
IL-10 Mediated Immunomodulation Limits Subepithelial Fibrosis and Repairs Airway Epithelium in Rejecting Airway Allografts

 , and
, and
Abstract
:1. Background
2. Materials and Methods
2.1. Mice Strains
2.2. Experimental Planning
2.3. Surgical Procedure
2.4. IL-10 Depletion (−) and Reconstitution (+)
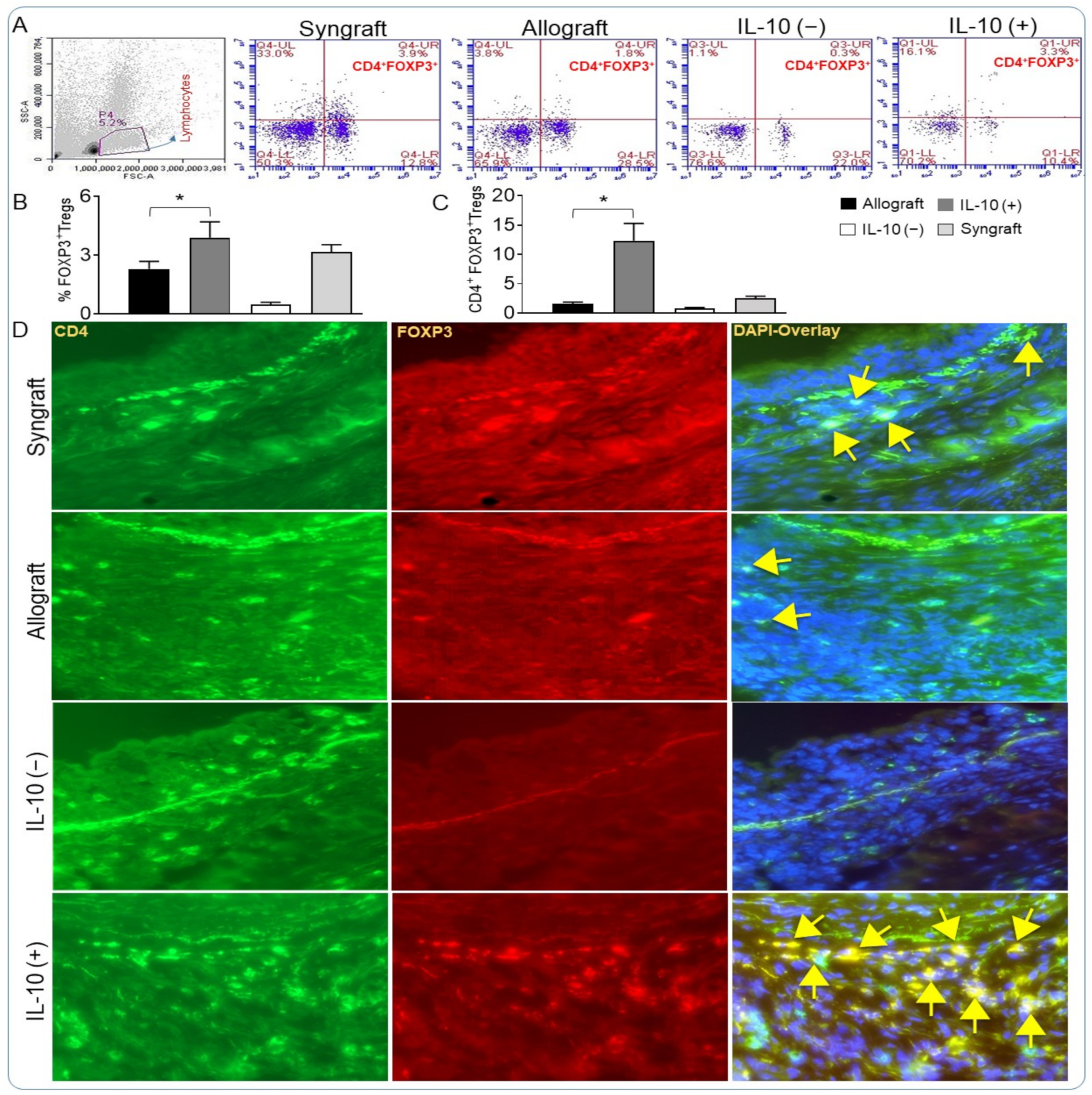
2.5. Analysis of Regulatory T Cells
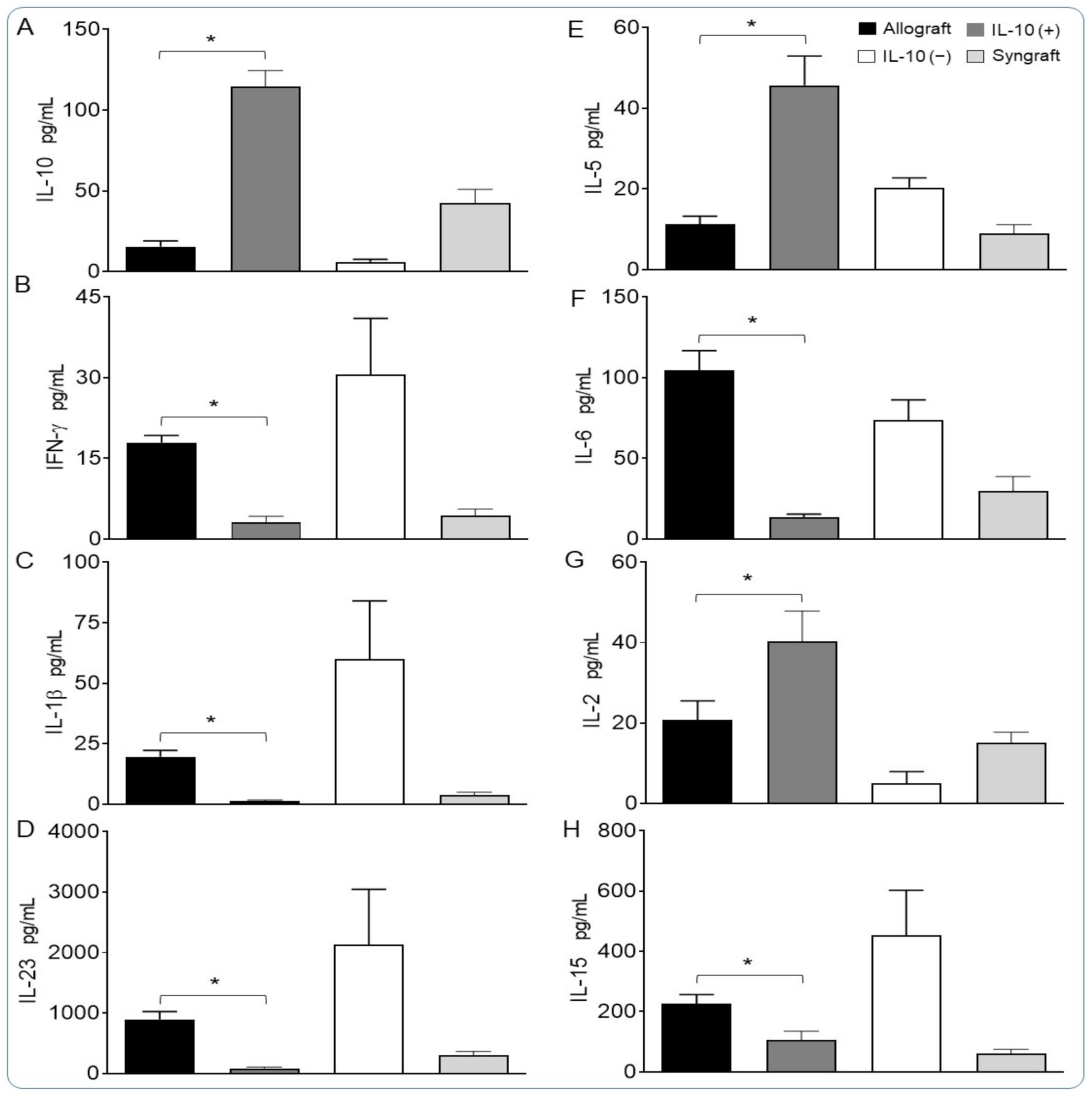
2.6. Analysis of Serum Cytokines
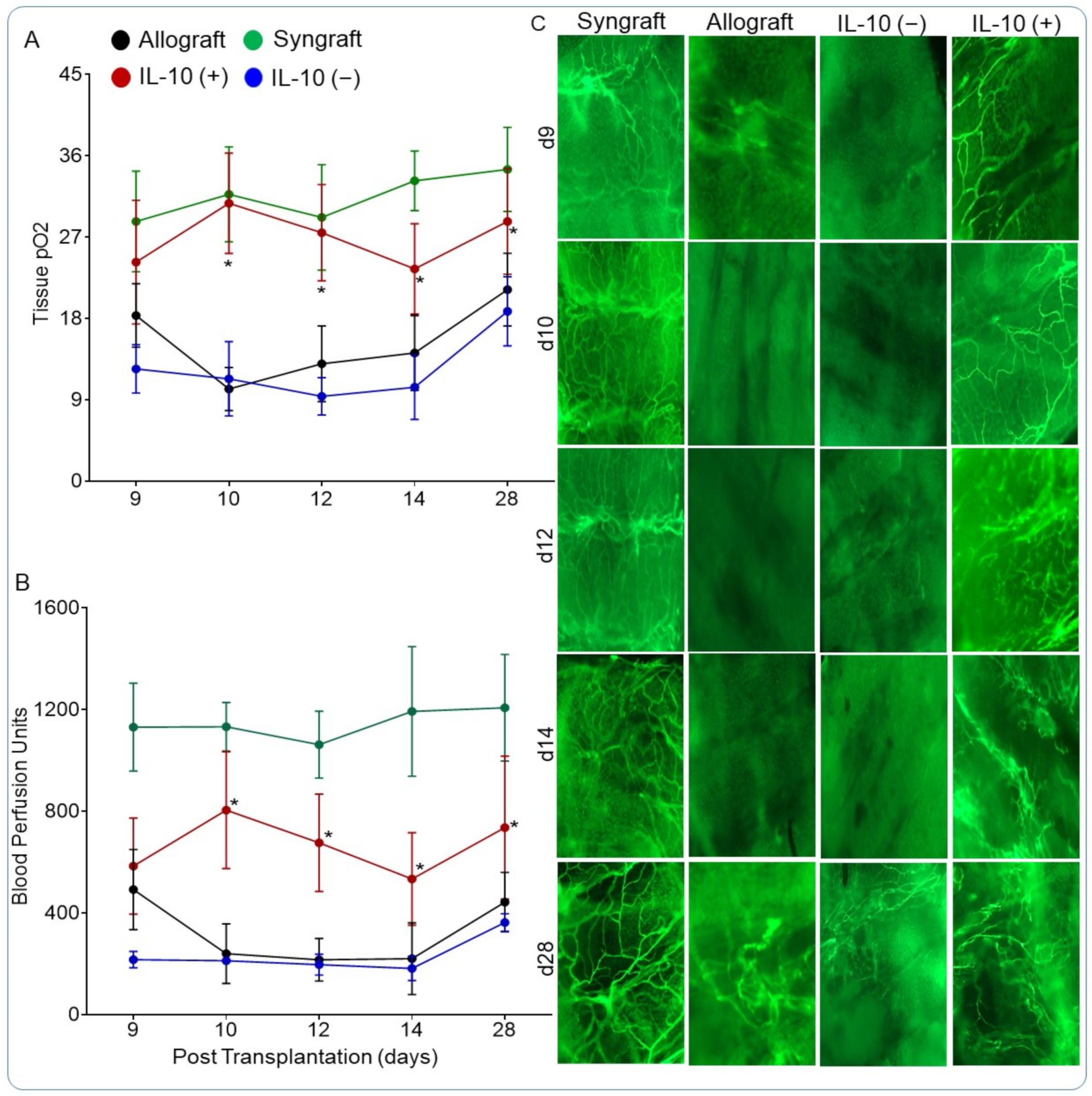
2.7. Analysis of Graft Blood Flow, Oxygenation, and Microvasculature
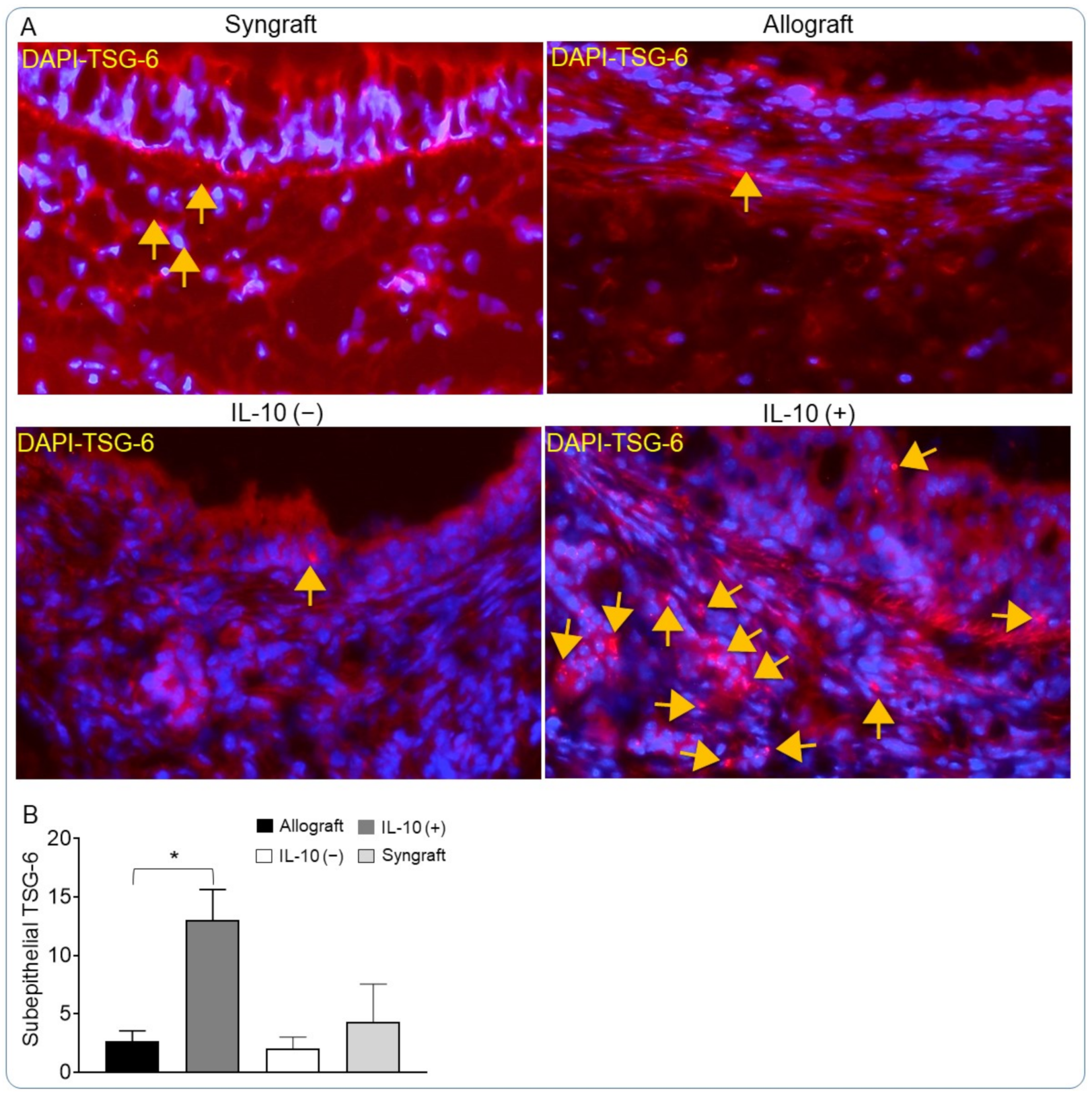
2.8. Analysis of Graft Epithelium and Collagen Deposition
2.9. Statistical Analysis
3. Results
3.1. IL-10 Is Sufficient to Establish Immunotolerance
3.2. IL-10 Is Sufficient to Augment TSG-6 Deposition
3.3. IL-10 Is Sufficient to Suppress Pro-Inflammatory Cytokines
3.4. IL-10 Is Sufficient to Restore Graft Oxygenation and Microvascular Blood Flow
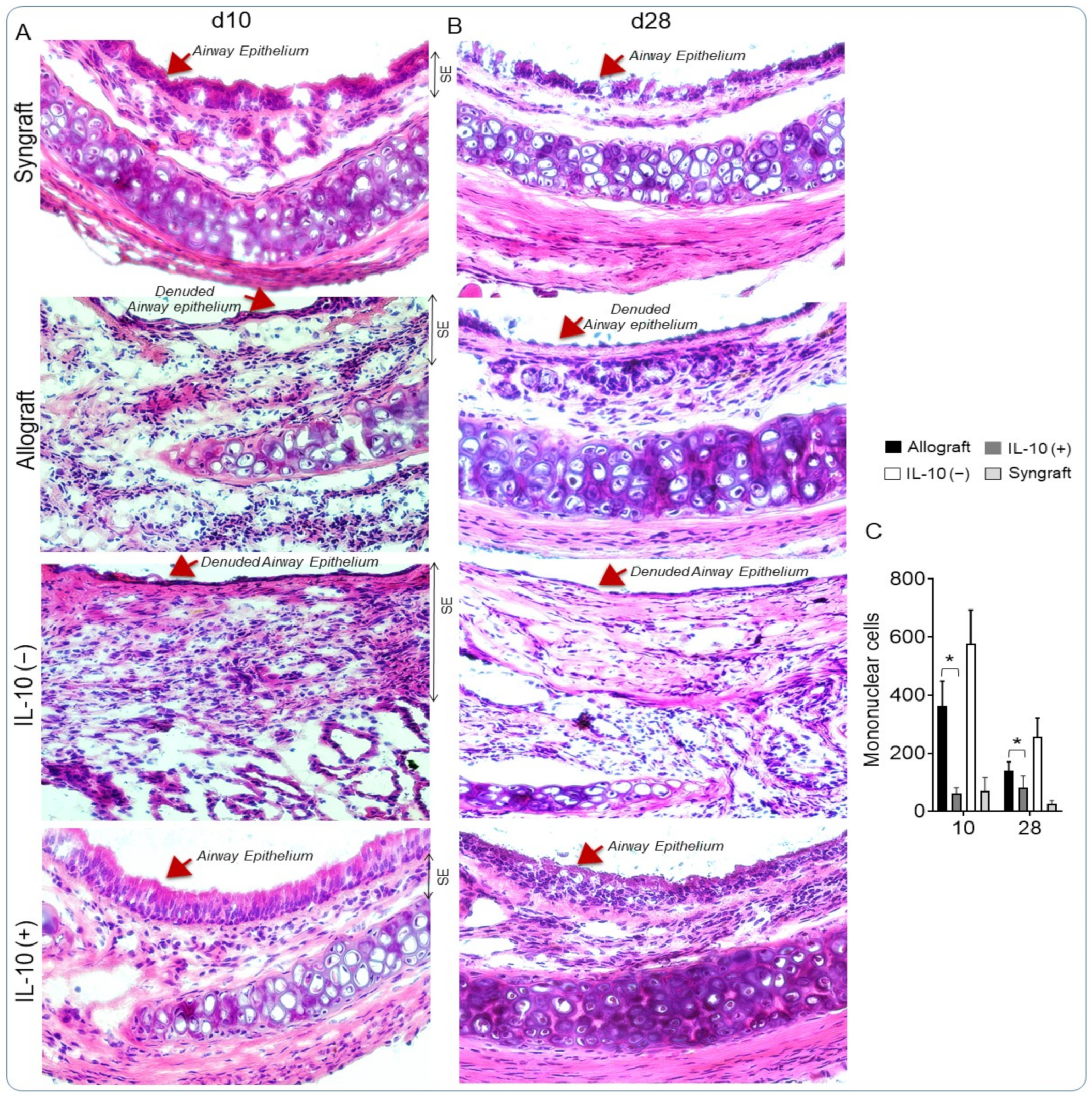
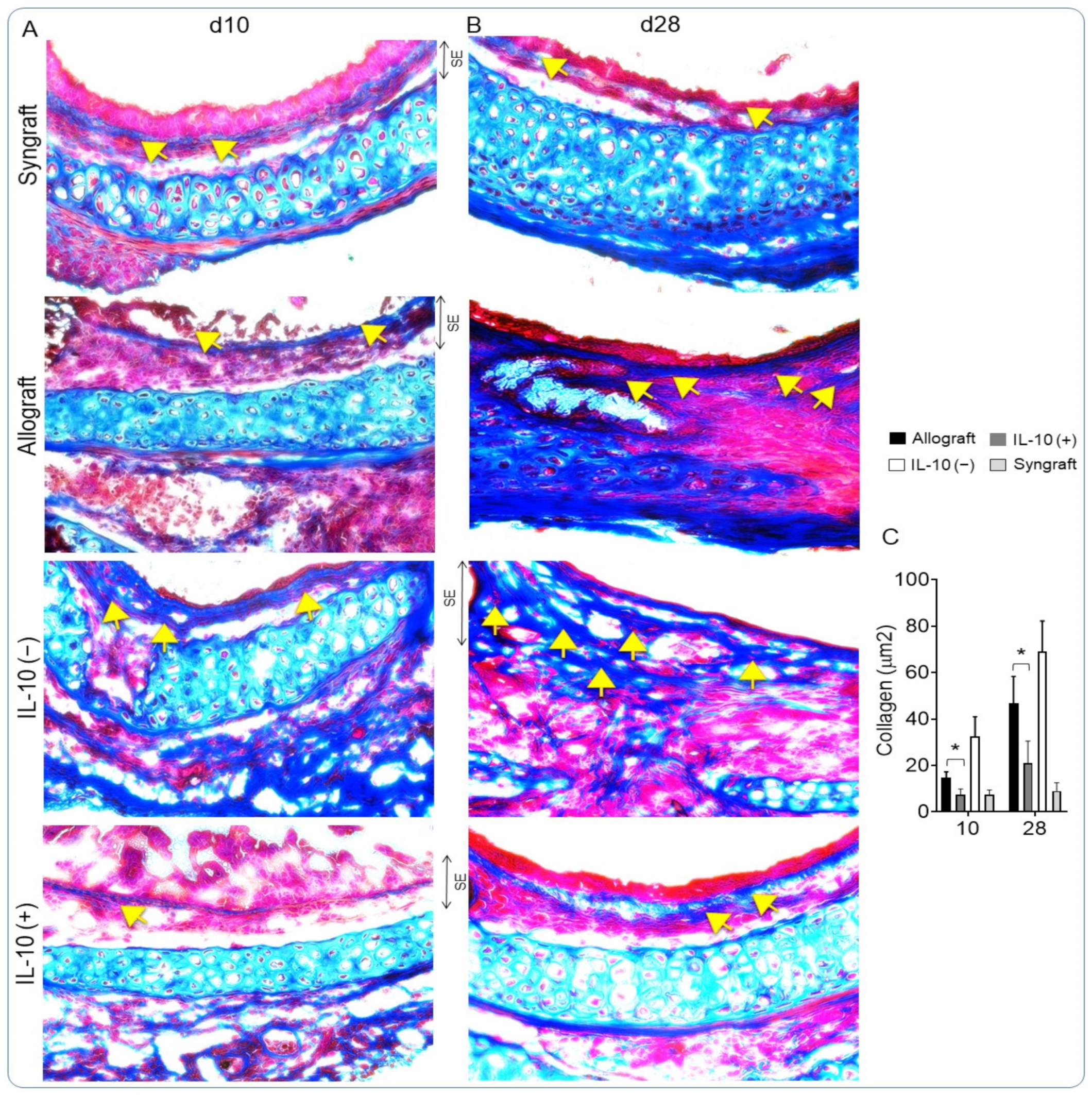
3.5. IL-10 Is Sufficient to Augment Airway Epithelial Repair and Subdue Fibrosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Babu, A.N.; Murakawa, T.; Thurman, J.M.; Miller, E.J.; Henson, P.M.; Zamora, M.R.; Voelkel, N.F.; Nicolls, M.R. Microvascular destruction identifies murine allografts that cannot be rescued from airway fibrosis. J. Clin. Investig. 2007, 117, 3774–3785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doreille, A.; Dieudé, M.; Cardinal, H. The determinants, biomarkers, and consequences of microvascular injury in kidney transplant recipients. Am. J. Physiol. Ren. Physiol. 2019, 316, F9–F19. [Google Scholar] [CrossRef] [PubMed]
- Heim, C.; Khan, M.A.; von Silva-Tarouca, B.; Kuckhahn, A.; Stamminger, T.; Ramsperger-Gleixner, M.; Nicolls, M.R.; Weyand, M.; Ensminger, S.M. Preservation of Microvascular Integrity in Murine Orthotopic Tracheal Allografts by Clopidogrel. Transplantation 2019, 103, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Nguyen, T.T.; Tian, W.; Sung, Y.K.; Yuan, K.; Qian, J.; Rajadas, J.; Sallenave, J.M.; Nickel, N.P.; de Jesus Perez, V.; et al. Cyclosporine Does Not Prevent Microvascular Loss in Transplantation but Can Synergize With a Neutrophil Elastase Inhibitor, Elafin, to Maintain Graft Perfusion During Acute Rejection. Am. J. Transplant. 2015. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Sung, Y.K.; Tian, W.; Qian, J.; Semenza, G.L.; Nicolls, M.R. Graft microvascular disease in solid organ transplantation. J. Mol. Med. (Berl) 2014, 92, 797–810. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Shamma, T.; Kazmi, S.; Altuhami, A.; Ahmed, H.A.; Assiri, A.M.; Broering, D.C. Hypoxia-induced complement dysregulation is associated with microvascular impairments in mouse tracheal transplants. J. Transl. Med. 2020, 18, 147. [Google Scholar] [CrossRef]
- Luckraz, H.; Goddard, M.; McNeil, K.; Atkinson, C.; Charman, S.C.; Stewart, S.; Wallwork, J. Microvascular changes in small airways predispose to obliterative bronchiolitis after lung transplantation. J. Heart Lung Transplant. 2004, 23, 527–531. [Google Scholar] [CrossRef]
