
The Autophagy Machinery in Human-Parasitic Protists; Diverse Functions for Universally Conserved Proteins



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
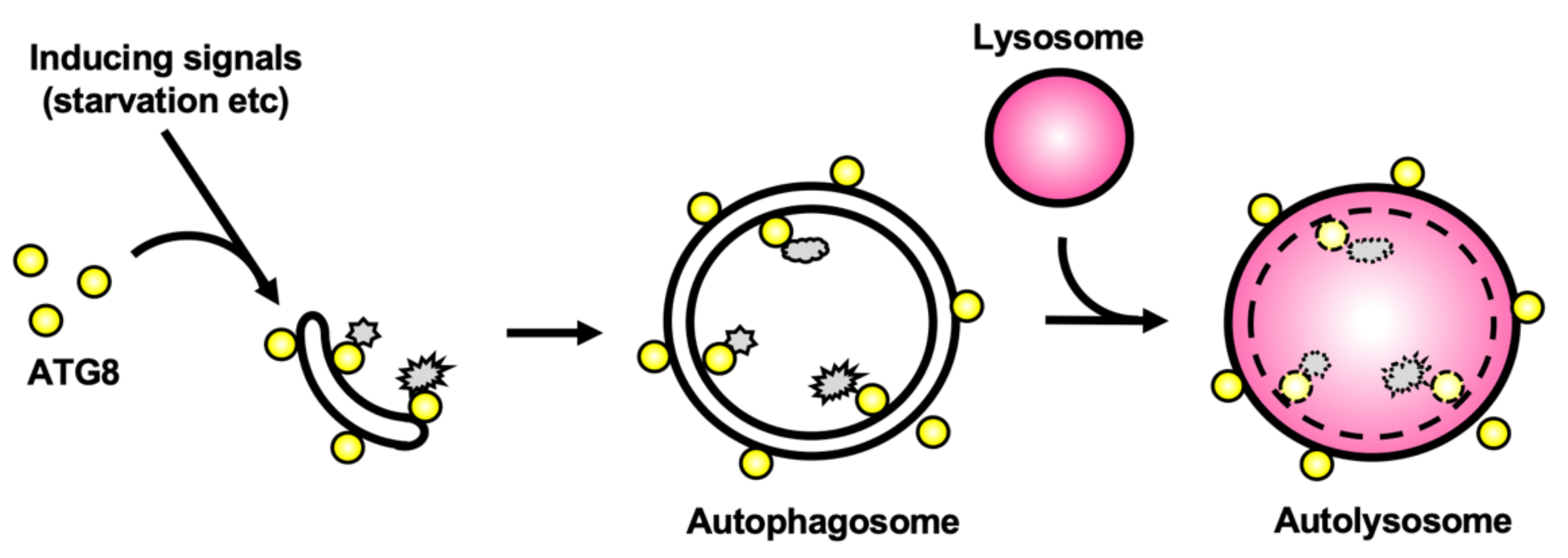
:1. Introduction
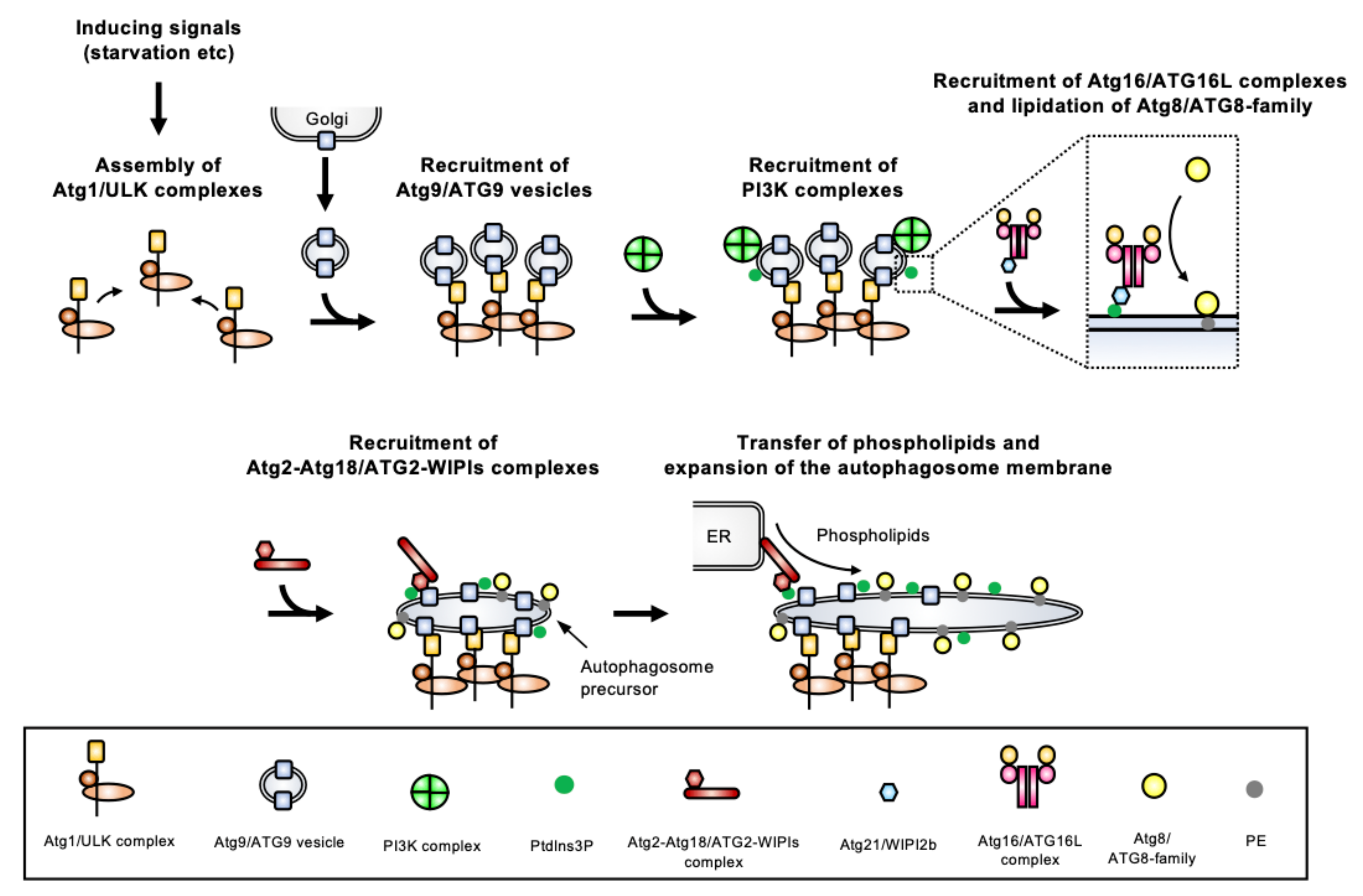
2. The ‘Core’ ATGs for Autophagosome Formation
2.1. Atg1/ULK Complex
2.2. Atg9/ATG9 Vesicles
2.3. PI3K Complex
2.4. Atg2-Atg18/ATG2-WIPI Complex
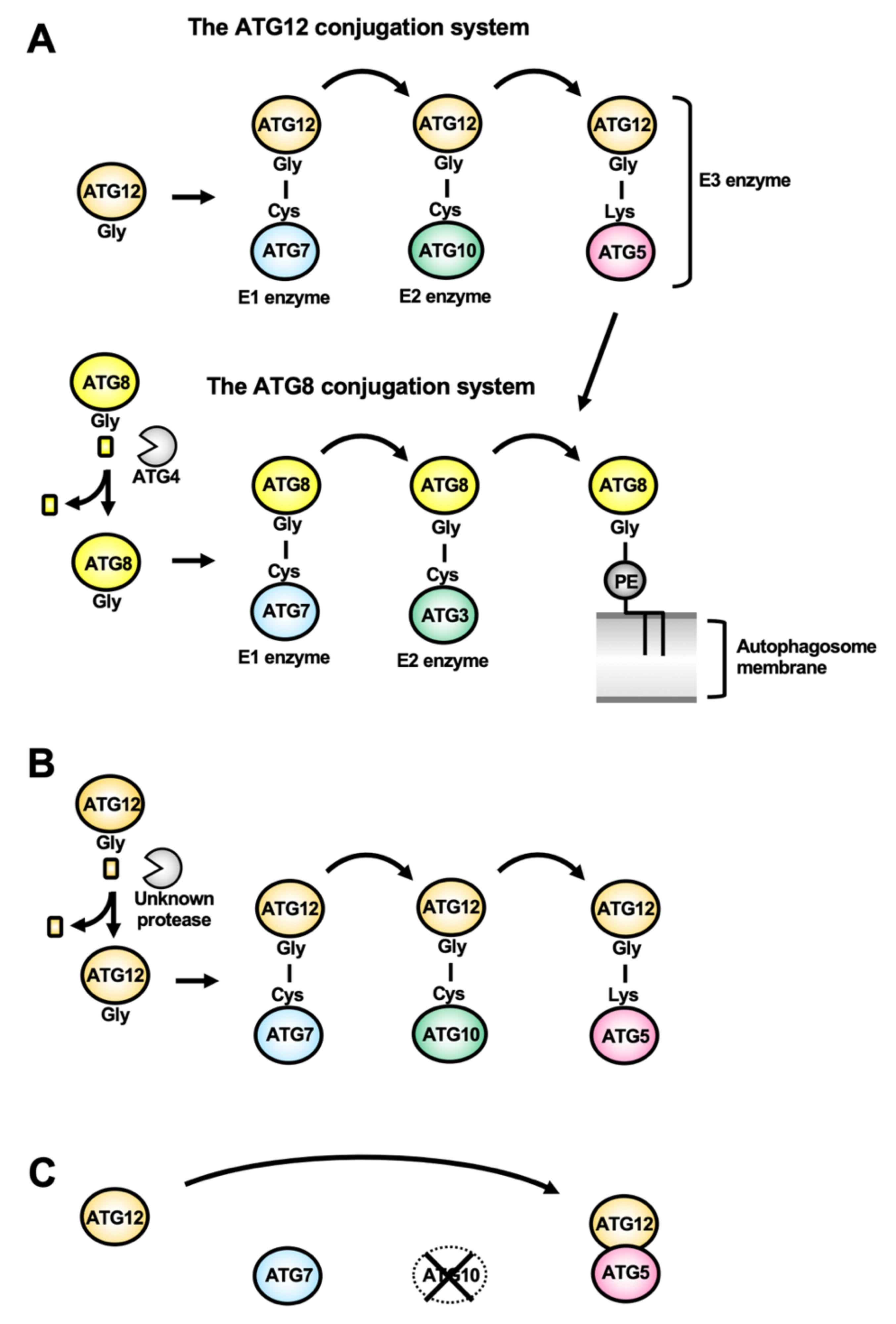
2.5. Atg12/ATG12 Conjugation System (Atg16/ATG16L Complex)
2.6. Atg8/ATG8-Family Conjugation System
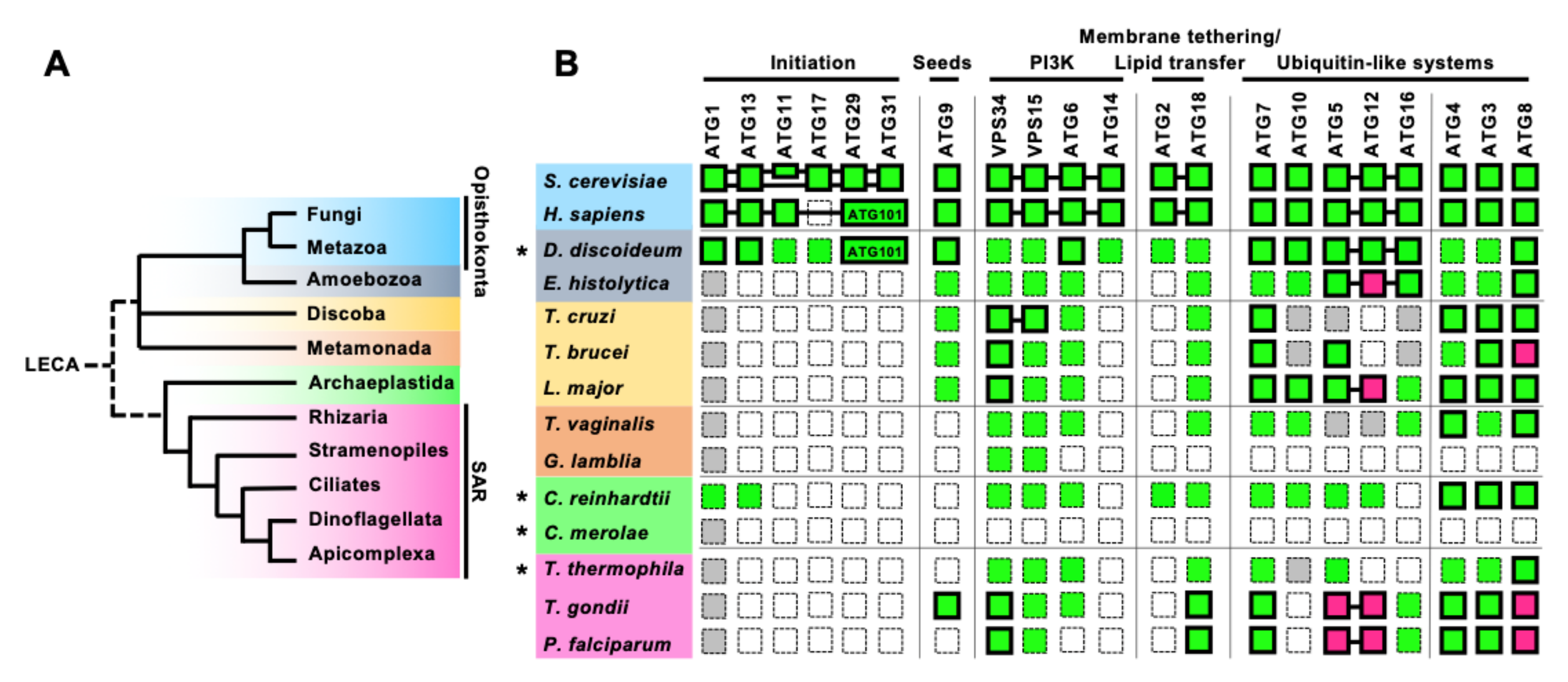
3. Overview of Conservation of the Core ATGs in Protists
3.1. Poor Conservation of the Early Components of the Autophagy Machinery in Protists
3.2. Conservation of the PtdIns3P Signaling Complex
3.3. The ATG8 Conjugation System Is Conserved in Almost All Protists
4. Autophagy in Parasitic Protists
4.1. Trypanosomatidae
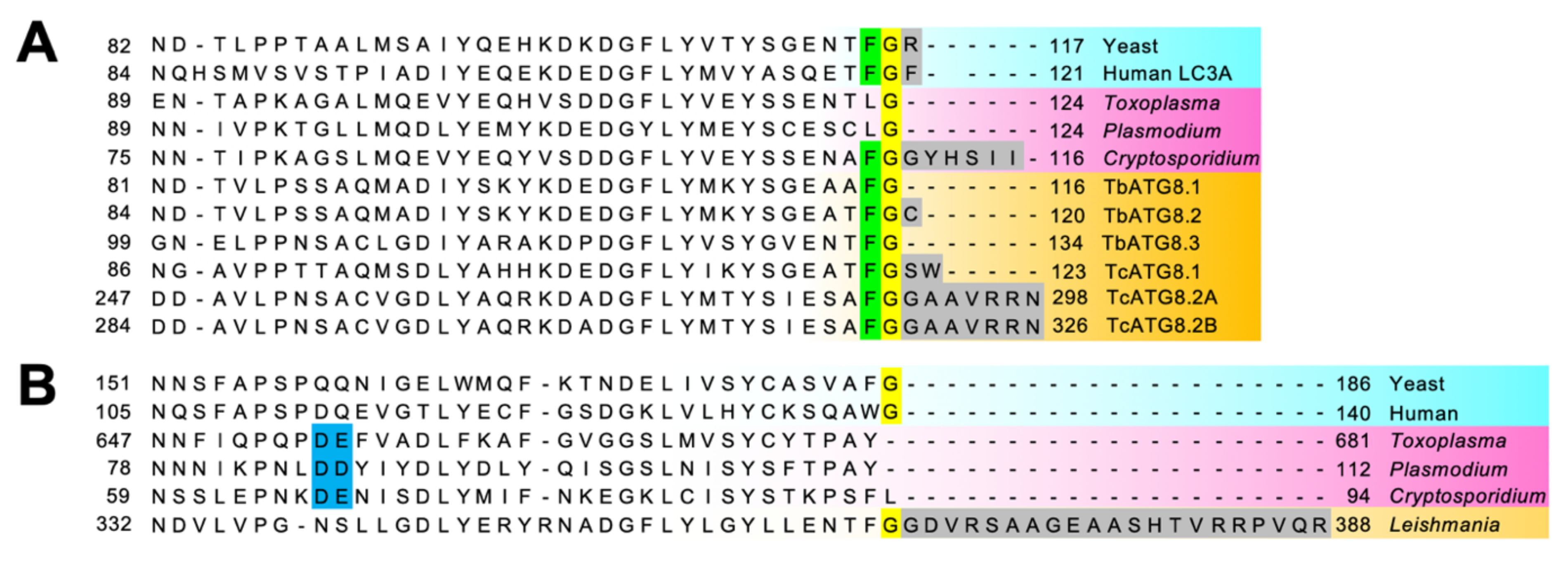
4.1.1. The ATG8 Conjugation System: ATG8s Are Multicopy and Some of Them Are Atypical
4.1.2. The ATG12 Conjugation System: Originality and Uncertainty
4.1.3. Autophagosome-Like Vesicles in Trypanosomatidae
4.1.4. Physiological Significance of the ATGs in the Life Cycle of Trypanosomatidae
4.1.5. Autophagy in Organelle Turnover
4.1.6. Alternative Ways of Regulating Autophagy in Trypanosomatidae?
4.2. Apicomplexa
4.2.1. ATG8 Has Atypical Features in Apicomplexa
4.2.2. An Atypical ATG12-ATG5 “Complex”: The First Evidence for Evolutionary Loss of Covalent Conjugation in Ubiquitin-Like ATG12
4.2.3. Autophagosome-Like Vesicles in Apicomplexa
4.2.4. ATGs Are Essential for Apicoplast Biogenesis
4.3. Entamoeba
4.3.1. ATG12 and ATG8 Conjugation Systems in Entamoeba spp
4.3.2. Function of the ATGs in Entamoeba spp
4.4. Metamonada (Giardia, Trichomonas)
4.4.1. The ATG Machinery in Metamonada
4.4.2. Function of the ATGs in Metamonada
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Duve, C.; Wattiaux, R. Functions of Lysosomes. Annu. Rev. Physiol. 1966, 28, 435–492. [Google Scholar] [CrossRef]
- Yang, Z.; Klionsky, D.J. Eaten alive: A history of macroautophagy. Nat. Cell Biol. 2010, 12, 814–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohsumi, Y. Historical landmarks of autophagy research. Cell Res. 2014, 24, 9–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukada, M.; Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 1993, 333, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Dikic, I.; Elazar, Z. Mechanism and medical implications of mammalian autophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 349–364. [Google Scholar] [CrossRef]
- Levine, B.; Kroemer, G. Biological Functions of Autophagy Genes: A Disease Perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [Green Version]
- Pohl, C.; Dikic, I. Cellular quality control by the ubiquitin-proteasome system and autophagy. Science 2019, 366, 818–822. [Google Scholar] [CrossRef]
- Morishita, H.; Mizushima, N. Diverse Cellular Roles of Autophagy. Annu. Rev. Cell Dev. Biol. 2019, 35, 453–475. [Google Scholar] [CrossRef]
- Faktorová, D.; Nisbet, R.E.R.; Robledo, J.A.F.; Casacuberta, E.; Sudek, L.; Allen, A.E.; Ares, M.; Aresté, C.; Balestreri, C.; Barbrook, A.C.; et al. Genetic tool development in marine protists: Emerging model organisms for experimental cell biology. Nat. Methods 2020, 17, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Keeling, P.J.; Burki, F.; Wilcox, H.M.; Allam, B.; Allen, E.E.; Amaral-Zettler, L.A.; Armbrust, E.V.; Archibald, J.M.; Bharti, A.K.; Bell, C.J.; et al. The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): Illuminating the Functional Diversity of Eukaryotic Life in the Oceans through Transcriptome Sequencing. PLoS Biol. 2014, 12, e1001889. [Google Scholar] [CrossRef] [Green Version]
- Montagnes, D.; Roberts, E.; Lukeš, J.; Lowe, C. The rise of model protozoa. Trends Microbiol. 2012, 20, 184–191. [Google Scholar] [CrossRef]
