
Cellular Mechanisms of FGF-Stimulated Tissue Repair


Abstract
:1. Introduction
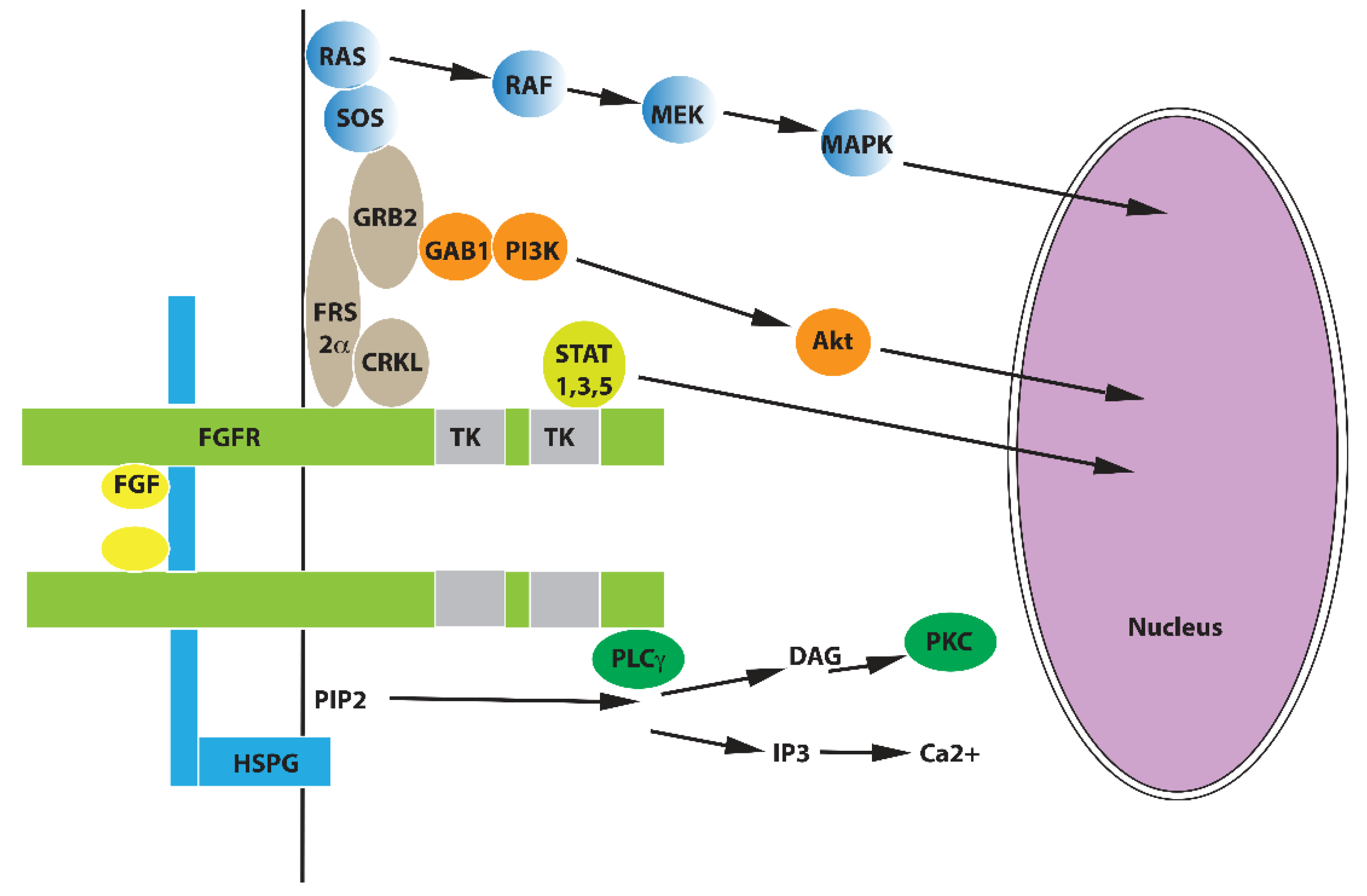
2. FGF Family and FGF Signaling
3. FGF as Stimulators of Regeneration and Repair
3.1. FGF and Regeneration in Invertebrates
3.2. FGF and Regeneration in Lower Vertebrates
3.3. FGF and Tissue Repair in Higher Vertebrates
3.4. Use of Recombinant FGFs for Tissue Repair
3.5. Potential Participation of Intracellular and Hormone-Like FGFs in Repair Processes
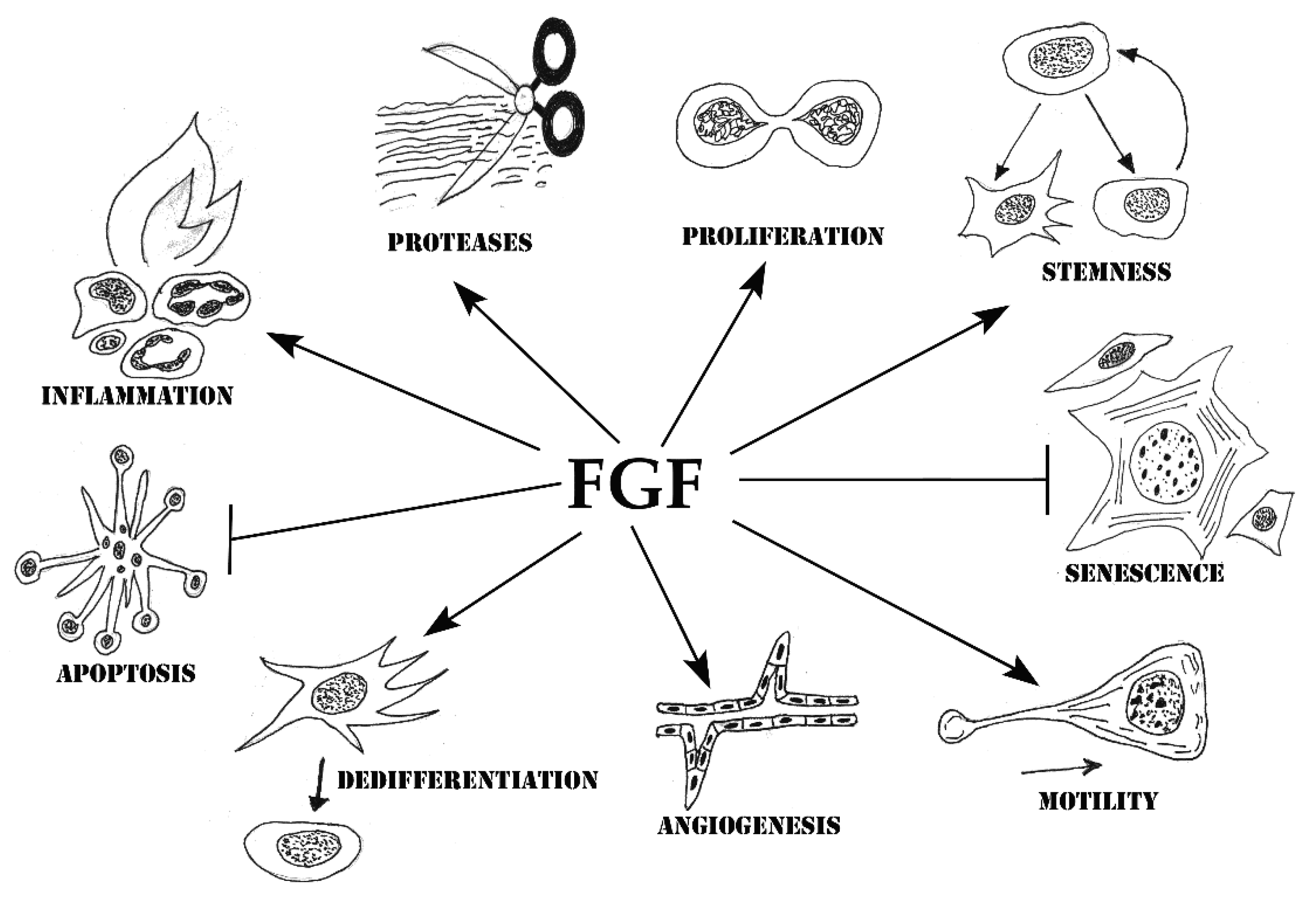
4. Cell Processes Underlying the Stimulation of Tissue Repair by FGF
4.1. Maintenance of Cell Stemness
4.2. Induction of Cell De-Differentiation
4.3. Proliferative Stimulation and Its Limitation
4.4. Suppression of Cell Senescence
4.5. Suppression of Cell Death
4.6. Regulation of Inflammation
4.7. Stimulation of Angiogenesis
4.8. Enhancement of Proteases Expression
5. Tissue Stress and Stimulation of FGF Expression and Release
5.1. FGF Expression
5.2. Release of Signal Peptide-Less FGFs
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ECM | extracellular matrix |
| FGF | fibroblast growth factor |
| FGFR | FGF receptor |
| HSPG | heparan sulfate proteoglycan |
| MMP | matrix metalloproteinase |
| VSMC | vascular smooth muscle cell |
References
- Schein, C.H. The Shape of the Messenger: Using Protein Structure Information to Design Novel Cytokine-based Therapeutics. Curr. Pharm. Des. 2002, 8, 2113–2129. [Google Scholar] [CrossRef]
- Wei, E.Q.; Barnett, A.S.; Pitt, G.S.; Hennessey, J.A. Fibroblast Growth Factor Homologous Factors in the Heart: A Potential Locus for Cardiac Arrhythmias. Trends Cardiovasc. Med. 2011, 21, 199–203. [Google Scholar] [CrossRef] [Green Version]
- Degirolamo, C.; Sabbà, C.; Moschetta, A. Therapeutic potential of the endocrine fibroblast growth factors FGF19, FGF21 and FGF23. Nat. Rev. Drug Discov. 2016, 15, 51–69. [Google Scholar] [CrossRef]
- Friesel, R.; Maciag, T. Fibroblast growth factor prototype release and fibroblast growth factor receptor signaling. Thromb. Haemost. 1999, 82, 748–754. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef] [Green Version]
- Kuro, O.M. The Klotho proteins in health and disease. Nat. Rev. Nephrol. 2019, 15, 27–44. [Google Scholar] [CrossRef]
- Klagsbrun, M. Mediators of Angiogenesis: The Biological Significance of Basic Fibroblast Growth Factor (Bfgf)-Heparin and Heparan Sulfate Interactions. Semin. Cancer Biol. 1992, 3, 81–87. [Google Scholar] [PubMed]
- Pande, S.; Yang, X.; Friesel, R. Interleukin-17 receptor D (Sef) is a multi-functional regulator of cell signaling. Cell Commun. Signal. 2021, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lange, E.; Bertrand, S.; Holz, O.; Rebscher, N.; Hassel, M. Dynamic expression of a Hydra FGF at boundaries and termini. Dev. Genes Evol. 2014, 224, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Turwankar, A.; Ghaskadbi, S. VEGF and FGF signaling during head regeneration in hydra. Gene 2019, 717, 144047. [Google Scholar] [CrossRef] [PubMed]
- Auwal, M.A.; Kashima, M.; Nishimura, O.; Hosoda, K.; Motoishi, M.; Kamimura, A.; Okumura, A.; Agata, K.; Umesono, Y. Identification and characterization of a fibroblast growth factor gene in the planarian Dugesia japonica. Dev. Growth Differ. 2020, 62, 527–539. [Google Scholar] [CrossRef]
- Cancre, I.; Van Wormhoudt, A.; le Gal, Y. Heparin-binding molecules with growth factor activities in regenerating-tissues of the starfish Asterias rubens. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1999, 123, 285–292. [Google Scholar] [CrossRef]
- Czarkwiani, A.; Dylus, D.V.; Carballo, L.; Oliveri, P. FGF signalling plays similar roles in development and regeneration of the skeleton in the brittle star Amphiura filiformis. Development 2021, 148. [Google Scholar] [CrossRef]
- Poss, K.D.; Shen, J.; Nechiporuk, A.; McMahon, G.; Thisse, B.; Thisse, C.; Keating, M.T. Roles for Fgf Signaling During Zebrafish Fin Regeneration. Dev. Biol. 2000, 222, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duszynski, R.J.; Topczewski, J.; LeClair, E.E. Divergent requirements for fibroblast growth factor signaling in zebrafish maxillary barbel and caudal fin regeneration. Dev. Growth Differ. 2013, 55, 282–300. [Google Scholar] [CrossRef] [Green Version]
- Whitehead, G.G.; Makino, S.; Lien, C.-L.; Keating, M.T. fgf20 Is Essential for Initiating Zebrafish Fin Regeneration. Science 2005, 310, 1957–1960. [Google Scholar] [CrossRef] [Green Version]
- Goldshmit, Y.; Sztal, T.E.; Jusuf, P.R.; Hall, T.E.; Nguyen-Chi, M.; Currie, P.D. Fgf-Dependent Glial Cell Bridges Facilitate Spinal Cord Regeneration in Zebrafish. J. Neurosci. 2012, 32, 7477–7492. [Google Scholar] [CrossRef] [Green Version]
- Kan, N.G.; Junghans, D.; Belmonte, J.C.I. Compensatory growth mechanisms regulated by BMP and FGF signaling mediate liver regeneration in zebrafish after partial hepatectomy. FASEB J. 2009, 23, 3516–3525. [Google Scholar] [CrossRef] [Green Version]
- Lepilina, A.; Coon, A.N.; Kikuchi, K.; Holdway, J.E.; Roberts, R.W.; Burns, C.G.; Poss, K.D. A Dynamic Epicardial Injury Response Supports Progenitor Cell Activity during Zebrafish Heart Regeneration. Cell 2006, 127, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.G.; Huang, M.; Obholzer, N.D.; Sun, S.; Li, W.; Petrillo, M.; Dai, P.; Zhou, Y.; Cotanche, U.A.; Megason, S.G.; et al. Myc and Fgf Are Required for Zebrafish Neuromast Hair Cell Regeneration. PLoS ONE 2016, 11, e0157768. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Kidd, A.R., 3rd; Thomas, J.L.; Poss, K.D.; Hyde, D.R.; Raymond, P.A.; Thummel, R. Fgf Signaling Regulates Rod Photoreceptor Cell Maintenance and Regeneration in Zebrafish. Exp. Eye Res. 2011, 93, 726–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saera-Vila, A.; Kish, P.E.; Kahana, A. Fgf regulates dedifferentiation during skeletal muscle regeneration in adult zebrafish. Cell. Signal. 2016, 28, 1196–1204. [Google Scholar] [CrossRef] [Green Version]
- Christen, B.; Slack, J.M. FGF-8Is Associated with Anteroposterior Patterning and Limb Regeneration inXenopus. Dev. Biol. 1997, 192, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Han, M.-J.; An, J.-Y.; Kim, W.-S. Expression patterns ofFgf-8 during development and limb regeneration of the axolotl. Dev. Dyn. 2001, 220, 40–48. [Google Scholar] [CrossRef]
