoptile phototropism protein 1 [Glycine max] | MDYFVKTLSGIKAK-GV | 57–72 | MDVFMKGLS--KAKEGV | 65 |
| Proteinaceous RNase P 2 [Glycine max] | VGEAEFDAGRVKEGVL | 219–234 | VAEA---AGKTKEGVL | 63 |
| Uncharacterized protein At4g13230 [Glycine max] | KTKES-AEHAKDNVVGKTKESAEYV | 121–144 | KTKQGVAEAA-----GKTKEGVLYV | 56 |
| Soyasapogenol B glucuronide galactosyltransferase-like [Glycine max] | KT-QG--EEEGWLEWLNKKKEGSVLYV | 268–291 | KTKQGVAEAAG------KTKEG-VLYV | 52 |
| Peptidyl-prolyl cis-trans isomerase FKBP62 [Glycine max] | EAAGKKKEEGNVLF | 389–402 | EAAGKTKE--GVLY | 64 |
| 51 kDa seed maturation protein isoform X1 [Glycine max] | VGDAAQKTKE---YV | 164–175 | VAEAAGKTKEGVLYV | 60 |
| Binding partner of ACD11 1 [Glycine max] | KTKEKVLAQDNQGKTEEG | 228–245 | KTKQGV-AEAA-GKTKEG | 56 |
| ATP-dependent DNA helicase Q-like 3 isoform X1 [Glycine max] | SKIK--VV--VATVA | 307–317 | SKTKEGVVHGVATVA | 67 |
| Hypothetical protein GLYMA_15G213400 [Glycine max] | GVVQHGPGGTMATVAE | 1008–1023 | GVV-HG----VATVAE | 63 |
| Proteasome subunit alpha type-5 [Glycine max] | KTKEGVVLAV----EK | 41–51 | KTKEGVVHGVATVAEK | 63 |
| Subtilisin-like protease SBT1.2 [Glycine max] | VTNVGEANSSYVVT-VSA | 692–708 | VTNVGGA----VVTGVTA | 61 |
| AT-hook motif nuclear-localized protein 23 [Glycine max] | VTNVSLRQPASAGAVVT | 112–128 | VTNVG-------GAVVT | 53 |
| NAD-dependent malic enzyme 59 kDa isoform, mitochondrial [Glycine max] | VTAEVGAAVV--CAAVAEK | 543–559 | VT-NVGGAVVTGVTAVAQK | 58 |
| Subtilisin-like protease SBT5.3 [Glycine max] | VTNVGKARSIYKAVVVSPTGVNVTVV | 679–704 | VTNVG------GAVV---TG--VTAV | 50 |
| Pyruvate decarboxylase 2 [Glycine max] | VE---AIQTATG-VKKD | 555–567 | VEGAGSIAAATGFVKKD | 59 |
| 2-oxoglutarate dehydrogenase, mitochondrial [Glycine max] | SAATATGFLKVHQKEQ | 993–1008 | SIAAATGFVK---KDQ | 56 |
| Pyruvate decarboxylase 2 [Glycine max] | VE---AIATATG-PKKDSL | 560–574 | VEGAGSIAAATGFVKKDQL | 58 |
| Autophagy-related protein 18f isoform X2 [Glycine max] | TVSGAA--AAATG--RKNAL | 572–587 | TVEGAGSIAAATGFVKKDQL | 55 |
| Latex Hevein Protein | Latex Hevein Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
|---|---|---|---|---|
| Hypothetical protein GH714_008467 | LSKAKGGVVAEEE | 28–40 | LSKAKEGVVAAAE | 77 |
| Hypothetical protein GH714_000288 | KGVLSRKAKEAAVAAAQ | 176–192 | KG-LS-KAKEGVVAAAE * | 71 |
| Hypothetical protein GH714_000288 | KENVVEA | 587–593 | KEGVVAA * | 71 |
| Uncharacterized protein LOC110639732 | LSLKAHDGGVVA | 257–268 | LS-KAKEG-VVA | 67 |
| Lysine-specific histone demethylase 1 homolog 1-like | MKG-----DGVVAAAD | 288–298 | MKGLSKAKEGVVAAAE * | 56 |
| Proteasome subunit alpha type-5 | GL-KTKEGVVLAVE | 39–51 | GLSKAKEGVVAAAE | 71 |
| Hypothetical protein GH714_036586 | LDVFMKGI--AAEG | 223–234 | MDVFMKGLSKAKEG | 64 |
| Hypothetical protein GH714_025383 | DVFMKGI--AAEG | 282–292 | DVFMKGLSKAKEG * | 69 |
| Hypothetical protein GH714_028032 | DVFTKSLSQAK | 237–247 | DVFMKGLSKAK * | 73 |
| ABC transporter B family member 2-like | KEGASEGEVVEAA | 1096–1108 | KEG-----VVAAA | 54 |
| E3 ubiquitin-protein ligase CIP8 | YELPTDD--QDYE | 288–298 | YEMPSEEGYQDYE | 54 |
| The article Zinc finger protein BRUTUS-like At1g18910 | EAYEMPYASE | 583–592 | EAYEMP—SE * | 80 |