- Plattner, H. Evolutionary Cell Biology of Proteins from Protists to Humans and Plants. J. Eukaryot. Microbiol. 2017, 65, 255–289. [Google Scholar] [CrossRef]
- Lander, N.; Chiurillo, M.A.; Docampo, R. Genome Editing by CRISPR/Cas9: A Game Change in the Genetic Manipulation of Protists. J. Eukaryot. Microbiol. 2016, 63, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Yoshimoto, K.; Ohsumi, Y. Unveiling the molecular mechanisms of plant autophagy—From autophagosomes to vacuoles in plants. Plant Cell Physiol. 2018, 59, 1337–1344. [Google Scholar] [CrossRef]
- Nakatogawa, H. Mechanisms governing autophagosome biogenesis. Nat. Rev. Mol. Cell Biol. 2020, 21, 439–458. [Google Scholar] [CrossRef]
- Noda, N.N.; Wang, Z.; Zhang, H. Liquid–liquid phase separation in autophagy. J. Cell Biol. 2020, 219, 219. [Google Scholar] [CrossRef]
- Dudley, L.J.; Makar, A.N.; Gammoh, N. Membrane targeting of core autophagy players during autophagosome biogenesis. FEBS J. 2020, 287, 4806–4821. [Google Scholar] [CrossRef]
- Nascimbeni, A.C.; Codogno, P.; Morel, E. Phosphatidylinositol-3-phosphate in the regulation of autophagy membrane dynamics. FEBS J. 2017, 284, 1267–1278. [Google Scholar] [CrossRef]
- Proikas-Cezanne, T.; Takacs, Z.; Dönnes, P.; Kohlbacher, O. WIPI proteins: Essential PtdIns3P effectors at the nascent autophagosome. J. Cell Sci. 2015, 128, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N. The ATG conjugation systems in autophagy. Curr. Opin. Cell Biol. 2020, 63, 1–10. [Google Scholar] [CrossRef]
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [Green Version]
- Burki, F.; Roger, A.J.; Brown, M.W.; Simpson, A.G. The New Tree of Eukaryotes. Trends Ecol. Evol. 2020, 35, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Calvo-Garrido, J.; Carilla-Latorre, S.; Kubohara, Y.; Santos-Rodrigo, N.; Mesquita, A.; Soldati, T.; Golstein, P.; Escalante, R. Autophagy in Dictyostelium: Genes and pathways, cell death and infection. Autophagy 2010, 6, 686–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesquita, A.; Cardenal-Muñoz, E.; Dominguez, E.; Muñoz-Braceras, S.; Nuñez-Corcuera, B.; Phillips, B.A.; Tábara, L.C.; Xiong, Q.; Coria, R.; Eichinger, L.; et al. Autophagy in Dictyostelium: Mechanisms, regulation and disease in a simple biomedical model. Autophagy 2017, 13, 24–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shemi, A.; Ben-Dor, S.; Vardi, A. Elucidating the composition and conservation of the autophagy pathway in photosynthetic eukaryotes. Autophagy 2015, 11, 701–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslan, E.; Küçükoğlu, N.; Arslanyolu, M. A comparative in-silico analysis of autophagy proteins in ciliates. PeerJ 2017, 5, e2878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delorme-Axford, E.; Klionsky, D.J. A missing piece of the puzzle: Atg11 functions as a scaffold to activate Atg1 for selective autophagy. Autophagy 2015, 11, 2139–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, H.; Fujioka, Y.; Suzuki, S.W.; Noshiro, D.; Suzuki, H.; Kondo-Kakuta, C.; Kimura, Y.; Hirano, H.; Ando, T.; Noda, N.N.; et al. The Intrinsically Disordered Protein Atg13 Mediates Supramolecular Assembly of Autophagy Initiation Complexes. Dev. Cell 2016, 38, 86–99. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Kakuta, S.; Watanabe, T.M.; Kitamura, A.; Sekito, T.; Kondo-Kakuta, C.; Ichikawa, R.; Kinjo, M.; Ohsumi, Y. Atg9 vesicles are an important membrane source during early steps of autophagosome formation. J. Cell Biol. 2012, 198, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Sawa-Makarska, J.; Baumann, V.; Coudevylle, N.; Von Bülow, S.; Nogellova, V.; Abert, C.; Schuschnig, M.; Graef, M.; Hummer, G.; Martens, S. Reconstitution of autophagosome nucleation defines Atg9 vesicles as seeds for membrane formation. Science 2020, 369, eaaz7714. [Google Scholar] [CrossRef] [PubMed]
