
Establishing a 3D In Vitro Hepatic Model Mimicking Physiologically Relevant to In Vivo State

 ,
,  ,
,  and
and 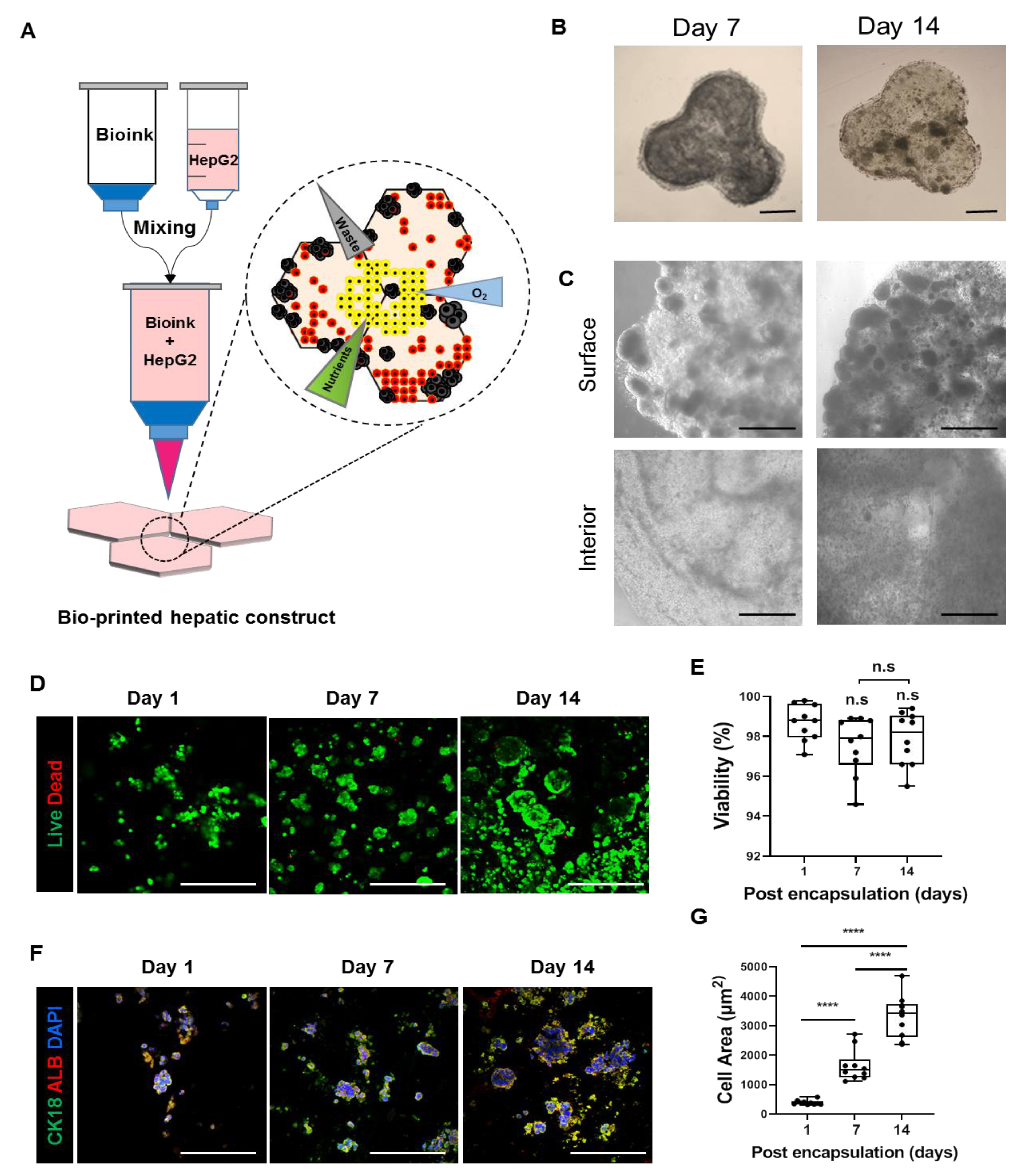{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. HepG2 Cell Line Culture
2.2. Bioprinting of 3D Hepatic Constructs
2.3. Live/Dead Cell Staining
2.4. Quantitative RT-PCR
2.5. Histological Characterization (H&E Staining)
2.6. Immunocytochemistry
2.7. Functional Analysis
2.8. Western Blot Analysis
2.9. Drug (Acetaminophen, N-Acetyl-L-cysteine) Treatment
2.10. Statistical Analysis
3. Results
3.1. Fabrication of 3D Bioprinted Human Liver Tissue
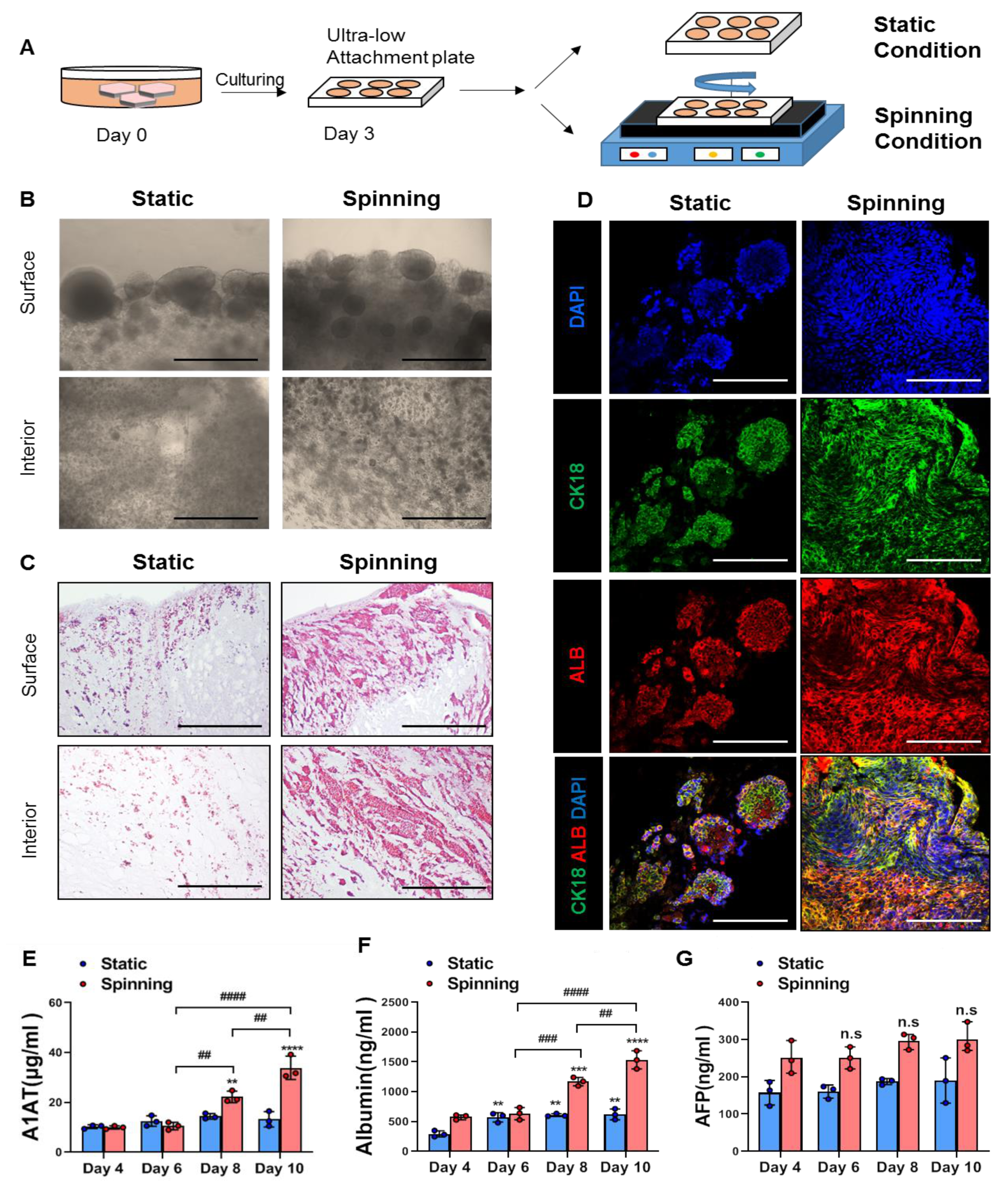
3.2. Spinning Culture Conditions Enable a Long-Term Culture Period of Bioprinted Hepatic Constructs with Consistent Hepatic Expression and Functionality of Encapsulated Cells
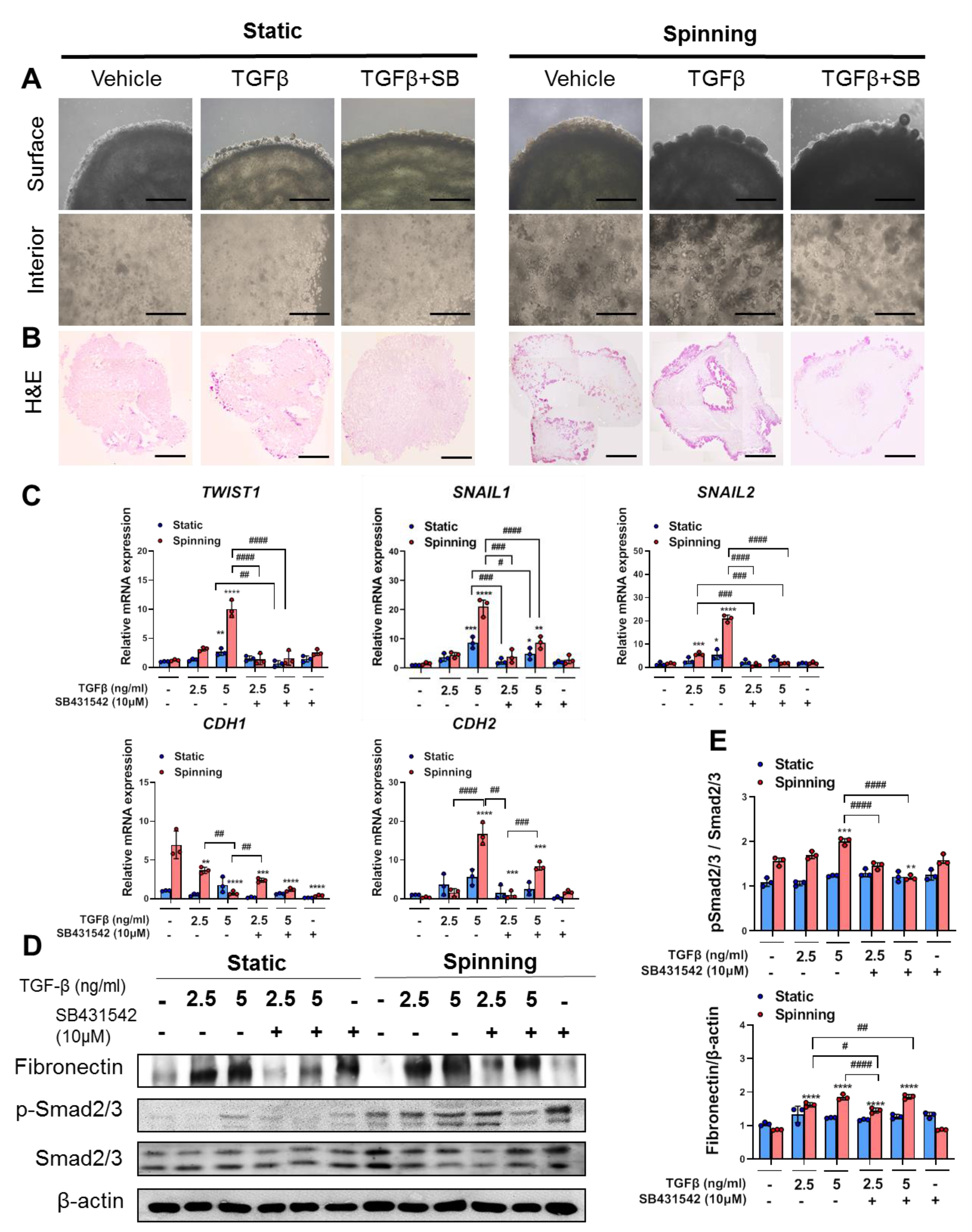
3.3. 3D Bioprinted Hepatic Constructs Show Efficient Changes in Cellular Characteristics under Spinning Conditions
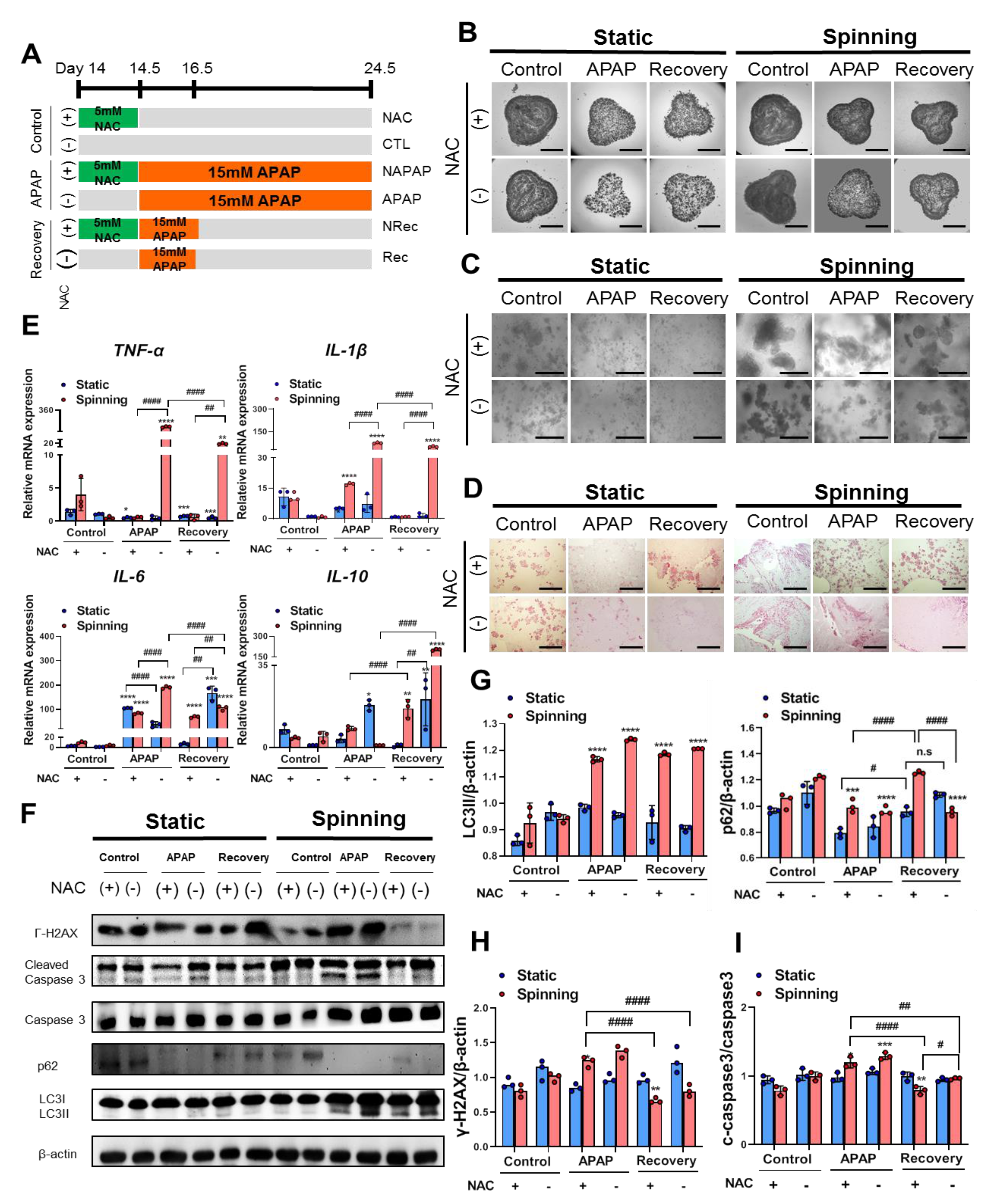
3.4. The Dynamic Microenvironment Generated by an Orbital Shaker Enables Artificial Liver Construction as a Liver Toxicity Test Model
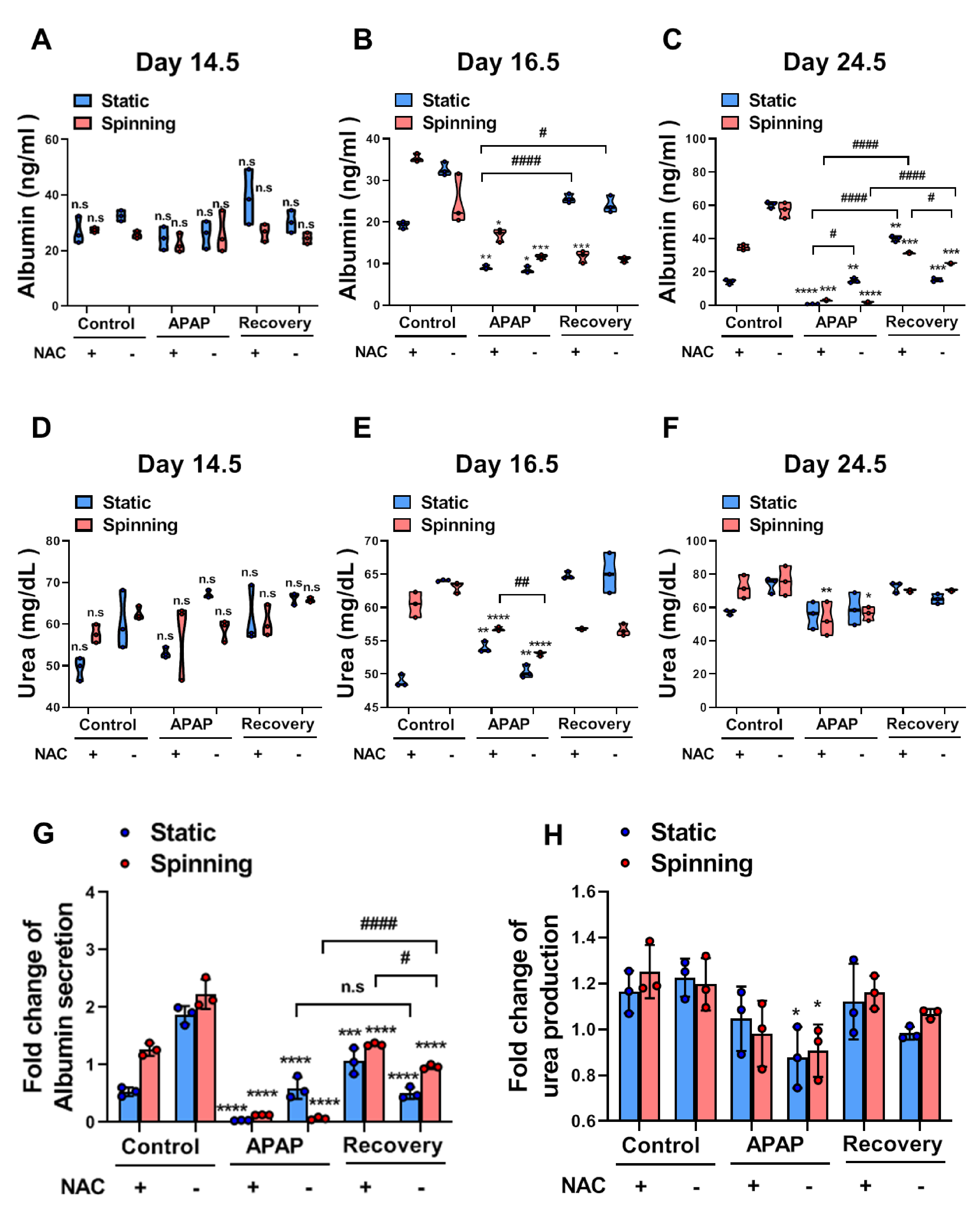
3.5. Functional Evaluation of Bioprinted Hepatic Constructs Revealed That Hepatotoxicity Induced by APAP and Prevented by NAC Was Enhanced under Spinning Conditions
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cui, X.; Li, J.; Hartanto, Y.; Durham, M.; Tang, J.; Zhang, H.; Hooper, G.; Lim, K.; Woodfield, T. Advances in Extrusion 3D Bioprinting: A Focus on Multicomponent Hydrogel-Based Bioinks. Adv. Healthc. Mater. 2020, 9, 1901648. [Google Scholar] [CrossRef] [PubMed]
