
The RHO Family GTPases: Mechanisms of Regulation and Signaling


Abstract
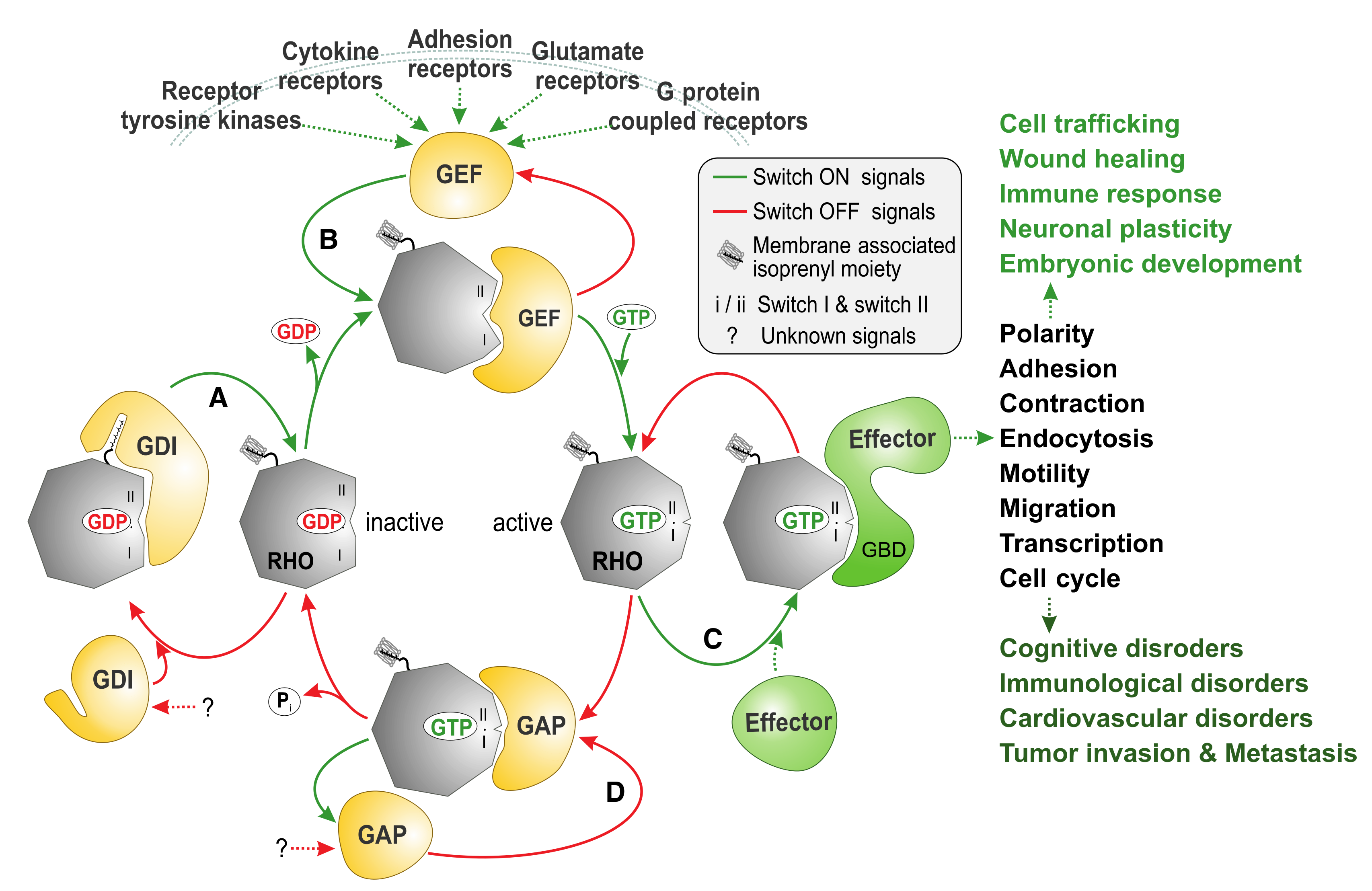
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RHO GTPases | Effector Proteins | Function | Functions and Effects | References |
|---|---|---|---|---|
| RHOA | ROCK I/II | Ser /Thr kinase | Actin myosin contraction, Stress fiber formation | [10,11] |
| Citron kinase | Ser /Thr kinase | Cytokinesis | [12] | |
| MBS | Phosphatase subunit | MLC inactivation | [13] | |
| DIA 1/2 | Formin-like proteins | Actin polymerization | [10] | |
| RHOB | Integrin β1 | Cell surface receptor | Cell adhesion and migration | [14] |
| RHOC | FMNL3 | Formin like proteins | Migration, Invasion | [15] |
| RHOH | Kaiso | Transcription factor | TCR activation | [16] |
| RAC1 | PAK1/2/3 * | Ser /Thr kinase | JNK activation, Actin filament stabilization | [17] |
| MLK 2/3 * | Ser /Thr kinase | JNK activation | [18,19] | |
| WAVE | Scaffold | Actin organization | [20] | |
| p70 S6 kinase * | Ser /Thr kinase | Translation regulation | [21] | |
| IQGAP1/2 * | Scaffold | Actin/cell-cell contacts | [22,23] | |
| MEKK1/4 * | Ser /Thr kinase | JNK activation | [24] | |
| POR1 | Scaffold | Actin organization | [25] | |
| p67phox* | Scaffold | ROS generation | [26] | |
| PI3 kinase | Lipid kinase | PIP3 levels | [27] | |
| DAG kinase | Lipid kinase | PA levels | [28,29] | |
| PLCβ2 * | Lipase | DAG and IP3 levels | [30] | |
| RAC1B | p120ctn | Catenin | Cellular transformation | [31] |
| RAC2 | LFA-1 | Cell surface receptor | B cell adhesion | [32] |
| RAC3 | GIT1 | ARF GAP and scaffold | Regulation of cell adhesion and differentiation | [33] |
| RHOG | Kinectin | Kinesin receptor | Microtubule dependent transport | [34] |