- Luckraz, H.; Goddard, M.; McNeil, K.; Atkinson, C.; Sharples, L.D.; Wallwork, J. Is obliterative bronchiolitis in lung transplantation associated with microvascular damage to small airways? Ann. Thorac. Surg. 2006, 82, 1212–1218. [Google Scholar] [CrossRef]
- Nicolls, M.R.; Voelkel, N.F. Hypoxia and the lung: Beyond hypoxic vasoconstriction. Antioxid. Redox Signal. 2007, 9, 741–743. [Google Scholar] [CrossRef] [Green Version]
- Nicolls, M.R.; Zamora, M.R. Bronchial blood supply after lung transplantation without bronchial artery revascularization. Curr. Opin. Organ Transplant. 2010, 15, 563–567. [Google Scholar] [CrossRef] [Green Version]
- Wilkes, D.S. Chronic lung allograft rejection and airway microvasculature: Is HIF-1 the missing link? J. Clin. Investig. 2011, 121, 2155–2157. [Google Scholar] [CrossRef]
- Khan, M.A.; Alanazi, F.; Ahmed, H.A.; Al-Mohanna, F.H.; Assiri, A.M.; Broering, D.C. FOXP3+ regulatory T cell ameliorates microvasculature in the rejection of mouse orthotopic tracheal transplants. Clin. Immunol. 2017, 174, 84–98. [Google Scholar] [CrossRef]
- Ferreira, L.M.R.; Muller, Y.D.; Bluestone, J.A.; Tang, Q. Next-generation regulatory T cell therapy. Nat. Rev. Drug Discov. 2019, 18, 749–769. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Zhang, M.; Li, J.; Yang, M.; Liu, Y.; Guo, X.; Li, H.; Liu, Z.; Zhao, J. Circulating FoxP3+ Regulatory T and Interleukin17-Producing Th17 Cells Actively Influence HBV Clearance in De Novo Hepatitis B Virus Infected Patients after Orthotopic Liver Transplantation. PLoS ONE 2015, 10, e0137881. [Google Scholar] [CrossRef] [Green Version]
- Ge, W.; Jiang, J.; Liu, W.; Lian, D.; Saito, A.; Garcia, B.; Li, X.C.; Wang, H. Regulatory T cells are critical to tolerance induction in presensitized mouse transplant recipients through targeting memory T cells. Am. J. Transplant. 2010, 10, 1760–1773. [Google Scholar] [CrossRef]
- Horch, M.; Nguyen, V.H. Regulatory T-cell immunotherapy for allogeneic hematopoietic stem-cell transplantation. Ther. Adv. Hematol. 2012, 3, 29–44. [Google Scholar] [CrossRef] [Green Version]
- Issa, F.; Wood, K.J. CD4+ regulatory T cells in solid organ transplantation. Curr. Opin. Organ Transplant. 2010, 15, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Lechler, R.I.; He, X.-S.; Huang, J.-F. Regulatory T cells and transplantation tolerance. Hum. Immunol. 2006, 67, 765–776. [Google Scholar] [CrossRef] [PubMed]
- June, C.H.; Blazar, B.R. Clinical application of expanded CD4+25+ cells. Semin. Immunol. 2006, 18, 78–88. [Google Scholar] [CrossRef]
- Li, W.; Carper, K.; Liang, Y.; Zheng, X.X.; Kuhr, C.S.; Reyes, J.D.; Perkins, D.L.; Thomson, A.W.; Perkins, J.D. Anti-CD25 mAb administration prevents spontaneous liver transplant tolerance. Transplant. Proc. 2006, 38, 3207–3208. [Google Scholar] [CrossRef]
- Akimova, T.; Xiao, H.; Liu, Y.; Bhatti, T.R.; Jiao, J.; Eruslanov, E.; Singhal, S.; Wang, L.; Han, R.; Zacharia, K.; et al. Targeting sirtuin-1 alleviates experimental autoimmune colitis by induction of Foxp3+ T-regulatory cells. Mucosal Immunol. 2014, 7, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Jäger, A.; Kuchroo, V.K. Effector and regulatory T-cell subsets in autoimmunity and tissue inflammation. Scand. J. Immunol. 2010, 72, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Oberle, N.; Krammer, P.H. Molecular mechanisms of treg-mediated T cell suppression. Front. Immunol. 