
Polyamine Metabolism in Scots Pine Embryogenic Cells under Potassium Deficiency

Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of Embryogenic Cell Line
2.2. Liquid Cultures
2.3. Measurement of pH and Conductivity in Culture Medium
2.4. Potassium Treatments
2.5. The Cell Mass Growth and Cell Viability
2.6. Analysis of Chemical Elements
2.7. Polyamine Analysis
2.8. RNA Isolation and Reverse Transcription
2.9. Quantitative Real-Time PCR Analysis
2.10. Statistical Analysis
3. Results
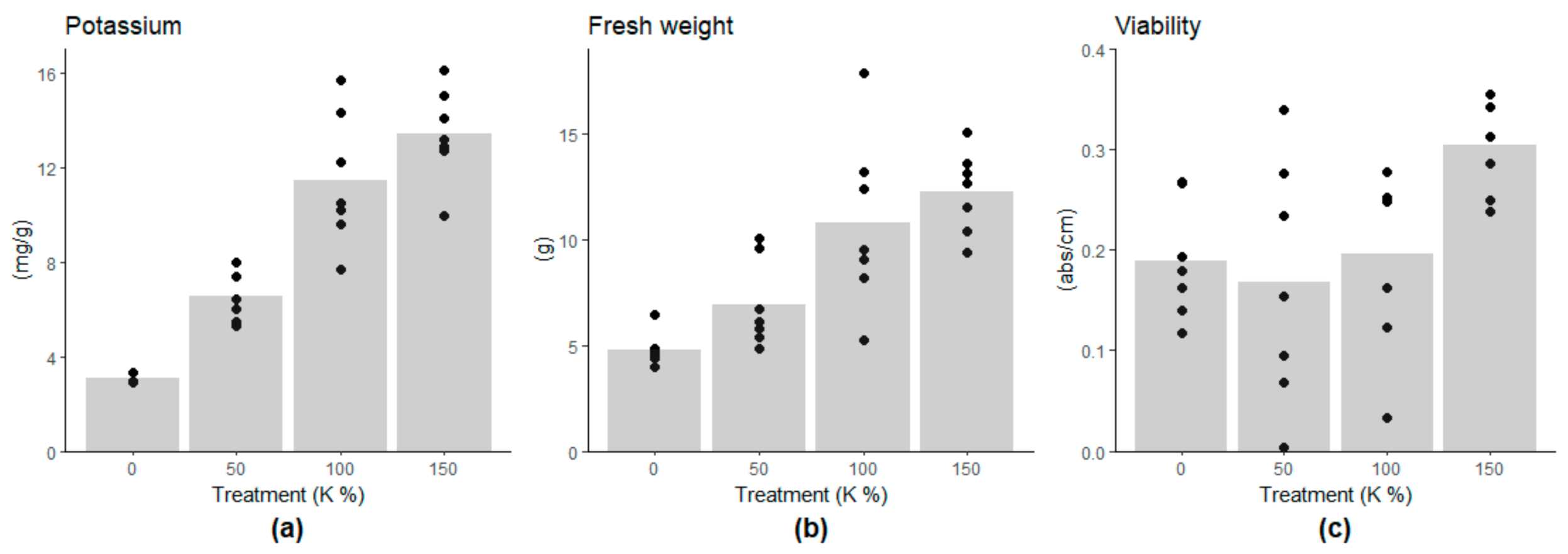
3.1. Effect of Potassium Deficiency on Viability and Growth of Embryogenic Cell Mass
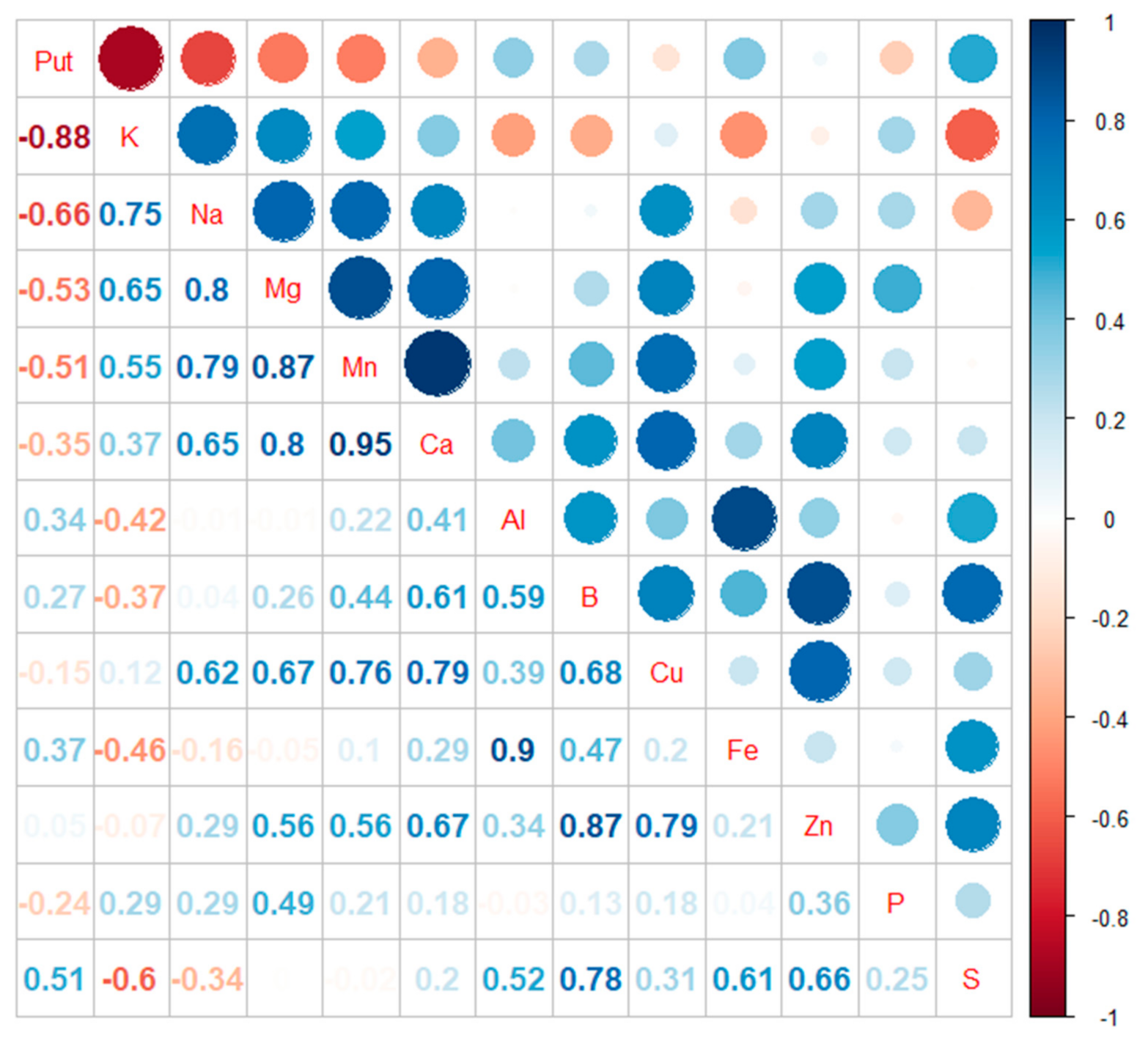
3.2. Content of Chemical Elements in Embryogenic Cells
3.3. Changes in pH and Conductivity of Culture Medium
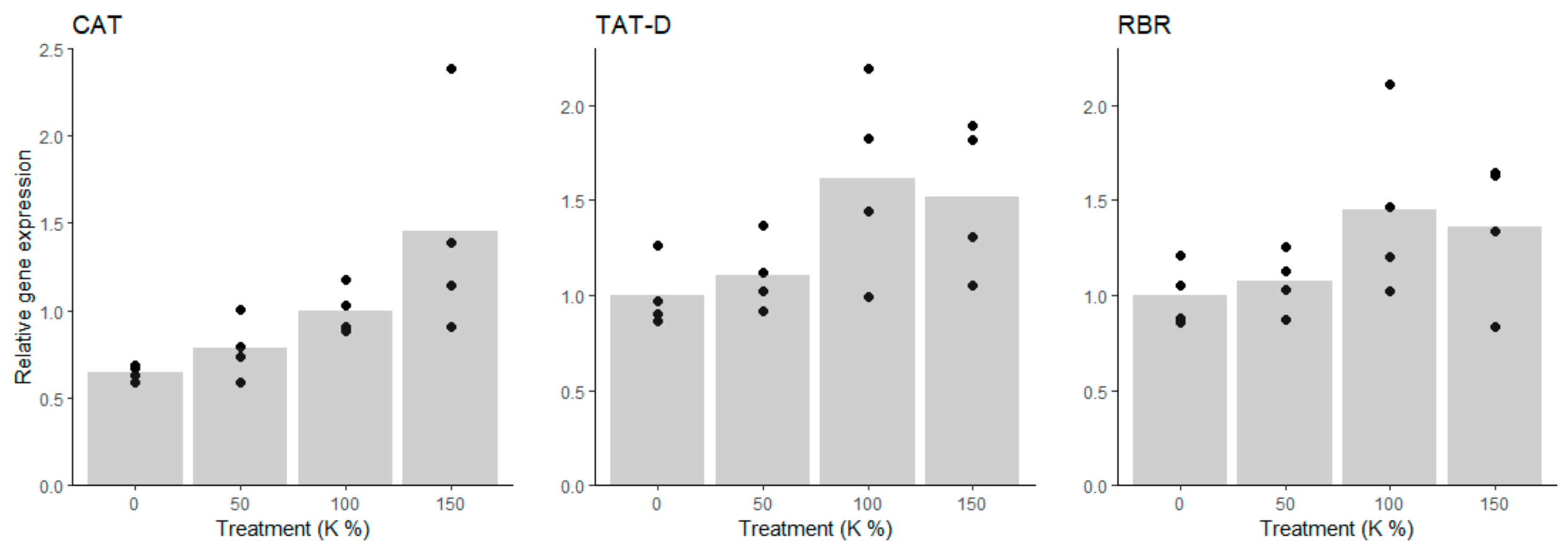
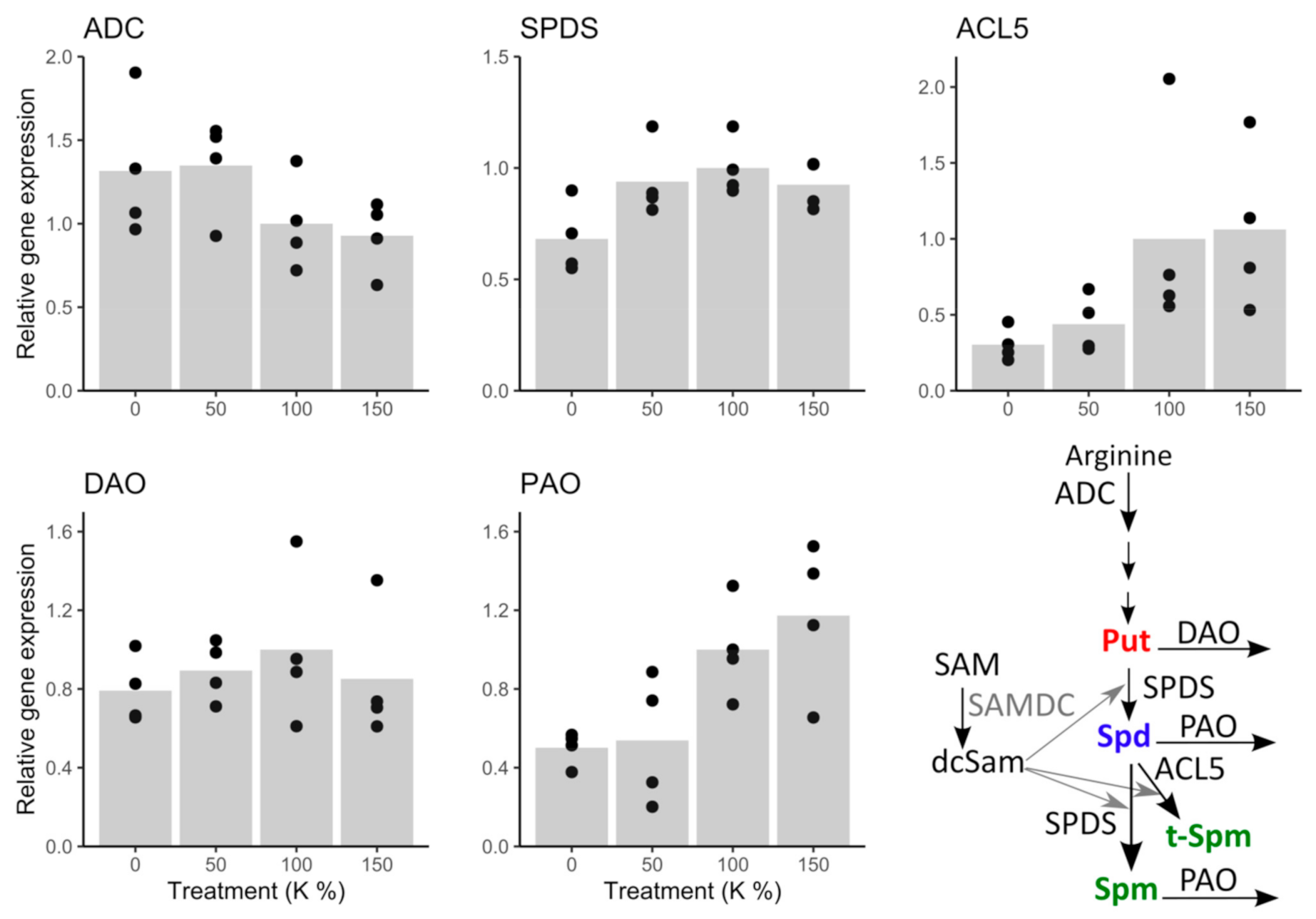
3.4. Transcriptional Regulation of Put Accumulation under Potassium Deficiency
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moilanen, M.; Saarinen, M.; Silfverberg, K. Foliar nitrogen, phosphorus and potassium concentrations of Scots pine in drained mires in Finland. Silva Fenn. 2010, 44, 583–601. [Google Scholar] [CrossRef] [Green Version]
- Kaunisto, S.; Tukeva, J. Kalilannoituksen tarve avosoille perustetuissa riukuasteen männiköissä (Need for potassium fertilization in pole stage pine stands established on bogs). Folia For 1984, 585, 40. [Google Scholar]
- Sarjala, T.; Kaunisto, S. Needle polyamine levels and potassium nutrition in Scots pine. Tree Physiol. 1993, 13, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Sarjala, T. Growth, Potassium and Polyamine Concentrations of Scots Pine seedlings in Relation to Potassium Availability under Controlled Growth Conditions. J. Plant Physiol. 1996, 147, 593–598. [Google Scholar] [CrossRef]