| CDC42 | N-WASP | Scaffold | Actin organization | [35] |
| PAK4 | Ser/Thr kinase | Actin organization | [36] | |
| MRCKα/β | Ser/Thr kinase | Actin organization | [37] | |
| TCL | GIT-PIX complex | Scaffold | Stabilization of focal adhesion | [38,39] |
| RHOD | Plexin A1/B1 | Semaphorin co-receptor | Growth cone formation | [40] |
| RIF | DIA 1/2 | Formin-like proteins | Actin organization | [40,41] |
| RND1 | Stathmin2 | Neuronal growth associated proteins | Microtubule depolymerization, Neurite extension | [42] |
| RND2 | Rapostlin | Formin-binding protein | Neurite branching | [43] |
| RND3 | Socius | Scaffold | Loss of stress fibers | [44] |
| ROCKI | Ser/Thr kinase | Actomyosin contractility | [45,46] |
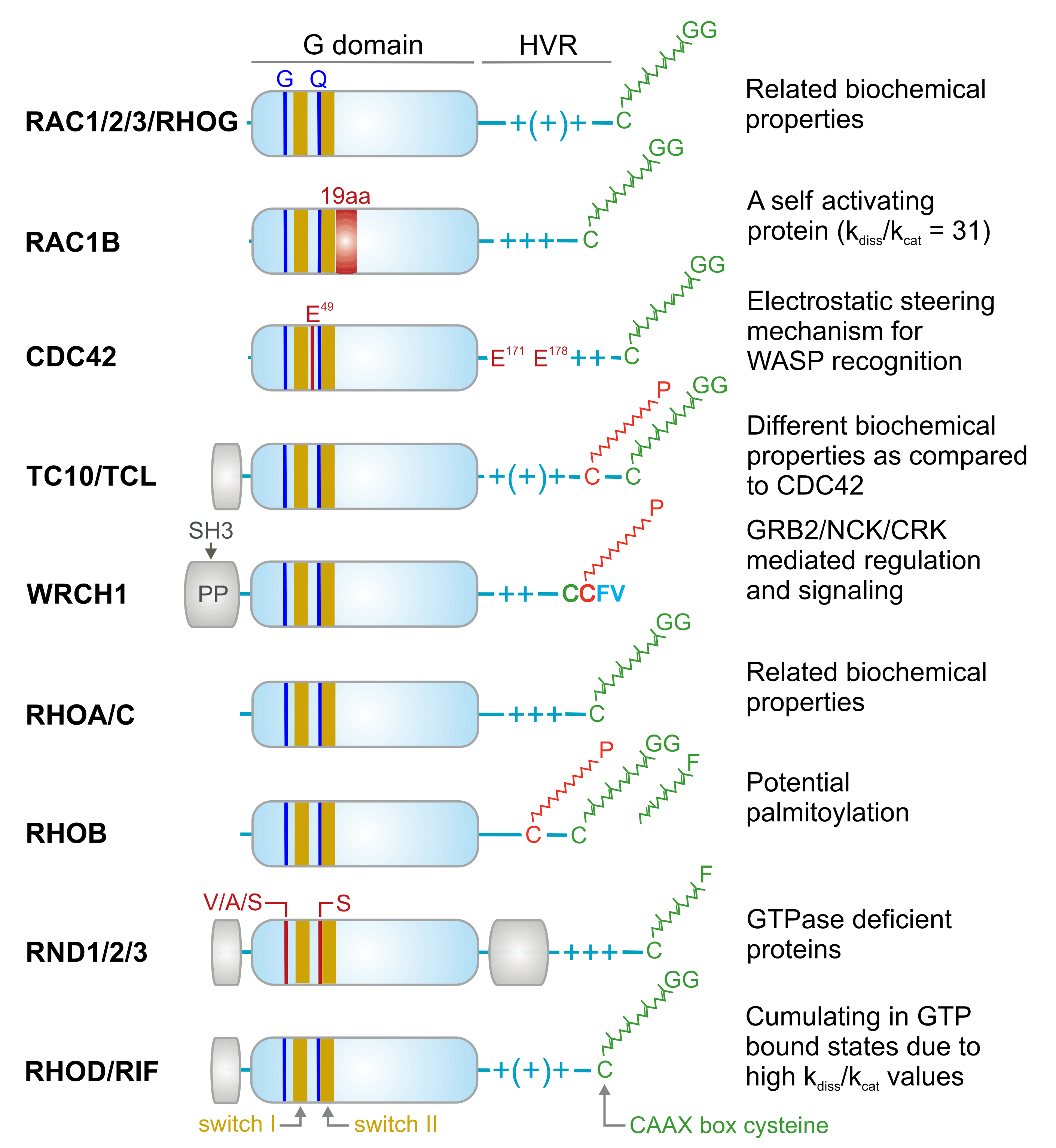
2. The RHO Family and the Molecular Switch Mechanism

3. Regulation of RHO Family GTPases
3.1. Guanine Nucleotide Dissociation Inhibitors (GDIs)
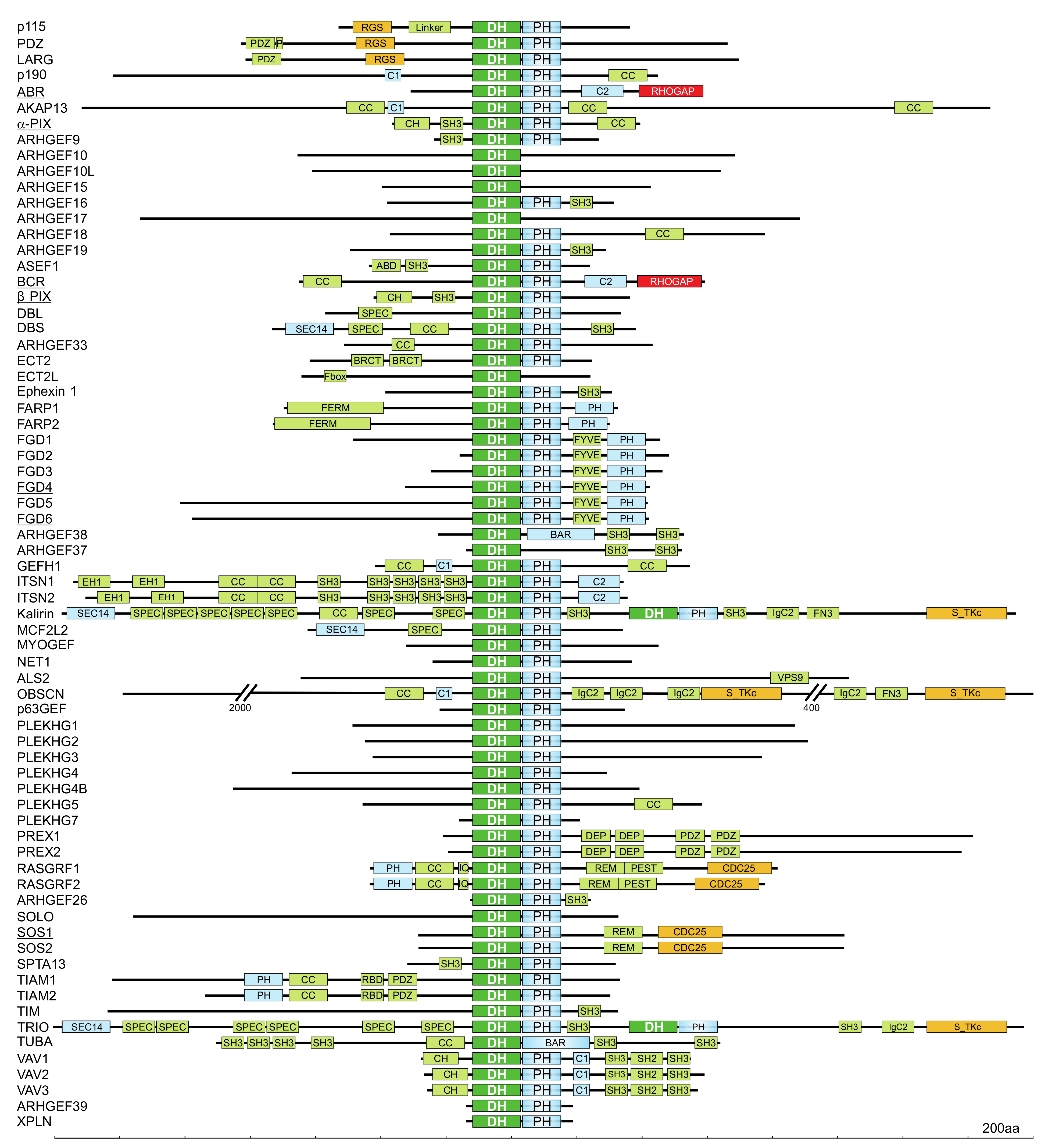
3.2. Guanine Nucleotide Exchange Factors (GEFs)
3.2.1. DBL Family GEFs
3.2.2. Structural and Functional Characteristics of the DH domain
3.2.3. The Tandem PH Domain in DBL Proteins
3.2.4. A Plethora of DBL Family Proteins
3.3. DOCK Family of RHOGEFs
3.4. GTPase-Activating Proteins (GAPs)
3.4.1. RHOGAP Family Proteins
3.4.2. Structural and Functional Characteristics of the RHOGAP Domain
3.4.3. The Mechanism by Which the GAP Domain Mediates GTP Hydrolysis
3.4.4. Overabundance and Diversity
3.4.5. Regulation and GAP Proteins Functions
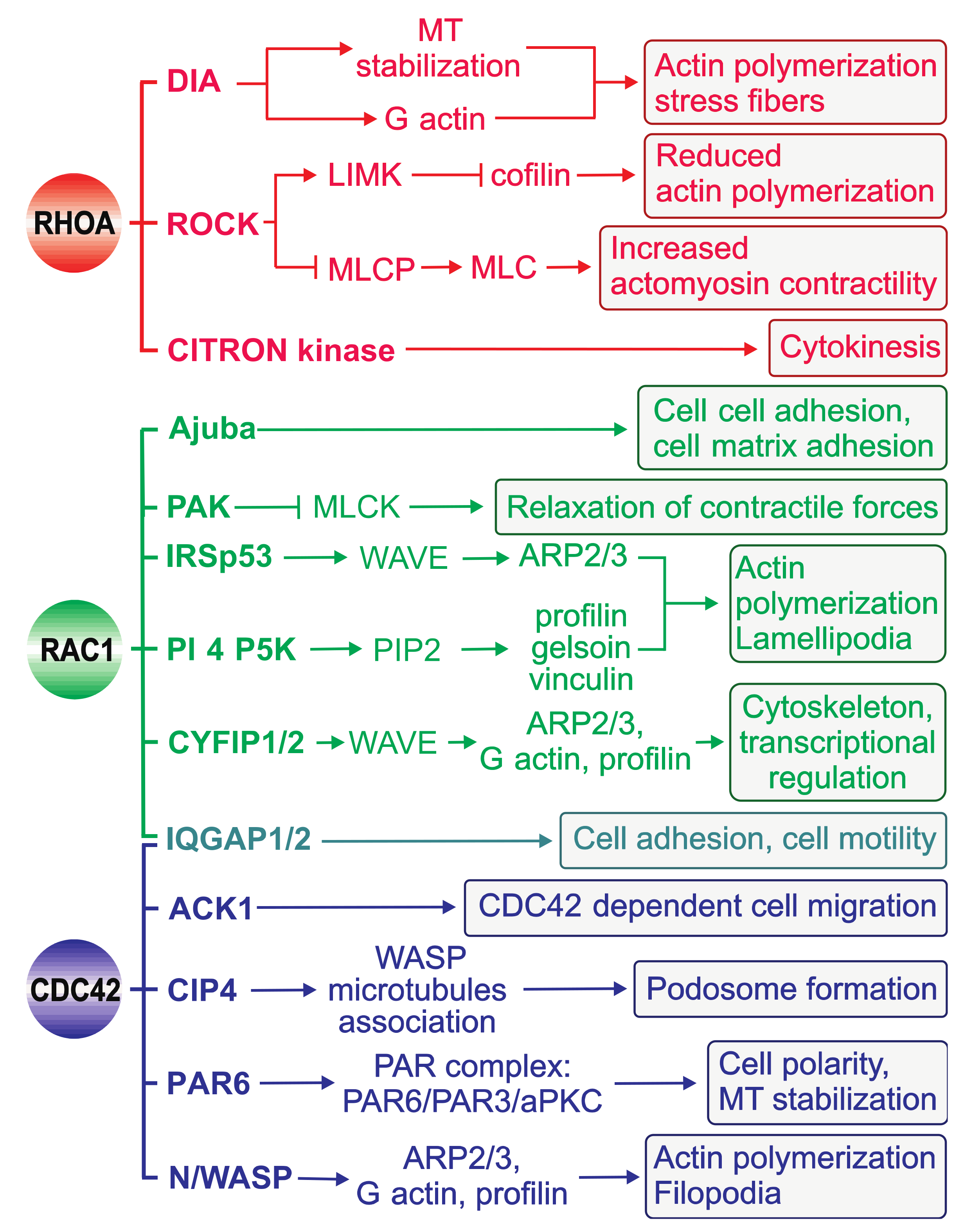
4. Downstream Effectors of RHO GTPases
4.1. Structural Characteristics of RHO GTPase-effector Interactions
4.2. RHO GTPase-Mediated Effector Signaling
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABD | APC binding domain |
