
Hymenoptera Venom Immunotherapy: Immune Mechanisms of Induced Protection and Tolerance



Abstract
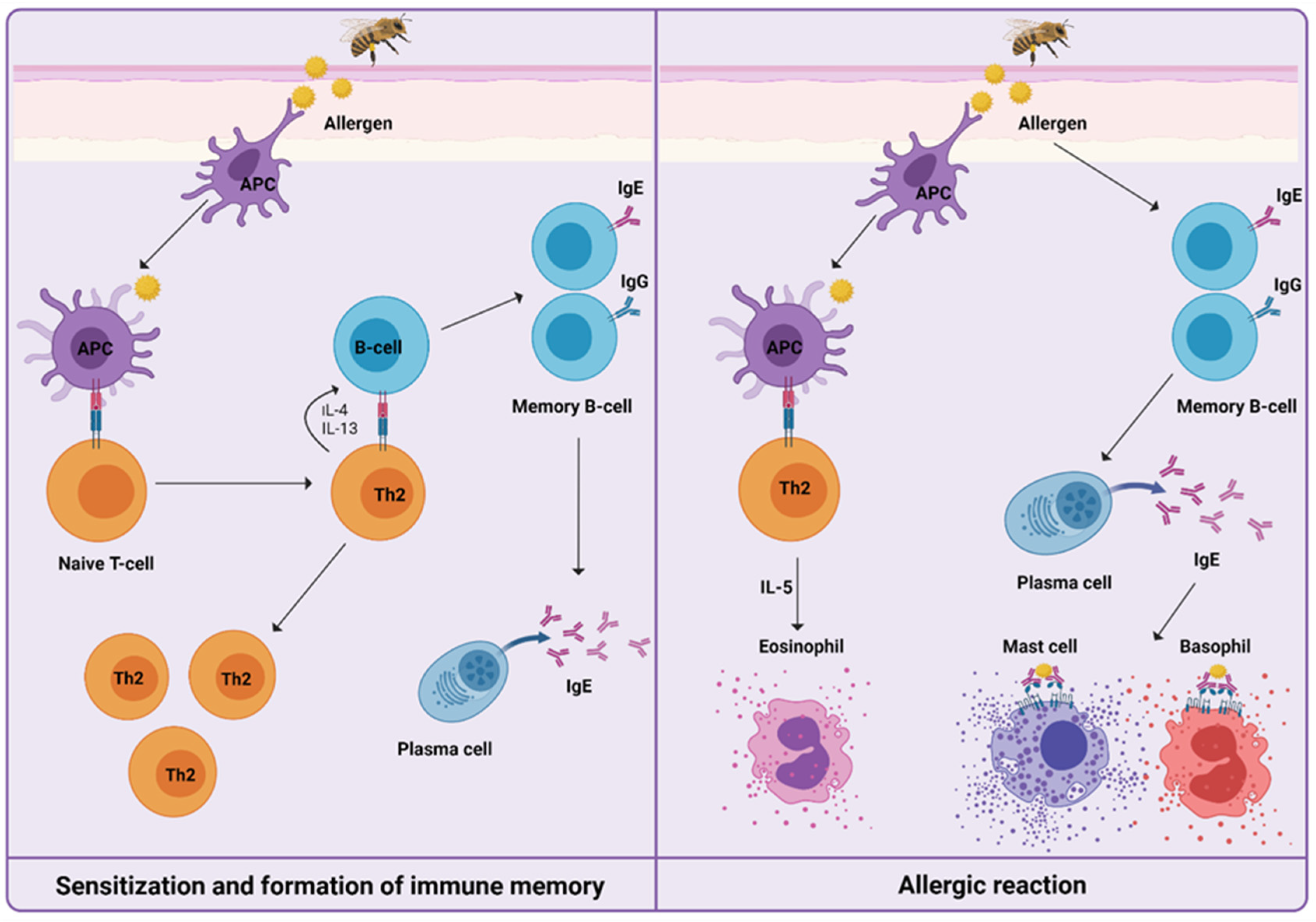
:1. Introduction
2. Venom Immunotherapy
3. Immune Mechanisms during Venom Immunotherapy
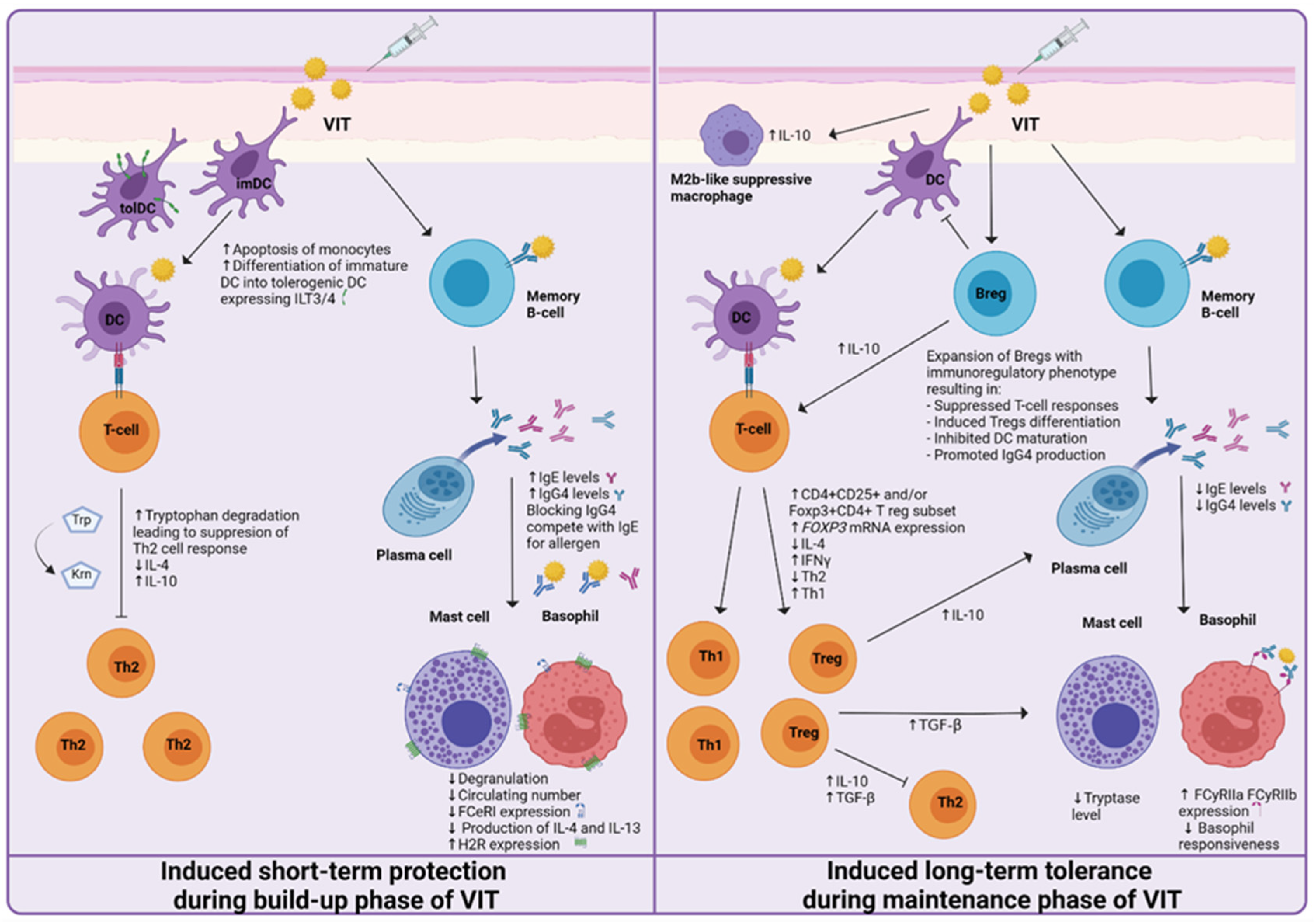
4. Mechanisms of Short-Term Protection
4.1. Basophils and Mast Cells
4.2. Regulatory Cells of the Immune System
4.3. Other Immune Cells
4.4. Antibodies
4.5. Cytokines
5. Mechanisms of Induced Long-Term Tolerance
5.1. Basophils and Mast Cells
5.2. Regulatory Cells of the Immune System
5.3. Other Immune Cells
5.4. Antibodies
5.5. Cytokines
6. Recurrent Systemic Reaction to a Sting after Stopping VIT
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HVA | Hymenoptera venom allergy |
| LLR | Large local reaction |
| SR | Systemic reaction |
| VIT | Venom immunotherapy |
| MD | Maintenance dose |
| Th2 | T helper cell type 2 |
| Tregs | Regulatory T cells |
| Bregs | Regulatory B cells |
| Tr1 | IL-10 producing Tregs |
| Br1 | CD73- CD25+ CD71+ Bregs secreting IL-10 |
| IL | Interleukin |
| Ig | Immunoglobulin |
| sIgE | Allergen specific immunoglobulin E |
| FCƐRI | High-affinity IgE receptor |
| FcγRIIa | Low-affinity IgG receptors |
| FcγRIIb | Low-affinity IgG receptors |
| DC | Dendritic cell |
| APCs | Antigen presenting cells |
| ILT | Inhibitory receptor immunoglobulin-like transcript |
| Foxp3 | Transcript factor forkhead box protein 3 |
| PBMC | Peripheral blood mononuclear cells |
| BAT | Basophil activation test |
References
- Akdis, C.A. Allergy and hypersensitivity: Mechanisms of allergic disease. Curr. Opin. Immunol. 2006, 18, 718–726. [Google Scholar] [CrossRef]
- Fujita, H.; Soyka, M.B.; Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy. Clin. Transl. Allergy 2012, 2, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Głobińska, A.; Boonpiyathad, T.; Satitsuksanoa, P.; Kleuskens, M.; van de Veen, W.; Sokolowska, M.; Akdis, M. Mechanisms of allergen-specific immunotherapy: Diverse mechanisms of immune tolerance to allergens. Ann. Allergy Asthma Immunol. 2018, 121, 306–312. [Google Scholar] [CrossRef]
- Biló, B.M.; Rueff, F.; Mosbech, H.; Bonifazi, F.; Oude-Elberink, J.N. EAACI Interest Group on Insect Venom Hypersensitivity. Diagnosis of Hymenoptera venom allergy. Allergy 2005, 60, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Bilò, B.M.; Bonifazi, F. Epidemiology of insect-venom anaphylaxis. Curr. Opin. Allergy Clin. Immunol. 2008, 8, 330–337. [Google Scholar] [CrossRef]
- Bilò, M.B. Anaphylaxis caused by Hymenoptera stings: From epidemiology to treatment. Allergy 2011, 66, 35–37. [Google Scholar] [CrossRef]