- Jokela, A.; Sarjala, T.; Kaunisto, S.; Huttunen, S. Effects of foliar potassium concentration on morphology, ultrastructure and polyamine concentrations of Scots pine needles. Tree Physiol. 1997, 17, 677–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, A.K.; Shankar, A.; Chandran, A.K.N.; Sharma, M.; Jung, K.H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2020, 71, 608–619. [Google Scholar] [CrossRef]
- Cui, J.; Pottosin, I.; Lamade, E.; Tcherkez, G. What is the role of putrescine accumulated under potassium deficiency? Plant Cell Environ. 2020, 43, 1331–1347. [Google Scholar] [CrossRef]
- Raddatz, N.; Morales de los Ríos, L.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated Transport of Nitrate, Potassium, and Sodium. Front. Plant Sci. 2020, 11, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Q.; Lyu, C.C.; Li, J.H.; Wan, C.Y.; Liu, L.; Xie, M.Q.; Zuo, R.J.; Ni, S.; Liu, F.; Zeng, F.C.; et al. Quantitative proteomic analysis of alligator weed leaves reveals that cationic peroxidase 1 plays vital roles in the potassium deficiency stress response. Int. J. Mol. Sci. 2020, 21, 2537. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, G.; Xue, H.; Zhang, J.; Wang, Q.; Zhang, Z.; Zhang, B. Metabolite Profile of Xylem Sap in Cotton Seedlings Is Changed by K Deficiency. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Richards, F.J.; Coleman, R.G. Occurrence of putrescine in potassium-deficient barley. Nature 1952, 170, 460. [Google Scholar] [CrossRef]
- Kaunisto, S.; Sarjala, T. Critical Needle Potassium Concentrations Indicated by Diamine Putrescine in Norway Spruce Growing on Peat Soils. Silva Fenn. 1997, 31, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Alcázar, R.; Planas, J.; Saxena, T.; Zazra, X.; Bortolotti, C.; Cuevas, J.; Bitrian, M.; Tiburcio, A.; Altabella, T. Putrecine accumulation confers drought tolerance in transgenic Arabidopsis plants over-expressing the homologous arginine decarboxylase 2 gene. Plant Physiol. Biochem. 2010, 48, 547–552. [Google Scholar] [CrossRef]
