
The Feasibility and Applicability of Stem Cell Therapy for the Cure of Type 1 Diabetes

 and
and
Abstract
:1. Introduction
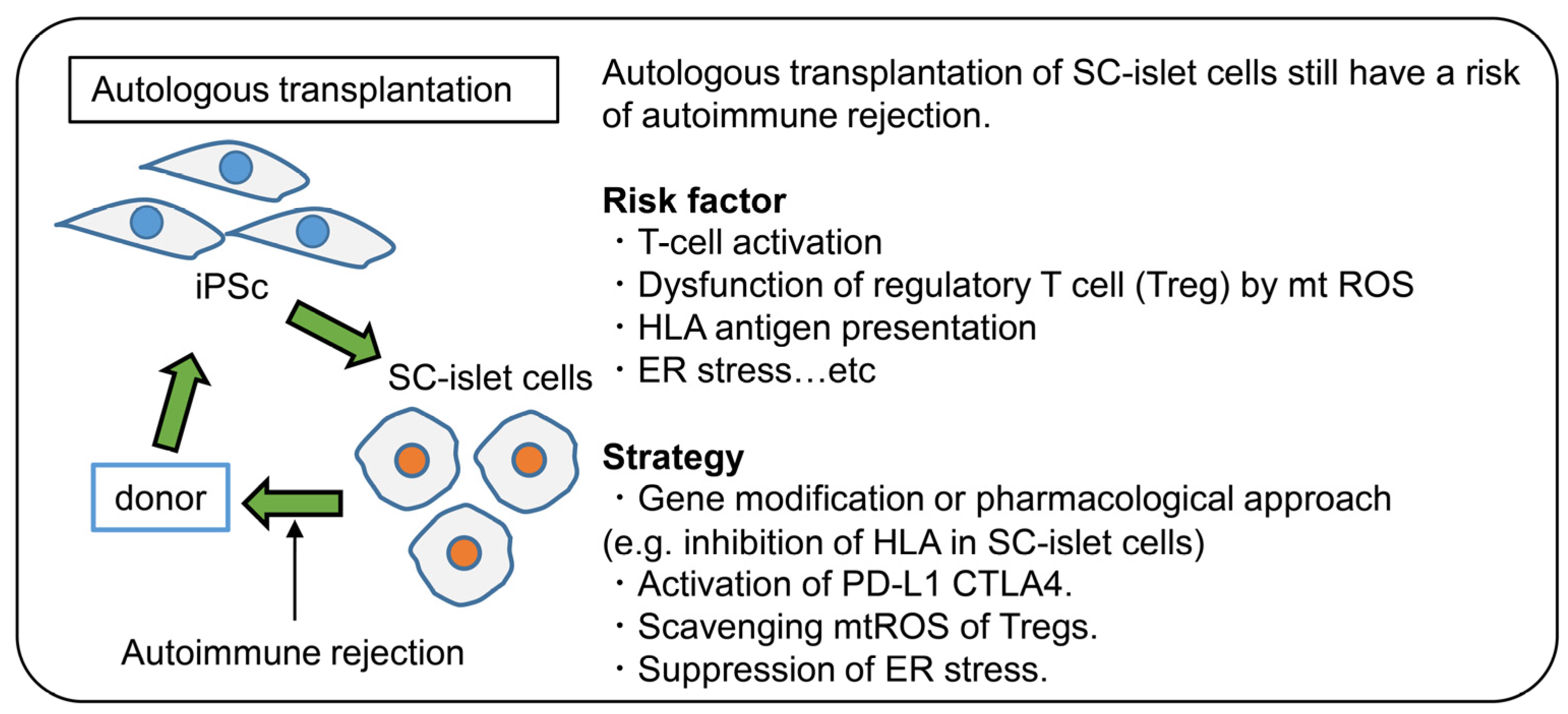
2. Autoimmune Responses in Stem Cell Therapy for T1D
3. Comparison of Stem Cell Therapy for Pancreatic β-Cells with Automated Insulin Delivery System
3.1. Stem Cell Therapy
3.2. Artificial Pancreas
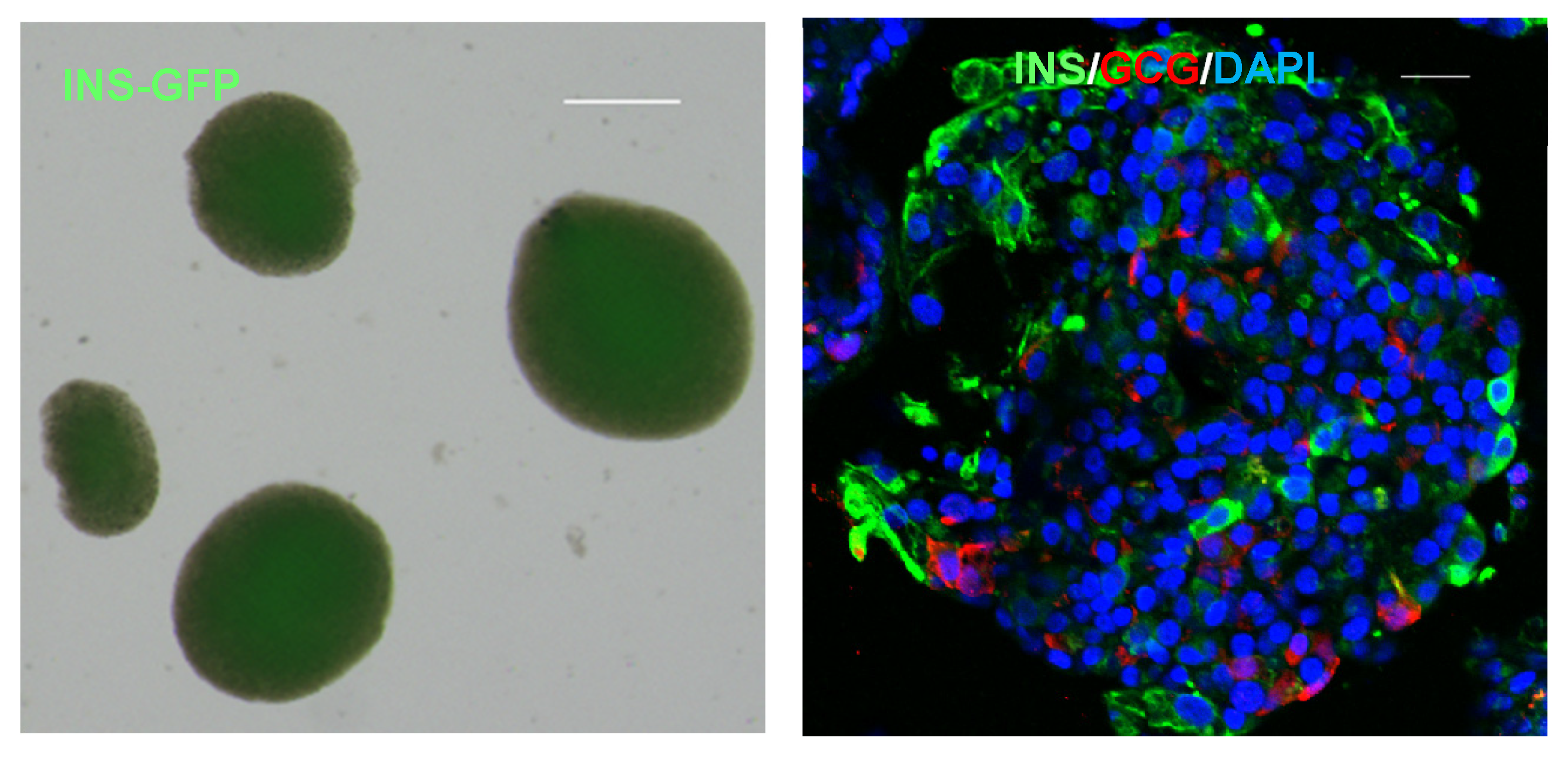
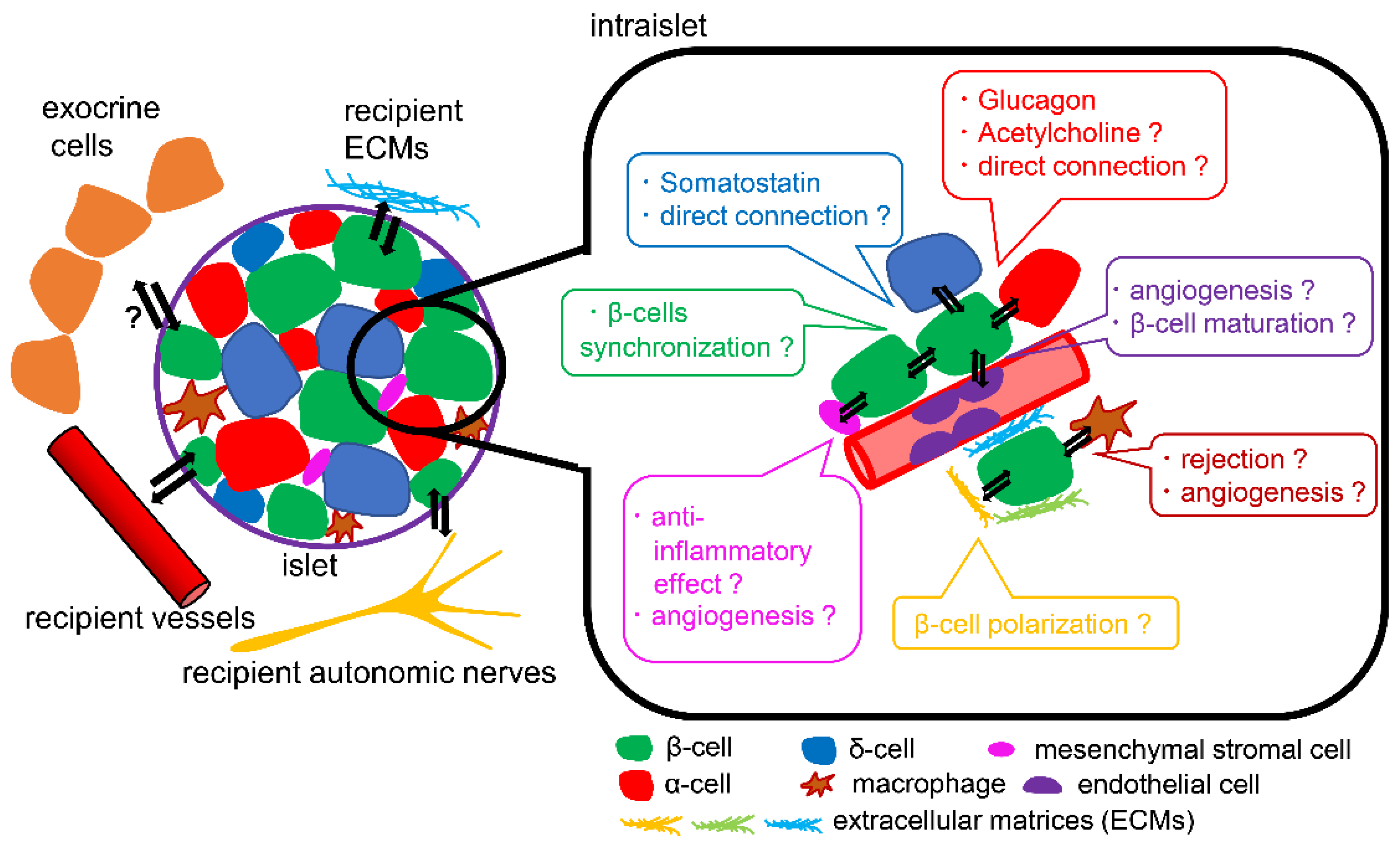
4. Islet Microenvironment and SC-Islet Cells
4.1. Endocrine Cells
4.2. Exocrine Cells
4.3. Mesenchymal Stromal Cells, Endothelial Cells, Nerves, and Macrophages
4.4. Extracellular Matrices
4.5. Transplantation Site
5. Cost-Effectiveness of Stem Cell-Derived Islet Cells Transplantation Therapy for T1D
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Todd, J.A. Etiology of type 1 diabetes. Immunity 2010, 32, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, M.A.; Eisenbarth, G.S.; Michels, A.W. Type 1 diabetes. Lancet 2014, 383, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Geho, W.B. The importance of the liver in insulin replacement therapy in insulin-deficient diabetes. Diabetes 2014, 63, 1445–1447. [Google Scholar] [CrossRef] [Green Version]
- Bergenstal, R.M.; Klonoff, D.C.; Garg, S.K.; Bode, B.W.; Meredith, M.; Slover, R.H.; Ahmann, A.J.; Welsh, J.B.; Lee, S.W.; Kaufman, F.R. Threshold-based insulin-pump interruption for reduction of hypoglycemia. N. Engl. J. Med. 2013, 369, 224–232. [Google Scholar] [CrossRef] [Green Version]
- Wojtusciszyn, A.; Branchereau, J.; Esposito, L.; Badet, L.; Buron, F.; Chetboun, M.; Kessler, L.; Morelon, E.; Berney, T.; Pattou, F.; et al. Indications for islet or pancreatic transplantation: Statement of the TREPID working group on behalf of the Société francophone du diabète (SFD), Société francaise d’endocrinologie (SFE), Société francophone de transplantation (SFT) and Société française de néphrologie-dialyse -transplantation (SFNDT). Diabetes Metab. 2019, 45, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Scalea, J.R.; Pettinato, L.; Fiscella, B.; Bartosic, A.; Piedmonte, A.; Paran, J.; Todi, N.; Siskind, E.J.; Bartlett, S.T. Successful pancreas transplantation alone is associated with excellent self-identified health score and glucose control: A retrospective study from a high-volume center in the United States. Clin. Transplant. 2018, 32. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.M.; Lakey, J.R.; Ryan, E.A.; Korbutt, G.S.; Toth, E.; Warnock, G.L.; Kneteman, N.M.; Rajotte, R.V. Islet transplantation in seven patients with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive regimen. N. Engl. J. Med. 2000, 343, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Lablanche, S.; Vantyghem, M.C.; Kessler, L.; Wojtusciszyn, A.; Borot, S.; Thivolet, C.; Girerd, S.; Bosco, D.; Bosson, J.L.; Colin, C.; et al. Islet transplantation versus insulin therapy in patients with type 1 diabetes with severe hypoglycaemia or poorly controlled glycaemia after kidney transplantation (TRIMECO): A multicentre, randomised controlled trial. Lancet Diabetes Endocrinol. 2018, 6, 527–537. [Google Scholar] [CrossRef]
- Bellin, M.D.; Dunn, T.B. Transplant strategies for type 1 diabetes: Whole pancreas, islet and porcine beta cell therapies. Diabetologia 2020, 63, 2049–2056. [Google Scholar] [CrossRef] [PubMed]
- Kevin, A.; D’Amour, A.G.B.; Susan, E.; Olivia, G.K.; Alan, D.A.; Nora, G.S.; Mark, A.M.; Evert, K.; Melissa, K.C.; Emmanuel, E.B. Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells. Nat. Biotechnol. 2006, 24, 1392–1401. [Google Scholar]
- Tingxia, G.; Na, L.; Li, L.; Matthias, H. Factors expressed by murine embryonic pancreatic mesenchyme enhance generation of insulin-producing cells from hESCs. Diabetes 2013, 23, 1581–1592. [Google Scholar] [CrossRef]
