
foxm1 Modulates Cell Non-Autonomous Response in Zebrafish Skeletal Muscle Homeostasis



Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Maintenance
2.2. Production of sgRNA and Cas9 mRNA
2.3. Microinjection in Zebrafish Embryos
2.4. Detection of Mutations
2.5. Identification of Founders and Stable Mutant Lines
2.6. Fluorescence-Activated Cell Sorting (FACS)
2.7. Tissue-Specific CRISPR/Cas9
2.8. Tol2 Transposase Synthesis
2.9. Tissue-Specific Mutagenesis Assay
2.10. RNA Extraction and Reverse Transcription
2.11. Real Time Quantitative PCR (RT-qPCR)
2.12. Cleaved Caspase-3 Immunostaining and Quantification
2.13. Pax7 Immunostaining and Quantification
2.14. Statistical Analysis
3. Results and Discussion
3.1. foxm1 Loss-of-Function Is Deleterious during Zebrafish Embryogenesis
3.2. foxm1 Is Expressed in Skeletal Muscle Cells
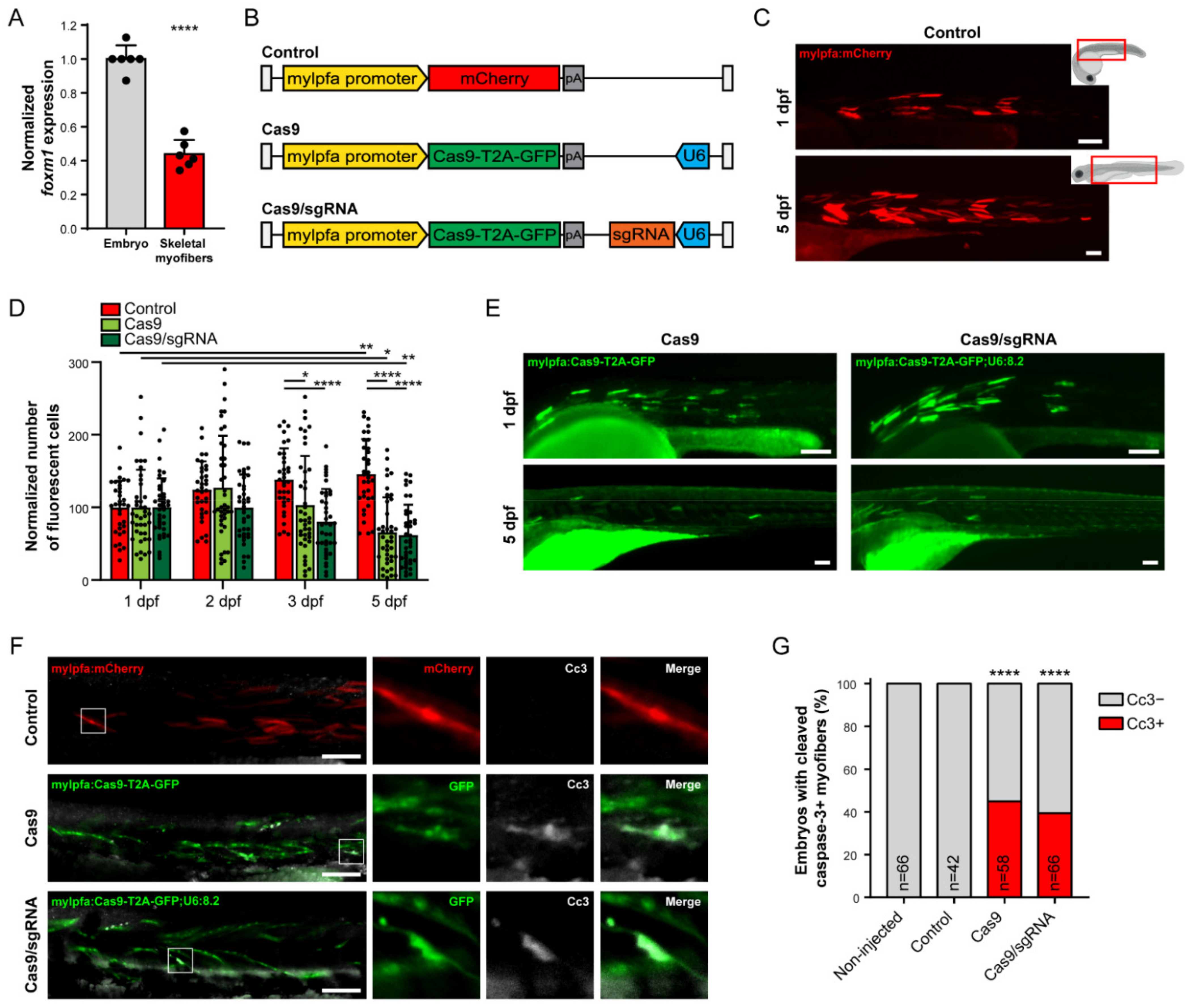
3.3. foxm1 Loss-of-Function and Strong Cas9 Expression Impair Muscle Cell Viability
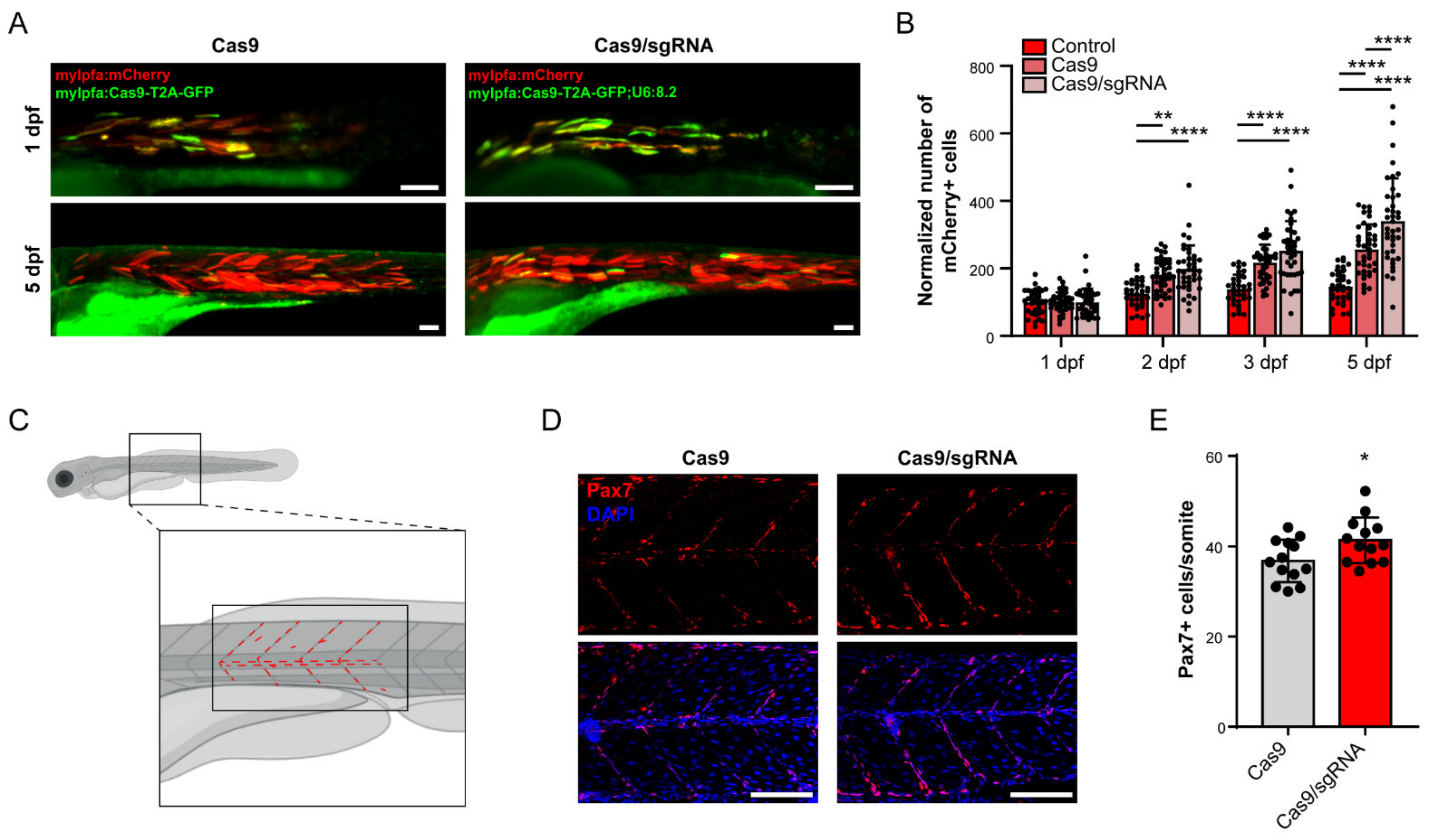
3.4. foxm1 in Muscle Cells Contributes to Non-Autonomous Tissue Repair
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Laoukili, J.; Kooistra, M.R.H.; Brás, A.; Kauw, J.; Kerkhoven, R.M.; Morrison, A.; Clevers, H.; Medema, R.H. FoxM1 Is Required for Execution of the Mitotic Programme and Chromosome Stability. Nat. Cell Biol. 2005, 7, 126–136. [Google Scholar] [CrossRef]
- Wang, X.; Kiyokawa, H.; Dennewitz, M.B.; Costa, R.H. The Forkhead Box M1b Transcription Factor Is Essential for Hepatocyte DNA Replication and Mitosis during Mouse Liver Regeneration. Proc. Natl. Acad. Sci. USA 2002, 99, 16881–16886. [Google Scholar] [CrossRef] [Green Version]
- Macedo, J.C.; Vaz, S.; Bakker, B.; Ribeiro, R.; Bakker, P.L.; Escandell, J.M.; Ferreira, M.G.; Medema, R.; Foijer, F.; Logarinho, E. FoxM1 Repression during Human Aging Leads to Mitotic Decline and Aneuploidy-Driven Full Senescence. Nat. Commun. 2018, 9, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ustiyan, V.; Wang, I.-C.; Ren, X.; Zhang, Y.; Snyder, J.; Xu, Y.; Wert, S.E.; Lessard, J.L.; Kalin, T.V.; Kalinichenko, V.V. Forkhead Box M1 Transcriptional Factor Is Required for Smooth Muscle Cells during Embryonic Development of Blood Vessels and Esophagus. Dev. Biol. 2009, 336, 266–279. [Google Scholar] [CrossRef] [Green Version]
- Kalin, T.V.; Ustiyan, V.; Kalinichenko, V.V. Multiple Faces of FoxM1 Transcription Factor: Lessons from Transgenic Mouse Models. Cell Cycle 2011, 10, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Chang-Panesso, M.; Kadyrov, F.F.; Lalli, M.; Wu, H.; Ikeda, S.; Kefaloyianni, E.; Abdelmageed, M.M.; Herrlich, A.; Kobayashi, A.; Humphreys, B.D. FOXM1 Drives Proximal Tubule Proliferation during Repair from Acute Ischemic Kidney Injury. J. Clin. Investig. 2019, 129, 5501–5517. [Google Scholar] [CrossRef]
- Korver, W.; Roose, J.; Clevers, H. The Winged-Helix Transcription Factor Trident Is Expressed in Cycling Cells. Nucleic Acids Res. 1997, 25, 1715–1719. [Google Scholar] [CrossRef] [Green Version]