- Sturm, G.J.; Varga, E.M.; Roberts, G.; Mosbech, H.; Bilò, M.B.; Akdis, C.A.; Antolín-Amérigo, D.; Cichocka-Jarosz, E.; Gawlik, R.; Jakob, T.; et al. EAACI guidelines on allergen immunotherapy: Hymenoptera venom allergy. Allergy 2018, 73, 744–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenstein, L.M.; Valentine, M.D.; Sobotka, A.K. A case for venom treatment in anaphylactic sensitivity to hymenoptera sting. N. Engl. J. Med. 1974, 290, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Golden, D.B.; Valentine, M.D.; Kagey-Sobotka, A.; Lichtenstein, L.M. Regimens of Hymenoptera venom immunotherapy. Ann. Intern. Med. 1980, 92, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Sturm, G.; Kränke, B.; Rudolph, C.; Aberer, W. Rush Hymenoptera venom immunotherapy: A safe and practical protocol for high-risk patients. J. Allergy Clin. Immunol. 2002, 110, 928–933. [Google Scholar] [CrossRef]
- Bonifazi, F.; Jutel, M.; Biló, B.M.; Birnbaum, J.; Muller, U. EAACI Interest Group on Insect Venom Hypersensitivity. Prevention and treatment of hymenoptera venom allergy: Guidelines for clinical practice. Allergy 2005, 60, 1459–1470. [Google Scholar] [CrossRef]
- (EAACI) The European Academy of Allergology and Clinical Immunology. Position paper: Immunotherapy with hymenoptera venoms. Allergy 1993, 48, 36–46. [Google Scholar]
- Patella, V.; Florio, G.; Giuliano, A.; Oricchio, C.; Spadaro, G.; Marone, G.; Genovese, A. Hymenoptera Venom Immunotherapy: Tolerance and Efficacy of an Ultrarush Protocol versus a Rush and a Slow Conventional Protocol. J. Allergy 2012, 2012, 192192. [Google Scholar] [CrossRef] [Green Version]
- Pospischil, I.M.; Kagerer, M.; Cozzio, A.; Angelova-Fischer, I.; Guenova, E.; Ballmer-Weber, B.; Hoetzenecker, W. Comparison of the Safety Profiles of 3 Different Hymenoptera Venom Immunotherapy Protocols: A Retrospective 2-Center Study of 143 Patients. Int. Arch. Allergy Immunol. 2020, 181, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Müller, U.; Helbling, A.; Berchtold, E. Immunotherapy with honeybee venom and yellow jacket venom is different regarding efficacy and safety. J. Allergy Clin. Immunol. 1992, 89, 529–535. [Google Scholar] [CrossRef]
- Ruëff, F.; Vos, B.; Oude Elberink, J.; Bender, A.; Chatelain, R.; Dugas-Breit, S.; Horny, H.P.; Küchenhoff, H.; Linhardt, A.; Mastnik, S.; et al. Predictors of clinical effectiveness of Hymenoptera venom immunotherapy. Clin. Exp. Allergy 2014, 44, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.; Hawranek, T. Hymenoptera Venom Immunotherapy and Field Stings. J. Investig. Allergol. Clin. Immunol. 2006, 16, 224–231. [Google Scholar] [PubMed]
- Sindher, S.B.; Long, A.; Acharya, S.; Sampath, V.; Nadeau, K.C. The Use of Biomarkers to Predict Aero-Allergen and Food Immunotherapy Responses. Clin. Rev. Allergy Immunol. 2018, 55, 190–204. [Google Scholar] [CrossRef]