- Alireza, R.; Rhonda, M.J.R.; Wideman, D.; Karanu, F.; Ao, Z.; Garth, L.W.; Timothy, J.K. Production of functional glucagon-secreting a-cells from human embryonic stem cells. Diabetes 2011, 60, 239–247. [Google Scholar]
- Rezania, A.; Bruin, J.E.; Riedel, M.J.; Mojibian, M.; Asadi, A.; Xu, J.; Gauvin, R.; Narayan, K.; Karanu, F.; O’Neil, J.J. Maturation of human embryonic stem cell-derived pancreatic progenitors into functional islets capable of treating pre-existing diabetes in mice. Diabetes 2012, 61, 2016–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Melton, D.A. Pancreas regeneration. Nature 2018, 557, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Loo, L.S.W.; Soetedjo, A.A.P.; Lau, H.H.; Ng, N.H.J.; Ghosh, S.; Nguyen, L.; Krishnan, V.G.; Choi, H.; Roca, X.; Hoon, S.; et al. BCL-xL/BCL2L1 is a critical anti-apoptotic protein that promotes the survival of differentiating pancreatic cells from human pluripotent stem cells. Cell Death Disease 2020, 11, 378–396. [Google Scholar] [CrossRef]
- Julie, B.; Sneddon, Q.T.; Peter, S.; Jeffrey, A.; Bluestone, S.R.; Tejal, D.; Matthias, H. Stem Cell Therapies for Treating Diabetes: Progress and Remaining Challenges. Cell Stem Cell 2018, 22, 811–823. [Google Scholar]
- Nair, G.G.; Liu, J.S.; Russ, H.A.; Tran, S.; Saxton, M.S.; Chen, R.; Juang, C.; Li, M.L.; Nguyen, V.Q.; Giacometti, S. Recapitulating endocrine cell clustering in culture promotes maturation of human stem-cell-derived β cells. Nat. Cell Biol. 2019, 21, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Bader, E.; Migliorini, A.; Gegg, M.; Moruzzi, N.; Sara, J.C.; Roscioni, S.S.; Bakhti, M.; Brandl, E.; Irmler, M.; Beckers, J.; et al. Identification of proliferative and mature β-cells in the islets of Langerhans. Nature 2016, 535, 430–441. [Google Scholar] [CrossRef]
- Britt, L.D.; Stojeba, P.C.; Scharp, C.R.; Greider, M.H.; Scharp, D.W. Neonatal pig pseudo-islets. A product of selective aggregation. Diabetes 1981, 30, 580–583. [Google Scholar] [CrossRef]
- Gingerich, R.L.; Scharp, D.W.; Greider, M.H.; Dye, E.S.; Mousel, K.A. A new in vitro model for studies of pancreatic polypeptide secretion and biochemistry. Regul. Pept. 1982, 5, 13–25. [Google Scholar] [CrossRef]
- Micallef, S.J.; Li, X.; Schiesser, J.V.; Hirst, C.E.; Yu, Q.C.; Lim, S.M.; Nostro, M.C.; Elliott, D.A.; Sarangi, F.; Harrison, L.C.; et al. INSGFP_w human embryonic stem cells facilitate isolation of in vitro derived insulin-producing cells. Diabetologia 2012, 55, 694–706. [Google Scholar] [CrossRef] [Green Version]
- Eiji Yoshihara, C.O.C.; Gasser, E.; Wei, Z.; Oh, T.G.; Tiffany, W.T.; Wang, D.; Cayabyab, F.; Dai, Y.; Ruth, T.Y.; Liddle, C.; et al. Immune-evasive human islet-like organoids ameliorate diabetes. Nature 2020. [Google Scholar] [CrossRef]
- Araki, R.; Uda, M.; Hoki, Y.; Sunayama, M.; Nakamura, M.; Ando, S.; Sugiura, M.; Ideno, H.; Shimada, A.; Nifuji, A.; et al. Negligible immunogenicity of terminally differentiated cells derived from induced pluripotent or embryonic stem cells. Nature 2013, 494, 100–104. [Google Scholar] [CrossRef]
- Guha, P.; Morgan, J.W.; Mostoslavsky, G.; Rodrigues, N.P.; Boyd, A.S. Lack of immune response to differentiated cells derived from syngeneic induced pluripotent stem cells. Cell Stem Cell 2013, 12, 407–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Vermilyea, S.C.; Zammit, M.; Lu, J.; Olsen, M.; Metzger, J.M.; Yao, L.; Chen, Y.; Phillips, S.; Holden, J.E.; et al. Autologous transplant therapy alleviates motor and depressive behaviors in parkinsonian monkeys. Nat. Med. 2021, 27, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Zhang, Z.-N.; Rong, Z.; Xu, Y. Immunogenicity of induced pluripotent stem cells. Nature 2011, 474, 212–215. [Google Scholar] [CrossRef] [Green Version]