| DEAD-box ATP-dependent RNA helicase 56-like isoform X1 | DNDAYE---EE-LLDYEEE | 6–20 | DNEAYEMPSEEGYQDYEPE * | 58 |
| Hypothetical protein GH714_022075 [Hevea brasiliensis] | EAYESSSEESFKD | 180–192 | EAYEMPSEEGYQD | 62 |
| Hypothetical protein GH714_005670 | DNERFDF-NEEPYQQYE | 231–246 | DNEAYEMPSEEGYQDYE | 53 |
| Splicing factor U2af large subunit B-like isoform X4 | YE---EEGYQGNGEDFE | 4–17 | YEMPSEEGYQ----DYE | 53 |
| Wheat Germ Agglutinin Protein | Wheat Germ Agglutinin Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
|---|---|---|---|---|
| Chain A, Glutaredoxin | LAKAKE-IVASA | 53–63 | LSKAKEGVVAAA | 67 |
| Chain Y, 60s Ribosomal Protein L24 | KAKGKFTAEDV-AAA | 129–142 | KAK-----EGVVAAA | 53 |
| Chain I, 60s Ribosomal Protein L16 | DVGMK-----KKGV | 35–43 | DVFMKGLSKAKEGV | 50 |
| Chain F, 60s Ribosomal Protein L30 | KQKVAAEKIKAAENTK--VIY | 29–47 | KQGV-AE---AAGKTKEGVLY | 52 |
| Chain u, 60s Ribosomal Protein P1 | GGAAAAEEKKE | 80–90 | GVAEAAGKTKE | 55 |
| Chain A, Xylanase Inhibitor Protein I | VHPKNVYYGVAPVAQK | 231–246 | VH------GVATVAEK | 50 |
| Chain H, 60s Ribosomal Protein L6 | EGVTVQ-VAAKVVTV | 14–27 | EGV-VHGVA----TV | 53 |
| Chain M, 40s Ribosomal Protein S12e | VHLV-TVPSAKT | 114–124 | VHGVATV-AEKT | 58 |
| Chain X, Xylanase Inhibitor | GSKPVSKVNV--GV | 109–120 | GSK—TKEGVVHGV * | 50 |
| Chain A, Eukaryotic Translation Initiation Factor 4e-1 | GAVV----SVRQK | 114–122 | GAVVTGVTAVAQK | 54 |
| Chain A, Ribulose bisphosphate carboxylase large chain | EEAGAAVAAESSTG | 51–64 | EGAGS-IAA--ATG | 50 |
| Chain J, 60s Ribosomal Protein L5 | GIQEHIDLGMKYDP | 107–120 | GILE--D--MPVDP | 50 |
| Pea Lectin Protein | Pea Lectin Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
| Chain A, Ferredoxin—nadp reductase, leaf isozyme, chloroplastic | VYMMGL-KGME | 263–272 | VFMKGLSKAKE* | 55 |
| Chain A, Putative aminoaldehyde dehydrogenase | KEDVDVAVAAA | 44–54 | KEGV---VAAA | 64 |
| Chain B, Photosystem II CP47 reaction center protein | EGVAAA | 94–99 | EGVVAA | 83 |
| Chain X, Photosystem II reaction center protein X | KASLKEKVVTGLTAAA | 16–31 | KA--KEGVV----AAA | 56 |
| Chain A, Dihydrolipoamide dehydrogenase | KAEEDGVA | 327–334 | KAKEGVVA | 63 |
| Chain A, Dihydrolipoamide dehydrogenase | FTSGLNLDKIGV | 280–291 | FMKGLSKAKEGV | 50 |
| Chain A, Protein (ferredoxin: nadp + reductase) | VYMCGL-KGME | 263–272 | VFMKGLSKAKE * | 55 |
| Concanavalin-A Protein | Concanavalin-A Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
| Chain A, Alpha-mannosidase | KAYEGEV | 518–524 | KAKEGVV | 71 |
| Chain A, Alpha-mannosidase | VENVLDSVV | 48–56 | VTNVGGAVV | 56 |
| Chain A, Urease | GALSIA----FVSKAALDQ | 758–772 | GAGSIAAATGFVKK---DQ * | 53 |
| Chain A, Refined crystal structure of cavavalin from jack bean | EE---QEGVIVKMP | 168–178 | EEGAPQEGILEDMP * | 50 |
| Chain A, Urease | KEEEDA-SEGV | 24–33 | KNEEGAPQEGI | 55 |
| Chain A, Alpha-mannosidase | YE--SSEGDFSDYQ | 609–620 | YEMPSEEG-YQDYE * | 50 |
| Lentil Lectin Protein | Lentil Lectin Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
| Solution structure of Lipid Transfer Protein | AGSITKLNTNNAAA | 56–69 | AGSI-------AAA * | 50 |
| Potato Protein | Potato Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
|---|---|---|---|---|