- Krishna, M.T.; Ewan, P.W.; Diwakar, L.; Durham, S.R.; Frew, A.J.; Leech, S.C.; Nasser, S.M. Diagnosis and management of hymenoptera venom allergy: British Society for Allergy and Clinical Immunology (BSACI) guidelines. Clin. Exp. Allergy 2011, 41, 1201–1220. [Google Scholar] [CrossRef] [PubMed]
- Aßmus, K.; Meissner, M.; Kaufmann, R.; Valesky, E.M. Benefits and limitations of sting challenge in hymenoptera venom allergy. Allergol. Select. 2021, 5, 45–50. [Google Scholar] [CrossRef]
- Bonadonna, P.; Zanotti, R.; Pagani, M.; Bonifacio, M.; Scaffidi, L.; Oliveri, E.; Franchini, M.; Reccardini, F.; Constantino, M.T.; Roncallo, C.; et al. Anaphylactic Reactions After Discontinuation of Hymenoptera Venom Immunotherapy: A Clonal Mast Cell Disorder Should Be Suspected. J. Allergy Clin. Immunol. Pract. 2018, 6, 1368–1372. [Google Scholar] [CrossRef]
- Šelb, J.; Rijavec, M.; Eržen, R.; Zidarn, M.; Kopač, P.; Škerget, M.; Bajrović, N.; Luzar, A.D.; Park, Y.H.; Liu, Y.; et al. Routine KIT p.D816V screening identifies clonal mast cell disease in patients with Hymenoptera allergy regularly missed using baseline tryptase levels alone. J. Allergy Clin. Immunol. 2021, 19, S0091–S6749. [Google Scholar] [CrossRef]
- Patella, V.; Florio, G.; Spadaro, G. Hymenoptera venom allergy: A new ultra-rush immunotherapy. In Cellular and Molecular Targets in Allergy and Clinical Immunology; Holgate, S.T., Marone, G., Ring, J., Eds.; Hogrefe and Huber: Göttingen, Germany, 2008; p. 354. [Google Scholar]
- Poulsen, L.K.; Hummelshoj, L. Triggers of IgE class switching and allergy development. Ann. Med. 2007, 39, 440–456. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradding, P.; Walls, A.F.; Holgate, S.T. The role of the mast cell in the pathophysiology of asthma. J. Allergy Clin. Immunol. 2006, 117, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Van Zelm, M.C.; McKenzie, C.I.; Varese, N.; Rolland, J.M.; O’Hehir, R.E. Recent developments and highlights in immune monitoring of allergen immunotherapy. Allergy 2019, 74, 2342–2354. [Google Scholar] [CrossRef] [Green Version]
- Sahiner, U.M.; Durham, S.R. Hymenoptera Venom Allergy: How Does Venom Immunotherapy Prevent Anaphylaxis From Bee and Wasp Stings? Front. Immunol. 2019, 10, 1959. [Google Scholar] [CrossRef]
- Goldberg, A.; Confino-Cohen, R. Bee venom immunotherapy - how early is it effective? Allergy 2010, 65, 391–395. [Google Scholar] [CrossRef]
- Chirumbolo, S.; Bjorklund, G.; Sboarina, A.; Vella, A. The role of basophils as innate immune regulatory cells in allergy and immunotherapy. Hum. Vaccin. Immunother. 2018, 14, 815–831. [Google Scholar] [CrossRef]