- Pagliuca, F.W.; Millman, J.R.; Gürtler, M.; Segel, M.; Van Dervort, A.; Ryu, J.H.; Peterson, Q.P.; Greiner, D.; Melton, D.A. Generation of functional human pancreatic β cells in vitro. Cell 2014, 159, 428–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okita, K.; Matsumura, Y.; Sato, Y.; Okada, A.; Morizane, A.; Okamoto, S.; Hong, H.; Nakagawa, M.; Tanabe, K.; Tezuka, K.; et al. A more efficient method to generate integration-free human iPS cells. Nat. Methods 2011, 8, 409–412. [Google Scholar] [CrossRef] [Green Version]
- Alagpulinsa, D.A.; Cao, J.J.L.; Driscoll, R.K.; Sîrbulescu, R.F.; Penson, M.F.E.; Sremac, M.; Engquist, E.N.; Brauns, T.A.; Markmann, J.F.; Melton, D.A.; et al. Alginate-microencapsulation of human stem cell-derived β cells with CXCL12 prolongs their survival and function in immunocompetent mice without systemic immunosuppression. Am. J. Transplant. 2019, 19, 1930–1940. [Google Scholar]
- Kim, K.; Doi, A.; Wen, B.; Ng, K.; Zhao, R.; Cahan, P.; Kim, J.; Aryee, M.J.; Ji, H.; Ehrlich, L.I.; et al. Epigenetic memory in induced pluripotent stem cells. Nature 2010, 467, 285–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, K.; Semi, K.; Yamamoto, T.; Shimizu, M.; Tanaka, A.; Mitsunaga, K.; Okita, K.; Osafune, K.; Arioka, Y.; Maeda, T.; et al. Premature termination of reprogramming in vivo leads to cancer development through altered epigenetic regulation. Cell 2014, 156, 663–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osada, N.; Kikuchi, J.; Umehara, T.; Sato, S.; Urabe, M.; Abe, T.; Hayashi, N.; Sugitani, M.; Hanazono, Y.; Furukawa, Y. Lysine-specific demethylase 1 inhibitors prevent teratoma development from human induced pluripotent stem cells. Oncotarget 2018, 9, 6450–6462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. Structure of the human class I histocompatibility antigen, HLA-A2. Nature 1987, 329, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Amaya, M.; Mellins, E.; Wiley, D.C. The structure of an intermediate in class II MHC maturation: CLIP bound to HLA-DR3. Nature 1995, 378, 457–462. [Google Scholar] [CrossRef]
- Jones, E.Y.; Fugger, L.; Strominger, J.L.; Siebold, C. MHC class II proteins and disease: A structural perspective. Nat. Rev. Immunol. 2006, 6, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, Y.; Ikegami, H.; Kawaguchi, Y.; Fujisawa, T.; Shintani, M.; Ono, M.; Nishino, M.; Uchigata, Y.; Lee, I.; Ogihara, T. Asian-specific HLA haplotypes reveal heterogeneity of the contribution of HLA-DR and -DQ haplotypes to susceptibility to type 1 diabetes. Diabetes 2002, 51, 545–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoneim, M.A.; Refaie, A.F.; Elbassiouny, B.L.; Gabr, M.M.; Zakaria, M.M. From Mesenchymal Stromal/Stem Cells to Insulin-Producing Cells: Progress and Challenges. Stem Cell Rev. Rep. 2020, 16, 1156–1172. [Google Scholar] [CrossRef]
- Yang, X.F.; Chen, T.; Ren, L.W.; Yang, L.; Qi, H.; Li, F.R. Immunogenicity of insulin-producing cells derived from human umbilical cord mesenchymal stem cells. Exp. Ther. Med. 2017, 13, 1456–1464. [Google Scholar] [CrossRef]
- Cai, J.; Wu, Z.; Xu, X.; Liao, L.; Chen, J.; Huang, L.; Wu, W.; Luo, F.; Wu, C.; Pugliese, A.; et al. Umbilical Cord Mesenchymal Stromal Cell With Autologous Bone Marrow Cell Transplantation in Established Type 1 Diabetes: A Pilot Randomized Controlled Open-Label Clinical Study to Assess Safety and Impact on Insulin Secretion. Diabetes Care 2016, 39, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dassau, E.; Renard, E.; Place, J.; Farret, A.; Pelletier, M.J.; Lee, J.; Huyett, L.M.; Chakrabarty, A.; Doyle, F.J., 3rd; Zisser, H.C. Intraperitoneal insulin delivery provides superior glycaemic regulation to subcutaneous insulin delivery in model predictive control-based fully-automated artificial pancreas in patients with type 1 diabetes: A pilot study. Diabetes Obes. Metab. 2017, 19, 1698–1705. [Google Scholar] [CrossRef] [PubMed]
- Leite, N.C.; Sintov, E.; Meissner, T.B.; Brehm, M.A.; Greiner, D.L.; Harlan, D.M.; Melton, D.A. Modeling Type 1 Diabetes In Vitro Using Human Pluripotent Stem Cells. Cell Rep. 2020, 32, 107894. [Google Scholar] [CrossRef] [PubMed]