- Tasoglu, S.; Demirci, U. Bioprinting for stem cell research. Trends Biotechnol. 2013, 31, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Matai, I.; Kaur, G.; Seyedsalehi, A.; McClinton, A.; Laurencin, C.T. Progress in 3D bioprinting technology for tissue/organ regenerative engineering. Biomaterials 2020, 226, 119536. [Google Scholar] [CrossRef] [PubMed]
- Mandrycky, C.; Wang, Z.; Kim, K.; Kim, D.-H. 3D bioprinting for engineering complex tissues. Biotechnol. Adv. 2016, 34, 422–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.N.; Underhill, G.H.; Zaret, K.S.; Fox, I.J. Cell and tissue engineering for liver disease. Sci. Transl. Med. 2014, 6, 245sr2. [Google Scholar] [CrossRef] [Green Version]
- Ozbolat, I.T.; Hospodiuk, M. Current advances and future perspectives in extrusion-based bioprinting. Biomaterials 2016, 76, 321–343. [Google Scholar] [CrossRef] [Green Version]
- Davoodi, E.; Sarikhani, E.; Montazerian, H.; Ahadian, S.; Costantini, M.; Swieszkowski, W.; Willerth, S.; Walus, K.; Mofidfar, M.; Toyserkani, E.; et al. Extrusion and Microfluidic-Based Bioprinting to Fabricate Biomimetic Tissues and Organs. Adv. Mater. Technol. 2020, 5, 1901044. [Google Scholar] [CrossRef]
- Ozbolat, I.T. Scaffold-Based or Scaffold-Free Bioprinting: Competing or Complementing Approaches? J. Nanotechnol. Eng. Med. 2015, 6, 024701. [Google Scholar] [CrossRef]
- Lee, S.W.; Jung, D.J.; Jeong, G.S. Gaining New Biological and Therapeutic Applications into the Liver with 3D In Vitro Liver Models. Tissue Eng. Regen. Med. 2020, 17, 731–745. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Gibeley, S.B.; Xu, C.; Xiao, Y.; Celik, O.; Ginsberg, H.N.; Leong, K.W. Engineering Liver Microtissues for Disease Modeling and Regenerative Medicine. Adv. Funct. Mater. 2020, 30, 1909553. [Google Scholar] [CrossRef] [PubMed]