- Krupczak-Hollis, K.; Wang, X.; Kalinichenko, V.V.; Gusarova, G.A.; Wang, I.-C.; Dennewitz, M.B.; Yoder, H.M.; Kiyokawa, H.; Kaestner, K.H.; Costa, R.H. The Mouse Forkhead Box M1 Transcription Factor Is Essential for Hepatoblast Mitosis and De-velopment of Intrahepatic Bile Ducts and Vessels during Liver Morphogenesis. Dev. Biol. 2004, 276, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, S.; Kim, I.-M.; Petrovic, V.; Malin, D.; Wang, I.-C.; Kalin, T.V.; Meliton, L.; Zhao, Y.-Y.; Ackerson, T.; Qin, Y.; et al. Myocardium Defects and Ventricular Hypoplasia in Mice Homozygous Null for TheForkhead Box M1 Transcription Factor. Dev. Dyn. 2007, 236, 1000–1013. [Google Scholar] [CrossRef]
- Korver, W.; Schilham, M.W.; Moerer, P.; van den Hoff, M.J.; Dam, K.; Lamers, W.H.; Medema, R.H.; Clevers, H. Uncoupling of S Phase and Mitosis in Cardiomyocytes and Hepatocytes Lacking the Winged-Helix Transcription Factor Trident. Curr. Biol. 1998, 8, 1327–1330. [Google Scholar] [CrossRef] [Green Version]
- Eckfeldt, C.E.; Mendenhall, E.M.; Flynn, C.M.; Wang, T.F.; Pickart, M.A.; Grindle, S.M.; Ekker, S.C.; Verfaillie, C.M. Functional Analysis of Human Hematopoietic Stem Cell Gene Expression Using Zebrafish. PLoS Biol. 2005, 3, e254. [Google Scholar] [CrossRef] [Green Version]
- Sasagawa, S.; Nishimura, Y.; Hayakawa, Y.; Murakami, S.; Ashikawa, Y.; Yuge, M.; Okabe, S.; Kawaguchi, K.; Kawase, R.; Tanaka, T. E2F4 Promotes Neuronal Regeneration and Functional Recovery after Spinal Cord Injury in Zebrafish. Front. Pharmacol. 2016, 7, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadler, K.C.; Krahn, K.N.; Gaur, N.A.; Ukomadu, C. Liver Growth in the Embryo and during Liver Regeneration in Zebrafish Requires the Cell Cycle Regulator, Uhrf1. PNAS 2007, 104, 1570–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marceau, A.H.; Brison, C.M.; Nerli, S.; Arsenault, H.E.; McShan, A.C.; Chen, E.; Lee, H.-W.; Benanti, J.A.; Sgourakis, N.G.; Rubin, S.M. An Order-to-Disorder Structural Switch Activates the FoxM1 Transcription Factor. eLife 2019, 8, e46131. [Google Scholar] [CrossRef]
- Laoukili, J.; Alvarez, M.; Meijer, L.A.T.; Stahl, M.; Mohammed, S.; Kleij, L.; Heck, A.J.R.; Medema, R.H. Activation of FoxM1 during G2 Requires Cyclin A/Cdk-Dependent Relief of Autorepression by the FoxM1 N-Terminal Domain. Mol. Cell. Biol. 2008, 28, 3076–3087. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Wang, Z.; Costa, R.H.; Tyner, A.; Lau, L.F.; Raychaudhuri, P. An N-Terminal Inhibitory Domain Modulates Activity of FoxM1 during Cell Cycle. Oncogene 2008, 27, 1696–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Major, M.L.; Lepe, R.; Costa, R.H. Forkhead Box M1B Transcriptional Activity Requires Binding of Cdk-Cyclin Complexes for Phosphorylation-Dependent Recruitment of P300/CBP Coactivators. Mol. Cell. Biol. 2004, 24, 2649–2661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, G.B.; Li, X.Z.; Zeng, S.; Liu, C.; Yang, S.M.; Yang, L.; Hu, C.J.; Bai, J.Y. Regulation of the Master Regulator FOXM1 in Cancer. Cell Commun. Signal. 2018, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.H.; Choi, M.H.; Kim, J.H.; Lim, I.K.; Park, T.J. C-Terminus-Deleted FoxM1 Is Expressed in Cancer Cell Lines and Induces Chromosome Instability. Carcinogenesis 2013, 34, 1907–1917. [Google Scholar] [CrossRef] [Green Version]