| ACK1 | Activated CDC42 kinase 1 |
| AlF4 | Tetrafluoroaluminate |
| ANK | Ankyrin-repeat |
| ARP2/3 | Actin related protein 2/3 |
| ASH | ASPM-SPD-2-Hydin |
| BAR | BIN–Amphiphysin–RVS |
| BCR | Breakpoint cluster region |
| BRCT | BRCA1 C-terminal domain |
| C1 | DAG-binding domain |
| C2 | Calcium/lipid-binding domain |
| CC | Coiled-coil |
| CDC25 | Catalytic RASGEF domain |
| CDC42 | Cell division control protein 42 |
| CH | Calponin homology |
| CIP4 | CDC42 interacting protein |
| CNTD1 | Cyclin N-terminal domain-containing protein |
| CYFIP1/2 | Cytoplasmic FMR-1 interacting protein 1/2 |
| DAG kinase | Diacyglycerol kinase |
| DAG | Diacylglycerol |
| DBL | Diffuse B-cell lymphoma |
| DEP | Dishevelled, Egl-10 and Pleckstrin domain |
| DH | DBL homology |
| DIA | Diaphanous-related formin |
| DLC1 | Deleted in liver Cancer |
| EH1 | EPS15 homology |
| F-BAR | Extended Fes-CIP4 homology (FCH)-BAR |
| Far | Farnesyl moiety |
| FERM | 4.1 protein/ezrin/radixin/moesin |
| FF | Conserved phenylalanines |
| FMNL3 | Formin-like protein 3 |
| FYVE | FAB1/YOTB/VAC1/EEA1 |
| GAPs | GTPase activating proteins |
| GDIs | Guanine nucleotide dissociation inhibitors |
| GDP | Guanosine-5′-diphosphate |
| GEFs | Guanine nucleotide exchange factors |
| GG | Geranylgeranyl moiety |
| GIT | G protein-coupled receptor kinase-interactor 1 |
| GppNHp | Guanosine-5′- [(β, γ)-imido] triphosphate, Trisodium salt |
| GTP | Guanosine-5′-triphosphate |
| GTPase | GTP-binding and hydrolyzing domain |
| GTPase | Guanine triphosphatase |
| HVR | Hypervariable region |
| ICMT | Isoprenylcystein carboxyl methyltransferase |
| IP3 | Inositol triphosphate |
| IPP5 | Inositol polyphosphate 5-phosphatase |