- Lauschke, V.M.; Hendriks, D.F.G.; Bell, C.C.; Andersson, T.B.; Ingelman-Sundberg, M. Novel 3D Culture Systems for Studies of Human Liver Function and Assessments of the Hepatotoxicity of Drugs and Drug Candidates. Chem. Res. Toxicol. 2016, 29, 1936–1955. [Google Scholar] [CrossRef]
- Hosseini, V.; Maroufi, N.F.; Saghati, S.; Asadi, N.; Darabi, M.; Ahmad, S.N.S.; Hosseinkhani, H.; Rahbarghazi, R. Current progress in hepatic tissue regeneration by tissue engineering. J. Transl. Med. 2019, 17, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Rashidi, H.; Alhaque, S.; Szkolnicka, D.; Flint, O.; Hay, D.C. Fluid shear stress modulation of hepatocyte-like cell function. Arch. Toxicol. 2016, 90, 1757–1761. [Google Scholar] [CrossRef] [Green Version]
- Bhise, N.S.; Manoharan, V.; Massa, S.; Tamayol, A.; Ghaderi, M.; Miscuglio, M.; Lang, Q.; Zhang, Y.S.; Shin, S.R.; Calzone, G.; et al. A liver-on-a-chip platform with bioprinted hepatic spheroids. Biofabrication 2016, 8, 14101. [Google Scholar] [CrossRef]
- No, D.Y.; Lee, K.-H.; Lee, J.; Lee, S.-H. 3D liver models on a microplatform: Well-defined culture, engineering of liver tissue and liver-on-a-chip. Lab Chip 2015, 15, 3822–3837. [Google Scholar] [CrossRef]
- Cho, S.; Yoon, J.-Y. Organ-on-a-chip for assessing environmental toxicants. Curr. Opin. Biotechnol. 2017, 45, 34–42. [Google Scholar] [CrossRef]
- Ahmed, H.M.M.; Salerno, S.; Morelli, S.; Giorno, L.; De Bartolo, L. 3D liver membrane system by co-culturing human hepatocytes, sinusoidal endothelial and stellate cells. Biofabrication 2017, 9, 25022. [Google Scholar] [CrossRef]
- Lee, H.; Chae, S.; Kim, J.Y.; Han, W.; Kim, J.; Choi, Y.; Cho, D.W. Cell-printed 3D liver-on-a-chip possessing a liver microen-vironment and biliary system. Biofabrication 2019, 11, 025001. [Google Scholar] [CrossRef]