- Plewako, H.; Wosińska, K.; Arvidsson, M.; Björkander, J.; Skov, P.S.; Håkansson, L.; Rak, S. Basophil Interleukin 4 and Interleukin 13 Production Is Suppressed during the Early Phase of Rush Immunotherapy. Int. Arch. Allergy Immunol. 2006, 141, 346–353. [Google Scholar] [CrossRef]
- Novak, N.; Mete, N.; Bussmann, M.D.; Maintz, L.; Bieber, T.; Akdis, M.; Zumkehr, J.; Jutel, M.; Akdis, C. Early supression of basophil activation during allergen-specific immunotherapy by histamine receptor 2. J. Allergy Clin. Immunol. 2012, 130, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Dy, M.; Schneider, E. Histamine–cytokine connection in immunity and hematopoiesis. Cytokine Growth Factor Rev. 2004, 15, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Čelesnik, N.; Vesel, T.; Rijavec, M.; Šilar, M.; Eržen, R.; Košnik, M.; Kloft Žitnik, S.E.; Avčin, T.; Korošec, P. Short-term venom immunotherapy induces desensitization of FcεRI-mediated basophil response. Allergy 2012, 67, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- MacGlashan, D., Jr. IgE receptor and signal transduction in mast cells and basophils. Curr. Opin. Immunol. 2008, 20, 717–723. [Google Scholar] [CrossRef]
- Rodriguez Trabado, A.; Camara Hijon, C. Short-, Intermediate-, and Long-Term Changes in Basophil Reactivity Induced by Venom Immunotherapy. Allergy Asthma Immunol. Res. 2016, 8, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Jutel, M.; Müller, U.R.; Fricker, M.; Rihs, S.; Pichler, W.J.; Dahinden, C. Influence of bee venom immunotherapy on degranulation and leukotriene generation in human blood basophils. Clin. Exp. Allergy 1996, 26, 1112–1118. [Google Scholar] [CrossRef]
- Čelesnik Smodiš, N.; Šilar, M.; Eržen, R.; Rijavec, M.; Košnik, M.; Korošec, P. Down-regulation of FcεRI-mediated CD63 basophil response during short-term VIT determined venom-nonspecific desensitization. PLoS ONE 2014, 14, e94762. [Google Scholar] [CrossRef]
- Palomares, O.; Akdis, M.; Martin-Fontecha, M.; Akdis, C.A. Mechanisms of immune regulation in allergic diseases: The role of regulatory T and B cells. Immunol Rev. 2017, 278, 219–236. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [Green Version]
- Palomares, O.; Martin-Fontecha, M.; Lauener, R.; Traidl-Hoffmann, C.; Cavakaytar, O.; Akdis, M.; Akdis, C.A. Regulatory T cells and immune regulation of allergic diseases: Roles of IL-10 and TGF- beta. Genes Immun. 2014, 15, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Zakrzewski, A.; Kruszewski, J.; Chciałowski, A.; Kłos, K.; Rzeszotarska, A.; Korsak, J.; Nowosielska, E.M.; Cheda, A.; Wrembel-Wargocka, J.; Janiak, M.K. How fast does wasp venom immunotherapy affect a regulatory T cell subpopulation (CD4+ CD25+ Foxp3+) and the synthesis of interleukins 10, 21 and transforming growth factor β1? Postepy Dermatol. Alergol. 2019, 36, 82–85. [Google Scholar] [CrossRef]