| The structure of a glutathione synthetase (StGSS1) from Solanum tuberosum in ADP and y-EC bound closed conformation | SSSNEGGVAA | 463–472 | SKAKEGVVAA | 60 |
| Crystal structure of S-adenosylmethionine decarboxylase | KGLRSLSKA | 35–43 | KGL---SKA | 67 |
| Crystal structure of potato Rx-CC domain in complex with RanGAP2-WPP domain | LSKE-E---AAKNAE | 36–46 | LSKAKEGVVAA--AE | 53 |
| Structure determination and refinement at 1.8 A resolution of Disproportionating Enzyme | EGAVSSVARIA | 458–468 | EGVVHGVATVA * | 55 |
| The structure of a glutathione synthetase (StGSS1) from Solanum tuberosum in ADP and y-EC bound closed conformation | GVDMVH--APVA | 54–63 | GV--VHGVATVA | 58 |
| Characterization of Solanum tuberosum Multicystatin and Significance of Core Domains | AATDGG--KK | 60–67 | AAT--GFVKK | 60 |
| >Crystal Structure of Potato Multicystatin | >AATDDAG--KK | >55–63 | >AAT---GFVKK | >55 |
| Peanut Protein | Peanut Protein Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
|---|---|---|---|---|
| Chain A, PR 10 protein | GLFRAIEGYVLA | 140–151 | GLSKAKEG-VVA * | 58 |
| Chain A, Stilbene synthase, | AGLKTTGEGLDWGVLF | 357–372 | AG-KTK-E----GVLY | 50 |
| Chain A, PR 10 protein, | VVGGVALPPT-AEKITFETK | 83–101 | VVHGVA---TVAEK----TK * | 55 |
| Chain A, Protein (peanut lectin) | VSGAVVK-VT | 156–164 | VGGAVVTGVT | 70 |
| Chain A, Arachin Arah3 isoform | EQ-----GAIVT | 274–280 | EQVTNVGGAVVT | 50 |
| Chain A, Arachin Arah3 isoform | AVPTGV | 137–142 | AVVTGV | 83 |
| Chain A, Protein (peanut lectin) | AGSIGGGT | 98–105 | AGSIAAAT | 63 |
| Shrimp Protein | Shrimp Protein Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
| Chain A, Triosephosphate isomerase | FMKTGPLSPNTEVVV | 28–42 | FMK-G-LSKAKEGVV | 60 |
| Chain A, Proliferating cell nuclear antigen | TKEGVKFSAA | 163–172 | TKEGVVHGVA * | 60 |
| Chain A, Triosephosphate isomerase | AIGTGKTA | 170–177 | AVVTGVTA | 63 |
| Chain A, Arginine kinase | QDGILE | 343–348 | QEGILE | 83 |
| Pineapple Bromelain Protein | Pineapple Bromelain Sequence | Mapped Start to End | Alpha-Synuclein Sequence | ID (%) |
| Three-Dimensional Structure of Pineapple Cystatin | KAKEQVV | 85–91 | KAKEGVV * | 86 |
| Three-Dimensional Structure of Pineapple Cystatin | AEAEAEEEEG | 19–28 | AEAAGKTKEG | 50 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vojdani, A.; Lerner, A.; Vojdani, E. Cross-Reactivity and Sequence Homology Between Alpha-Synuclein and Food Products: A Step Further for Parkinson’s Disease Synucleinopathy. Cells 2021, 10, 1111. https://doi.org/10.3390/cells10051111
Vojdani A, Lerner A, Vojdani E. Cross-Reactivity and Sequence Homology Between Alpha-Synuclein and Food Products: A Step Further for Parkinson’s Disease Synucleinopathy. Cells. 2021; 10(5):1111. https://doi.org/10.3390/cells10051111
Chicago/Turabian StyleVojdani, Aristo, Aaron Lerner, and Elroy Vojdani. 2021. "Cross-Reactivity and Sequence Homology Between Alpha-Synuclein and Food Products: A Step Further for Parkinson’s Disease Synucleinopathy" Cells 10, no. 5: 1111. https://doi.org/10.3390/cells10051111
APA StyleVojdani, A., Lerner, A., & Vojdani, E. (2021). Cross-Reactivity and Sequence Homology Between Alpha-Synuclein and Food Products: A Step Further for Parkinson’s Disease Synucleinopathy. Cells, 10(5), 1111. https://doi.org/10.3390/cells10051111