- Zona, S.; Bella, L.; Burton, M.J.; Nestal de Moraes, G.; Lam, E.W.F. FOXM1: An Emerging Master Regulator of DNA Damage Response and Genotoxic Agent Resistance. Biochim. Biophys. Acta Gene Regul. Mech. 2014, 1839, 1316–1322. [Google Scholar] [CrossRef] [Green Version]
- Black, M.; Arumugam, P.; Shukla, S.; Pradhan, A.; Ustiyan, V.; Milewski, D.; Kalinichenko, V.V.; Kalin, T.V. FOXM1 Nuclear Transcription Factor Translocates into Mitochondria and Inhibits Oxidative Phosphorylation. Mol. Biol. Cell 2020, 31, 1411–1424. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Messina, G. Comparative Myogenesis in Teleosts and Mammals. Cell. Mol. Life Sci. 2014, 71, 3081–3099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, H.E.; Ingham, P.W. Control of Muscle Fibre-Type Diversity during Embryonic Development: The Zebrafish Paradigm. Mech. Dev. 2013, 130, 447–457. [Google Scholar] [CrossRef]
- Stickney, H.L.; Barresi, M.J.F.; Devoto, S.H. Somite Development in Zebrafish. Dev. Dyn. 2000, 219, 287–303. [Google Scholar] [CrossRef]
- Keenan, S.R.; Currie, P.D. The Developmental Phases of Zebrafish Myogenesis. J. Dev. Biol. 2019, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.D.; Gurevich, D.B.; Sonntag, C.; Hersey, L.; Alaei, S.; Nim, H.T.; Siegel, A.; Hall, T.E.; Rossello, F.J.; Boyd, S.E.; et al. Muscle Stem Cells Undergo Extensive Clonal Drift during Tissue Growth via Meox1-Mediated Induction of G2 Cell-Cycle Arrest. Cell Stem Cell 2017, 21, 107–119.e6. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, D.B.; Nguyen, P.D.; Siegel, A.L.; Ehrlich, O.V.; Sonntag, C.; Phan, J.M.N.; Berger, S.; Ratnayake, D.; Hersey, L.; Berger, J.; et al. Asymmetric Division of Clonal Muscle Stem Cells Coordinates Muscle Regeneration in Vivo. Science 2016, 353, aad9969. [Google Scholar] [CrossRef]
- Maves, L. Recent Advances Using Zebrafish Animal Models for Muscle Disease Drug Discovery. Expert Opin. Drug Discov. 2014, 9, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.E.; Wood, A.J.; Ehrlich, O.; Li, M.; Sonntag, C.S.; Cole, N.J.; Huttner, I.G.; Sztal, T.E.; Currie, P.D. Cellular Rescue in a Zebrafish Model of Congenital Muscular Dystrophy Type 1A. npj Regen. Med. 2019, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, J.; Currie, P.D. Zebrafish Models Flex Their Muscles to Shed Light on Muscular Dystrophies. DMM Dis. Models Mech. 2012, 5, 726–732. [Google Scholar] [CrossRef] [Green Version]
- Knappe, S.; Zammit, P.S.; Knight, R.D. A Population of Pax7-Expressing Muscle Progenitor Cells Show Differential Responses to Muscle Injury Dependent on Developmental Stage and Injury Extent. Front. Aging Neurosci. 2015, 7, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berberoglu, M.A.; Gallagher, T.L.; Morrow, Z.T.; Talbot, J.C.; Hromowyk, K.J.; Tenente, I.M.; Langenau, D.M.; Amacher, S.L. Satellite-like Cells Contribute to Pax7-Dependent Skeletal Muscle Repair in Adult Zebrafish. Dev. Biol. 2017, 424, 162–180. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bu, N.; Qiao, X.; Zuo, Z.; Shu, Y.; Liu, Z.; Qian, Z.; Chen, J.; Hou, Y. Forkhead Box M1 Transcriptionally Regulates the Expression of Long Noncoding RNAs Snhg8 and Gm26917 to Promote Proliferation and Survival of Muscle Satellite Cells. Stem Cells 2018, 36, 1097–1108. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Li, L.; Xu, S.; Liu, Z.; Zhou, C.; Li, Z.; Liu, Y.; Wu, W.; Huang, Y.; Kuang, M.; et al. A Cdh1–FoxM1–Apc Axis Controls Muscle Development and Regeneration. Cell Death Dis. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, D.M.; Lazure, F.; Corchado, A.H.; Perkins, T.J.; Najafabadi, H.S.; Soleimani, V.D. High-Resolution Genome-Wide Expression Analysis of Single Myofibers Using Smart-Seq. J. Biol. Chem. 2019, 294, 20097–20108. [Google Scholar] [CrossRef] [PubMed]