2012, 3, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhry, A.; Samstein, R.M.; Treuting, P.; Liang, Y.; Pils, M.C.; Heinrich, J.M.; Jack, R.S.; Wunderlich, F.T.; Bruning, J.C.; Muller, W.; et al. Interleukin-10 signaling in regulatory T cells is required for suppression of Th17 cell-mediated inflammation. Immunity 2011, 34, 566–578. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Kingsley, C.I.; Niimi, M.; Read, S.; Turvey, S.E.; Bushell, A.R.; Morris, P.J.; Powrie, F.; Wood, K.J. IL-10 is required for regulatory T cells to mediate tolerance to alloantigens in vivo. J. Immunol. 2001, 166, 3789–3796. [Google Scholar] [CrossRef]
- Ng, T.H.S.; Britton, G.; Hill, E.; Verhagen, J.; Burton, B.; Wraith, D. Regulation of Adaptive Immunity; The Role of Interleukin-10. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Böhm, L.; Maxeiner, J.; Meyer-Martin, H.; Reuter, S.; Finotto, S.; Klein, M.; Schild, H.; Schmitt, E.; Bopp, T.; Taube, C. IL-10 and Regulatory T Cells Cooperate in Allergen-Specific Immunotherapy To Ameliorate Allergic Asthma. J. Immunol. 2015, 194, 887–897. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.; Santner-Nanan, B.; Hu, M.; Skarratt, K.; Lee, C.H.; Stormon, M.; Wong, M.; Fuller, S.J.; Nanan, R. IL-10 Potentiates Differentiation of Human Induced Regulatory T Cells via STAT3 and Foxo1. J. Immunol. 2015, 195, 3665–3674. [Google Scholar] [CrossRef] [Green Version]
- Bharat, A.; Fields, R.C.; Trulock, E.P.; Patterson, G.A.; Mohanakumar, T. Induction of IL-10 suppressors in lung transplant patients by CD4+25+ regulatory T cells through CTLA-4 signaling. J. Immunol. 2006, 177, 5631–5638. [Google Scholar] [CrossRef] [Green Version]
- Brockmann, L.; Soukou, S.; Steglich, B.; Czarnewski, P.; Zhao, L.; Wende, S.; Bedke, T.; Ergen, C.; Manthey, C.; Agalioti, T.; et al. Molecular and functional heterogeneity of IL-10-producing CD4(+) T cells. Nat. Commun. 2018, 9, 5457. [Google Scholar] [CrossRef] [Green Version]
- Cypel, M.; Liu, M.; Rubacha, M.; Yeung, J.C.; Hirayama, S.; Anraku, M.; Sato, M.; Medin, J.; Davidson, B.L.; de Perrot, M.; et al. Functional Repair of Human Donor Lungs by IL-10 Gene Therapy. Sci. Transl. Med. 2009, 1, 4ra9. [Google Scholar] [CrossRef]
- DeBruyne, L.A.; Li, K.; Chan, S.Y.; Qin, L.; Bishop, D.K.; Bromberg, J.S. Lipid-mediated gene transfer of viral IL-10 prolongs vascularized cardiac allograft survival by inhibiting donor-specific cellular and humoral immune responses. Gene Ther. 1998, 5, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Murai, M.; Turovskaya, O.; Kim, G.; Madan, R.; Karp, C.L.; Cheroutre, H.; Kronenberg, M. Interleukin 10 acts on regulatory T cells to maintain expression of the transcription factor Foxp3 and suppressive function in mice with colitis. Nat. Immunol. 2009, 10, 1178–1184. [Google Scholar] [CrossRef]
- Nitta, Y.; Tashiro, F.; Tokui, M.; Shimada, A.; Takei, I.; Tabayashi, K.; Miyazaki, J. Systemic delivery of interleukin 10 by intramuscular injection of expression plasmid DNA prevents autoimmune diabetes in nonobese diabetic mice. Hum. Gene Ther. 1998, 9, 1701–1707. [Google Scholar] [CrossRef]