| IQ | Isoleucine-glutamine-rich |
| IQGAP | IQ motif-containing GTPase activating proteins |
| IRSp53 | Insulin receptor substrate 53 kDa |
| kDa | Kilodalton |
| LFA-1 | Leukocyte function associated molecule 1 alpha |
| lgC2 | Immunoglobulin C-2 Type |
| LIMK | LIM domain kinase |
| MBS | Myosin binding subunit |
| MEKK | Mitogen Activated Protein kinase kinase kinase |
| MgF3 | Magnesium fluoride |
| MLC | Myosin light chain |
| MLCK | Myosin light chain kinase |
| MLCP | Myosin light chain phosphatase |
| MLK | Mixed lineage kinase |
| MRCK | Myotonic dystrophy kinase related- CDC42 binding Kinase |
| MT | Microtubule |
| MYSc | Myosin head domain |
| MyTH4 | Myosin tail homology 4 |
| OCRL1 | Lowe oculocerebrorenal syndrome protein |
| OPHN1 | Oligophrenin |
| P | Proline-rich |
| PA | Phosphatidic acid |
| PAK | p21 activated kinase |
| P | Palmitoyl moiety |
| PAR6 | Partitioning defective 6 homolog |
| PBR | Polybasic region |
| PDZ | PSD95/DLG/ZO-1 |
| PEST | Proline-glutamic acid-serine-threonine-rich |
| PH | Pleckstrin homology |
| PI3 kinase | Phosphatidylinositol 3-kinase |
| PI4P5K | Phosphatidylinositol 4, phosphate 5 kinase |
| PIP2 | Phosphatidylinositol 4,5 biphosphate |
| PIP3 | Phosphatidylinosiltol-3,4,5-triphosphate |
| PIX | p21-activated protein kinase exchange factor |
| PLC | Phospholipase C |
| POR1 | Partner of RAC1 |
| RA | RAS association domain |
| RAC | RAS-related C3 botulinum toxin substrate |
| RALBD | RAL-binding domain |
| RBD | RAS-binding domain |
| Rce1 | RAS converting enzyme |
| REM | RAS exchanger motif |
| RGS | Regulator of G protein signaling |
| RHO | RAS homolog |
| ROCK | RHO-associated protein kinase |
| S_TKc | Serine/Threonine protein kinase catalytic domain |