- Bussmann, C.; Xia, J.; Allam, J.P.; Maintz, L.; Bieber, T.; Novak, N. Early markers for protective mechanisms during rush venom immunotherapy. Allergy 2010, 65, 1558–1565. [Google Scholar] [CrossRef]
- Müller, U.; Helbling, A.; Bischof, M. Predictive value of venom-specific IgE, IgG and IgG subclass antibodies in patients on immunotherapy with honey bee venom. Allergy 1989, 44, 412–418. [Google Scholar] [CrossRef]
- Flicker, S.; Valenta, R. Renaissance of the blocking antibody concept in type I allergy. Int Arch. Allergy Immunol. 2003, 132, 13–24. [Google Scholar] [CrossRef]
- van Neerven, R.J.; Wikborg, T.; Lund, G.; Jacobsen, B.; Brinch-Nielsen, A.; Arnved, J.; Ipsen, H. Blocking antibodies induced by specific allergy vaccination prevent the activation of CD4+ T cells by inhibiting serum-IgE-facilitated allergen presentation. J. Immunol. 1999, 163, 2944–2952. [Google Scholar] [PubMed]
- Golden, D.B.; Meyers, D.A.; Kagey-Sobotka, A.; Valentine, M.D.; Lichtenstein, L.M. Clinical relevance of the venom-specific immunoglobulin G antibody level during immunotherapy. J. Allergy Clin. Immunol. 1982, 69, 489–493. [Google Scholar] [CrossRef]
- Akdis, C.A.; Blesken, T.; Akdis, M.; Wüthrich, B.; Blaser, K. Role of interleukin 10 in specific immunotherapy. J. Clin. Invest. 1998, 102, 98–106. [Google Scholar] [CrossRef] [PubMed]
- McHugh, S.M.; Deighton, J.; Stewart, A.G.; Lachmann, P.J.; Ewan, P.W. Bee venom immunotherapy induces a shift in cytokine responses from a TH-2 to a TH-1 dominant pattern: Comparison of rush and conventional immunotherapy. Clin. Exp. Allergy 1995, 25, 828–838. [Google Scholar] [CrossRef] [PubMed]
- Randolph, C.C.; Reisman, R.E. Evaluation of decline in serum venom-specific IgE as a criterion for stopping venom immunotherapy. J. Allergy Clin. Immunol. 1986, 77, 823–827. [Google Scholar] [CrossRef]
- Reisman, R.E. Duration of venom immunotherapy: Relationship to the severity of symptoms of initial insect sting anaphylaxis. J. Allergy Clin. Immunol. 1993, 92, 831–836. [Google Scholar] [CrossRef]
- Keating, M.U.; Kagey-Sobotka, A.; Hamilton, R.G.; Yunginger, J.W. Clinical and immunologic follow-up of patients who stop venom immunotherapy. J. Allergy Clin. Immunol. 1991, 88, 339–348. [Google Scholar] [CrossRef]
- Golden, D.B.; Kwiterovich, K.A.; Kagey-Sobotka, A.; Valentine, M.D.; Lichtenstein, L.M. Discontinuing venom immunotherapy: Outcome after five years. J. Allergy Clin. Immunol. 1996, 97, 579–587. [Google Scholar] [CrossRef]