- Cornet, C.; di Donato, V.; Terriente, J. Combining Zebrafish and CRISPR/Cas9: Toward a More Efficient Drug Discovery Pipeline. Front. Pharmacol. 2018, 9, 703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.H.Y.; Pannunzio, N.R.; Adachi, N.; Lieber, M.R. Non-Homologous DNA End Joining and Alternative Pathways to Double-Strand Break Repair. Nat. Rev. Mol. Cell Biol. 2017, 18, 495–506. [Google Scholar] [CrossRef]
- Burger, A.; Lindsay, H.; Felker, A.; Hess, C.; Anders, C.; Chiavacci, E.; Zaugg, J.; Weber, L.M.; Catena, R.; Jinek, M.; et al. Maximizing Mutagenesis with Solubilized CRISPR-Cas9 Ribonucleoprotein Complexes. Development 2016, 143, 2025–2037. [Google Scholar] [CrossRef] [Green Version]
- Jao, L.E.; Wente, S.R.; Chen, W. Efficient Multiplex Biallelic Zebrafish Genome Editing Using a CRISPR Nuclease System. Proc. Natl. Acad. Sci. USA 2013, 110, 13904–13909. [Google Scholar] [CrossRef] [Green Version]
- Ablain, J.; Durand, E.M.; Yang, S.; Zhou, Y.; Zon, L.I. A CRISPR/Cas9 Vector System for Tissue-Specific Gene Disruption in Zebrafish. Dev. Cell 2015, 32, 756–764. [Google Scholar] [CrossRef] [Green Version]
- Ihry, R.J.; Worringer, K.A.; Salick, M.R.; Frias, E.; Ho, D.; Theriault, K.; Kommineni, S.; Chen, J.; Sondey, M.; Ye, C.; et al. P53 Inhibits CRISPR-Cas9 Engineering in Human Pluripotent Stem Cells. Nat. Med. 2018, 24, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Kim, J.; Tang, Q.; Chen, Q.; Liu, J.; Xu, Y.; Fu, X. CAS9 Is a Genome Mutator by Directly Disrupting DNA-PK Dependent DNA Repair Pathway. Protein Cell 2020, 11, 352–365. [Google Scholar] [CrossRef] [Green Version]
- Benchling [Biology Software]. Available online: https://benchling.com (accessed on 1 April 2018).
- Doench, J.G.; Fusi, N.; Sullender, M.; Hegde, M.; Vaimberg, E.W.; Donovan, K.F.; Smith, I.; Tothova, Z.; Wilen, C.; Orchard, R.; et al. Optimized SgRNA Design to Maximize Activity and Minimize Off-Target Effects of CRISPR-Cas9. Nat. Biotechnol. 2016, 34, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA Targeting Specificity of RNA-Guided Cas9 Nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.J.; Joung, J.K. Efficient Genome Editing in Zebrafish Using a CRISPR-Cas System. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef]
- Bessa, J.; Gómez-Skarmeta, J.L. Making Reporter Gene Constructs to Analyze Cis-Regulatory Elements. Methods Mol. Biol. 2011, 772, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkman, E.K.; Chen, T.; Amendola, M.; van Steensel, B. Easy Quantitative Assessment of Genome Editing by Sequence Trace Decomposition. Nucleic Acids Res. 2014, 42, e168. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Sorrells, S.; Toruno, C.; Stewart, R.A.; Jette, C. Analysis of Apoptosis in Zebrafish Embryos by Whole-Mount Immunofluorescence to Detect Activated Caspase 3. J. Vis. Exp. 2013, 82, 51060. [Google Scholar] [CrossRef]