- Rodríguez-Kessler, M.; Alpuche-Solís, A.G.; Ruiz, O.A.; Jiménez-Bremont, J.F. Effect of salt stress on the regulation of maize (Zea mays L.) genes involved in polyamine biosynthesis. Plant Growth Regul. 2006, 48, 175–185. [Google Scholar] [CrossRef]
- Pottosin, I.; Olivas-Aguirre, M.; Dobrovinskaya, O.; Zepeda-Jazo, I.; Shabala, S. Modulation of Ion Transport Across Plant Membranes by Polyamines: Understanding Specific Modes of Action Under Stress. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef]
- Shabala, S.; Cuin, T.A.; Pottosin, I. Polyamine prevent NaCl-induced K+ efflux from pea mesophyll by blocking non-selective cation channels. FEBS Lett. 2007, 581, 1993–1999. [Google Scholar] [CrossRef] [Green Version]
- Nichols, C.G.; Sun-joo, L. Polyamines and potassium channels:A25-year romance. J. Biol. Chem. 2018, 293, 18779–18788. [Google Scholar] [CrossRef] [Green Version]
- Pál, M.; Tajti, J.; Szalai, G.; Peeva, V.; Végh, B.; Janda, T. Interaction of polyamines, abscisic acid and proline under osmotic stress in the leaves of wheat plants. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Moschou, P.N.; Paschalidis, K.A.; Roubelakis-Angelakis, K.A. Plant polyamine catabolism: The state of the art. Plant Signal. Behav. 2008, 3, 1061–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cona, A.; Rea, G.; Angelini, R.; Federico, R.; Tavladoraki, P. Functions of amine oxidases in plant development and defence. Trends Plant Sci. 2006, 11, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tan, C.; Cheng, X.; Zhao, X.; Li, T.; Jiang, J. miR168 targets Argonaute1A mediated miRNAs regulation pathways in response to potassium deficiency stress in tomato. BMC Plant Biol. 2020, 20, 1–17. [Google Scholar] [CrossRef]