- Niu, J.; Yue, W.; Song, Y.; Zhang, Y.; Qi, X.; Wang, Z.; Liu, B.; Shen, H.; Hu, X. Prevention of acute liver allograft rejection by IL-10-engineered mesenchymal stem cells. Clin. Exp. Immunol. 2014, 176, 473–484. [Google Scholar] [CrossRef]
- Branchett, W.J.; Lloyd, C.M. Regulatory cytokine function in the respiratory tract. Mucosal Immunol. 2019, 12, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Steen, E.H.; Wang, X.; Balaji, S.; Butte, M.J.; Bollyky, P.L.; Keswani, S.G. The Role of the Anti-Inflammatory Cytokine Interleukin-10 in Tissue Fibrosis. Adv. Wound Care (New Rochelle) 2020, 9, 184–198. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Alanazi, F.; Ahmed, H.A.; Shamma, T.; Kelly, K.; Hammad, M.A.; Alawad, A.O.; Assiri, A.M.; Broering, D.C. iPSC-derived MSC therapy induces immune tolerance and supports long-term graft survival in mouse orthotopic tracheal transplants. Stem Cell Res. Ther. 2019, 10, 290. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Alanazi, F.; Ahmed, H.A.; Vater, A.; Assiri, A.M.; Broering, D.C. C5a Blockade Increases Regulatory T Cell Numbers and Protects Against Microvascular Loss and Epithelial Damage in Mouse Airway Allografts. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]
- Khan, M.A.; Jiang, X.; Dhillon, G.; Beilke, J.; Holers, V.M.; Atkinson, C.; Tomlinson, S.; Nicolls, M.R. CD4+ T cells and complement independently mediate graft ischemia in the rejection of mouse orthotopic tracheal transplants. Circ. Res. 2011, 109, 1290–1301. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Maasch, C.; Vater, A.; Klussmann, S.; Morser, J.; Leung, L.L.; Atkinson, C.; Tomlinson, S.; Heeger, P.S.; Nicolls, M.R. Targeting complement component 5a promotes vascular integrity and limits airway remodeling. Proc. Natl. Acad. Sci. USA 2013, 110, 6061–6066. [Google Scholar] [CrossRef] [Green Version]
- Boehler, A. The role of interleukin-10 in lung transplantation. Transplant. Immunol. 2002, 9, 121–124. [Google Scholar] [CrossRef]
- Battaglia, M.; Stabilini, A.; Draghici, E.; Gregori, S.; Mocchetti, C.; Bonifacio, E.; Roncarolo, M.G. Rapamycin and interleukin-10 treatment induces T regulatory type 1 cells that mediate antigen-specific transplantation tolerance. Diabetes 2006, 55, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Dhillon, G.; Jiang, X.; Lin, Y.C.; Nicolls, M.R. New Methods for Monitoring Dynamic Airway Tissue Oxygenation and Perfusion in Experimental and Clinical Transplantation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L861–L869. [Google Scholar] [CrossRef] [Green Version]
- Komai, T.; Inoue, M.; Okamura, T.; Morita, K.; Iwasaki, Y.; Sumitomo, S.; Shoda, H.; Yamamoto, K.; Fujio, K. Transforming Growth Factor-beta and Interleukin-10 Synergistically Regulate Humoral Immunity via Modulating Metabolic Signals. Front. Immunol. 2018, 9, 1364. [Google Scholar] [CrossRef]
- Goto, Y.; Lamichhane, A.; Kamioka, M.; Sato, S.; Honda, K.; Kunisawa, J.; Kiyono, H. IL-10-producing CD4(+) T cells negatively regulate fucosylation of epithelial cells in the gut. Sci. Rep. 2015, 5, 15918. [Google Scholar] [CrossRef] [Green Version]
- Kota, D.J.; Wiggins, L.L.; Yoon, N.; Lee, R.H. TSG-6 produced by hMSCs delays the onset of autoimmune diabetes by suppressing Th1 development and enhancing tolerogenicity. Diabetes 2013, 62, 2048–2058. [Google Scholar] [CrossRef] [Green Version]