- Lin, G.; Slack, J.M. Requirement for Wnt and FGF signaling in Xenopus tadpole tail regeneration. Dev. Biol. 2008, 316, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Makanae, A.; Mitogawa, K.; Satoh, A. Co-operative Bmp- and Fgf-signaling inputs convert skin wound healing to limb formation in urodele amphibians. Dev. Biol. 2014, 396, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Mullen, L.M.; Bryant, S.V.; Torok, M.A.; Blumberg, B.; Gardiner, D.M. Nerve dependency of regeneration: The role of Distal-less and FGF signaling in amphibian limb regeneration. Development 1996, 122, 3487–3497. [Google Scholar] [CrossRef]
- Satoh, A.; Makanae, A.; Nishimoto, Y.; Mitogawa, K. FGF and BMP derived from dorsal root ganglia regulate blastema induction in limb regeneration in Ambystoma mexicanum. Dev. Biol. 2016, 417, 114–125. [Google Scholar] [CrossRef]
- Choi, Y.; Meng, F.; Cox, C.S.; Lally, K.P.; Huard, J.; Li, Y. Regeneration and Regrowth Potentials of Digit Tips in Amphibians and Mammals. Int. J. Cell Biol. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mao, S.A.; Glorioso, J.M.; Nyberg, S.L. Liver regeneration. Transl. Res. 2014, 163, 352–362. [Google Scholar] [CrossRef] [Green Version]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6, 265sr6. [Google Scholar] [CrossRef] [Green Version]
- Maddaluno, L.; Urwyler, C.; Werner, S. Fibroblast growth factors: Key players in regeneration and tissue repair. Development 2017, 144, 4047–4060. [Google Scholar] [CrossRef] [Green Version]
- Ortega, S.; Ittmann, M.; Tsang, S.H.; Ehrlich, M.; Basilico, C. Neuronal defects and delayed wound healing in mice lacking fibroblast growth factor 2. Proc. Natl. Acad. Sci. USA 1998, 95, 5672–5677. [Google Scholar] [CrossRef] [Green Version]
- Meyer, M.; Müller, A.-K.; Yang, J.; Moik, D.; Ponzio, G.; Ornitz, D.; Grose, R.; Werner, S. FGF receptors 1 and 2 are key regulators of keratinocyte migration in vitro and in wounded skin. J. Cell Sci. 2012, 125, 5690–5701. [Google Scholar] [CrossRef] [Green Version]
- Furusho, M.; Roulois, A.J.; Franklin, R.J.M.; Bansal, R. Fibroblast growth factor signaling in oligodendrocyte-lineage cells facilitates recovery of chronically demyelinated lesions but is redundant in acute lesions. Glia 2015, 63, 1714–1728. [Google Scholar] [CrossRef] [Green Version]
- Narla, S.T.; Bushnell, D.S.; Schaefer, C.M.; Nouraie, M.; Tometich, J.T.; Hand, Y.W.; Bates, C.M. Loss of Fibroblast Growth Factor Receptor 2 (Fgfr2) Leads to Defective Bladder Urothelial Regeneration after Cyclophosphamide Injury. Am. J. Pathol. 2021, 191, 631–651. [Google Scholar] [CrossRef]
- Guzy, R.D.; Stoilov, I.; Elton, T.J.; Mecham, R.P.; Ornitz, D.M. Fibroblast Growth Factor 2 Is Required for Epithelial Recovery, but Not for Pulmonary Fibrosis, in Response to Bleomycin. Am. J. Respir. Cell Mol. Biol. 2015, 52, 116–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, H.Y.; El-Baz, L.M.; House, S.L.; Cilvik, S.; Dorry, S.J.; Shoukry, N.M.; Salem, M.L.; Hafez, H.S.; Dulin, N.O.; Ornitz, D.M.; et al. Fibroblast growth factor 2 decreases bleomycin-induced pulmonary fibrosis and inhibits fibroblast collagen production and myofibroblast differentiation. J. Pathol. 2018, 246, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xu, B.; Wang, Y.; Li, Y.; Si, H.; Zheng, X.; Chen, Z.; Chen, F.; Fan, D. Dramatic promotion of wound healing using a recombinant human-like collagen and bFGF cross-linked hydrogel by transglutaminase. J. Biomater. Sci. Polym. Ed. 2019, 30, 1591–1603. [Google Scholar] [CrossRef] [PubMed]
- Zubair, M.; Ahmad, J. Role of growth factors and cytokines in diabetic foot ulcer healing: A detailed review. Rev. Endocr. Metab. Disord. 2019, 20, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Haastert, K.; Ying, Z.; Grothe, C.; Gomez-Pinilla, F. The effects of FGF-2 gene therapy combined with voluntary exercise on axonal regeneration across peripheral nerve gaps. Neurosci. Lett. 2008, 443, 179–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-J.; Jingushi, S.; Aoyama, I.; Anzai, J.; Hirata, G.; Tamura, M.; Iwamoto, Y. Effects of FGF-2 on metaphyseal fracture repair in rabbit tibiae. J. Bone Miner. Metab. 2004, 22, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewska, M.; Marcinkowska, E.; Wiedlocha, A. FGF-1: From Biology Through Engineering to Potential Medical Applications. Crit. Rev. Clin. Lab. Sci. 2008, 45, 91–135. [Google Scholar] [CrossRef] [PubMed]
- Braund, R.; Hook, S.M.; Greenhill, N.; Medlicott, N.J. Distribution of Fibroblast Growth Factor-2 (Fgf-2) within Model Excisional Wounds Following Topical Application. J. Pharm. Pharmacol. 2009, 61, 193–200. [Google Scholar] [CrossRef]
- Hui, Q.; Jin, Z.; Li, X.; Liu, C.; Wang, X. FGF Family: From Drug Development to Clinical Application. Int. J. Mol. Sci. 2018, 19, 1875. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, Y.; Kawai, T.; Goto, K.; Matsuda, S. Clinical application of injectable growth factor for bone regeneration: A systematic review. Inflamm. Regen. 2019, 39, 1–10. [Google Scholar] [CrossRef]
- Nunes, Q.; Li, Y.; Sun, C.; Kinnunen, T.K.; Fernig, D.G. Fibroblast growth factors as tissue repair and regeneration therapeutics. PeerJ 2016, 4, e1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Kim, W.J.; Jun, H.O.; Lee, E.J.; Lee, K.W.; Jeong, J.-Y.; Lee, S.-W. Hypoxia-induced fibroblast growth factor 11 stimulates capillary-like endothelial tube formation. Oncol. Rep. 2015, 34, 2745–2751. [Google Scholar] [CrossRef] [Green Version]
- Knowles, H.J. Hypoxia-Induced Fibroblast Growth Factor 11 Stimulates Osteoclast-Mediated Resorption of Bone. Calcif. Tissue Int. 2017, 100, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Yim, H.; Shin, J.; Lee, C.; Lee, J.-H.; Jeong, J. FGF11 induced by hypoxia interacts with HIF-1α and enhances its stability. FEBS Lett. 2017, 591, 348–357. [Google Scholar] [CrossRef]
- Yamauchi, M.; Hirohashi, Y.; Torigoe, T.; Matsumoto, Y.; Yamashita, K.; Kayama, M.; Sato, N.; Yotsuyanagi, T. Wound Healing Delays in Alpha-Klotho-Deficient Mice That Have Skin Appearance Similar to That in Aged Humans—Study of Delayed Wound Healing Mechanism. Biochem. Biophys. Res. Commun. 2016, 473, 845–852. [Google Scholar] [CrossRef]