- Kawakami, A.; Kimura-Kawakami, M.; Nomura, T.; Fujisawa, H. Distributions of PAX6 and PAX7 Proteins Suggest Their Involvement in Both Early and Late Phases of Chick Brain Development. Mech. Dev. 1997, 66, 119–130. [Google Scholar] [CrossRef]
- Seger, C.; Hargrave, M.; Wang, X.; Chai, R.J.; Elworthy, S.; Ingham, P.W. Analysis of Pax7 Expressing Myogenic Cells in Zebrafish Muscle Development, Injury, and Models of Disease. Dev. Dyn. 2011, 240, 2440–2451. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Mateos, M.A.; Vejnar, C.E.; Beaudoin, J.D.; Fernandez, J.P.; Mis, E.K.; Khokha, M.K.; Giraldez, A.J. CRISPRscan: Designing Highly Efficient SgRNAs for CRISPR-Cas9 Targeting in Vivo. Nat. Methods 2015, 12, 982–988. [Google Scholar] [CrossRef] [Green Version]
- Amrani, N.; Sachs, M.S.; Jacobson, A. Early Nonsense: MRNA Decay Solves a Translational Problem. Nat. Rev. Mol. Cell Biol. 2006, 7, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Lindeboom, R.G.H.; Supek, F.; Lehner, B. The Rules and Impact of Nonsense-Mediated MRNA Decay in Human Cancers. Nat. Genet. 2016, 48, 1112–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wittkopp, N.; Huntzinger, E.; Weiler, C.; Saulière, J.; Schmidt, S.; Sonawane, M.; Izaurralde, E. Nonsense-Mediated MRNA Decay Effectors Are Essential for Zebrafish Embryonic Development and Survival. Mol. Cell. Biol. 2009, 29, 3517–3528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, E.; Maquat, L.E. A Rule for Termination-Codon Position within Intron-Containing Genes: When Nonsense Affects RNA Abundance. Trends Biochem. Sci. 1998, 23, 198–199. [Google Scholar] [CrossRef]
- Myatt, S.S.; Kongsema, M.; Man, C.W.Y.; Kelly, D.J.; Gomes, A.R.; Khongkow, P.; Karunarathna, U.; Zona, S.; Langer, J.K.; Dunsby, C.W.; et al. SUMOylation Inhibits FOXM1 Activity and Delays Mitotic Transition. Oncogene 2014, 33, 4316–4329. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Zhao, G.; Sun, X.; Wang, P.; Xie, N.; Luo, J.; Tong, T. Acetylation of FOXM1 Is Essential for Its Transactivation and Tumor Growth Stimulation. Oncotarget 2016, 7, 60366–60382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Xu, Y.; Yu, S.; Lu, L.; Ding, M.; Cheng, J.; Song, G.; Gao, X.; Yao, L.; Fan, D.; et al. An Efficient Genotyping Method for Genome-Modified Animals and Human Cells Generated with CRISPR/Cas9 System. Sci. Rep. 2014, 4, 6420. [Google Scholar] [CrossRef] [PubMed]
- El-Brolosy, M.A.; Kontarakis, Z.; Rossi, A.; Kuenne, C.; Günther, S.; Fukuda, N.; Kikhi, K.; Boezio, G.L.M.; Takacs, C.M.; Lai, S.L.; et al. Genetic Compensation Triggered by Mutant MRNA Degradation. Nature 2019, 568, 193–197. [Google Scholar] [CrossRef]
- Golson, M.L.; Kaestner, K.H. Fox Transcription Factors: From Development to Disease. Development 2016, 143, 4558–4570. [Google Scholar] [CrossRef] [Green Version]