- Millman, J.R.; Xie, C.; Van Dervort, A.; Gürtler, M.; Pagliuca, F.W.; Melton, D.A. Generation of stem cell-derived β-cells from patients with type 1 diabetes. Nat. Commun. 2016, 7, 11463. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Lee, E.Y.; You, Y.H.; Yang, H.K.; Yoon, K.H.; Kim, J.W. Generation of iPSC-derived insulin-producing cells from patients with type 1 and type 2 diabetes compared with healthy control. Stem Cell Res. 2020, 48, 101958. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.; Toyoda, T.; Fukui, K.; Baden, M.Y.; Funato, M.; Kondo, Y.; Sudo, T.; Iwahashi, H.; Kishida, M.; Okada, C.; et al. Insulin-producing cells derived from ‘induced pluripotent stem cells’ of patients with fulminant type 1 diabetes: Vulnerability to cytokine insults and increased expression of apoptosis-related genes. J. Diabetes Investig. 2017, 9, 481–493. [Google Scholar] [CrossRef] [Green Version]
- Torikai, H.; Mi, T.; Gragert, L.; Maiers, M.; Najjar, A.; Ang, S.; Maiti, S.; Dai, J.; Switzer, K.C.; Huls, H.; et al. Genetic editing of HLA expression in hematopoietic stem cells to broaden their human application. Sci. Rep. 2016, 6, 21757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falcone, M.; Fousteri, G. Role of the PD-1/PD-L1 Dyad in the Maintenance of Pancreatic Immune Tolerance for Prevention of Type 1 Diabetes. Front. Endocrinol. 2020, 11, 569. [Google Scholar] [CrossRef] [PubMed]
- El Khatib, M.M.; Sakuma, T.; Tonne, J.M.; Mohamed, M.S.; Holditch, S.J.; Lu, B.; Kudva, Y.C.; Ikeda, Y. β-Cell-targeted blockage of PD1 and CTLA4 pathways prevents development of autoimmune diabetes and acute allogeneic islets rejection. Gene Ther. 2015, 22, 430–438. [Google Scholar] [CrossRef] [Green Version]
- Alissafi, T.; Kalafati, L.; Lazari, M.; Filia, A.; Kloukina, I.; Manifava, M.; Lim, J.H.; Alexaki, V.I.; Ktistakis, N.T.; Doskas, T.; et al. Mitochondrial Oxidative Damage Underlies Regulatory T Cell Defects in Autoimmunity. Cell Metab. 2020, 32, 591–604.e597. [Google Scholar] [CrossRef]
- Joshi, K.; Elso, C.; Motazedian, A.; Labonne, T.; Schiesser, J.V.; Cameron, F.; Mannering, S.I.; Elefanty, A.G.; Stanley, E.G. Induced pluripotent stem cell macrophages present antigen to proinsulin-specific T cell receptors from donor-matched islet-infiltrating T cells in type 1 diabetes. Diabetologia 2019, 62, 2245–2251. [Google Scholar] [CrossRef] [Green Version]
- Battelino, T.; Danne, T.; Bergenstal, R.M.; Amiel, S.A.; Beck, R.; Biester, T.; Bosi, E.; Buckingham, B.A.; Cefalu, W.T.; Close, K.L.; et al. Clinical Targets for Continuous Glucose Monitoring Data Interpretation: Recommendations From the International Consensus on Time in Range. Diabetes Care 2019, 42, 1593–1603. [Google Scholar] [CrossRef] [Green Version]
- Mancini, G.; Berioli, M.G.; Santi, E.; Rogari, F.; Toni, G.; Tascini, G.; Crispoldi, R.; Ceccarini, G.; Esposito, S. Flash Glucose Monitoring: A Review of the Literature with a Special Focus on Type 1 Diabetes. Nutrients 2018, 10, 992. [Google Scholar] [CrossRef] [Green Version]
- Forlenza, G.P.; Li, Z.; Buckingham, B.A.; Pinsker, J.E.; Cengiz, E.; Wadwa, R.P.; Ekhlaspour, L.; Church, M.M.; Weinzimer, S.A.; Jost, E.; et al. Predictive Low-Glucose Suspend Reduces Hypoglycemia in Adults, Adolescents, and Children With Type 1 Diabetes in an At-Home Randomized Crossover Study: Results of the PROLOG Trial. Diabetes Care 2018, 41, 2155–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breton, M.D.; Kanapka, L.G.; Beck, R.W.; Ekhlaspour, L.; Forlenza, G.P.; Cengiz, E.; Schoelwer, M.; Ruedy, K.J.; Jost, E.; Carria, L.; et al. A Randomized Trial of Closed-Loop Control in Children with Type 1 Diabetes. N. Engl. J. Med. 2020, 383, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Tanosaki, S.; Tohyama, S.; Fujita, J.; Someya, S.; Hishiki, T.; Matsuura, T.; Nakanishi, H.; Ohto-Nakanishi, T.; Akiyama, T.; Morita, Y.; et al. Fatty Acid Synthesis Is Indispensable for Survival of Human Pluripotent Stem Cells. iScience 2020, 23, 101535. [Google Scholar] [CrossRef] [PubMed]