- Kang, K.; Xu, P.; Wang, M.; Chunyu, J.; Sun, X.; Ren, G.; Xiao, W.; Li, D. FGF21 attenuates neurodegeneration through modulating neuroinflammation and oxidant-stress. Biomed. Pharmacother. 2020, 129, 110439. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Ying, Y.; Ye, L.; Ying, W.; Ye, J.; Wu, Q.; Chen, M.; Zhu, H.; Li, X.; Dou, H.; et al. Systemic Administration of Fibroblast Growth Factor 21 Improves the Recovery of Spinal Cord Injury (SCI) in Rats and Attenuates SCI-Induced Autophagy. Front. Pharmacol. 2021, 11, 628369. [Google Scholar] [CrossRef]
- Kong, B.; Huang, J.; Zhu, Y.; Li, G.; Williams, J.; Shen, S.; Aleksunes, L.M.; Richardson, J.R.; Apte, U.; Rudnick, D.A.; et al. Fibroblast growth factor 15 deficiency impairs liver regeneration in mice. Am. J. Physiol. Liver Physiol. 2014, 306, G893–G902. [Google Scholar] [CrossRef] [Green Version]
- Trivier, E.; Kurz, D.J.; Hong, Y.; Huang, H.-L.; Erusalimsky, J.D. Differential Regulation of Telomerase in Endothelial Cells by Fibroblast Growth Factor-2 and Vascular Endothelial Growth Factor-A: Association with Replicative Life Span. Ann. N. Y. Acad. Sci. 2004, 1019, 111–115. [Google Scholar] [CrossRef]
- Rieck, P.W.; von Stockhausen, R.M.; Metzner, S.; Hartmann, C.; Courtois, Y. Fibroblast Growth Factor-2 Protects Endothelial Cells from Damage after Corneal Storage at 4 Degrees C. Graefes. Arch. Clin. Exp. Ophthalmol. 2003, 241, 757–764. [Google Scholar] [CrossRef]
- Partridge, C.R.; Hawker, J.R., Jr.; Forough, R. Overexpression of a Secretory Form of Fgf-1 Promotes Mmp-1-Mediated Endothelial Cell Migration. J. Cell Biochem. 2000, 78, 487–499. [Google Scholar] [CrossRef]
- Kim, B.S.; Park, J.-Y.; Kang, H.-J.; Kim, H.-J.; Lee, J. Fucoidan/FGF-2 induces angiogenesis through JNK- and p38-mediated activation of AKT/MMP-2 signalling. Biochem. Biophys. Res. Commun. 2014, 450, 1333–1338. [Google Scholar] [CrossRef]
- Rusnati, M.; Dell’Era, P.; Urbinati, C.; Tanghetti, E.; Massardi, M.L.; Nagamine, Y.; Monti, E.; Presta, M. A Distinct Basic Fibroblast Growth Factor (Fgf-2)/Fgf Receptor Interaction Distinguishes Urokinase-Type Plasminogen Activator Induction from Mitogenicity in Endothelial Cells. Mol. Biol. Cell 1996, 7, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Behr, B.; Leucht, P.; Longaker, M.T.; Quarto, N. Fgf-9 is required for angiogenesis and osteogenesis in long bone repair. Proc. Natl. Acad. Sci. USA 2010, 107, 11853–11858. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, B.; Von Specht, B.U.; Haberstroh, J.; Pecher, P. The stimulation of neo-angiogenesis in the ischemic heart by the human growth factor FGF. J. Cardiovasc. Surg. 1998, 39, 445–453. [Google Scholar]
- Ishihara, M.; Obara, K.; Nakamura, S.; Fujita, M.; Masuoka, K.; Kanatani, Y.; Takase, B.; Hattori, H.; Morimoto, Y.; Ishihara, M.; et al. Chitosan Hydrogel as a Drug Delivery Carrier to Control Angiogenesis. J. Artif. Organs 2006, 9, 8–16. [Google Scholar] [CrossRef]
- Fujita, M.; Ishihara, M.; Shimizu, M.; Obara, K.; Nakamura, S.; Kanatani, Y.; Morimoto, Y.; Takase, B.; Matsui, T.; Kikuchi, M.; et al. Therapeutic angiogenesis induced by controlled release of fibroblast growth factor-2 from injectable chitosan/non-anticoagulant heparin hydrogel in a rat hindlimb ischemia model. Wound Repair Regen. 2007, 15, 58–65. [Google Scholar] [CrossRef]
- Li, J.; Wei, Y.; Liu, K.; Yuan, C.; Tang, Y.; Quan, Q.; Chen, P.; Wang, W.; Hu, H.; Yang, L. Synergistic effects of FGF-2 and PDGF-BB on angiogenesis and muscle regeneration in rabbit hindlimb ischemia model. Microvasc. Res. 2010, 80, 10–17. [Google Scholar] [CrossRef]
- Takikawa, M.; Nakamura, S.; Ishihara, M.; Takabayashi, Y.; Fujita, M.; Hattori, H.; Kushibiki, T.; Ishihara, M. Improved angiogenesis and healing in crush syndrome by fibroblast growth factor-2–containing low-molecular-weight heparin (Fragmin)/protamine nanoparticles. J. Surg. Res. 2015, 196, 247–257. [Google Scholar] [CrossRef]
- Qu, D.; Li, J.; Li, Y.; Gao, Y.; Zuo, Y.; Hsu, Y.; Hu, J. Angiogenesis and osteogenesis enhanced by bFGF ex vivo gene therapy for bone tissue engineering in reconstruction of calvarial defects. J. Biomed. Mater. Res. Part A 2011, 96, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Kelpke, S.S.; Zinn, K.R.; Rue, L.W.; Thompson, J.A. Site-Specific Delivery of Acidic Fibroblast Growth Factor Stimulates Angiogenic and Osteogenic Responses in vivo. J. Biomed. Mater. Res. A 2004, 71, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Pandit, A.S.; Feldman, D.S.; Caulfield, J.; Thompson, A. Stimulation of Angiogenesis by Fgf-1 Delivered through a Modified Fibrin Scaffold. Growth Factors 1998, 15, 113–123. [Google Scholar] [CrossRef]
- Iwakura, A.; Tabata, Y.; Tamura, N.; Doi, K.; Nishimura, K.; Nakamura, T.; Shimizu, Y.; Fujita, M.; Komeda, M. Gelatin Sheet Incorporating Basic Fibroblast Growth Factor Enhances Healing of Devascularized Sternum in Diabetic Rats. Circulation 2001, 104, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Xin, S. FGF signaling contributes to atherosclerosis by enhancing the inflammatory response in vascular smooth muscle cells. Mol. Med. Rep. 2019, 20, 162–170. [Google Scholar] [CrossRef]
- Chen, P.Y.; Qin, L.; Li, G.; Tellides, G.; Simons, M. Fibroblast Growth Factor (Fgf) Signaling Regulates Transforming Growth Factor Beta (Tgfbeta)-Dependent Smooth Muscle Cell Phenotype Modulation. Sci. Rep. 2016, 6, 33407. [Google Scholar] [CrossRef] [Green Version]
- Dolivo, D.; Larson, S.; Dominko, T. Fibroblast Growth Factor 2 as an Antifibrotic: Antagonism of Myofibroblast Differentiation and Suppression of Pro-Fibrotic Gene Expression. Cytokine Growth Factor Rev. 2017, 38, 49–58. [Google Scholar] [CrossRef]
- Page, R.L.; Ambady, S.; Holmes, W.F.; Vilner, L.; Kole, D.; Kashpur, O.; Huntress, V.; Vojtic, I.; Whitton, H.; Dominko, T. Induction of Stem Cell Gene Expression in Adult Human Fibroblasts without Transgenes. Cloning Stem Cells 2009, 11, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Kashpur, O.; Lapointe, D.; Ambady, S.; Ryder, E.F.; Dominko, T. FGF2-induced effects on transcriptome associated with regeneration competence in adult human fibroblasts. BMC Genom. 2013, 14, 656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglia, C.; Berlingieri, M.T.; Martelli, M.L.; Trapasso, F.; Bovi, P.D.; Fusco, A. Mitogenic and dedifferentiating effect of the K-fgf/hst oncogene on rat thyroid PC clone 3 epithelial cells. Cell Growth Differ. Mol. Boil. J. Am. Assoc. Cancer Res. 1993, 4, 185–192. [Google Scholar]