- Shimeld, S.M.; Degnan, B.; Luke, G.N. Evolutionary Genomics of the Fox Genes: Origin of Gene Families and the Ancestry of Gene Clusters. Genomics 2010, 95, 256–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Overbeek, M.; Capurso, D.; Carter, M.M.; Thompson, M.S.; Frias, E.; Russ, C.; Reece-Hoyes, J.S.; Nye, C.; Gradia, S.; Vidal, B.; et al. DNA Repair Profiling Reveals Nonrandom Outcomes at Cas9-Mediated Breaks. Mol. Cell 2016, 63, 633–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taheri-Ghahfarokhi, A.; Taylor, B.J.M.; Nitsch, R.; Lundin, A.; Cavallo, A.L.; Madeyski-Bengtson, K.; Karlsson, F.; Clausen, M.; Hicks, R.; Mayr, L.M.; et al. Decoding Non-Random Mutational Signatures at Cas9 Targeted Sites. Nucleic Acids Res. 2018, 46, 8417–8434. [Google Scholar] [CrossRef] [Green Version]
- Sacco, A.; Pajalunga, D.; Latella, L.; Siepi, F.; Rufini, A.; Crescenzi, M. Cell Cycle Reactivation in Skeletal Muscle and Other Terminally Differentiated Cells; Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Bessa, J.; Tena, J.J.; de La Calle-Mustienes, E.; Fernández-Miñán, A.; Naranjo, S.; Fernández, A.; Montoliu, L.; Akalin, A.; Lenhard, B.; Casares, F.; et al. Zebrafish Enhancer Detection (ZED) Vector: A New Tool to Facilitate Transgenesis and the Functional Analysis of Cis-Regulatory Regions in Zebrafish. Dev. Dyn. 2009, 238, 2409–2417. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Ju, B.; Chong, S.W.; He, J.; Wang, X.; Xu, Y.; Wan, H.; Tong, Y.; Yan, T.; Korzh, V.; Gong, Z. Recapitulation of Fast Skeletal Muscle Development in Zebrafish by Transgenic Expression of GFP under the Mylz2 Promoter. Dev. Dyn. 2003, 227, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Roy, S. The Zebrafish Fast Myosin Light Chain Mylpfa:H2B-GFP Transgene Is a Useful Tool for In Vivo Imaging of Myocyte Fusion in the Vertebrate Embryo. Gene Expr. Patterns 2016, 20, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Pourghadamyari, H.; Rezaei, M.; Ipakchi-Azimi, A.; Eisa-Beygi, S.; Basiri, M.; Tahamtani, Y.; Baharvand, H. Establishing a New Animal Model for Muscle Regeneration Studies. Mol. Biol. Res. Commun. 2019, 8, 171–179. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell Death: Critical Control Points. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.G.; Cullen, S.P.; Sheridan, C.; Lüthi, A.U.; Gerner, C.; Martin, S.J. Executioner Caspase-3 and Caspase-7 Are Functionally Distinct Proteases. Proc. Natl. Acad. Sci. USA 2008, 105, 12815–12819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spead, O.; Verreet, T.; Donelson, C.J.; Poulain, F.E. Characterization of the Caspase Family in Zebrafish. PLoS ONE 2018, 13, e0197966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Zhao, L.; Tian, T.; Zhang, Y.; Tong, J.; Zheng, X.; Meng, A. Interruption of Cenph Causes Mitotic Failure and Embryonic Death, and Its Haploinsufficiency Suppresses Cancer in Zebrafish. J. Biol. Chem. 2010, 285, 27924–27934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratz, E.; Eimon, P.M.; Mukhyala, K.; Stern, H.; Zha, J.; Strasser, A.; Hart, R.; Ashkenazi, A. Functional Characterization of the Bcl-2 Gene Family in the Zebrafish. Cell Death Differ. 2006, 13, 1631–1640. [Google Scholar] [CrossRef]