- Hsu, J.L.; Jiang, X.; Khan, M.A.; Sobel, R.A.; Clemons, K.V.; Stevens, D.A.; Nicolls, M.R. Aspergillus Invasion Increases with Progressive Airway Ischemia. Ann. Am. Thorac. Soc. 2014, 11, S79. [Google Scholar] [CrossRef]
- Fehervari, Z.; Sakaguchi, S. CD4+ Tregs and immune control. J. Clin. Investig. 2004, 114, 1209–1217. [Google Scholar] [CrossRef]
- Picarda, E.; Bezie, S.; Usero, L.; Ossart, J.; Besnard, M.; Halim, H.; Echasserieau, K.; Usal, C.; Rossjohn, J.; Bernardeau, K.; et al. Cross-Reactive Donor-Specific CD8(+) Tregs Efficiently Prevent Transplant Rejection. Cell Rep. 2019, 29, 4245.e4246–4255.e4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.; Lee, R.H.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. Anti-inflammatory protein TSG-6 secreted by activated MSCs attenuates zymosan-induced mouse peritonitis by decreasing TLR2/NF-kappaB signaling in resident macrophages. Blood 2011, 118, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kui, L.; Chan, G.C.; Lee, P.P. TSG-6 Downregulates IFN-Alpha and TNF-Alpha Expression by Suppressing IRF7 Phosphorylation in Human Plasmacytoid Dendritic Cells. Mediators Inflamm. 2017, 2017, 7462945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Song, H.; Duan, H.; Chai, J.; Yang, J.; Li, X.; Yu, Y.; Zhang, X.; Hu, X.; Xiao, M.; et al. TSG-6 secreted by human umbilical cord-MSCs attenuates severe burn-induced excessive inflammation via inhibiting activations of P38 and JNK signaling. Sci. Rep. 2016, 6, 30121. [Google Scholar] [CrossRef]
- Milner, C.M.; Day, A.J. TSG-6: A multifunctional protein associated with inflammation. J. Cell Sci. 2003, 116, 1863–1873. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.; Verhagen, J.; Blaser, K.; Akdis, M.; Akdis, C.A. Mechanisms of immune suppression by interleukin-10 and transforming growth factor-beta: The role of T regulatory cells. Immunology 2006, 117, 433–442. [Google Scholar] [CrossRef]
- Khan, M.A.; Hsu, J.L.; Assiri, A.M.; Broering, D.C. Targeted complement inhibition and microvasculature in transplants: A therapeutic perspective. Clin. Exp. Immunol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Belperio, J.A.; Weigt, S.S.; Fishbein, M.C.; Lynch, J.P., 3rd. Chronic lung allograft rejection: Mechanisms and therapy. Proc. Am. Thorac. Soc. 2009, 6, 108–121. [Google Scholar] [CrossRef]
- Nicolls, M.R.; Hsu, J.L.; Jiang, X. Microvascular injury after lung transplantation. Curr. Opin. Organ Transplant. 2016, 21, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Ji, L.; Wang, W.; Hua, F.; Zhan, Y.; Zou, S.; Yuan, L.; Ke, Y.; Min, Z.; Song, D.; et al. Insufficient secretion of IL-10 by Tregs compromised its control on over-activated CD4+ T effector cells in newly diagnosed adult immune thrombocytopenia patients. Immunol. Res. 2015, 61, 269–280. [Google Scholar] [CrossRef]
- Liu, K.S.; Fan, X.Q.; Zhang, L.; Wen, Q.N.; Feng, J.H.; Chen, F.C.; Luo, J.M.; Sun, W.B. Effects of recombinant human interleukin-10 on Treg cells, IL-10 and TGF-beta in transplantation of rabbit skin. Mol. Med. Rep. 2014, 9, 639–644. [Google Scholar] [CrossRef] [Green Version]