- Fei, Y.; Xiao, B.; Yang, M.; Ding, Q.; Tang, W. MicroRNAs, polyamines, and the activities antioxidant enzymes are associated with in vitro rooting in white pine (Pinus strobus L.). Springerplus 2016, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vuosku, J.; Jokela, A.; Läärä, E.; Sääskilahti, M.; Muilu, R.; Sutela, S.; Altabella, T.; Sarjala, T.; Häggman, H. Consistency of polyamine profiles and expression of arginine decarboxylase in mitosis during zygotic embryogenesis of scots pine. Plant Physiol. 2006, 142, 1027–1038. [Google Scholar] [CrossRef] [Green Version]
- Vuosku, J.; Muilu-Mäkelä, R.; Avia, K.; Suokas, M.; Kestilä, J.; Läärä, E.; Häggman, H.; Savolainen, O.; Sarjala, T. Thermospermine Synthase (ACL5) and Diamine Oxidase (DAO) Expression Is Needed for Zygotic Embryogenesis and Vascular Development in Scots Pine. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Galloway, G.L.; Malmberg, R.L.; Price, R.A. Phylogenetic utility of the nuclear gene arginine decarboxylase: An example from brassicaceae. Mol. Biol. Evol. 1998, 15, 1312–1320. [Google Scholar] [CrossRef] [Green Version]
- Hummel, I.; Bourdais, G.; Gouesbet, G.; Couée, I.; Malmberg, R.L.; El Amrani, A. Differential gene expression of Arginine decarboxylase ADC1 and ADC2 in Arabidopsis thaliana: Characterization of transcriptional regulation during seed germination and seedling development. New Phytol. 2004, 163, 519–531. [Google Scholar] [CrossRef]
- Tavladoraki, P.; Cona, A.; Angelini, R. Copper-containing amine oxidases and FAD-dependent polyamine oxidases are key players in plant tissue differentiation and organ development. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Vuosku, J.; Karppinen, K.; Muilu-Mäkelä, R.; Kusano, T.; Sagor, G.H.M.; Avia, K.; Alakärppä, E.; Kestilä, J.; Suokas, M.; Nickolov, K.; et al. Scots pine aminopropyltransferases shed new light on evolution of the polyamine biosynthesis pathway in seed plants. Ann. Bot. 2018, 121, 1243–1256. [Google Scholar] [CrossRef] [Green Version]