- Chang, R.; Emami, K.; Wu, H.; Sun, W. Biofabrication of a three-dimensional liver micro-organ as an in vitro drug metabolism model. Biofabrication 2010, 2, 045004. [Google Scholar] [CrossRef]
- Jin, Y.; Kim, J.; Lee, J.S.; Min, S.; Kim, S.; Ahn, D.H.; Kim, Y.G.; Cho, S.W. Vascularized liver organoids generated using induced hepatic tissue and dynamic liver-specific microenvironment as a drug testing platform. Adv. Funct. Mater. 2018, 28, 1801954. [Google Scholar] [CrossRef]
- Nguyen, D.G.; Funk, J.; Robbins, J.B.; Crogan-Grundy, C.; Presnell, S.C.; Singer, T.; Roth, A.B. Bioprinted 3D Primary Liver Tissues Allow Assessment of Organ-Level Response to Clinical Drug Induced Toxicity In Vitro. PLoS ONE 2016, 11, e0158674. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.-C.; Lee, J.Y.; Kim, J.; Yoo, J.M.; Kang, I.; Kim, J.-J.; Shin, N.; Kim, D.J.; Choi, S.W.; Kim, D.; et al. Graphene quantum dots as anti-inflammatory therapy for colitis. Sci. Adv. 2020, 6, eaaz2630. [Google Scholar] [CrossRef] [PubMed]
- Sung, E.A.; Yu, K.R.; Shin, J.H.; Seo, Y.; Kim, H.S.; Koog, M.G.; Kang, I.; Kim, J.J.; Lee, B.C.; Shin, T.H.; et al. Generation of patient specific human neural stem cells from Niemann-Pick disease type C patient-derived fibroblasts. Oncotarget 2017, 8, 85428. [Google Scholar] [CrossRef] [PubMed]
- Cameron, K.; Tan, R.; Schmidt-Heck, W.; Campos, G.; Lyall, M.J.; Wang, Y.; Lucendo-Villarin, B.; Szkolnicka, D.; Bates, N.; Kimber, S.J.; et al. Recombinant laminins drive the differentiation and self-organization of hESC-derived hepatocytes. Stem Cell Rep. 2015, 5, 1250–1262. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.K.; Schmid, R.; Arkudas, A.; Kengelbach-Weigand, A.; Bosserhoff, A.K. Tumor Cells Develop Defined Cellular Phenotypes after 3D-Bioprinting in Different Bioinks. Cells 2019, 8, 1295. [Google Scholar] [CrossRef] [Green Version]