- Matoba, K.; Kotani, T.; Tsutsumi, A.; Tsuji, T.; Mori, T.; Noshiro, D.; Sugita, Y.; Nomura, N.; Iwata, S.; Ohsumi, Y.; et al. Atg9 is a lipid scramblase that mediates autophagosomal membrane expansion. Nat. Struct. Mol. Biol. 2020, 27, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Yamamoto, H.; Kinch, L.N.; Garza, C.M.; Takahashi, S.; Otomo, C.; Grishin, N.V.; Forli, S.; Mizushima, N.; Otomo, T. Structure, lipid scrambling activity and role in autophagosome formation of ATG9A. Nat. Struct. Mol. Biol. 2020, 27, 1194–1201. [Google Scholar] [CrossRef] [PubMed]
- Osawa, T.; Kotani, T.; Kawaoka, T.; Hirata, E.; Suzuki, K.; Nakatogawa, H.; Ohsumi, Y.; Noda, N.N. Atg2 mediates direct lipid transfer between membranes for autophagosome formation. Nat. Struct. Mol. Biol. 2019, 26, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Otomo, C.; Otomo, T. The autophagic membrane tether ATG2A transfers lipids between membranes. eLife 2019, 8, 8. [Google Scholar] [CrossRef]
- Valverde, D.P.; Yu, S.; Boggavarapu, V.; Kumar, N.; Lees, J.A.; Walz, T.; Reinisch, K.M.; Melia, T.J. ATG2 transports lipids to promote autophagosome biogenesis. J. Cell Biol. 2019, 218, 1787–1798. [Google Scholar] [CrossRef] [Green Version]
- Wesch, N.; Kirkin, V.; Rogov, V.V. Atg8-Family Proteins—Structural Features and Molecular Interactions in Autophagy and Beyond. Cells 2020, 9, 2008. [Google Scholar] [CrossRef]
- Maruyama, T.; Noda, N. Autophagy-regulating protease Atg4: Structure, function, regulation and inhibition. J. Antibiot. 2018, 71, 72–78. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in Mammalian Autophagy Research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef] [Green Version]
- Yoshii, S.R.; Mizushima, N. Monitoring and Measuring Autophagy. Int. J. Mol. Sci. 2017, 18, 1865. [Google Scholar] [CrossRef]
- Kuma, A.; Komatsu, M.; Mizushima, N. Autophagy-monitoring and autophagy-deficient mice. Autophagy 2017, 13, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Murphy, L.O. Autophagy Assays for Biological Discovery and Therapeutic Development. Trends Biochem. Sci. 2020, 45, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Kaizuka, T.; Morishita, H.; Hama, Y.; Tsukamoto, S.; Matsui, T.; Toyota, Y.; Kodama, A.; Ishihara, T.; Mizushima, T.; Mizushima, N. An Autophagic Flux Probe that Releases an Internal Control. Mol. Cell 2016, 64, 835–849. [Google Scholar] [CrossRef] [Green Version]
- Rigden, D.J.; Michels, P.; Ginger, M.L. Autophagy in protists: Examples of secondary loss, lineage-specific innovations, and the conundrum of remodeling a single mitochondrion. Autophagy 2009, 5, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Brennand, A.; Gualdrón-López, M.; Coppens, I.; Rigden, D.J.; Ginger, M.L.; Michels, P.A. Autophagy in parasitic protists: Unique features and drug targets. Mol. Biochem. Parasitol. 2011, 177, 83–99. [Google Scholar] [CrossRef]
- Duszenko, M.; Ginger, M.L.; Brennand, A.; Gualdron-López, M.; Colombo, M.I.; Coombs, G.H.; Coppens, I.; Jayabalasingham, B.; Langsley, G.; De Castro, S.L.; et al. Autophagy in protists. Autophagy 2011, 7, 127–158. [Google Scholar] [CrossRef] [Green Version]
- Besteiro, S. Autophagy in apicomplexan parasites. Curr. Opin. Microbiol. 2017, 40, 14–20. [Google Scholar] [CrossRef]
- De Laté, P.L.; Pineda, M.; Harnett, M.; Harnett, W.; Besteiro, S.; Langsley, G. Apicomplexan autophagy and modulation of autophagy in parasite-infected host cells. Biomed. J. 2017, 40, 23–30. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, M.A.; Leger, M.M.; Wideman, J.G.; Ruiz-Trillo, I. Concepts of the last eukaryotic common ancestor. Nat. Ecol. Evol. 2019, 3, 338–344. [Google Scholar] [CrossRef]
- Misumi, O.; Matsuzaki, M.; Nozaki, H.; Miyagishima, S.-Y.; Mori, T.; Nishida, K.; Yagisawa, F.; Yoshida, Y.; Kuroiwa, H.; Kuroiwa, T. Cyanidioschyzon merolae Genome. A Tool for Facilitating Comparable Studies on Organelle Biogenesis in Photosynthetic Eukaryotes. Plant Physiol. 2005, 137, 567–585. [Google Scholar] [CrossRef] [Green Version]