- Minguet, E.G.; Vera-Sirera, F.; Marina, A.; Carbonell, J.; Blázquez, M.A. Evolutionary diversification in polyamine biosynthesis. Mol. Biol. Evol. 2008, 25, 2119–2128. [Google Scholar] [CrossRef] [Green Version]
- Urano, K.; Hobo, T.; Shinozaki, K. Arabidopsis ADC genes involved in polyamine biosynthesis areb essential for seed development. FEBS Lett. 2005, 579, 1557–1564. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Takahashi, Y.; Berberich, T.; Imai, A.; Miyazaki, A.; Takahashi, T.; Michael, A.; Kusano, T. The polyamine spermine protects against high salt stress in Arabidopsis thaliana. FEBS Lett. 2006, 580, 6783–6788. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Takahashi, Y.; Berberich, T.; Imai, A.; Takahashi, T.; Michael, A.J.; Kusano, T. A protective role for the polyamine spermine against drought stress in Arabidopsis. Biochem. Biophys. Res. Commun. 2006, 352, 486–490. [Google Scholar] [CrossRef]
- Muilu-Mäkelä, R.; Vuosku, J.; Läärä, E.; Saarinen, M.; Heiskanen, J.; Häggman, H.; Sarjala, T. Water availability influences morphology, mycorrhizal associations, PSII efficiency and polyamine metabolism at early growth phase of Scots pine seedlings. Plant Physiol. Biochem. 2015, 88, 70–81. [Google Scholar] [CrossRef]
- Sarjala, T.; Häggman, H.; Aronen, T. Effect of Exogenous Polyamines and Inhibitors of Polyamine Biosynthesis on Growth and Free Polyamine Contents of Embryogenic Scots Pine Callus. J. Plant Physiol. 1997, 150, 597–602. [Google Scholar] [CrossRef]