- Talari, A.C.S.; Raza, A.; Rehman, S.; Rehman, I.U. Analyzing normal proliferating, hypoxic and necrotic regions of T-47D human breast cancer spheroids using Raman spectroscopy. Appl. Spectrosc. Rev. 2017, 52, 909–924. [Google Scholar] [CrossRef]
- Pinzani, M. Epithelial-mesenchymal transition in chronic liver disease: Fibrogenesis or escape from death? J. Hepatol. 2011, 55, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.S.; Diehl, A.M. Epithelial-to-mesenchymal transitions in the liver. Hepatology 2009, 50, 2007–2013. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Mao, S.; Yao, R.; He, J.; Zhou, Z.; Feng, L.; Zhang, K.; Cheng, S.; Sun, W. TGF-β induced epithelial–mesenchymal transition in an advanced cervical tumor model by 3D printing. Biofabrication 2018, 10, 044102. [Google Scholar] [CrossRef]
- Yang, R.; Miki, K.; He, X.; Killeen, M.E.; Fink, M.P. Prolonged treatment with N-acetylcystine delays liver recovery from ac-etaminophen hepatotoxicity. Crit. Care 2009, 13, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Palabiyik, S.S.; Karakus, E.; Halici, Z.; Cadirci, E.; Bayir, Y.; Ayaz, G.; Cinar, I. The protective effects of carvacrol and thymol against paracetamol–induced toxicity on human hepatocellular carcinoma cell lines (HepG2). Hum. Exp. Toxicol. 2016, 35, 1252–1263. [Google Scholar] [CrossRef]
- Liang, Y.-L.; Zhang, Z.-H.; Liu, X.-J.; Liu, X.-Q.; Tao, L.; Zhang, Y.-F.; Wang, H.; Zhang, C.; Chen, X.; Xu, D.-X. Melatonin Protects against Apoptosis-Inducing Factor (AIF)-Dependent Cell Death during Acetaminophen-Induced Acute Liver Failure. PLoS ONE 2012, 7, e51911. [Google Scholar] [CrossRef] [Green Version]