- Yabe, S.G.; Fukuda, S.; Nishida, J.; Takeda, F.; Nashiro, K.; Okochi, H. Induction of functional islet-like cells from human iPS cells by suspension culture. Regen. Ther. 2019, 10, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Thabit, H.; Tauschmann, M.; Allen, J.M.; Leelarathna, L.; Hartnell, S.; Wilinska, M.E.; Acerini, C.L.; Dellweg, S.; Benesch, C.; Heinemann, L.; et al. Home Use of an Artificial Beta Cell in Type 1 Diabetes. N. Engl. J. Med. 2015, 373, 2129–2140. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.K.; Thorsen, S.U.; Thyssen, J.P.; Zachariae, C.; Keiding, H.; Svensson, J. Cost of Treating Skin Problems in Patients with Diabetes Who Use Insulin Pumps and/or Glucose Sensors. Diabetes Technol. Ther. 2020, 22, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.; Kerr, D. Skin-related complications of insulin therapy: Epidemiology and emerging management strategies. Am. J. Clin. Dermatol. 2003, 4, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Rezania, A.; Bruin, J.E.; Arora, P.; Rubin, A.; Batushansky, I.; Asadi, A.; O’Dwyer, S.; Quiskamp, N.; Mojibian, M.; Albrecht, T.; et al. Reversal of diabetes with insulin-producing cells derived in vitro from human pluripotent stem cells. Nat. Biotechnol. 2014, 32, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Russ, H.A.; Parent, A.V.; Ringler, J.J.; Hennings, T.G.; Nair, G.G.; Shveygert, M.; Guo, T.; Puri, S.; Haataja, L.; Cirulli, V.; et al. Controlled induction of human pancreatic progenitors produces functional beta-like cells in vitro. Embo J. 2015, 34, 1759–1772. [Google Scholar] [CrossRef] [Green Version]
- Sambathkumar, R.; Migliorini, A.; Nostro, M.C. Pluripotent Stem Cell-Derived Pancreatic Progenitors and β-Like Cells for Type 1 Diabetes Treatment. Physiology 2018, 33, 394–402. [Google Scholar] [CrossRef]
- Veres, A.; Faust, A.L.; Bushnell, H.L.; Engquist, E.N.; Kenty, J.H.; Harb, G.; Poh, Y.C.; Sintov, E.; Gürtler, M.; Pagliuca, F.W.; et al. Charting cellular identity during human in vitro β-cell differentiation. Nature 2019, 569, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.M. State of the art of clinical islet transplantation and novel protocols of immunosuppression. Curr. Diab. Rep. 2011, 11, 345–354. [Google Scholar] [CrossRef]
- Korsgren, O. Islet Encapsulation: Physiological Possibilities and Limitations. Diabetes 2017, 66, 1748–1754. [Google Scholar] [CrossRef] [Green Version]
- Galli, A.; Algerta, M.; Marciani, P.; Schulte, C.; Lenardi, C.; Milani, P.; Maffioli, E.; Tedeschi, G.; Perego, C. Shaping Pancreatic β-Cell Differentiation and Functioning: The Influence of Mechanotransduction. Cells 2020, 9, 413. [Google Scholar]
- Huising, M.O. Paracrine regulation of insulin secretion. Diabetologia 2020, 63, 2057–2063. [Google Scholar] [CrossRef]
- Moede, T.; Leibiger, I.B.; Berggren, P.O. Alpha cell regulation of beta cell function. Diabetologia 2020, 63, 2064–2075. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Diaz, R.; Tamayo, A.; Hara, M.; Caicedo, A. The Local Paracrine Actions of the Pancreatic α-Cell. Diabetes 2020, 69, 550–558. [Google Scholar] [CrossRef]
- Hartig, S.M.; Cox, A.R. Paracrine signaling in islet function and survival. J. Mol. Med. 2020, 98, 451–467. [Google Scholar] [CrossRef] [PubMed]
- van der Meulen, T.; Donaldson, C.J.; Cáceres, E.; Hunter, A.E.; Cowing-Zitron, C.; Pound, L.D.; Adams, M.W.; Zembrzycki, A.; Grove, K.L.; Huising, M.O. Urocortin3 mediates somatostatin-dependent negative feedback control of insulin secretion. Nat. Med. 2015, 21, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Vivot, K.; Moullé, V.S.; Zarrouki, B.; Tremblay, C.; Mancini, A.D.; Maachi, H.; Ghislain, J.; Poitout, V. The regulator of G-protein signaling RGS16 promotes insulin secretion and β-cell proliferation in rodent and human islets. Mol. Metab. 2016, 5, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, G.; Dawra, R.K.; Pugazhenthi, S.; Guo, Z.; Soltani, S.M.; Wiseman, A.; Sanders, M.A.; Papas, K.K.; Velayutham, K.; Saluja, A.K.; et al. Insulin degradation by acinar cell proteases creates a dysfunctional environment for human islets before/after transplantation: Benefits of α-1 antitrypsin treatment. Transplantation 2011, 92, 1222–1230. [Google Scholar] [CrossRef] [Green Version]