- Gupta, P.; Durzan, D. Shoot multiplication from mature trees of Douglas-fir (Pseudotsuga menziesii) and sugar pine (Pinus iambertiana). Plant Cell Rep. 1985, 4, 177–179. [Google Scholar] [CrossRef]
- Becwar, M.R.; Nagmani, R.; Wann, S.R. Initiation of embryogenic cultures and somatic embryo development in loblolly pine (Pinustaeda). Can. J. For. Res. 1990, 20, 810–817. [Google Scholar] [CrossRef]
- Towill, L.E.; Mazur, P. Studies on the reduction of 2,3,5-triphenyltetrazolium chloride as a viability assay for plant tissue cultures. Can. J. Bot. 1975, 53, 1097–1102. [Google Scholar] [CrossRef]
- Zapata, J.; Salinas, A.A.; Calderon, R.; Barcelo, R. Reduction of 2,3,5-triphenyltetrazolium chloride by the KCN-insensitive, salicylhydroxamic acid-sensitive alternative respiratory pathway of mitochondria from cultured grapevine cells. Plant Cell Rep. 1991, 10, 579–582. [Google Scholar] [CrossRef] [PubMed]
- Mikuła, A.; Niedzielski, M.; Rybczyński, J.J. The use of TTC reduction assay for assessment of Gentiana spp. cell suspension viability after cryopreservation. Acta Physiol. Plant. 2006, 28, 315–324. [Google Scholar] [CrossRef]
- Kalra, Y.P. Handbook of Reference Methods for Plant Analysis; Kalra, Y.P., Ed.; CRC Press, Taylor&Francis Group, LLC: Boca Raton, FL, USA, 1998; ISBN 9781574441246. [Google Scholar]
- Fornalé, S.; Sarjala, T.; Bagni, N. Endogenous polyamine content and metabolism in the ectomycorrhizal fungus Paxillus involutus. New Phytol. 1999, 143, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Chambers, J.M. Linear Models. In Statistical Models in S; Chambers, J.M., Hastie, T.J., Eds.; Wadsworth & Brooks/Cole Advanced Books and Software: Pacific Grove, CA, USA, 1992; Chapter 4; ISBN 0-534-16765-9. [Google Scholar]
- R Core Team R. A Language and Environment for Statistical Computing 2020; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 15 February 2021).