- Peternelj, A.; Silar, M.; Erzen, R.; Kosnik, M.; Korosec, P. Basophil sensitivity in patients not responding to venom immunotherapy. Int. Arch. Allergy Immunol. 2008, 146, 248–254. [Google Scholar] [CrossRef]
- Eržen, R.; Košnik, M.; Silar, M.; Korošec, P. Basophil response and the induction of a tolerance in venom immunotherapy: A long-term sting challenge study. Allergy 2012, 67, 822–830. [Google Scholar] [CrossRef]
- Hausmann, O.; Diaz, C.; Schneider, M.; Weber, J.; Pecaric-Petkovic, T.; Helbling, A. Correlation of sting challenge outcome and change in EC50 in basophil activation test (BAT) in bee venom allergic patients after 2-5 years of venom immunotherapy. Allergy 2014, 69, 111. [Google Scholar]
- Cady, C.T.; Powell, M.S.; Harbeck, R.J.; Giclas, P.C.; Murphy, J.R.; Katial, R.K.; Weber, R.W.; Hogarth, P.M.; Johnson, S.; Bonvini, E.; et al. IgG antibodies produced during subcutaneous allergen immunotherapy mediate inhibition of basophil activation via a mechanism involving both FcgammaRIIA and FcgammaRIIB. Immunol. Lett. 2010, 130, 57–65. [Google Scholar] [CrossRef] [Green Version]
- MacGlashan, D., Jr.; Hamilton, R.G. Parameters determining the efficacy of CD32 to inhibit activation of FcεRI in human basophils. J. Allergy Clin. Immunol. 2016, 137, 1256–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, K.; Rydent, F.; Broos, S.; Andersson, M.; Greiff, L.; Lindstedt, M. C-type lectin receptor expression on human basophils and effects of allergen-specific immunotherapy. Scand. J. Immunol. 2016, 84, 150–157. [Google Scholar] [CrossRef]
- Dugas-Breit, S.; Przybilla, B.; Dugas, M.; Arnold, A.; Pfundstein, G.; Küchenhoff, H.; Ruëff, F. Serum concentration of baseline mast cell tryptase: Evidence for a decline during long-term immunotherapy for Hymenoptera venom allergy. Clin. Exp. Allergy 2010, 40, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Santos, M.C.; Baptista, A.P.; Melo, A.; Alves, R.R.; Soares, R.S.; Pedro, E.; Pereira-Barbosa, M.; Victorino, R.M.; Sousa, A.E. Expansion of circulating Foxp3+) D25bright CD4+ T cells during specific venom immunotherapy. Clin. Exp. Allergy 2008, 38, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Ramsdell, F.; Rudensky, A.Y. Foxp3: A genetic foundation for regulatory T cell differentiation and function. Nat. Immunol. 2020, 21, 708–709. [Google Scholar] [CrossRef]
- Plitas, G.; Rudensky, A.Y. Regulatory T Cells: Differentiation and Function. Cancer Immunol. Res. 2016, 4, 721–725. [Google Scholar] [CrossRef] [Green Version]
- Meiler, F.; Klunker, S.; Zimmermann, M.; Akdis, C.A.; Akdis, M. Distinct regulation of IgE, IgG4 and IgA by T regulatory cells and toll-like receptors. Allergy 2008, 63, 1455–1463. [Google Scholar] [CrossRef] [PubMed]