- Lungu, G.; Covaleda, L.; Mendes, O.; Martini-Stoica, H.; Stoica, G. Fgf-1-Induced Matrix Metalloproteinase-9 Expression in Breast Cancer Cells Is Mediated by Increased Activities of Nf-Kappab and Activating Protein-1. Mol. Carcinog. 2008, 47, 424–435. [Google Scholar] [CrossRef]
- Koike, Y.; Yozaki, M.; Utani, A.; Murota, H. Fibroblast growth factor 2 accelerates the epithelial–mesenchymal transition in keratinocytes during wound healing process. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pu, J.; Wang, R.; Zhang, G.; Wang, J. Fgf-7 Facilitates the Process of Psoriasis by Inducing Tnf-Alpha Expression in Hacat Cells. Acta. Biochim. Biophys. Sin. 2019, 51, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chen, X.; Han, C.; Wang, Y.; Huang, T.; Du, Y.; Dong, Z. FGF-2 Transcriptionally Down-Regulates the Expression of BNIP3L via PI3K/Akt/FoxO3a Signaling and Inhibits Necrosis and Mitochondrial Dysfunction Induced by High Concentrations of Hydrogen Peroxide in H9c2 Cells. Cell. Physiol. Biochem. 2016, 40, 1678–1691. [Google Scholar] [CrossRef] [PubMed]
- Koleini, N.; Nickel, B.E.; Wang, J.; Roveimiab, Z.; Fandrich, R.R.; Kirshenbaum, L.A.; Cattini, P.A.; Kardami, E. Fibroblast growth factor-2-mediated protection of cardiomyocytes from the toxic effects of doxorubicin requires the mTOR/Nrf-2/HO-1 pathway. Oncotarget 2017, 8, 87415–87430. [Google Scholar] [CrossRef] [Green Version]
- Martin, I.; Novakovica, G.; Yang, J.; Langer, R.; Freed, L. Mammalian Chondrocytes Expanded in the Presence of Fibroblast Growth Factor 2 Maintain the Ability to Differentiate and Regenerate Three-Dimensional Cartilaginous Tissue. Exp. Cell Res. 1999, 253, 681–688. [Google Scholar] [CrossRef]
- Im, H.-J.; Li, X.; Muddasani, P.; Kim, G.-H.; Davis, F.; Rangan, J.; Forsyth, C.B.; Ellman, M.; Thonar, E.J. Basic fibroblast growth factor accelerates matrix degradation via a neuro-endocrine pathway in human adult articular chondrocytes. J. Cell. Physiol. 2008, 215, 452–463. [Google Scholar] [CrossRef] [Green Version]
- Chikazu, D.; Hakeda, Y.; Ogata, N.; Nemoto, K.; Itabashi, A.; Takato, T.; Kumegawa, M.; Nakamura, K.; Kawaguchi, H. Fibroblast Growth Factor (FGF)-2 Directly Stimulates Mature Osteoclast Function through Activation of FGF Receptor 1 and p42/p44 MAP Kinase. J. Biol. Chem. 2000, 275, 31444–31450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.C.; Szuflita, N.; Khatri, R.; Laterra, J.; Hossain, M.A. Transgenic expression of human FGF-1 protects against hypoxic–ischemic injury in perinatal brain by intervening at caspase-XIAP signaling cascades. Neurobiol. Dis. 2006, 22, 677–690. [Google Scholar] [CrossRef]
- Okada, T.; Enkhjargal, B.; Travis, Z.D.; Ocak, U.; Tang, J.; Suzuki, H.; Zhang, J.H. Fgf-2 Attenuates Neuronal Apoptosis Via Fgfr3/Pi3k/Akt Signaling Pathway after Subarachnoid Hemorrhage. Mol. Neurobiol. 2019, 56, 8203–8219. [Google Scholar] [CrossRef]
- Yasuda, T.; Grinspan, J.; Stern, J.; Franceschini, B.; Bannerman, P.; Pleasure, D. Apoptosis occurs in the oligodendroglial lineage, and is prevented by basic fibroblast growth factor. J. Neurosci. Res. 1995, 40, 306–317. [Google Scholar] [CrossRef]
- Song, H.; Kwon, K.; Lim, S.; Kang, S.-M.; Ko, Y.-G.; Xu, Z.; Chung, J.H.; Kim, B.-S.; Lee, H.; Joung, B.; et al. Transfection of mesenchymal stem cells with the FGF-2 gene improves their survival under hypoxic conditions. Mol. Cells 2005, 19, 402–407. [Google Scholar]
- Ito, T.; Sawada, R.; Fujiwara, Y.; Seyama, Y.; Tsuchiya, T. Fgf-2 Suppresses Cellular Senescence of Human Mesenchymal Stem Cells by Down-Regulation of Tgf-Beta2. Biochem. Biophys. Res. Commun. 2007, 359, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Hirai, K.; Sasaki, H.; Yamamoto, H.; Sakamoto, H.; Kubota, Y.; Kakizoe, T.; Terada, M.; Ochiya, T. HST-1/FGF-4 protects male germ cells from apoptosis under heat-stress condition. Exp. Cell Res. 2004, 294, 77–85. [Google Scholar] [CrossRef]
- Hsieh, Y.-C.; Intawicha, P.; Lee, K.-H.; Chiu, Y.-T.; Lo, N.-W.; Ju, J.-C. LIF and FGF Cooperatively Support Stemness of Rabbit Embryonic Stem Cells Derived from Parthenogenetically Activated Embryos. Cell. Reprogramming 2011, 13, 241–255. [Google Scholar] [CrossRef]
- Lee, E.J.; Kang, H.-J.; Lee, H.N.; Kang, S.K.; Kim, K.-H.; Lee, S.-W.; Lee, G.; Park, Y.-B.; Kim, H.-S. New culture system for human embryonic stem cells: Autologous mesenchymal stem cell feeder without exogenous fibroblast growth factor 2. Differentiation 2012, 83, 92–100. [Google Scholar] [CrossRef]
- Galderisi, U.; Peluso, G.; Di Bernardo, G.; Calarco, A.; D’Apolito, M.; Petillo, O.; Cipollaro, M.; Fusco, F.; Melone, M.A.B. Efficient cultivation of neural stem cells with controlled delivery of FGF-2. Stem Cell Res. 2013, 10, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Latos, P.A.; Goncalves, A.; Oxley, D.; Mohammed, H.; Turro, E.; Hemberger, M. Fgf and Esrrb integrate epigenetic and transcriptional networks that regulate self-renewal of trophoblast stem cells. Nat. Commun. 2015, 6, 7776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Guo, H.; Yang, C.; Chen, Q.; Huang, J.; Liu, L.; Zhang, Y.; Jin, S.; Song, A.; Yang, P. The biological behavior optimization of human periodontal ligament stem cells via preconditioning by the combined application of fibroblast growth factor-2 and A83-01 in in vitro culture expansion. J. Transl. Med. 2019, 17, 66. [Google Scholar] [CrossRef] [PubMed]
- Eom, Y.W.; Oh, J.-E.; Lee, J.I.; Baik, S.K.; Rhee, K.-J.; Shin, H.C.; Kim, Y.M.; Ahn, C.M.; Kong, J.H.; Kim, H.S.; et al. The role of growth factors in maintenance of stemness in bone marrow-derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2014, 445, 16–22. [Google Scholar] [CrossRef]
- Pawlikowski, B.; Vogler, T.O.; Gadek, K.; Olwin, B.B. Regulation of skeletal muscle stem cells by fibroblast growth factors. Dev. Dyn. 2017, 246, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, S.-H.; Ha, L.; Jang, H.-S. Nephron Progenitor Maintenance Is Controlled through Fibroblast Growth Factors and Sprouty1 Interaction. J. Am. Soc. Nephrol. 2020, 31, 2559–2572. [Google Scholar] [CrossRef]