| SAM | Sterile alpha motif |
| SEC14 | Phosphoinositide binding |
| SH | SRC homology |
| SH2 | SRC homology 2 |
| SH3 | SRC homology 3 |
| SPEC | Spectrin repeat |
| StART | Steroidogenic acute regulatory protein-related lipid transfer |
| TCR | T cell receptor |
| VPS9 | Vacuolar protein sorting-associated 9 |
| WASP | Wiskott-Aldrich Syndrome Protein |
| WAVE | WASP family verprolin homologous protein |
| WW | Conserved tryptophans |
References
- Jaffe, A.B.; Hall, A. Rho GTPases: Biochemistry and biology. Annu. Rev. Cell Dev. Biol. 2005, 21, 247–269. [Google Scholar] [CrossRef] [Green Version]
- Ellenbroek, S.I.J.; Collard, J.G. Rho GTPases: Functions and association with cancer. Clin. Exp. Metastasis 2007, 24, 657–672. [Google Scholar] [CrossRef]
- Dvorsky, R.; Ahmadian, M.R. Always look on the bright site of Rho: Structural implications for a conserved intermolecular interface. EMBO Rep. 2004, 5, 1130–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherfils, J.; Zeghouf, M. Regulation of small GTPases by GEFs, GAPs, and GDIs. Physiol. Rev. 2013, 93, 269–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, A.L.; Hall, A. Rho GTPases and their effector proteins. Biochem. J. 2000, 348, 241–255. [Google Scholar] [CrossRef]
- Burridge, K.; Wennerberg, K. Rho and Rac Take Center Stage. Cell 2004, 116, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Etienne-Manneville, S.; Hall, A. Rho GTPases in cell biology. Nature 2002, 420, 629–635. [Google Scholar] [CrossRef]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef] [PubMed]