- Soyer, O.U.; Akdis, M.; Ring, J.; Behrendt, H.; Crameri, R.; Lauener, R.; Akdis, C.A. Mechanisms of peripheral tolerance to allergens. Allergy 2013, 68, 161–170. [Google Scholar] [CrossRef]
- van de Veen, W.; Stanic, B.; Wirz, O.F.; Jansen, K.; Globinska, A.; Akdis, M. Role of regulatory B cells in immune tolerance to allergens and beyond. J. Allergy Clin. Immunol. 2016, 138, 654–665. [Google Scholar] [CrossRef] [Green Version]
- van de Veen, W.; Stanic, B.; Yaman, G.; Wawrzyniak, M.; Sollner, S.; Akdis, D.G.; Ruckert, B.; Akdis, C.A.; Akdis, M. IgG4 production is confined to human IL-10- producing regulatory B cells that suppress antigen-specific immune responses. J. Allergy Clin. Immunol. 2013, 131, 1204–1212. [Google Scholar] [CrossRef]
- Kucuksezer, U.C.; Ozdemir, C.; Cevhertas, L.; Ogulur, I.; Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy and allergen tolerance. Allergol. Int. 2020, 69, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Van de Veen, W. The role of regulatory B cells in allergen immunotherapy. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 447–452. [Google Scholar] [CrossRef]
- Boonpiyathad, T.; Meyer, N.; Moniuszko, M.; Sokolovska, M.; Eljaszewicz, A.; Wirz, O.F.; Tomasiak-Lozowska, M.M.; Bondzenta-Lukaszyk, A.; Ruxrungtham, K.; van de Veen, W. High-dose bee venom exposure induces similar tolerogenic B-cell responses in allergic patients and healthy beekeepers. Allergy 2017, 72, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, R.; Roth-Walter, F.; Ohradanova-Repic, A.; Flicker, S.; Hufnagl, K.; Fischer, M.B.; Stockinger, H.; Jensen-Jarolim, E. IgG4 drives M2a macrophages to a regulatory M2b-like phenotype: Potential implication in immune tolerance. Allergy 2019, 74, 483–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoos, A.M.; Bullens, D.; Chawes, B.L.; Costa, J.; De Vlieger, L.; DunnGalvin, A.; Epstein, M.M.; Garssen, J.; Hilger, C.; Knipping, K.; et al. Immunological Outcomes of Allergen-Specific Immunotherapy in Food Allergy. Front. Immunol. 2020, 11, 568–598. [Google Scholar] [CrossRef] [PubMed]
- Scadding, G.W.; Calderon, M.A.; Shamji, M.H.; Eifan, A.O.; Penagos, M.; Dumitru, F.; Sever, M.L.; Bahnson, H.T.; Lawson, K.; Harris, K.M.; et al. Effect of 2 years of treatment with sublingual grass pollen immunotherapy on nasal reponse to allergen challenge at 3 years among patients with moderate to severe seasonal allergic rhinitis: The GRASS randomized clinical trial. JAMA 2017, 317, 615–625. [Google Scholar] [CrossRef]
- James, L.K.; Shamji, M.H.; Walker, S.M.; Wilson, D.R.; Wachholz, P.A.; Francis, J.N.; Durham, S.R. Long-term tolerance after allergen immunotherapy is accompanied by selective persistence of blocking antibodies. J. Allergy Clin. Immunol. 2011, 127, 509–516. [Google Scholar] [CrossRef]