- Huang, Y.; Hamana, T.; Liu, J.; Wang, C.; An, L.; You, P.; Chang, J.Y.F.; Xu, J.; Jin, C.; Zhang, Z.; et al. Type 2 Fibroblast Growth Factor Receptor Signaling Preserves Stemness and Prevents Differentiation of Prostate Stem Cells from the Basal Compartment. J. Biol. Chem. 2015, 290, 17753–17761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasegawa, K.; Saga, Y. FGF8-FGFR1 Signaling Acts as a Niche Factor for Maintaining Undifferentiated Spermatogonia in the Mouse1. Biol. Reprod. 2014, 91, 145. [Google Scholar] [CrossRef] [Green Version]
- Adachi, K.; Nikaido, I.; Ohta, H.; Ohtsuka, S.; Ura, H.; Kadota, M.; Wakayama, T.; Ueda, H.; Niwa, H. Context-Dependent Wiring of Sox2 Regulatory Networks for Self-Renewal of Embryonic and Trophoblast Stem Cells. Mol. Cell 2013, 52, 380–392. [Google Scholar] [CrossRef] [Green Version]
- Jaroonwitchawan, T.; Muangchan, P.; Noisa, P. nhibition of Fgf Signaling Accelerates Neural Crest Cell Differentiation of Human Pluripotent Stem Cells. Biochem. Biophys. Res. Commun. 2016, 481, 176–181. [Google Scholar] [CrossRef]
- Yu, P.; Pan, G.; Yu, J.; Thomson, J.A. FGF2 Sustains NANOG and Switches the Outcome of BMP4-Induced Human Embryonic Stem Cell Differentiation. Cell Stem Cell 2011, 8, 326–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang-Panesso, M.; Humphreys, B.D. Cellular plasticity in kidney injury and repair. Nat. Rev. Nephrol. 2016, 13, 39–46. [Google Scholar] [CrossRef]
- Kleiderman, S.; Gutbier, S.; Tufekci, K.U.; Ortega, F.; Sa, J.V.; Teixeira, A.P.; Brito, C.; Glaab, E.; Berninger, B.; Alves, P.M.; et al. Conversion of Nonproliferating Astrocytes into Neurogenic Neural Stem Cells: Control by Fgf2 and Interferon-Gamma. Stem Cells 2016, 34, 2861–2874. [Google Scholar] [CrossRef] [Green Version]
- Raju, R.; Palapetta, S.M.; Sandhya, V.K.; Sahu, A.; Alipoor, A.; Balakrishnan, L.; Advani, J.; George, B.; Kini, K.R.; Geetha, N.P.; et al. A Network Map of FGF-1/FGFR Signaling System. J. Signal Transduct. 2014, 2014, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Quinlan, D.C.; Hochstadt, J. The regulation by fibroblast growth factor of early transport changes in quiescent 3T3 cells. J. Cell. Physiol. 1977, 93, 237–246. [Google Scholar] [CrossRef]
- Poole, A.; Kacer, R.; Cooper, E.; Tarantini, F.; Prudovsky, I. Sustained Inhibition of Proliferative Response After Transient FGF Stimulation Is Mediated by Interleukin 1 Signaling. J. Cell. Physiol. 2015, 231, 650–658. [Google Scholar] [CrossRef] [Green Version]
- Orth, P.; Kaul, G.; Cucchiarini, M.; Zurakowski, D.; Menger, M.D.; Kohn, D.; Madry, H. Transplanted articular chondrocytes co-overexpressing IGF-I and FGF-2 stimulate cartilage repair in vivo. Knee Surg. Sports Traumatol. Arthrosc. 2011, 19, 2119–2130. [Google Scholar] [CrossRef]
- Madry, H.; Kohn, D.; Cucchiarini, M. Direct FGF-2 Gene Transfer via Recombinant Adeno-Associated Virus Vectors Stimulates Cell Proliferation, Collagen Production, and the Repair of Experimental Lesions in the Human ACL. Am. J. Sports Med. 2013, 41, 194–202. [Google Scholar] [CrossRef]
- Lavallee, T.M.; Prudovsky, I.A.; McMahon, G.A.; Hu, X.; Maciag, T. Activation of the MAP Kinase Pathway by FGF-1 Correlates with Cell Proliferation Induction While Activation of the Src Pathway Correlates with Migration. J. Cell Biol. 1998, 141, 1647–1658. [Google Scholar] [CrossRef]
- Borghesan, M.; Hoogaars, W.; Varela-Eirin, M.; Talma, N.; Demaria, M. A Senescence-Centric View of Aging: Implications for Longevity and Disease. Trends Cell Biol. 2020, 30, 777–791. [Google Scholar] [CrossRef]
- Irfan-Maqsood, M.; Matin, M.M.; Bahrami, A.R.; Ghasroldasht, M.M. Immortality of cell lines: Challenges and advantages of establishment. Cell Biol. Int. 2013, 37, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Garfinkel, S.; Hu, X.; A Prudovsky, I.; A McMahon, G.; Kapnik, E.M.; McDowell, S.D.; Maciag, T. FGF-1-dependent proliferative and migratory responses are impaired in senescent human umbilical vein endothelial cells and correlate with the inability to signal tyrosine phosphorylation of fibroblast growth factor receptor-1 substrates. J. Cell Biol. 1996, 134, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Segura, A.; Nehme, J.; Demaria, M. Hallmarks of Cellular Senescence. Trends Cell Biol. 2018, 28, 436–453. [Google Scholar] [CrossRef]
- Tchkonia, T.; Zhu, Y.; Van Deursen, J.; Campisi, J.; Kirkland, J.L. Cellular senescence and the senescent secretory phenotype: Therapeutic opportunities. J. Clin. Investig. 2013, 123, 966–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laberge, R.-M.; Sun, Y.; Orjalo, A.V.; Patil, C.K.; Freund, A.; Zhou, L.; Curran, S.C.; Davalos, A.R.; Wilson-Edell, K.A.; Liu, S.; et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat. Cell Biol. 2015, 17, 1049–1061. [Google Scholar] [CrossRef]
- Lee, N.J.; Wang, S.J.; KayDurairaj, K.; Srivatsan, E.S.; Wang, M.B. Increased Expression of Transforming Growth Factor-β1, Acidic Fibroblast Growth Factor, and Basic Fibroblast Growth Factor in Fetal Versus Adult Fibroblast Cell Lines. Laryngoscope 2000, 110, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Tong, H.J.; Li, M.; Tan, K.S.; Cao, T. Telomere length is regulated by FGF-2 in human embryonic stem cells and affects the life span of its differentiated progenies. Biogerontology 2016, 18, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.; Choi, J.; Nagy, A.; Sung, H.; Kim, J. Antisenescence effect of mouse embryonic stem cell conditioned medium through a PDGF/FGF pathway. FASEB J. 2016, 30, 1276–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, S.; Krampert, M.; Bodo, E.; Kumin, A.; Born-Berclaz, C.; Paus, R.; Werner, S. Keratinocyte Growth Factor Protects Epidermis and Hair Follicles from Cell Death Induced by Uv Irradiation, Chemotherapeutic or Cytotoxic Agents. J. Cell Sci. 2006, 119, 4841–4849. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Chen, H.; Han, C.; Fu, D.; Wang, F.; Wang, D.; Ma, L.; Zhou, L.; Han, D. The anti-apoptotic and prognostic value of fibroblast growth factor 9 in gastric cancer. Oncotarget 2016, 7, 36655–36665. [Google Scholar] [CrossRef]
- Fukushima, S.; Kato, S.; Maeda, M.; Shigemori, M. Caspase-9 pathway activation by inhibiting endogenous fibroblast growth factor signaling in human glioma cells. Int. J. Oncol. 2008, 32, 467–473. [Google Scholar] [CrossRef]