- Knyphausen, P.; Kuhlmann, N.; de Boor, S.; Lammers, M. Lysine-acetylation as a fundamental regulator of ran function: Implications for signaling of proteins of the ras-superfamily. Small GTPases 2015, 6, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, N.; Kato, T.; Fujita, A.; Ishizaki, T.; Narumiya, S. Cooperation between mDia1 and ROCK in Rho-induced actin reorganization. Nat. Cell Biol. 1999, 1, 136–143. [Google Scholar] [CrossRef]
- Maekawa, M.; Ishizaki, T.; Boku, S.; Watanabe, N.; Fujita, A.; Iwamatsu, A.; Obinata, T.; Ohashi, K.; Mizuno, K.; Narumiya, S. Signaling from Rho to the actin cytoskeleton through protein kinases ROCK and LIM-kinase. Science 1999, 285, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Madaule, P.; Eda, M.; Watanabe, N.; Fujisawa, K.; Matsuoka, T.; Bito, H.; Ishizaki, T.; Narumiya, S. Role of citron kinase as a target of the small GTPase Rho in cytokinesis. Nature 1998, 394, 491–494. [Google Scholar] [CrossRef]
- Kimura, K.; Ito, M.; Amano, M.; Chihara, K.; Fukata, Y.; Nakafuku, M.; Yamamori, B.; Feng, J.; Nakano, T.; Okawa, K.; et al. Regulation of myosin phosphatase by Rho and Rho-associated kinase (Rho- kinase). Science 1996, 273, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.M.; Colomba, A.; Reymond, N.; Thomas, M.; Ridley, A.J. RhoB regulates cell migration through altered focal adhesion dynamics. Open Biol. 2012, 2. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.M.; Fruhwirth, G.; Ng, T.; Ridley, A.J. RhoA and RhoC have distinct roles in migration and invasion by acting through different targets. J. Cell Biol. 2011, 193, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mino, A.; Troeger, A.; Brendel, C.; Cantor, A.; Harris, C.; Ciuculescu, M.F.; Williams, D.A. Small GTPases RhoH participates in a multi-protein complex with the zinc finger protein kaiso that regulates both cytoskeletal structures and chemokine-induced T cells. Small GTPases 2018. [Google Scholar] [CrossRef] [Green Version]