- Földvári-Nagy, L.; Ari, E.; Csermely, P.; Korcsmáros, T.; Vellai, T. Starvation-response may not involve Atg1-dependent autophagy induction in non-unikont parasites. Sci. Rep. 2014, 4, srep05829. [Google Scholar] [CrossRef] [Green Version]
- Kihara, A.; Noda, T.; Ishihara, N.; Ohsumi, Y. Two Distinct Vps34 Phosphatidylinositol 3–Kinase Complexes Function in Autophagy and Carboxypeptidase Y Sorting inSaccharomyces cerevisiae. J. Cell Biol. 2001, 152, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Itakura, E.; Kishi, C.; Inoue, K.; Mizushima, N. Beclin 1 Forms Two Distinct Phosphatidylinositol 3-Kinase Complexes with Mammalian Atg14 and UVRAG. Mol. Biol. Cell 2008, 19, 5360–5372. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, V.E.; Kosec, G.; Sant’Anna, C.; Turk, V.; Cazzulo, J.J.; Turk, B. Autophagy Is Involved in Nutritional Stress Response and Differentiation in Trypanosoma cruzi. J. Biol. Chem. 2008, 283, 3454–3464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.A.; Woods, K.L.; Juliano, L.; Mottram, J.C.; Coombs, G.H. Characterisation of unusual families of ATG8-like proteins and ATG12 in the protozoan parasite Leishmania major. Autophagy 2009, 5, 159–172. [Google Scholar] [CrossRef] [Green Version]
- Besteiro, S.; Williams, R.A.; Coombs, G.H.; Mottram, J.C. Protein turnover and differentiation in Leishmania. Int. J. Parasitol. 2007, 37, 1063–1075. [Google Scholar] [CrossRef] [Green Version]
- Williams, R.A.M.; Smith, T.K.; Cull, B.; Mottram, J.C.; Coombs, G.H. ATG5 Is Essential for ATG8-Dependent Autophagy and Mitochondrial Homeostasis in Leishmania major. PLoS Pathog. 2012, 8, e1002695. [Google Scholar] [CrossRef] [Green Version]
- Salassa, B.N.; Romano, P.S. Autophagy: A necessary process during the Trypanosoma cruzi life-cycle. Virulence 2019, 10, 460–469. [Google Scholar] [CrossRef] [Green Version]
- Proto, W.R.; Jones, N.G.; Coombs, G.H.; Mottram, J.C. Tracking autophagy during proliferation and differentiation of Trypanosoma brucei. Microb. Cell 2014, 1, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shen, Q.; Wang, C.; Sun, Y.; Yuan, A.Y.; He, C.Y. A role of autophagy in Trypanosoma brucei cell death. Cell. Microbiol. 2012, 14, 1242–1256. [Google Scholar] [CrossRef]
- Santa-Rita, R.M.; Lira, R.; Barbosa, H.S.; Urbina, J.A.; De Castro, S.L. Anti-proliferative synergy of lysophospholipid analogues and ketoconazole against Trypanosoma cruzi (Kinetoplastida: Trypanosomatidae): Cellular and ultrastructural analysis. J. Antimicrob. Chemother. 2005, 55, 780–784. [Google Scholar] [CrossRef] [Green Version]
- Braga, M.V.; De Souza, W. Effects of protein kinase and phosphatidylinositol-3 kinase inhibitors on growth and ultrastructure of Trypanosoma cruzi. FEMS Microbiol. Lett. 2006, 256, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Vanrell, M.C.; Losinno, A.D.; Cueto, J.A.; Balcazar, D.; Fraccaroli, L.V.; Carrillo, C.; Romano, P.S. The regulation of autophagy differentially affects Trypanosoma cruzi metacyclogenesis. PLoS Neglected Trop. Dis. 2017, 11, e0006049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledezma, E.; Jorquera, A.; Bendezú, H.; Vivas, J.; Pérez, G. Antiproliferative and leishmanicidal effect of ajoene on various Leishmania species: Ultrastructural study. Parasitol. Res. 2002, 88, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Santa-Rita, R.M.; Henriques-Pons, A.; Barbosa, H.S.; De Castro, S.L. Effect of the lysophospholipid analogues edelfosine, ilmofosine and miltefosine against Leishmania amazonensis. J. Antimicrob. Chemother. 2004, 54, 704–710. [Google Scholar] [CrossRef]
- Williams, R.A.; Tetley, L.; Mottram, J.C.; Coombs, G.H. Cysteine peptidases CPA and CPB are vital for autophagy and differentiation in Leishmania mexicana. Mol. Microbiol. 2006, 61, 655–674. [Google Scholar] [CrossRef]