- Rifkin, D.B.; Moscatelli, D.; Roghani, M.; Nagano, Y.; Quarto, N.; Klein, S.; Bikfalvi, A. Studies on FGF-2: Nuclear localization and function of high molecular weight forms and receptor binding in the absence of heparin. Mol. Reprod. Dev. 1994, 39, 102–105. [Google Scholar] [CrossRef]
- Koleini, N.; Santiago, J.-J.; Nickel, B.E.; Sequiera, G.L.; Wang, J.; Fandrich, R.R.; Jassal, D.S.; Dhingra, S.; Kirshenbaum, L.A.; Cattini, P.A.; et al. Elimination or neutralization of endogenous high-molecular-weight FGF2 mitigates doxorubicin-induced cardiotoxicity. Am. J. Physiol. Circ. Physiol. 2019, 316, H279–H288. [Google Scholar] [CrossRef]
- Peluso, J.J. Basic fibroblast growth factor (bFGF) regulation of the plasma membrane calcium ATPase (PMCA) as part of an anti-apoptotic mechanism of action. Biochem. Pharmacol. 2003, 66, 1363–1369. [Google Scholar] [CrossRef]
- Kim, H.R.; Heo, Y.M.; Jeong, K.I.; Kim, Y.M.; Jang, H.L.; Lee, K.Y.; Yeo, C.Y.; Kim, S.H.; Lee, H.K.; Kim, S.R.; et al. Choi. Fgf-2 Inhibits Tnf-Alpha Mediated Apoptosis through Upregulation of Bcl2-A1 and Bcl-Xl in Atdc5 Cells. BMB Rep. 2012, 45, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Tahara, N.; Akiyama, R.; Wang, J.; Kawakami, H.; Bessho, Y.; Kawakami, Y. The FGF-AKT pathway is necessary for cardiomyocyte survival for heart regeneration in zebrafish. Dev. Biol. 2021, 472, 30–37. [Google Scholar] [CrossRef]
- Cooke, J.P. Inflammation and Its Role in Regeneration and Repair. Circ. Res. 2019, 124, 1166–1168. [Google Scholar] [CrossRef]
- Wang, C.; Li, Y.; Li, H.; Zhang, Y.; Ying, Z.; Wang, X.; Zhang, T.; Zhang, W.; Fan, Z.; Li, X.; et al. Disruption of FGF Signaling Ameliorates Inflammatory Response in Hepatic Stellate Cells. Front. Cell Dev. Biol. 2020, 8, 601. [Google Scholar] [CrossRef]
- Wang, C.; Ke, Y.; Liu, S.; Pan, S.; Liu, Z.; Zhang, H.; Fan, Z.; Zhou, C.; Liu, J.; Wang, F. Ectopic Fibroblast Growth Factor Receptor 1 Promotes Inflammation by Promoting Nuclear Factor-Kappab Signaling in Prostate Cancer Cells. J. Biol. Chem. 2018, 293, 14839–14849. [Google Scholar] [CrossRef] [Green Version]
- Tsunoda, S.; Sakurai, H.; Saito, Y.; Ueno, Y.; Koizumi, K.; Saiki, I. Massive T-Lymphocyte Infiltration into the Host Stroma Is Essential for Fibroblast Growth Factor-2-Promoted Growth and Metastasis of Mammary Tumors via Neovascular Stability. Am. J. Pathol. 2009, 174, 671–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Wang, Q.; Ma, H.; Yan, W.; Yang, J. Knockout of Low Molecular Weight FGF2 Attenuates Atherosclerosis by Reducing Macrophage Infiltration and Oxidative Stress in Mice. Cell. Physiol. Biochem. 2018, 45, 1434–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirov, A.; Duarte, M.; Guay, J.; Karolak, M.; Yan, C.; Oxburgh, L.; Prudovsky, I. Transgenic Expression of Nonclassically Secreted FGF Suppresses Kidney Repair. PLoS ONE 2012, 7, e36485. [Google Scholar] [CrossRef] [PubMed]
- Meij, J.T.A.; Sheikh, F.; Jimenez, S.K.; Nickerson, P.W.; Kardami, E.; Cattini, P.A. Exacerbation of myocardial injury in transgenic mice overexpressing FGF-2 is T cell dependent. Am. J. Physiol. Circ. Physiol. 2002, 282, H547–H555. [Google Scholar] [CrossRef] [Green Version]
- Bovolenta, R.; Zucchini, S.; Paradiso, B.; Rodi, D.; Merigo, F.; Mora, G.N.; Osculati, F.; Berto, E.; Marconi, P.; Marzola, A.; et al. Hippocampal Fgf-2 and Bdnf Overexpression Attenuates Epileptogenesis-Associated Neuroinflammation and Reduces Spontaneous Recurrent Seizures. J. Neuroinflamm. 2010, 7, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, H.-J.; Zhao, C.-F.; Chen, Z.-W.; Lin, W.; Jiang, Y.-C. Fibroblast Growth Factor (FGF) Signaling Protects Against Acute Pancreatitis-Induced Damage by Modulating Inflammatory Responses. Med. Sci. Monit. 2020, 26, e920684-1. [Google Scholar] [CrossRef]
- Fan, L.; Ding, L.; Lan, J.; Niu, J.; He, Y.; Song, L. Fibroblast Growth Factor-1 Improves Insulin Resistance via Repression of JNK-Mediated Inflammation. Front. Pharmacol. 2019, 10, 1478. [Google Scholar] [CrossRef] [Green Version]
- Maciag, T.; A Hoover, G.; Stemerman, M.B.; Weinstein, R. Serial propagation of human endothelial cells in vitro. J. Cell Biol. 1981, 91, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Gospodarowicz, D.; Zetter, B.R. The use of fibroblast and epidermal growth factors to lower the serum requirement for growth of normal diploid cells in early passage: A new method for cloning. Dev. Boil. Stand. 1976, 37, 109–130. [Google Scholar]
- Korc, M.; Friesel, R.E. The Role of Fibroblast Growth Factors in Tumor Growth. Curr. Cancer Drug Targets 2009, 9, 639–651. [Google Scholar] [CrossRef]
- Presta, M.; Dell’Era, P.; Mitola, S.; Moroni, E.; Ronca, R.; Rusnati, M. Fibroblast Growth Factor/Fibroblast Growth Factor Receptor System in Angiogenesis. Cytokine Growth Factor Rev. 2005, 16, 159–178. [Google Scholar] [CrossRef] [Green Version]
- Oladipupo, S.S.; Smith, C.; Santeford, A.; Park, C.; Sene, A.; Wiley, L.A.; Osei-Owusu, P.; Hsu, J.; Zapata, N.; Liu, F.; et al. Endothelial cell FGF signaling is required for injury response but not for vascular homeostasis. Proc. Natl. Acad. Sci. USA 2014, 111, 13379–13384. [Google Scholar] [CrossRef] [Green Version]
- Ribatti, D.; Nico, B.; Vacca, A.; Roncali, L.; Presta, M. Endogenous and exogenous fibroblast growth factor-2 modulate wound healing in the chick embryo chorioallantoic membrane. Angiogenesis 1999, 3, 89–95. [Google Scholar] [CrossRef]
- Nagaraja, S.; Chen, L.; DiPietro, L.A.; Reifman, J.; Mitrophanov, A.Y. Predictive Approach Identifies Molecular Targets and Interventions to Restore Angiogenesis in Wounds with Delayed Healing. Front. Physiol. 2019, 10, 636. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Khalil, R.A. Matrix Metalloproteinases, Vascular Remodeling, and Vascular Disease. Adv. Pharmacol. 2018, 81, 241–330. [Google Scholar] [CrossRef]
- Yasui, H.; Andoh, A.; Bamba, S.; Inatomi, O.; Ishida, H.; Fujiyama, Y. Role of Fibroblast Growth Factor-2 in the Expression of Matrix Metalloproteinases and Tissue Inhibitors of Metalloproteinases in Human Intestinal Myofibroblasts. Digestion 2004, 69, 34–44. [Google Scholar] [CrossRef]