- Mathias, J.R.; Dodd, M.E.; Walters, K.B.; Yoo, S.K.; Ranheim, E.A.; Huttenlocher, A. Characterization of Zebrafish Larval Inflammatory Macrophages. Dev. Comp. Immunol. 2009, 33, 1212–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, M.R.; Ravichandran, K.S. The Dynamics of Apoptotic Cell Clearance. Dev. Cell 2016, 38, 147–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Li, S.; Li, X.J. Shortening the Half-Life of Cas9 Maintains Its Gene Editing Ability and Reduces Neuronal Toxicity. Cell Rep. 2018, 25, 2653–2659.e3. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an Endocrine Organ: Focus on Muscle-Derived Interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.K. Muscle as a Secretory Organ. In Comprehensive Physiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; Volume 3, pp. 1337–1362. [Google Scholar]
- Pillon, N.J.; Bilan, P.J.; Fink, L.N.; Klip, A. Cross-Talk between Skeletal Muscle and Immune Cells: Muscle-Derived Mediators and Metabolic Implications. Am. J. Physiol. Endocrinol. Metab. 2013, 304, 453–465. [Google Scholar] [CrossRef]
- Palstra, A.P.; Rovira, M.; Rizo-Roca, D.; Torrella, J.R.; Spaink, H.P.; Planas, J.V. Swimming-Induced Exercise Promotes Hy-pertrophy and Vascularization of Fast Skeletal Muscle Fibres and Activation of Myogenic and Angiogenic Transcriptional Programs in Adult Zebrafish. BMC Genom. 2014, 15, 1136. [Google Scholar] [CrossRef] [Green Version]
- Rovira, M.; Arrey, G.; Planas, J.V. Exercise-Induced Hypertrophic and Oxidative Signaling Pathways and Myokine Expression in Fast Muscle of Adult Zebrafish. Front. Physiol. 2017, 8, 1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pipalia, T.G.; Koth, J.; Roy, S.D.; Hammond, C.L.; Kawakami, K.; Hughes, S.M. Cellular Dynamics of Regeneration Reveals Role of Two Distinct Pax7 Stem Cell Populations in Larval Zebrafish Muscle Repair. DMM Dis. Models Mech. 2016, 9, 671–684. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.D.; Williams, V.C.; Pipalia, T.G.; Li, K.; Hammond, C.L.; Knappe, S.; Knight, R.D.; Hughes, S.M. Myotome Adaptability Confers Developmental Robustness to Somitic Myogenesis in Response to Fibre Number Alteration. Dev. Biol. 2017, 431, 321–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balli, D.; Ustiyan, V.; Zhang, Y.; Wang, I.-C.; Masino, A.J.; Ren, X.; Whitsett, J.A.; Kalinichenko, V.V.; Kalin, T.V. Foxm1 Transcription Factor Is Required for Lung Fibrosis and Epithelial-to-Mesenchymal Transition. EMBO J. 2013, 32, 231–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, R.-M.; Lu, X.-H.; Lin, J.; Hu, J.; Rong, Z.-J.; Xu, W.-C.; Liu, Z.-W.; Zeng, W.-T. Knockdown of FOXM1 Attenuates Inflammatory Response in Human Osteoarthritis Chondrocytes. Int. Immunopharmacol. 2019, 68, 74–80. [Google Scholar] [CrossRef]
- Toth, K.G.; McKay, B.R.; de Lisio, M.; Little, J.P.; Tarnopolsky, M.A.; Parise, G. IL-6 Induced STAT3 Signalling Is Associated with the Proliferation of Human Muscle Satellite Cells Following Acute Muscle Damage. PLoS ONE 2011, 6, e17392. [Google Scholar] [CrossRef]
- Steyn, P.J.; Dzobo, K.; Smith, R.I.; Myburgh, K.H. Interleukin-6 Induces Myogenic Differentiation via JAK2-STAT3 Signaling in Mouse C2C12 Myoblast Cell Line and Primary Human Myoblasts. Int. J. Mol. Sci. 2019, 20, 5273. [Google Scholar] [CrossRef] [Green Version]