- Schuster, S.; Cabrera, D.; Arrese, M.; Feldstein, A.E. Triggering and resolution of inflammation in NASH. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 349–364. [Google Scholar] [CrossRef]
- Borude, P.; Bhushan, B.; Apte, U. DNA damage response regulates initiation of liver regeneration following aceta-minophen overdose. Gene Expr. 2018, 18, 115. [Google Scholar] [CrossRef]
- Ma, L.; Wu, Y.; Li, Y.; Aazmi, A.; Zhou, H.; Zhang, B.; Yang, H. Current Advances on 3D-Bioprinted Liver Tissue Models. Adv. Healthc. Mater. 2020, 9, e2001517. [Google Scholar] [CrossRef]
- Morgan, F.L.C.; Moroni, L.; Baker, M.B. Dynamic Bioinks to Advance Bioprinting. Adv. Healthc. Mater. 2020, 9, 1901798. [Google Scholar] [CrossRef]
- Carlsson, R.; Engvall, E.; Freeman, A.; Ruoslahti, E. Laminin and fibronectin in cell adhesion: Enhanced adhesion of cells from regenerating liver to laminin. Proc. Natl. Acad. Sci. USA 1981, 78, 2403–2406. [Google Scholar] [CrossRef] [Green Version]
- Kanninen, L.K.; Harjumäki, R.; Peltoniemi, P.; Bogacheva, M.; Salmi, T.; Porola, P.; Niklander, J.; Smutný, T.; Urtti, A.; Yliperttula, M.L.; et al. Laminin-511 and laminin-521-based matrices for efficient hepatic specification of human pluripotent stem cells. Biomaterials 2016, 103, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-Dimensional Cell Culture Systems and Their Applications in Drug Discovery and Cell-Based Biosensors. ASSAY Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Datta, P.; Dey, M.; Ataie, Z.; Unutmaz, D.; Ozbolat, I.T. 3D bioprinting for reconstituting the cancer microenvironment. npj Precis. Oncol. 2020, 4, 1–13. [Google Scholar] [CrossRef]