- Pintucci, G.; Yu, P.J.; Sharony, R.; Baumann, F.G.; Saponara, F.; Frasca, A.; Galloway, A.C.; Moscatelli, D.; Mignatti, P. Induction of Stromelysin-1 (Mmp-3) by Fibroblast Growth Factor-2 (Fgf-2) in Fgf-2-/- Microvascular Endothelial Cells Requires Prolonged Activation of Extracellular Signal-Regulated Kinases-1 and -2 (Erk-1/2). J. Cell Biochem. 2003, 90, 1015–1025. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Vaughan, D.E. PAI-1 in tissue fibrosis. J. Cell. Physiol. 2012, 227, 493–507. [Google Scholar] [CrossRef] [Green Version]
- Sakaki, T.; Yamada, K.; Otsuki, H.; Yuguchi, T.; Kohmura, E.; Hayakawa, T. Brief exposure to hypoxia induces bFGF mRNA and protein and protects rat cortical neurons from prolonged hypoxic stress. Neurosci. Res. 1995, 23, 289–296. [Google Scholar] [CrossRef]
- Ganat, Y.; Soni, S.; Chacon, M.; Schwartz, M.L.; Vaccarino, F.M. Chronic hypoxia up-regulates fibroblast growth factor ligands in the perinatal brain and induces fibroblast growth factor-responsive radial glial cells in the sub-ependymal zone. Neuroscience 2002, 112, 977–991. [Google Scholar] [CrossRef]
- Kakudo, N.; Morimoto, N.; Ogawa, T.; Taketani, S.; Kusumoto, K. Hypoxia Enhances Proliferation of Human Adipose-Derived Stem Cells via HIF-1α Activation. PLoS ONE 2015, 10, e0139890. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Qiao, F.; Yin, X. Hypoxia Induces Fgf2 Production by Vascular Endothelial Cells and Alters Mmp9 and Timp1 Expression in Extravillous Trophoblasts and Their Invasiveness in a Cocultured Model. J. Reprod. Dev. 2011, 57, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.-S.; Zhu, H.-J.; Zhang, B.; Li, H.; Yan, H.; Wang, B. Inhibitive Effect of Genistein on Hypoxia-Induced Basic Fibroblast Growth Factor Expression in Human Retinal Pigment Epithelium Cells. J. Ocul. Pharmacol. Ther. 2006, 22, 103–109. [Google Scholar] [CrossRef]
- Kuwabara, K.; Ogawa, S.; Matsumoto, M.; Koga, S.; Clauss, M.; Pinsky, D.J.; Lyn, P.; Leavy, J.; Witte, L.; Joseph-Silverstein, J. Hypoxia-mediated induction of acidic/basic fibroblast growth factor and platelet-derived growth factor in mononuclear phagocytes stimulates growth of hypoxic endothelial cells. Proc. Natl. Acad. Sci. USA 1995, 92, 4606–4610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheraga, R.; Thompson, C.; Tulapurkar, M.E.; Nagarsekar, A.C.; Cowan, M.; Potla, R.; Sun, J.; Cai, R.; Logun, C.; Shelhamer, J.; et al. Activation of heat shock response augments fibroblast growth factor-1 expression in wounded lung epithelium. Am. J. Physiol. Cell. Mol. Physiol. 2016, 311, L941–L955. [Google Scholar] [CrossRef] [Green Version]
- Song, J.S.; Lee, L.J.; Kay, E.P. Induction of Fgf-2 Synthesis by Il-1beta in Aqueous Humor through P13-Kinase and P38 in Rabbit Corneal Endothelium. Investig. Ophthalmol. Vis. Sci. 2010, 51, 822–829. [Google Scholar] [CrossRef]
- Sobue, T.; Zhang, X.; Florkiewicz, R.; Hurley, M. Interleukin-1 Regulates FGF-2 mRNA and Localization of FGF-2 Protein in Human Osteoblasts. Biochem. Biophys. Res. Commun. 2001, 286, 33–40. [Google Scholar] [CrossRef]
- Chien, S.Y.; Huang, C.Y.; Tsai, C.H.; Wang, S.W.; Lin, Y.M.; Tang, C.H. Interleukin-1beta Induces Fibroblast Growth Factor 2 Expression and Subsequently Promotes Endothelial Progenitor Cell Angiogenesis in Chondrocytes. Clin. Sci. 2016, 130, 667–681. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.G.; Kay, E.P. Nf-Kappab is the Transcription Factor for Fgf-2 That Causes Endothelial Mesenchymal Transformation in Cornea. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1530–1538. [Google Scholar] [CrossRef]
- Chedid, M.; Rubin, J.S.; Csaky, K.G.; Aaronson, S.A. Regulation of keratinocyte growth factor gene expression by interleukin 1. J. Biol. Chem. 1994, 269, 10753–10757. [Google Scholar] [CrossRef]
- Prudovsky, I.; Mandinova, A.; Soldi, R.; Bagala, C.; Graziani, I.; Landriscina, M.; Tarantini, F.; Duarte, M.F.; Bellum, S.; Doherty, H.; et al. The non-classical export routes: FGF1 and IL-1α point the way. J. Cell Sci. 2003, 116, 4871–4881. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.; Friedman, S.; Zhan, X.; Engleka, K.A.; Forough, R.; Maciag, T.; Jackson, A.; Friedman, S.; Zhan, X.; Engleka, K.A.; et al. Heat shock induces the release of fibroblast growth factor 1 from NIH 3T3 cells. Proc. Natl. Acad. Sci. USA 1992, 89, 10691–10695. [Google Scholar] [CrossRef] [Green Version]
- Carreira, C.M.; Landriscina, M.; Bellum, S.; Prudovsky, I.; Maciag, T. The comparative release of FGF1 by hypoxia and temperature stress. Growth Factors 2001, 18, 277–285. [Google Scholar] [CrossRef]
- Di Serio, C.; Cozzi, A.; Angeli, I.; Doria, L.; Micucci, I.; Pellerito, S.; Mirone, P.; Masotti, G.; Moroni, F.; Tarantini, F. Kynurenic Acid Inhibits the Release of the Neurotrophic Fibroblast Growth Factor (FGF)-1 and Enhances Proliferation of Glia Cells, in vitro. Cell. Mol. Neurobiol. 2005, 25, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Landriscina, M.; Bagalá, C.; Mandinova, A.; Soldi, R.; Micucci, I.; Bellum, S.; Prudovsky, I.; Maciag, T. Copper Induces the Assembly of a Multiprotein Aggregate Implicated in the Release of Fibroblast Growth Factor 1 in Response to Stress. J. Biol. Chem. 2001, 276, 25549–25557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landriscina, M.; Soldi, R.; Bagala, C.; Micucci, I.; Bellum, S.; Tarantini, F.; Prudovsky, I.; Maciag, T. S100a13 Participates in the Release of Fibroblast Growth Factor 1 in Response to Heat Shock in Vitro. J. Biol. Chem. 2001, 276, 22544–22552. [Google Scholar] [CrossRef] [Green Version]
- Tarantini, F.; LaVallee, T.; Jackson, A.; Gamble, S.; Carreira, C.M.; Garfinkel, S.; Burgess, W.H.; Maciag, T. The Extravesicular Domain of Synaptotagmin-1 Is Released with the Latent Fibroblast Growth Factor-1 Homodimer in Response to Heat Shock. J. Biol. Chem. 1998, 273, 22209–22216. [Google Scholar] [CrossRef] [Green Version]
- Bagalá, C.; Kolev, V.; Mandinova, A.; Soldi, R.; Mouta, C.; Graziani, I.; Prudovsky, I.; Maciag, T. The alternative translation of synaptotagmin 1 mediates the non-classical release of FGF1. Biochem. Biophys. Res. Commun. 2003, 310, 1041–1047. [Google Scholar] [CrossRef]
- Soldi, R.; Mandinova, A.; Venkataraman, K.; Hla, T.; Vadas, M.; Pitson, S.; Duarte, M.; Graziani, I.; Kolev, V.; Kacer, D.; et al. Sphingosine kinase 1 is a critical component of the copper-dependent FGF1 export pathway. Exp. Cell Res. 2007, 313, 3308–3318. [Google Scholar] [CrossRef] [Green Version]