- Friendly, M. Corrgrams: Exploratory displays for correlatigon matrices. Am. Stat. 2002, 56, 316–324. [Google Scholar] [CrossRef]
- Muilu-Mäkelä, R.; Vuosku, J.; Saarinen, M.; Hamberg, L.; Ruotsalainen, S.; Häggman, H.; Sarjala, T. Coping with spring frost-effects on polyamine metabolism of scots pine seedlings. IForest 2017, 10, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Vuosku, J.; Sutela, S.; Kestilä, J.; Jokela, A.; Sarjala, T.; Häggman, H. Expression of catalase and retinoblastoma-related protein genes associates with cell death processes in Scots pine zygotic embryogenesis. BMC Plant Biol. 2015, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Yoon, J.H.; Shen, B. Search for apoptotic nucleases in yeast: Role of Tat-D nuclease in apoptotic DNA degradation. J. Biol. Chem. 2005, 280, 15370–15379. [Google Scholar] [CrossRef] [Green Version]
- Gutzat, R.; Borghi, L.; Gruissem, W. Emerging roles of RETINOBLASTOMA-RELATED proteins in evolution and plant development. Trends Plant Sci. 2012, 17, 139–148. [Google Scholar] [CrossRef]
- Hahlbrock, K.; Kuhlen, E. Relationship between growth of parsley and soybean cells in suspension cultures and changes in the conductivity of the culture medium. Planta 1972, 108, 271–278. [Google Scholar] [CrossRef]
- Adams, D.O.; Franke, K.E.; Christensen, L.P. Elevated putrescine levels in grapevine leaves that display symptoms of potassium deficiency. Am. J. Enol. Vitic. 1990, 41, 121–125. [Google Scholar]
- Okazaki, K.; Tanahashi, T.; Kato, Y.; Suzuki, I.; Tanaka, F.; Ohwaki, Y. Metabolic indices related to leaf marginal necrosis associated with potassium deficiency in tomato using GC/MS metabolite profiling. J. Biosci. Bioeng. 2020, 130, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Edfast, A.B.; Näsholm, T.; Aronsson, A.; Ericsson, A. Applications of mineral nutrients to heavily N-fertilized scots pine trees: Effects on arginine and mineral nutrient concentrations. Plant Soil 1996, 184, 57–65. [Google Scholar] [CrossRef]
- Takusagawa, F.; Kamitori, S.; Misaki, S.; Markham, G.D. chrystal Structure of S-Adenosylmethionine Synthetase. J. Biol. Chem. 1996, 271, 136–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Y.; Wu, Q.; Li, J.; Sun, S.; Sun, S. S-adenosylmethionine: A metabolite critical to the regulation of autophagy. Cell Prolif. 2020, 53. [Google Scholar] [CrossRef]
- Jensén, P. Effects of interrupted K+ supply on growth and uptake of K+, Ca2+, Mg2+ and Na+ in spring wheat. Physiol. Plant. 1982, 56, 259–265. [Google Scholar] [CrossRef]
- Luan, S.; Lan, W.; Lee, S.C. Potassium nutrition, sodium toxicity, and calcium signaling: Connections through the CBL-CIPK network. Curr. Opin. Plant Biol. 2009, 12, 339–346. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhong, M.; Du, N.; Shu, S.; Sun, J.; Guo, S. Putrescine enhances salt tolerance of cucumber seedlings by regulating ionhomeostasis. Environ. Exp. Bot. 2019, 165, 70–82. [Google Scholar] [CrossRef]
- Chen, Z.; Newman, I.; Zhou, M.; Mendham, N.; Zhang, G.; Shabala, S. Screening plants for salt tolerance by measuring K+ flux: A case study for barley. Plant Cell Environ. 2005, 28, 1230–1246. [Google Scholar] [CrossRef]
- Pottosin, I.; Velarde-Buendía, A.M.; Bose, J.; Fuglsang, A.T.; Shabala, S. Polyamines cause plasma membrane depolarization, activate Ca-, and modulate H-ATPase pump activity in pea roots. J. Exp. Bot. 2014, 65, 2463–2472. [Google Scholar] [CrossRef] [Green Version]
- Muilu-Mäkelä, R.; Vuosku, J.; Hamberg, L.; Latva-Mäenpää, H.; Häggman, H.; Sarjala, T. Osmotic stress affects polyamine homeostasis and phenolic content in proembryogenic liquid cell cultures of Scots pine. Plant Cell. Tissue Organ Cult. 2015, 122, 709–726. [Google Scholar] [CrossRef]