- Luckert, C.; Schulz, C.; Lehmann, N.; Thomas, M.; Hofmann, U.; Hammad, S.; Hengstler, J.G.; Braeuning, A.; Lampen, A.; Hessel, S. Comparative analysis of 3D culture methods on human HepG2 cells. Arch. Toxicol. 2017, 91, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Manov, I.; Hirsh, M.; Iancu, T.C. N-Acetylcysteine does not Protect HepG2 Cells against Acetaminophen-Induced Apoptosis. Basic Clin. Pharmacol. Toxicol. 2004, 94, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Duval, K.; Grover, H.; Han, L.-H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Jung, D.J.; Byeon, J.H.; Jeong, G.S. Flow enhances phenotypic and maturation of adult rat liver organoids. Biofabrication 2020, 12, 45035. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Wu, D.; Lo, J.; Wang, Y.; Wu, J.; Lu, S.; Xu, H.; Zhao, X.; He, Y.; Li, J.; et al. A bioartificial liver support system integrated with a DLM/GelMA-based bioengineered whole liver for prevention of hepatic encephalopathy via enhanced ammonia reduction. Biomater. Sci. 2020, 8, 2814–2824. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.J.; Lee, K.G.; Choi, D. Clinical implications of advances in liver regeneration. Clin. Mol. Hepatol. 2015, 21, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Romero, N.; Sainz-Arnal, P.; Pla-Palacín, I.; Dachary, P.R.; Almeida, H.; Pastor, C.; Soto, D.R.; Rodriguez, M.C.; Arbizu, E.O.; Martinez, L.B.; et al. The role of extracellular matrix on liver stem cell fate: A dynamic relationship in health and disease. Differentiation 2019, 106, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roskams, T.A.; Libbrecht, L.; Desmet, V.J. Progenitor Cells in Diseased Human Liver. Semin. Liver Dis. 2003, 23, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Vig, P.; Russo, F.P.; Edwards, R.J.; Tadrous, P.J.; Wright, N.A.; Thomas, H.C.; Alison, M.R.; Forbes, S.J. The