- Kirov, A.; Kacer, D.; Conley, B.A.; Vary, C.P.; Prudovsky, I. AHNAK2 Participates in the Stress-Induced Nonclassical FGF1 Secretion Pathway. J. Cell. Biochem. 2015, 116, 1522–1531. [Google Scholar] [CrossRef] [Green Version]
- Kirov, A.; Al-Hashimi, H.; Solomon, P.; Mazur, C.; Thorpe, P.E.; Sims, P.J.; Tarantini, F.; Kumar, S.T.; Prudovsky, I. Phosphatidylserine externalization and membrane blebbing are involved in the nonclassical export of FGF1. J. Cell. Biochem. 2012, 113, 956–966. [Google Scholar] [CrossRef] [Green Version]
- Prudovsky, I.; Kacer, D.; Davis, J.; Shah, V.; Jayanthi, S.; Huber, I.; Dakshinamurthy, R.; Ganter, O.; Soldi, R.; Neivandt, D.; et al. Folding of Fibroblast Growth Factor 1 Is Critical for Its Nonclassical Release. Biochemistry 2016, 55, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Small, D.; Kovalenko, D.; Kacer, D.; Liaw, L.; Landriscina, M.; Di Serio, C.; Prudovsky, I.; Maciag, T. Soluble Jagged 1 Represses the Function of Its Transmembrane Form to Induce the Formation of the Src-dependent Chord-like Phenotype. J. Biol. Chem. 2001, 276, 32022–32030. [Google Scholar] [CrossRef] [Green Version]
- Small, D.; Kovalenko, D.; Soldi, R.; Mandinova, A.; Kolev, V.; Trifonova, R.; Bagala, C.; Kacer, D.; Battelli, C.; Liaw, L.; et al. Notch Activation Suppresses Fibroblast Growth Factor-dependent Cellular Transformation. J. Biol. Chem. 2003, 278, 16405–16413. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.; Kolev, V.; Kacer, D.; Mouta-Bellum, C.; Soldi, R.; Graziani, I.; Kirov, A.; Friesel, R.; Liaw, L.; Small, D.; et al. Novel Cross-Talk between Three Cardiovascular Regulators: Thrombin Cleavage Fragment of Jagged1 Induces Fibroblast Growth Factor 1 Expression and Release. Mol. Biol. Cell 2008, 19, 4863–4874. [Google Scholar] [CrossRef] [Green Version]
- Pallotta, M.T.; Nickel, W. Fgf2 and Il-1beta—Explorers of Unconventional Secretory Pathways at a Glance. J. Cell Sci. 2020, 133, 21. [Google Scholar] [CrossRef]
- Hennig, T.; Mogensen, C.; Kirsch, J.; Pohl, U.; Gloe, T. Shear Stress Induces the Release of an Endothelial Elastase: Role in Integrin Alpha(V)Beta(3)-Mediated Fgf-2 Release. J. Vasc. Res. 2011, 48, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Tholozan, F.M.; Gribbon, C.; Li, Z.; Goldberg, M.W.; Prescott, A.R.; McKie, N.; Quinlan, R.A. FGF-2 Release from the Lens Capsule by MMP-2 Maintains Lens Epithelial Cell Viability. Mol. Biol. Cell 2007, 18, 4222–4231. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Maccarana, M.; Min, T.H.; Strate, I.; Pera, E.M. The Secreted Serine Protease xHtrA1 Stimulates Long-Range FGF Signaling in the Early Xenopus Embryo. Dev. Cell 2007, 13, 226–241. [Google Scholar] [CrossRef]
- Kole, D.; Grella, A.; Dolivo, D.; Shumaker, L.; Hermans, W.; Dominko, T. High molecular weight FGF2 isoforms demonstrate canonical receptor-mediated activity and support human embryonic stem cell self-renewal. Stem Cell Res. 2017, 21, 106–116. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cell Type | FGF | Biological Effect | Mechanism (If Known) | Ref. |
|---|---|---|---|---|
| Endothelial cells | FGF2 | Lifespan extension | Telomerase activity increase | [55] |
| Endothelial cells | FGF2 | Apoptosis suppression | Increase of Bcl2 expression | [56] |
| Endothelial cells | FGF1 | Increase of MMP1 and MMP3 expression | Increase of proinflammatory cytokines | [57] |
| Endothelial cells | FGF2 | Increase of MMP2 expression | Activation of FGFR/JNK pathway | [58] |
| Endothelial cells | FGF2 | Induction of uPA expression | [59] | |
| Endothelial cells | FGF1, FGF2, FGF9 | Enhancement of angiogenesis | Simulation of proliferation, increase of cell–cell adhesion | [60,61,62,63,64,65,66,67,68,69] |
| VSMC | FGF1, FGF2 | De-differentiation | Suppression of TGFβ signaling | [70,71] |
| VSMC | FGF1 | Increase of proinflammatory cytokines expression | [70] | |
| Fibroblasts | FGF2 | Suppression of differentiation to myofibroblasts | Suppression of TGFβ signaling | [72] (review) |
| Fibroblasts | FGF2 | Lifespan extension | Increase of OCT4, SOX2 and NANOG expression | [73] |
| Fibroblasts | FGF2 | Increase of MMP1, ADAMTS8, MMP27, MMP10, and MMP3 expression | [74] | |
| Thyroid Epithelial cells | FGF3 | Suppression of differentiation | [75] | |
| Mammary epithelial cells | FGF1 | Increase of MMP9 expression | Stimulation of NFκB signaling | [76] |
| Keratinocytes | FGF2 | De-differentiation | Increase of Snail 2 expression | [77] |
| Keratinocytes | FGF7 | Increase of TNFα expression | Activation of FGFR2/Akt/NFκB pathway | [78] |
| Cardiomyocytes | FGF2 | Protection from necrosis | Activation of PI3K/Akt pathway | [79] |
| Cardiomyocytes | FGF2 | Protection from doxorubicin toxic effect | Activation of mTOR/Nrf-2/HO1 pathway | [80] |
| Chondrocytes | FGF2 | De-differentiation | [81] | |
| Chondrocytes | FGF2 | Induction of inflammatory phenotype | Mediated by ILβ | [82] |
| Osteoclasts | FGF2 | Increase of MMP9 and cathepsin K expression | Activation of FGFR1/MAPK signaling | [83] |
| Neurons | FGF1 | Suppression of apoptosis | Decrease of the expression of XIAP and caspases 9 and 3 | [84] |
| Neurons | FGF2 | Suppression of apoptosis | Activation of PI3K/Akt pathway | [85] |
| Oligodendrocytes | FGF2 | Suppression of apoptosis | [86] | |
| Mesenchymal stem cells | FGF2 | Suppression of apoptosis | Increase of Bcl2 expression | [87] |
| Mesenchymal stem cells | FGF2 | Suppression of senescence | Decrease of p16, p21 and p53 expression | [88] |
| Male germ cells | FGF4 | Suppression of apoptosis | [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prudovsky, I. Cellular Mechanisms of FGF-Stimulated Tissue Repair. Cells 2021, 10, 1830. https://doi.org/10.3390/cells10071830
Prudovsky I. Cellular Mechanisms of FGF-Stimulated Tissue Repair. Cells. 2021; 10(7):1830. https://doi.org/10.3390/cells10071830
Chicago/Turabian StylePrudovsky, Igor. 2021. "Cellular Mechanisms of FGF-Stimulated Tissue Repair" Cells 10, no. 7: 1830. https://doi.org/10.3390/cells10071830
APA StylePrudovsky, I. (2021). Cellular Mechanisms of FGF-Stimulated Tissue Repair. Cells, 10(7), 1830. https://doi.org/10.3390/cells10071830