- Spangenburg, E.E.; Booth, F.W. Multiple Signaling Pathways Mediate LIF-Induced Skeletal Muscle Satellite Cell Proliferation. Am. J. Physiol. Cell Physiol. 2002, 283, C204–C211. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.J.; Thaloor, D.; Matteson, S.; Pavlath, G.K. Hepatocyte Growth Factor Affects Satellite Cell Activation and Differen-tiation in Regenerating Skeletal Muscle. Am. J. Physiol. Cell Physiol. 2000, 278, C174–C181. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Noulet, F.; Edom-Vovard, F.; le Grand, F.; Duprez, D. Bmp Signaling at the Tips of Skeletal Muscles Regulates the Number of Fetal Muscle Progenitors and Satellite Cells during Development. Dev. Cell 2010, 18, 643–654. [Google Scholar] [CrossRef] [Green Version]
- Ono, Y.; Calhabeu, F.; Morgan, J.E.; Katagiri, T.; Amthor, H.; Zammit, P.S. BMP Signalling Permits Population Expansion by Preventing Premature Myogenic Differentiation in Muscle Satellite Cells. Cell Death Differ. 2011, 18, 222–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stantzou, A.; Schirwis, E.; Swist, S.; Alonso-Martin, S.; Polydorou, I.; Zarrouki, F.; Mouisel, E.; Beley, C.; Julien, A.; le Grand, F.; et al. BMP Signaling Regulates Satellite Cell-Dependent Postnatal Muscle Growth. Development 2017, 144, 2737–2747. [Google Scholar] [CrossRef] [Green Version]
- Pawlikowski, B.; Vogler, T.O.; Gadek, K.; Olwin, B.B. Regulation of Skeletal Muscle Stem Cells by Fibroblast Growth Factors. Dev. Dyn. 2017, 246, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Ren, X.; Wang, I.C.; Pradhan, A.; Zhang, Y.; Flood, H.M.; Han, B.; Whitsett, J.A.; Kalin, T.V.; Kalinichenko, V.V. The FOXM1 Inhibitor RCM-1 Suppresses Goblet Cell Metaplasia and Prevents IL-13 and STAT6 Signaling in Allergen-Exposed Mice. Sci. Signal. 2017, 10, eaai8583. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, A.P.; Stone, R.C.; Brooks, S.R.; Pastar, I.; Jozic, I.; Hasneen, K.; O’Neill, K.; Mehdizadeh, S.; Head, C.R.; Strbo, N.; et al. Deregulated Immune Cell Recruitment Orchestrated by FOXM1 Impairs Human Diabetic Wound Healing. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genotype | DNA Sequence | aa Sequence | Frequency (N) |
|---|---|---|---|
| Wild type | …AAGATGAAGCCTCTACTGCCTCGGACTGAC… | …KMKPLLPRTD… | 36% (5) |
| 3 nts deletion | …AAGATGAAGCCTCTACTGCCTCGGACTGAC… | …KMKPLLPRTD… | 7% (1) |
| 9 nts deletion | …AAGATGAAGCCTCTACTGCCTCGGACTGAC… | …KMKPLLPRTD… | 43% (6) |
| 12 nts deletion | …AAGATGAAGCCTCTACTGCCTCGGACTGAC… | …KMKPLLPRTD… | 14% (2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, F.J.; Carvalho, L.; Logarinho, E.; Bessa, J. foxm1 Modulates Cell Non-Autonomous Response in Zebrafish Skeletal Muscle Homeostasis. Cells 2021, 10, 1241. https://doi.org/10.3390/cells10051241
Ferreira FJ, Carvalho L, Logarinho E, Bessa J. foxm1 Modulates Cell Non-Autonomous Response in Zebrafish Skeletal Muscle Homeostasis. Cells. 2021; 10(5):1241. https://doi.org/10.3390/cells10051241
Chicago/Turabian StyleFerreira, Fábio J., Leonor Carvalho, Elsa Logarinho, and José Bessa. 2021. "foxm1 Modulates Cell Non-Autonomous Response in Zebrafish Skeletal Muscle Homeostasis" Cells 10, no. 5: 1241. https://doi.org/10.3390/cells10051241
APA StyleFerreira, F. J., Carvalho, L., Logarinho, E., & Bessa, J. (2021). foxm1 Modulates Cell Non-Autonomous Response in Zebrafish Skeletal Muscle Homeostasis. Cells, 10(5), 1241. https://doi.org/10.3390/cells10051241