- Varga, E.M.; Francis, J.N.; Zach, M.S.; Klunker, S.; Aberer, W.; Durham, S.R. Time course of serum inhibitory activity for facilitated allergen-IgE binding during bee venom immunotherapy in children. Clin. Exp. Allergy 2009, 39, 1353–1357. [Google Scholar] [CrossRef] [PubMed]
- Albanesi, M.; Nico, A.; Sinisi, A.; Giliberti, L.; Rossi, M.P.; Rossini, M.; Kourtis, G.; Rucco, A.S.; Loconte, F.; Muolo, L.; et al. A 13-year real-life study on efficacy, safety and biological effects of Vespula venom immunotherapy. Clin. Mol. Allergy 2018, 16, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Jin, W.; Hardegen, N.; Lei, K.J.; Li, L.; Marinos, N.; McGrady, G.; Wahl, M.S. Conversion of peripheral CD4 + CD25- naive T cells to CD4 + CD25 + regulatory T cells by TGF-beta induction of transcription factor Foxp3. J. Exp. Med. 2003, 198, 1875–1886. [Google Scholar] [CrossRef] [PubMed]
- Adelmeyer, J.; Pickert, J.; Pfützner, W.; Möbs, C. Long-term impact of hymenoptera venom immunotherapy on clinical course, immune parameters, and psychosocial aspects. Allergol. Select. 2021, 5, 57–66. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Day 1 | Day 2 | Day 3 | Day 4 | Day 8 | Day 11 | Day 15 | Day 22 | Day 29 | Day 36 | Day 43 | Day 50 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ultra-rush | 0.1–* 100 µg | * 2 × 50 µg | * 100 µg | |||||||||
| Rush | 0.01–2 µg | 4–20 µg | 40–80 µg | * 100 µg | * 100 µg | |||||||
| Cluster | 0.001–0.1 µg | 1–10 µg | 20–30 µg | 2 × 50 µg | * 100 µg | * 100 µg | ||||||
| Conventional | 0.01–0.1 µg | 1–2 µg | 4–8 µg | 10–20 µg | 40 µg | 60 µg | 80 µg | * 100 µg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demšar Luzar, A.; Korošec, P.; Košnik, M.; Zidarn, M.; Rijavec, M. Hymenoptera Venom Immunotherapy: Immune Mechanisms of Induced Protection and Tolerance. Cells 2021, 10, 1575. https://doi.org/10.3390/cells10071575
Demšar Luzar A, Korošec P, Košnik M, Zidarn M, Rijavec M. Hymenoptera Venom Immunotherapy: Immune Mechanisms of Induced Protection and Tolerance. Cells. 2021; 10(7):1575. https://doi.org/10.3390/cells10071575
Chicago/Turabian StyleDemšar Luzar, Ajda, Peter Korošec, Mitja Košnik, Mihaela Zidarn, and Matija Rijavec. 2021. "Hymenoptera Venom Immunotherapy: Immune Mechanisms of Induced Protection and Tolerance" Cells 10, no. 7: 1575. https://doi.org/10.3390/cells10071575
APA StyleDemšar Luzar, A., Korošec, P., Košnik, M., Zidarn, M., & Rijavec, M. (2021). Hymenoptera Venom Immunotherapy: Immune Mechanisms of Induced Protection and Tolerance. Cells, 10(7), 1575. https://doi.org/10.3390/cells10071575