- O’Garra, A.; Vieira, P.L.; Vieira, P.; Goldfeld, A.E. IL-10–producing and naturally occurring CD4+ Tregs: Limiting collateral damage. J. Clin. Investig. 2004, 114, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Martins, S.; de Perrot, M.; Imai, Y.; Yamane, M.; Quadri, S.M.; Segall, L.; Dutly, A.; Sakiyama, S.; Chaparro, A.; Davidson, B.L.; et al. Transbronchial administration of adenoviral-mediated interleukin-10 gene to the donor improves function in a pig lung transplant model. Gene Ther. 2004, 11, 1786–1796. [Google Scholar] [CrossRef] [Green Version]
- Sendra Gisbert, L.; Miguel Matas, A.; Sabater Orti, L.; Herrero, M.J.; Sabater Olivas, L.; Montalva Oron, E.M.; Frasson, M.; Abargues Lopez, R.; Lopez-Andujar, R.; Garcia-Granero Ximenez, E.; et al. Efficacy of hydrodynamic interleukin 10 gene transfer in human liver segments with interest in transplantation. Liver Transpl. 2017, 23, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Pierog, J.; Tamo, L.; Fakin, R.; Kocher, G.; Gugger, M.; Grodzki, T.; Geiser, T.; Gazdhar, A.; Schmid, R.A. Bone marrow stem cells modified with human interleukin 10 attenuate acute rejection in rat lung allotransplantation. Eur. J. Cardiothorac. Surg. 2018, 53, 194–200. [Google Scholar] [CrossRef]
- Nicod, L.P. Mechanisms of Airway Obliteration after Lung Transplantation. Proc. Amer. Thorac. Soc. 2006, 3, 444–449. [Google Scholar] [CrossRef]
- Sato, M.; Keshavjee, S. Bronchiolitis obliterans syndrome: Alloimmune-dependent and -independent injury with aberrant tissue remodeling. Semin. Thorac. Cardiovasc. Surg. 2008, 20, 173–182. [Google Scholar] [CrossRef]
- Dhillon, G.S.; Zamora, M.R.; Roos, J.E.; Sheahan, D.; Sista, R.R.; Van der Starre, P.; Weill, D.; Nicolls, M.R. Lung transplant airway hypoxia: A diathesis to fibrosis? Am. J. Respir. Crit. Care Med. 2010, 182, 230–236. [Google Scholar] [CrossRef]
- Fernandez, I.E.; Eickelberg, O. New cellular and molecular mechanisms of lung injury and fibrosis in idiopathic pulmonary fibrosis. Lancet 2012, 380, 680–688. [Google Scholar] [CrossRef]
- Jeffery, P.K. Remodeling in asthma and chronic obstructive lung disease. Am. J. Respir. Crit. Care Med. 2001, 164, S28–S38. [Google Scholar] [CrossRef] [PubMed]
- Todd, N.W.; Luzina, I.G.; Atamas, S.P. Molecular and cellular mechanisms of pulmonary fibrosis. Fibrogenesis Tissue Repair 2012, 5, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.A.; Nicolls, M.R. Complement-mediated microvascular injury leads to chronic rejection. Adv. Exp. Med. Biol. 2013, 734, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Croasdell Lucchini, A.; Gachanja, N.N.; Rossi, A.G.; Dorward, D.A.; Lucas, C.D. Epithelial Cells and Inflammation in Pulmonary Wound Repair. Cells 2021, 10, 339. [Google Scholar] [CrossRef]
- Zhang, S.B.; Sun, X.; Wu, Q.; Wu, J.P.; Chen, H.Y. Impaired Capacity of Fibroblasts to Support Airway Epithelial Progenitors in Bronchiolitis Obliterans Syndrome. Chin. Med. J. (Engl.) 2016, 129, 2040–2044. [Google Scholar] [CrossRef]
- Dace, D.S.; Khan, A.A.; Kelly, J.; Apte, R.S. Interleukin-10 Promotes Pathological Angiogenesis by Regulating Macrophage Response to Hypoxia during Development. PLoS ONE 2008, 3, e3381. [Google Scholar] [CrossRef]