sources of parenchymal regeneration after chronic hepatocellular liver injury in mice. Hepatology 2006, 43, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Unutmaz, D.; Ozbolat, I.T. Bioprinting towards Physiologically Relevant Tissue Models for Pharmaceutics. Trends Biotechnol. 2016, 34, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, M.; Bansal, R.; Rouwkema, J. Bioengineered 3d models to recapitulate tissue Fibros. Trends Biotechnol. 2020, 38, 623–636. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.K.; Sarsenova, M.; Kim, D.-H.; Kim, M.S.; Lee, J.Y.; Sung, E.-A.; Kook, M.G.; Kim, N.G.; Choi, S.W.; Ogay, V.; et al. Establishing a 3D In Vitro Hepatic Model Mimicking Physiologically Relevant to In Vivo State. Cells 2021, 10, 1268. https://doi.org/10.3390/cells10051268
Kang HK, Sarsenova M, Kim D-H, Kim MS, Lee JY, Sung E-A, Kook MG, Kim NG, Choi SW, Ogay V, et al. Establishing a 3D In Vitro Hepatic Model Mimicking Physiologically Relevant to In Vivo State. Cells. 2021; 10(5):1268. https://doi.org/10.3390/cells10051268
Chicago/Turabian StyleKang, Hyun Kyoung, Madina Sarsenova, Da-Hyun Kim, Min Soo Kim, Jin Young Lee, Eun-Ah Sung, Myung Geun Kook, Nam Gyo Kim, Soon Won Choi, Vyacheslav Ogay, and et al. 2021. "Establishing a 3D In Vitro Hepatic Model Mimicking Physiologically Relevant to In Vivo State" Cells 10, no. 5: 1268. https://doi.org/10.3390/cells10051268
APA StyleKang, H. K., Sarsenova, M., Kim, D. -H., Kim, M. S., Lee, J. Y., Sung, E. -A., Kook, M. G., Kim, N. G., Choi, S. W., Ogay, V., & Kang, K. -S. (2021). Establishing a 3D In Vitro Hepatic Model Mimicking Physiologically Relevant to In Vivo State. Cells, 10(5), 1268. https://doi.org/10.3390/cells10051268