- Besteiro, S.; Williams, R.A.; Morrison, L.S.; Coombs, G.H.; Mottram, J.C. Endosome Sorting and Autophagy Are Essential for Differentiation and Virulence of Leishmania major. J. Biol. Chem. 2006, 281, 11384–11396. [Google Scholar] [CrossRef] [Green Version]
- Denny, P.W.; Goulding, D.; Ferguson, M.A.J.; Smith, D.F. Sphingolipid-free Leishmania are defective in membrane trafficking, differentiation and infectivity. Mol. Microbiol. 2004, 52, 313–327. [Google Scholar] [CrossRef]
- Williams, R.A.M.; Mottram, J.C.; Coombs, G.H. Distinct Roles in Autophagy and Importance in Infectivity of the Two ATG4 Cysteine Peptidases of Leishmania major. J. Biol. Chem. 2013, 288, 3678–3690. [Google Scholar] [CrossRef] [Green Version]
- Haanstra, J.R.; González-Marcano, E.B.; Gualdrón-López, M.; Michels, P.A. Biogenesis, maintenance and dynamics of glycosomes in trypanosomatid parasites. Biochim. Biophys. Acta Bioenerg. 2016, 1863, 1038–1048. [Google Scholar] [CrossRef]
- Bauer, S.; Morris, M.T. Glycosome biogenesis in trypanosomes and the de novo dilemma. PLoS Neglected Trop. Dis. 2017, 11, e0005333. [Google Scholar] [CrossRef]
- Oku, M.; Sakai, Y. Pexophagy in yeasts. Biochim. Biophys. Acta Bioenerg. 2016, 1863, 992–998. [Google Scholar] [CrossRef] [PubMed]
- Germain, K.; Kim, P.K. Pexophagy: A Model for Selective Autophagy. Int. J. Mol. Sci. 2020, 21, 578. [Google Scholar] [CrossRef] [Green Version]
- Cull, B.; Godinho, J.L.P.; Rodrigues, J.C.F.; Frank, B.; Schurigt, U.; Williams, R.A.; Coombs, G.H.; Mottram, J.C. Glycosome turnover in Leishmania major is mediated by autophagy. Autophagy 2014, 10, 2143–2157. [Google Scholar] [CrossRef] [Green Version]
- Herman, M.; Pérez-Morga, D.; Schtickzelle, N.; Michels, P.A. Turnover of glycosomes during life-cycle differentiation of Trypanosoma brucei. Autophagy 2008, 4, 294–308. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, S.; Honda, S.; Yamaguchi, H.; Shimizu, S. Molecular mechanisms and physiological roles of Atg5/Atg7-independent alternative autophagy. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Oku, M.; Sakai, Y. Three Distinct Types of Microautophagy Based on Membrane Dynamics and Molecular Machineries. BioEssays 2018, 40, e1800008. [Google Scholar] [CrossRef]
- Nakamura, S.; Izumi, M. Chlorophagy is ATG gene-dependent microautophagy process. Plant Signal. Behav. 2019, 14, 1554469. [Google Scholar] [CrossRef] [Green Version]
- Cunha-E-Silva, N.; Sant’Anna, C.; Pereira, M.G.; Porto-Carreiro, I.; Jeovanio, A.L.; De Souza, W. Reservosomes: Multipurpose organelles? Parasitol. Res. 2006, 99, 325–327. [Google Scholar] [CrossRef]
- Siqueira-Neto, J.L.; Debnath, A.; McCall, L.-I.; Bernatchez, J.A.; Ndao, M.; Reed, S.L.; Rosenthal, P.J. Cysteine proteases in protozoan parasites. PLoS Negl. Trop. Dis. 2018, 12, e0006512. [Google Scholar] [CrossRef]
- De Cazzulo, B.M.F.; Martínez, J.; North, M.J.; Coombs, G.H.; Cazzulo, J.-J. Effects of proteinase inhibitors on the growth and differentiation of Trypanosoma cruzi. FEMS Microbiol. Lett. 1994, 124, 81–86. [Google Scholar] [CrossRef]
- Losinno, A.D.; Martínez, S.J.; Labriola, C.A.; Carrillo, C.; Romano, P.S. Induction of autophagy increases the proteolytic activity of reservosomes during Trypanosoma cruzi metacyclogenesis. Autophagy 2021, 17, 439–456. [Google Scholar] [CrossRef]
- Vietri, M.; Radulovic, M.; Stenmark, H. The many functions of ESCRTs. Nat. Rev. Mol. Cell Biol. 2020, 21, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-J.; He, C.Y. Acidocalcisome is required for autophagy in Trypanosoma brucei. Autophagy 2014, 10, 1978–1988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Docampo, R.; Huang, G. Acidocalcisomes of eukaryotes. Curr. Opin. Cell Biol. 2016, 41, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Schoijet, A.C.; Sternlieb, T.; Alonso, G.D. The Phosphatidylinositol 3-kinase Class III Complex Containing TcVps15 and TcVps34 Participates in Autophagy in Trypanosoma cruzi. J. Eukaryot. Microbiol. 2017, 64, 308–321. [Google Scholar] [CrossRef]