- Heuser, M.; Wolf, B.; Vollmar, B.; Menger, M.D. Exocrine contamination of isolated islets of Langerhans deteriorates the process of revascularization after free transplantation. Transplantation 2000, 69, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Marín-Cañas, S.; Estil Les, E.; Llado, L.; San José, P.; Nacher, M.; Téllez, N.; Montanya, E. Pancreatic ductal cells may have a negative effect on human islet transplantation. PLoS ONE 2019, 14, e0220064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichii, H.; Miki, A.; Yamamoto, T.; Molano, R.D.; Barker, S.; Mita, A.; Rodriguez-Diaz, R.; Klein, D.; Pastori, R.; Alejandro, R.; et al. Characterization of pancreatic ductal cells in human islet preparations. Lab. Investig. 2008, 88, 1167–1177. [Google Scholar] [CrossRef] [Green Version]
- Movahedi, B.; Gysemans, C.; Jacobs-Tulleneers-Thevissen, D.; Mathieu, C.; Pipeleers, D. Pancreatic duct cells in human islet cell preparations are a source of angiogenic cytokines interleukin-8 and vascular endothelial growth factor. Diabetes 2008, 57, 2128–2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dybala, M.P.; Kuznetsov, A.; Motobu, M.; Hendren-Santiago, B.K.; Philipson, L.H.; Chervonsky, A.V.; Hara, M. Integrated Pancreatic Blood Flow: Bidirectional Microcirculation Between Endocrine and Exocrine Pancreas. Diabetes 2020, 69, 1439–1450. [Google Scholar] [CrossRef]
- Khiatah, B.; Qi, M.; Du, W.K.; van Megen, K.M.; Perez, R.G.; Isenberg, J.S.; Kandeel, F.; Roep, B.O.; Ku, H.T.; Al-Abdullah, I.H. Intra-pancreatic tissue-derived mesenchymal stromal cells: A promising therapeutic potential with anti-inflammatory and pro-angiogenic profiles. Stem Cell Res. Ther. 2019, 10, 322. [Google Scholar] [CrossRef]
- Talavera-Adame, D.; Woolcott, O.O.; Ignatius-Irudayam, J.; Arumugaswami, V.; Geller, D.H.; Dafoe, D.C. Effective endothelial cell and human pluripotent stem cell interactions generate functional insulin-producing beta cells. Diabetologia 2016, 59, 2378–2386. [Google Scholar] [CrossRef] [Green Version]
- Townsend, S.E.; Gannon, M. Extracellular Matrix-Associated Factors Play Critical Roles in Regulating Pancreatic β-Cell Proliferation and Survival. Endocrinology 2019, 160, 1885–1894. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Diaz, R.; Abdulreda, M.H.; Formoso, A.L.; Gans, I.; Ricordi, C.; Berggren, P.O.; Caicedo, A. Innervation patterns of autonomic axons in the human endocrine pancreas. Cell Metab. 2011, 14, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Criscimanna, A.; Coudriet, G.M.; Gittes, G.K.; Piganelli, J.D.; Esni, F. Activated macrophages create lineage-specific microenvironments for pancreatic acinar- and β-cell regeneration in mice. Gastroenterology 2014, 147, 1106–1118.e1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brissova, M.; Aamodt, K.; Brahmachary, P.; Prasad, N.; Hong, J.Y.; Dai, C.; Mellati, M.; Shostak, A.; Poffenberger, G.; Aramandla, R.; et al. Islet microenvironment, modulated by vascular endothelial growth factor-A signaling, promotes β cell regeneration. Cell Metab. 2014, 19, 498–511. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.; Plüddemann, A. Tissue macrophages: Heterogeneity and functions. BMC Biol. 2017, 15, 53. [Google Scholar] [CrossRef]