- Deng, B.; Wehling-Henricks, M.; Villalta, S.A.; Wang, Y.; Tidball, J.G. IL-10 triggers changes in macrophage phenotype that promote muscle growth and regeneration. J. Immunol. 2012, 189, 3669–3680. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Kohda, Y.; Chiao, H.; Wang, Y.; Hu, X.; Hewitt, S.M.; Miyaji, T.; McLeroy, P.; Nibhanupudy, B.; Li, S.; et al. Interleukin-10 inhibits ischemic and cisplatin-induced acute renal injury. Kidney Int. 2001, 60, 2118–2128. [Google Scholar] [CrossRef] [Green Version]
- King, A.; Balaji, S.; Le, L.D.; Crombleholme, T.M.; Keswani, S.G. Regenerative Wound Healing: The Role of Interleukin-10. Adv. Wound Care (New Rochelle) 2014, 3, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.J.; Zheng, W.D.; Chen, Y.X.; Huang, Y.H.; Chen, Z.X.; Zhang, S.J.; Shi, M.N.; Wang, X.Z. Antifibrotic effects of interleukin-10 on experimental hepatic fibrosis. Hepatogastroenterology 2007, 54, 2092–2098. [Google Scholar]
- Tran, G.T.; Hodgkinson, S.J.; Carter, N.M.; Verma, N.D.; Plain, K.M.; Boyd, R.; Robinson, C.M.; Nomura, M.; Killingsworth, M.; Hall, B.M. IL-5 promotes induction of antigen-specific CD4+CD25+ T regulatory cells that suppress autoimmunity. Blood 2012, 119, 4441–4450. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.M.; Plain, K.M.; Tran, G.T.; Verma, N.D.; Robinson, C.M.; Nomura, M.; Boyd, R.; Hodgkinson, S.J. Cytokines affecting CD4(+)T regulatory cells in transplant tolerance. III. Interleukin-5 (IL-5) promotes survival of alloantigen-specific CD4(+) T regulatory cells. Transplant. Immunol. 2017, 43–44, 33–41. [Google Scholar] [CrossRef]
- Sim, G.C.; Martin-Orozco, N.; Jin, L.; Yang, Y.; Wu, S.; Washington, E.; Sanders, D.; Lacey, C.; Wang, Y.; Vence, L.; et al. IL-2 therapy promotes suppressive ICOS+ Treg expansion in melanoma patients. J. Clin. Investig. 2014, 124, 99–110. [Google Scholar] [CrossRef] [Green Version]
- Roura, S.; Monguio-Tortajada, M.; Munizaga-Larroude, M.; Clos-Sansalvador, M.; Franquesa, M.; Rosell, A.; Borras, F.E. Potential of Extracellular Vesicle-Associated TSG-6 from Adipose Mesenchymal Stromal Cells in Traumatic Brain Injury. Int. J. Mol. Sci. 2020, 21, 6761. [Google Scholar] [CrossRef]
- Watanabe, R.; Sato, Y.; Ozawa, N.; Takahashi, Y.; Koba, S.; Watanabe, T. Emerging Roles of Tumor Necrosis Factor-Stimulated Gene-6 in the Pathophysiology and Treatment of Atherosclerosis. Int. J. Mol. Sci. 2018, 19, 465. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Li, G.; Zhang, Y.; Liu, N.; Zhang, P.; Pan, C.; Nie, H.; Li, Q.; Tang, Z. Upregulated TSG-6 Expression in ADSCs Inhibits the BV2 Microglia-Mediated Inflammatory Response. Biomed. Res. Int. 2018, 2018, 7239181. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lim, D.G.; Wee, Y.M.; Kim, J.H.; Yun, C.O.; Choi, M.Y.; Park, Y.H.; Kim, S.C.; Han, D.J. Viral IL-10 gene transfer prolongs rat islet allograft survival. Cell Transplant. 2008, 17, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Sakai, K.; Nozaki, Y.; Murao, Y.; Yano, T.; Ri, J.; Niki, K.; Kinoshita, K.; Funauchi, M.; Matsumura, I. Protective effect and mechanism of IL-10 on renal ischemia–reperfusion injury. Lab. Investig. 2019. [Google Scholar] [CrossRef]