- Kitamura, K.; Kishi-Itakura, C.; Tsuboi, T.; Sato, S.; Kita, K.; Ohta, N.; Mizushima, N. Autophagy-Related Atg8 Localizes to the Apicoplast of the Human Malaria Parasite Plasmodium falciparum. PLoS ONE 2012, 7, e42977. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Yamamoto, H.; Sakamoto, H.; Oku, M.; Mutungi, J.K.; Sahani, M.H.; Kurikawa, Y.; Kita, K.; Noda, N.N.; Sakai, Y.; et al. Evolution from covalent conjugation to non-covalent interaction in the ubiquitin-like ATG12 system. Nat. Struct. Mol. Biol. 2019, 26, 289–296. [Google Scholar] [CrossRef]
- Kong-Hap, M.A.; Mouammine, A.; Daher, W.; Berry, L.; Lebrun, M.; Dubremetz, J.-F.; Besteiro, S. Regulation of ATG8 membrane association by ATG4 in the parasitic protist Toxoplasma gondii. Autophagy 2013, 9, 1334–1348. [Google Scholar] [CrossRef] [Green Version]
- Besteiro, S.; Brooks, C.F.; Striepen, B.; Dubremetz, J.-F. Autophagy Protein Atg3 is Essential for Maintaining Mitochondrial Integrity and for Normal Intracellular Development of Toxoplasma gondii Tachyzoites. PLoS Pathog. 2011, 7, e1002416. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, D.; Walton, J.L.; Roepe, P.D.; Sinai, A.P. Autophagy is a cell death mechanism in Toxoplasma gondii. Cell. Microbiol. 2011, 14, 589–607. [Google Scholar] [CrossRef] [Green Version]
- LaVine, M.D.; Arrizabalaga, G. Analysis of Monensin Sensitivity in Toxoplasma gondii Reveals Autophagy as a Mechanism for Drug Induced Death. PLoS ONE 2012, 7, e42107. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.M.; Berry, L.; Sullivan, W.J.; Besteiro, S. Autophagy participates in the unfolded protein response in Toxoplasma gondii. FEMS Microbiol. Lett. 2017, 364, 364. [Google Scholar] [CrossRef] [PubMed]
- Tomlins, A.M.; Ben-Rached, F.; Williams, R.A.; Proto, W.R.; Coppens, I.; Ruch, U.; Gilberger, T.W.; Coombs, G.H.; Mottram, J.C.; Müller, S.; et al. Plasmodium falciparumATG8 implicated in both autophagy and apicoplast formation. Autophagy 2013, 9, 1540–1552. [Google Scholar] [CrossRef] [Green Version]
- Eickel, N.; Kaiser, G.; Prado, M.; Burda, P.-C.; Roelli, M.; Stanway, R.R.; Heussler, V.T. Features of autophagic cell death in Plasmodium liver-stage parasites. Autophagy 2013, 9, 568–580. [Google Scholar] [CrossRef] [Green Version]
- Jayabalasingham, B.; Bano, N.; Coppens, I. Metamorphosis of the malaria parasite in the liver is associated with organelle clearance. Cell Res. 2010, 20, 1043–1059. [Google Scholar] [CrossRef] [Green Version]
- Jayabalasingham, B.; Voss, C.; Ehrenman, K.; Romano, J.D.; Smith, M.E.; Fidock, D.A.; Bosch, J.; Coppens, I. Characterization of the ATG8-conjugation system in 2 Plasmodium species with special focus on the liver stage. Autophagy 2013, 10, 269–284. [Google Scholar] [CrossRef]
- Voss, C.; Ehrenman, K.; Mlambo, G.; Mishra, S.; Kumar, K.A.; Sacci, J.B.; Sinnis, P.; Coppens, I. Overexpression of Plasmodium berghei ATG8 by Liver Forms Leads to Cumulative Defects in Organelle Dynamics and to Generation of Noninfectious Merozoites. mBio 2016, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Ponpuak, M.; Mandell, M.A.; Kimura, T.; Chauhan, S.; Cleyrat, C.; Deretic, V. Secretory autophagy. Curr. Opin. Cell Biol. 2015, 35, 106–116. [Google Scholar] [CrossRef] [Green Version]
- Miranda, K.; Pace, D.A.; Cintron, R.; Rodrigues, J.; Fang, J.; Smith, A.; Rohloff, P.; Coelho, E.; De Haas, F.; De Souza, W.; et al. Characterization of a novel organelle in Toxoplasma gondii with similar composition and function to the plant vacuole. Mol. Microbiol. 2010, 76, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Parussini, F.; Coppens, I.; Shah, P.P.; Diamond, S.L.; Carruthers, V.B. Cathepsin L occupies a vacuolar compartment and is a protein maturase within the endo/exocytic system of Toxoplasma gondii. Mol. Microbiol. 2010, 76, 1340–1357. [Google Scholar] [CrossRef] [Green Version]