- Lammert, E.; Thorn, P. The Role of the Islet Niche on Beta Cell Structure and Function. J. Mol. Biol. 2020, 432, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Cottle, L.; Loudovaris, T.; Xiao, D.; Yang, P.; Thomas, H.E.; Kebede, M.A.; Thorn, P. Enhanced structure and function of human pluripotent stem cell-derived beta-cells cultured on extracellular matrix. Stem Cells Transl. Med. 2020. [Google Scholar] [CrossRef]
- Sackett, S.D.; Tremmel, D.M.; Ma, F.; Feeney, A.K.; Maguire, R.M.; Brown, M.E.; Zhou, Y.; Li, X.; O’Brien, C.; Li, L.; et al. Extracellular matrix scaffold and hydrogel derived from decellularized and delipidized human pancreas. Sci. Rep. 2018, 8, 10452. [Google Scholar] [CrossRef]
- Kasoju, N.; Pátíková, A.; Wawrzynska, E.; Vojtíšková, A.; Sedlačík, T.; Kumorek, M.; Pop-Georgievski, O.; Sticová, E.; KříŽ, J.; Kubies, D. Bioengineering a pre-vascularized pouch for subsequent islet transplantation using VEGF-loaded polylactide capsules. Biomater. Sci. 2020, 8, 631–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ackermann, M.; Neufurth, M.; Wang, S.; Li, Q.; Feng, Q.; Schröder, H.C.; Müller, W.E.G. Restoration of Impaired Metabolic Energy Balance (ATP Pool) and Tube Formation Potential of Endothelial Cells under "high glucose", Diabetic Conditions by the Bioinorganic Polymer Polyphosphate. Polymers 2017, 9, 575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, E.A.; Shandro, T.; Green, K.; Paty, B.W.; Senior, P.A.; Bigam, D.; Shapiro, A.M.; Vantyghem, M.C. Assessment of the severity of hypoglycemia and glycemic lability in type 1 diabetic subjects undergoing islet transplantation. Diabetes 2004, 53, 955–962. [Google Scholar] [CrossRef] [Green Version]
- Senior, P.A.; Kin, T.; Shapiro, J.; Koh, A. Islet Transplantation at the University of Alberta: Status Update and Review of Progress over the Last Decade. Can. J. Diabetes 2012, 36, 32–37. [Google Scholar] [CrossRef]
- Tao, B.; Pietropaolo, M.; Atkinson, M.; Schatz, D.; Taylor, D. Estimating the cost of type 1 diabetes in the U.S.: A propensity score matching method. PLoS ONE 2010, 5, e11501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beckwith, J.; Nyman, J.A.; Flanagan, B.; Schrover, R.; Schuurman, H.J. A health economic analysis of clinical islet transplantation. Clin. Transplant. 2012, 26, 23–33. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. Economic Costs of Diabetes in the U.S. in 2017. Diabetes Care 2018, 41, 917–928. [Google Scholar] [CrossRef] [Green Version]
- Balamurugan, A.N.; Naziruddin, B.; Lockridge, A.; Tiwari, M.; Loganathan, G.; Takita, M.; Matsumoto, S.; Papas, K.; Trieger, M.; Rainis, H.; et al. Islet product characteristics and factors related to successful human islet transplantation from the Collaborative Islet Transplant Registry (CITR) 1999-2010. Am. J. Transplant. 2014, 14, 2595–2606. [Google Scholar] [CrossRef] [PubMed]
- Bottino, R.; Knoll, M.F.; Knoll, C.A.; Bertera, S.; Trucco, M.M. The Future of Islet Transplantation Is Now. Front. Med. 2018, 5, 202. [Google Scholar] [CrossRef] [Green Version]
- Welsch, C.A.; Rust, W.L.; Csete, M. Concise Review: Lessons Learned from Islet Transplant Clinical Trials in Developing Stem Cell Therapies for Type 1 Diabetes. Stem Cells Transl. Med. 2019, 8, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, A.M.; Pokrywczynska, M.; Ricordi, C. Clinical pancreatic islet transplantation. Nat. Rev. Endocrinol. 2017, 13, 268–277. [Google Scholar] [CrossRef]
- Kroon, E.; Martinson, L.A.; Kadoya, K.; Bang, A.G.; Kelly, O.G.; Eliazer, S.; Young, H.; Richardson, M.; Smart, N.G.; Cunningham, J.; et al. Pancreatic endoderm derived from human embryonic stem cells generates glucose-responsive insulin-secreting cells in vivo. Nat. Biotechnol. 2008, 26, 443–452. [Google Scholar] [CrossRef]