- Kassan, M.; Galan, M.; Partyka, M.; Trebak, M.; Matrougui, K. Interleukin-10 released by CD4(+)CD25(+) natural regulatory T cells improves microvascular endothelial function through inhibition of NADPH oxidase activity in hypertensive mice. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2534–2542. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Gao, X.; Shen, G.; Wang, W.; Li, J.; Zhao, J.; Wei, Y.Q.; Edwards, C.K. Interleukin-10 deficiency impairs regulatory T cell-derived neuropilin-1 functions and promotes Th1 and Th17 immunity. Sci. Rep. 2016, 6, 24249. [Google Scholar] [CrossRef] [Green Version]
- Kingsley, C.I.; Karim, M.; Bushell, A.R.; Wood, K.J. CD25+CD4+ regulatory T cells prevent graft rejection: CTLA-4- and IL-10-dependent immunoregulation of alloresponses. J. Immunol. 2002, 168, 1080–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, S.; Gagliani, N.; Esplugues, E.; O’Connor, W., Jr.; Huber, F.J.; Chaudhry, A.; Kamanaka, M.; Kobayashi, Y.; Booth, C.J.; Rudensky, A.Y.; et al. Th17 cells express interleukin-10 receptor and are controlled by Foxp3(-) and Foxp3+ regulatory CD4+ T cells in an interleukin-10-dependent manner. Immunity 2011, 34, 554–565. [Google Scholar] [CrossRef] [Green Version]
- D’Alessio, F.R.; Zhong, Q.; Jenkins, J.; Moldobaeva, A.; Wagner, E.M. Lung Angiogenesis Requires CD4(+)Forkhead Homeobox Protein-3(+) Regulatory T Cells. Am. J. Respir. Cell Mol. Biol. 2015, 52, 603–610. [Google Scholar] [CrossRef] [Green Version]
- Mock, J.R.; Garibaldi, B.T.; Aggarwal, N.R.; Jenkins, J.; Limjunyawong, N.; Singer, B.D.; Chau, E.; Rabold, R.; Files, D.C.; Sidhaye, V.; et al. Foxp3+ regulatory T cells promote lung epithelial proliferation. Mucosal Immunol. 2014, 7, 1440–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | Recipient | Treatment Plan | Monitoring of Transplants |
|---|---|---|---|
| C57BL/6 | C57BL/6 | Vehicle-Treated Syngeneic Control | 9, 10, 12, 14, 28 |
| BALB/c | C57BL/6 | Vehicle-Treated Allogeneic Control | 9, 10, 12, 14, 28 |
| BALB/c | C57BL/6 | IL-10 Depletion | 9, 10, 12, 14, 28 |
| BALB/c | C57BL/6 | IL-10 Reconstitution | 9, 10, 12, 14, 28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.A.; Ashoor, G.A.; Shamma, T.; Alanazi, F.; Altuhami, A.; Kazmi, S.; Ahmed, H.A.; Mohammed Assiri, A.; Clemens Broering, D. IL-10 Mediated Immunomodulation Limits Subepithelial Fibrosis and Repairs Airway Epithelium in Rejecting Airway Allografts. Cells 2021, 10, 1248. https://doi.org/10.3390/cells10051248
Khan MA, Ashoor GA, Shamma T, Alanazi F, Altuhami A, Kazmi S, Ahmed HA, Mohammed Assiri A, Clemens Broering D. IL-10 Mediated Immunomodulation Limits Subepithelial Fibrosis and Repairs Airway Epithelium in Rejecting Airway Allografts. Cells. 2021; 10(5):1248. https://doi.org/10.3390/cells10051248
Chicago/Turabian StyleKhan, Mohammad Afzal, Ghazi Abdulmalik Ashoor, Talal Shamma, Fatimah Alanazi, Abdullah Altuhami, Shadab Kazmi, Hala Abdalrahman Ahmed, Abdullah Mohammed Assiri, and Dieter Clemens Broering. 2021. "IL-10 Mediated Immunomodulation Limits Subepithelial Fibrosis and Repairs Airway Epithelium in Rejecting Airway Allografts" Cells 10, no. 5: 1248. https://doi.org/10.3390/cells10051248
APA StyleKhan, M. A., Ashoor, G. A., Shamma, T., Alanazi, F., Altuhami, A., Kazmi, S., Ahmed, H. A., Mohammed Assiri, A., & Clemens Broering, D. (2021). IL-10 Mediated Immunomodulation Limits Subepithelial Fibrosis and Repairs Airway Epithelium in Rejecting Airway Allografts. Cells, 10(5), 1248. https://doi.org/10.3390/cells10051248