- Dou, Z.; Coppens, I.; Carruthers, V.B. Non-canonical Maturation of Two Papain-family Proteases in Toxoplasma gondii. J. Biol. Chem. 2013, 288, 3523–3534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.M.; El Hajj, H.; El Hajj, R.; Tawil, N.; Berry, L.; Lebrun, M.; Bordat, Y.; Besteiro, S. Toxoplasma gondii autophagy-related protein ATG9 is crucial for the survival of parasites in their host. Cell. Microbiol. 2017, 19, e12712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cristina, M.; Dou, Z.; Lunghi, M.; Kannan, G.; Huynh, M.-H.; McGovern, O.L.; Schultz, T.L.; Schultz, A.J.; Miller, A.J.; Hayes, B.M.; et al. Toxoplasma depends on lysosomal consumption of autophagosomes for persistent infection. Nat. Microbiol. 2017, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.; Kannan, G.; Coppens, I.; Wang, F.; Nguyen, H.M.; Cerutti, A.; Olafsson, E.B.; Rimple, P.A.; Schultz, T.L.; Soto, N.M.M.; et al. Toxoplasma TgATG9 is critical for autophagy and long-term persistence in tissue cysts. eLife 2021, 10, 59384. [Google Scholar] [CrossRef] [PubMed]
- Van Dooren, G.G.; Hapuarachchi, S.V. The Dark Side of the Chloroplast: Biogenesis, Metabolism and Membrane Biology of the Apicoplast. Adv. Bot. Res. 2017, 84, 145–185. [Google Scholar] [CrossRef]
- Mizushima, N.; Sahani, M.H. ATG8 localization in apicomplexan parasites. Autophagy 2014, 10, 1487–1494. [Google Scholar] [CrossRef] [Green Version]
- Tawk, L.; Dubremetz, J.-F.; Montcourrier, P.; Chicanne, G.; Merezegue, F.; Richard, V.; Payrastre, B.; Meissner, M.; Vial, H.J.; Roy, C.; et al. Phosphatidylinositol 3-Monophosphate Is Involved in Toxoplasma Apicoplast Biogenesis. PLoS Pathog. 2011, 7, e1001286. [Google Scholar] [CrossRef] [Green Version]
- Daher, W.; Morlon-Guyot, J.; Sheiner, L.; Lentini, G.; Berry, L.; Tawk, L.; Dubremetz, J.-F.; Wengelnik, K.; Striepen, B.; Lebrun, M. Lipid kinases are essential for apicoplast homeostasis in Toxoplasma gondii. Cell. Microbiol. 2014, 17, 559–578. [Google Scholar] [CrossRef] [Green Version]
- Bansal, P.; Tripathi, A.; Thakur, V.; Mohmmed, A.; Sharma, P. Autophagy-Related Protein ATG18 Regulates Apicoplast Biogenesis in Apicomplexan Parasites. mBio 2017, 8, e01468-17. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.M.; Liu, S.; Daher, W.; Tan, F.; Besteiro, S. Characterisation of two Toxoplasma PROPPINs homologous to Atg18/WIPI suggests they have evolved distinct specialised functions. PLoS ONE 2018, 13, e0195921. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Meister, T.R.; Walczak, M.; Pulkoski-Gross, M.J.; Hari, S.B.; Sauer, R.T.; Amberg-Johnson, K.; Yeh, E. A mutagenesis screen for essential plastid biogenesis genes in human malaria parasites. PLoS Biol. 2019, 17, e3000136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévêque, M.F.; Berry, L.; Cipriano, M.J.; Nguyen, H.-M.; Striepen, B.; Besteiro, S. Autophagy-Related Protein ATG8 Has a Noncanonical Function for Apicoplast Inheritance in Toxoplasma gondii. mBio 2015, 6, e01446-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walczak, M.; Ganesan, S.M.; Niles, J.C.; Yeh, E. ATG8 Is Essential Specifically for an Autophagy-Independent Function in Apicoplast Biogenesis in Blood-Stage Malaria Parasites. mBio 2018, 9, e02021-17. [Google Scholar] [CrossRef] [Green Version]
- Kaneda, Y.; Torii, M.; Tanaka, T.; Aikawa, M. In vitroeffects of berberine sulphate on the growth and structure of Entamoeba histolytica, Giardia lamblia and Trichomonas vaginalis. Ann. Trop. Med. Parasitol. 1991, 85, 417–425. [Google Scholar] [CrossRef]
- Picazarri, K.; Nakada-Tsukui, K.; Tsuboi, K.; Miyamoto, E.; Watanabe, N.; Kawakami, E.; Nozaki, T. Atg8 is involved in endosomal and phagosomal acidification in the parasitic protist Entamoeba histolytica. Cell. Microbiol. 2015, 17, 1510–1522. [Google Scholar] [CrossRef] [Green Version]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Ali, V.; Nozaki, T. A Retromerlike Complex Is a Novel Rab7 Effector That Is Involved in the Transport of the Virulence Factor Cysteine Protease in the Enteric Protozoan Parasite Entamoeba histolytica. Mol. Biol. Cell 2005, 16, 5294–5303. [Google Scholar] [CrossRef] [Green Version]