- Wallner, K.; Shapiro, A.M.; Senior, P.A.; McCabe, C. Cost effectiveness and value of information analyses of islet cell transplantation in the management of ‘unstable’ type 1 diabetes mellitus. BMC Endocr. Disord. 2016, 16, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieffer, T.J.; Woltjen, K.; Osafune, K.; Yabe, D.; Inagaki, N. Beta-cell replacement strategies for diabetes. J. Diabetes Investig. 2017, 9, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Bandeiras, C.; Cabral, J.M.S.; Gabbay, R.A.; Finkelstein, S.N.; Ferreira, F.C. Bringing Stem Cell-Based Therapies for Type 1 Diabetes to the Clinic: Early Insights from Bioprocess Economics and Cost-Effectiveness Analysis. Biotechnol. J. 2019, 14, e1800563. [Google Scholar] [CrossRef] [PubMed]
- Moassesfar, S.; Masharani, U.; Frassetto, L.A.; Szot, G.L.; Tavakol, M.; Stock, P.G.; Posselt, A.M. A Comparative Analysis of the Safety, Efficacy, and Cost of Islet Versus Pancreas Transplantation in Nonuremic Patients With Type 1 Diabetes. Am. J. Transplant. 2016, 16, 518–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Lin, P.J.; Cohen, J.T.; Winn, A.N.; Neumann, P.J. Cost-utility analyses in diabetes: A systematic review and implications from real-world evidence. Value Health 2015, 18, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.; Zacharias, R.L. Chapter 75-US Stem Cell Research Policy. In Principles of Regenerative Medicine, 3rd ed.; Atala, A., Lanza, R., Mikos, A.G., Nerem, R., Eds.; Academic Press: Boston, MA, USA, 2019; pp. 1309–1329. [Google Scholar] [CrossRef]
- Koike, H.; Iwasawa, K.; Ouchi, R.; Maezawa, M.; Giesbrecht, K.; Saiki, N.; Ferguson, A.; Kimura, M.; Thompson, W.L.; Wells, J.M.; et al. Modelling human hepato-biliary-pancreatic organogenesis from the foregut-midgut boundary. Nature 2019, 574, 112–116. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| SC-Islet Cells Transplantation | Automated Insulin Delivery Systems | |
|---|---|---|
| Advantages | Free from insulin injection | No immunosuppression |
| Improved time-in-range | Improved time-in-range | |
| Disadvantages | Risk of immunosuppression | Complicated maintenance |
| Risk of cancerization | Adjustment of dose according to diet | |
| Risk of insulin insufficiency | Risk of hypoglycemia, DKA or HHS | |
| Risk of re-transplantation | Local skin troubles |
| Source/Country | Study Population | Time Horizon | Intervention and Comparator | Per Additional QALY | Results |
|---|---|---|---|---|---|
| Wallner et al., 2018 (Canada) | Average age of 47 years, adults with hypoglycemia unawareness | 62.5 years | Intervention: Cadaveric islet cell transplantation Comparator: Intensive insulin therapy | $150,006 | Islet cell transplantation had a probability of being cost-effective of 0.5% at a willingness-to-pay (WTP) threshold of $100,000 per QALY. |
| Banderias et al., 2019 (USA) | Age of 18–35 years, hypoglycemia unawareness | 20 years | Intervention: Stem cell-derived islet cell transplantation Comparator: Intensive insulin therapy | $249,740 | Islet cell transplantation was cost-effective for 3.4% of patients when a WTP of up to $150,000 per QALY was considered. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, R.; Nishiyama, K.; Li, J.; Miyashita, D.; Ono, M.; Terauchi, Y.; Shirakawa, J. The Feasibility and Applicability of Stem Cell Therapy for the Cure of Type 1 Diabetes. Cells 2021, 10, 1589. https://doi.org/10.3390/cells10071589
Inoue R, Nishiyama K, Li J, Miyashita D, Ono M, Terauchi Y, Shirakawa J. The Feasibility and Applicability of Stem Cell Therapy for the Cure of Type 1 Diabetes. Cells. 2021; 10(7):1589. https://doi.org/10.3390/cells10071589
Chicago/Turabian StyleInoue, Ryota, Kuniyuki Nishiyama, Jinghe Li, Daisuke Miyashita, Masato Ono, Yasuo Terauchi, and Jun Shirakawa. 2021. "The Feasibility and Applicability of Stem Cell Therapy for the Cure of Type 1 Diabetes" Cells 10, no. 7: 1589. https://doi.org/10.3390/cells10071589
APA StyleInoue, R., Nishiyama, K., Li, J., Miyashita, D., Ono, M., Terauchi, Y., & Shirakawa, J. (2021). The Feasibility and Applicability of Stem Cell Therapy for the Cure of Type 1 Diabetes. Cells, 10(7), 1589. https://doi.org/10.3390/cells10071589