- Salo, H.M.; Sarjala, T.; Jokela, A.; Häggman, H.; Vuosku, J. Moderate stress responses and specific changes in polyamine metabolism characterize Scots pine somatic embryogenesis. Tree Physiol. 2016, 36, 392–402. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.M.; Skalicky, M.; Jahan, M.S.; Hossain, M.N.; Anwar, Z.; Nie, Z.F.; Alabdallah, N.M.; Brestic, M.; Hejnak, V.; Fang, X.W. Spermine: Its Emerging Role in Regulating Drought Stress Responses in Plants. Cells 2021, 10, 261. [Google Scholar] [CrossRef]
- Toumi, I.; Moschou, P.N.; Paschalidis, K.A.; Bouamama, B.; Salem-fnayou, A.B.; Ghorbel, A.W.; MLiki, A.; Roubelakis-Angelakis, K.A. Abscisic acid signals reorientation of polyamine metabolism to orchestratestress responses via the polyamine exodus pathway in grapevine. J. Plant Physiol. 2010, 167, 519–525. [Google Scholar] [CrossRef]
- Liu, K.; Fu, H.; Bei, Q.; Luan, S. Inward potassium channel in guard cells as a target for polyamine regulation of stomatal movements. Plant Physiol. 2000, 124, 1315–1325. [Google Scholar] [CrossRef] [Green Version]
- De Diego, N.; Sainz-Fernández, I.; Rodríques, J.L.; Pérez-Alfocea, P.; Sampedro, M.C.; Barrio, R.J.; Lacuesta, M.; Moncaleán, P. Metabolites and hormones are involved in the intraspecific variability of drought hardening in radiata pine. J. Plant Physiol. 2015, 188, 64–71. [Google Scholar] [CrossRef]
- Mittler, R. ROS Are Good. Trend Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Ródenas, R.; Lara, A.; Martínez, V.; Rubio, F. The combination of K + deficiency with other environmental stresses: What is the outcome? Physiol. Plant. 2019, 165, 264–276. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Cao, B.; Qi, L.; Yin, L.; Wang, S.; Deng, X. Silicon-moderated K-deficiency-induced leaf chlorosis by decreasing putrescine accumulation in sorghum. Ann. Bot. 2016, 118, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenach, C.; De Angeli, A. Ion transport at the vacuole during stomatal movements. Plant Physiol. 2017, 174, 520–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | Cond (μS/cm) | ||
|---|---|---|---|---|
| mean | range | mean | range | |
| 0% K | 3.5 | 3.4–3.5 | 1.6 | 1.4–1.9 |
| 50% K | 3.7 | 3.6–3.8 | 1.5 | 1.0–1.8 |
| 100% K | 4.4 | 3.8–5.6 | 1.4 | 1.0–1.7 |
| 150% K | 4.9 | 4.1–5.6 | 1.6 | 1.3–2.2 |
| PA Gene Expression | Free PA Content | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatment | Plant Material | ADC | SPDS | ACL5 | DAO | PAO | Put | Spd | Spm/ t-Spm |
| Potassium deficiency in liquid cell cultures | Proembryogenic cells | ⇧ | ⇩ | ⇩ | ⇩ | ⇧ | |||
| Potassium deficiency [4] | Needles | nd | nd | nd | nd | nd | ⇧ | ⇩ | ⇩ |
| Induction of osmotic stress by PEG treatment in liquid cell cultures [63] | Proembryogenic cells | ⇩ | ⇩ | ⇧ | |||||
| Triggering of SE by ABA + PEG treatment [64] | Embryogenic cells | ⇩ | ⇩ | ⇩ | ⇩ | nd | |||
| Transferring to culture medium with ABA and without PEG | ⇧ | ⇧ | ⇧ | ⇧ | nd | ||||
| Exposure of seedlings to drought stress [36] | Needles | ⇩ | ⇧ | ||||||
| Stems | ⇩ | ⇩ | ⇩ | ||||||
| Roots | ⇧ | ⇩ | ⇧ | ⇧ | |||||
| Exposure of seedlings to spring frost [48] | Needles | ⇩ | ⇩ | ||||||
| Roots | ⇧ | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muilu-Mäkelä, R.; Vuosku, J.; Häggman, H.; Sarjala, T. Polyamine Metabolism in Scots Pine Embryogenic Cells under Potassium Deficiency. Cells 2021, 10, 1244. https://doi.org/10.3390/cells10051244
Muilu-Mäkelä R, Vuosku J, Häggman H, Sarjala T. Polyamine Metabolism in Scots Pine Embryogenic Cells under Potassium Deficiency. Cells. 2021; 10(5):1244. https://doi.org/10.3390/cells10051244
Chicago/Turabian StyleMuilu-Mäkelä, Riina, Jaana Vuosku, Hely Häggman, and Tytti Sarjala. 2021. "Polyamine Metabolism in Scots Pine Embryogenic Cells under Potassium Deficiency" Cells 10, no. 5: 1244. https://doi.org/10.3390/cells10051244
APA StyleMuilu-Mäkelä, R., Vuosku, J., Häggman, H., & Sarjala, T. (2021). Polyamine Metabolism in Scots Pine Embryogenic Cells under Potassium Deficiency. Cells, 10(5), 1244. https://doi.org/10.3390/cells10051244