- Picazarri, K.; Nakada-Tsukui, K.; Nozaki, T. Autophagy during Proliferation and Encystation in the Protozoan Parasite Entamoeba invadens. Infect. Immun. 2007, 76, 278–288. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Moon, E.-K.; Hong, Y.; Chung, D.-I.; Kong, H.-H. Autophagy protein 12 plays an essential role in Acanthamoeba encystation. Exp. Parasitol. 2015, 159, 46–52. [Google Scholar] [CrossRef]
- Song, S.-M.; Han, B.-I.; Moon, E.-K.; Lee, Y.-R.; Yu, H.S.; Jha, B.K.; Danne, D.-B.S.; Kong, H.-H.; Chung, D.-I.; Hong, Y. Autophagy protein 16-mediated autophagy is required for the encystation of Acanthamoeba castellanii. Mol. Biochem. Parasitol. 2012, 183, 158–165. [Google Scholar] [CrossRef]
- Huang, K.-Y.; Chen, R.-M.; Lin, H.-C.; Cheng, W.-H.; Lin, W.-N.; Huang, P.-J.; Chiu, C.-H.; Tang, P. Potential role of autophagy in proteolysis in Trichomonas vaginalis. J. Microbiol. Immunol. Infect. 2019, 52, 336–344. [Google Scholar] [CrossRef]
- Hernández-García, M.S.; Miranda-Ozuna, J.F.T.; Salazar-Villatoro, L.; Vázquez-Calzada, C.; Ávila-González, L.; González-Robles, A.; Ortega-López, J.; Arroyo, R. Biogenesis of Autophagosome in Trichomonas vaginalis during Macroautophagy Induced by Rapamycin-treatment and Iron or Glucose Starvation Conditions. J. Eukaryot. Microbiol. 2019, 66, 654–669. [Google Scholar] [CrossRef] [PubMed]
- Cernikova, L.; Faso, C.; Hehl, A.B. Five facts about Giardia lamblia. PLoS Pathog. 2018, 14, e1007250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.-Y.; Chen, Y.-Y.M.; Fang, Y.-K.; Cheng, W.-H.; Cheng, C.-C.; Chen, Y.-C.; Wu, T.E.; Ku, F.-M.; Lin, R.; Tang, P. Adaptive responses to glucose restriction enhance cell survival, antioxidant capability, and autophagy of the protozoan parasite Trichomonas vaginalis. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 53–64. [Google Scholar] [CrossRef]
- Ishii, S.; Matsuura, A.; Itakura, E. Identification of a factor controlling lysosomal homeostasis using a novel lysosomal trafficking probe. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zheng, C.; Liu, F.; Yang, C.; Zheng, P.; Lu, X.; Tian, J.; Chung, T.; Otegui, M.S.; Xiao, S.; et al. Genetic Analyses of the Arabidopsis ATG1 Kinase Complex Reveal Both Kinase-Dependent and Independent Autophagic Routes during Fixed-Carbon Starvation. Plant Cell 2019, 31, 2973–2995. [Google Scholar] [CrossRef]
- Fujioka, Y.; Alam, J.M.; Noshiro, D.; Mouri, K.; Ando, T.; Okada, Y.; May, A.I.; Knorr, R.L.; Suzuki, K.; Ohsumi, Y.; et al. Phase separation organizes the site of autophagosome formation. Nat. Cell Biol. 2020, 578, 301–305. [Google Scholar] [CrossRef]
- Lystad, A.H.; Simonsen, A. Mechanisms and Pathophysiological Roles of the ATG8 Conjugation Machinery. Cells 2019, 8, 973. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakamoto, H.; Nakada-Tsukui, K.; Besteiro, S. The Autophagy Machinery in Human-Parasitic Protists; Diverse Functions for Universally Conserved Proteins. Cells 2021, 10, 1258. https://doi.org/10.3390/cells10051258
Sakamoto H, Nakada-Tsukui K, Besteiro S. The Autophagy Machinery in Human-Parasitic Protists; Diverse Functions for Universally Conserved Proteins. Cells. 2021; 10(5):1258. https://doi.org/10.3390/cells10051258
Chicago/Turabian StyleSakamoto, Hirokazu, Kumiko Nakada-Tsukui, and Sébastien Besteiro. 2021. "The Autophagy Machinery in Human-Parasitic Protists; Diverse Functions for Universally Conserved Proteins" Cells 10, no. 5: 1258. https://doi.org/10.3390/cells10051258
APA StyleSakamoto, H., Nakada-Tsukui, K., & Besteiro, S. (2021). The Autophagy Machinery in Human-Parasitic Protists; Diverse Functions for Universally Conserved Proteins. Cells, 10(5), 1258. https://doi.org/10.3390/cells10051258