- Bagrodia, S.; Cerione, R.A. PAK to the future. Trends Cell Biol. 1999, 9, 350–355. [Google Scholar] [CrossRef]
- Nagata, K.I.; Puls, A.; Futter, C.; Aspenstrom, P.; Schaefer, E.; Nakata, T.; Hirokawa, N.; Hall, A. The MAP kinase kinase kinase MLK2 co-localizes with activated JNK along microtubules and associates with kinesin superfamily motor KIF3. EMBO J. 1998, 17, 149–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teramoto, H.; Coso, O.A.; Miyata, H.; Igishi, T.; Miki, T.; Silvio Gutkind, J. Signaling from the small GTP-binding proteins Rac1 and Cdc42 to the c- Jun N-terminal kinase/stress-activated protein kinase pathway: A role for mixed lineage kinase 3/protein-tyrosine kinase 1, a novel member of the mixed lineage kinase family. J. Biol. Chem. 1996, 271, 27225–27228. [Google Scholar] [CrossRef] [Green Version]
- Miki, H.; Suetsugu, S.; Takenawa, T. WAVE, a novel WASP-family protein involved in actin reorganization induced by Rac. EMBO J. 1998, 17, 6932–6941. [Google Scholar] [CrossRef] [Green Version]
- Chou, M.M.; Blenis, J. The 70 kDa S6 kinase complexes with and is activated by the Rho family G proteins Cdc42 and Rac1. Cell 1996, 85, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Fukata, M.; Watanabe, T.; Noritake, J.; Nakagawa, M.; Yamaga, M.; Kuroda, S.; Matsuura, Y.; Iwamatsu, A.; Perez, F.; Kaibuchi, K. Rac1 and Cdc42 capture microtubules through IQGAP1 and CLIP-170. Cell 2002, 109, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Fukataf, M.; Kuroda, S.; Fujii, K.; Nakamura, T.; Shoji, I.; Matsuura, Y.; Okawa, K.; Iwamatsu, A.; Kikuchi, A.; Kaibuchi, K. Regulation of cross-linking of actin filament by IQGAP1, a target for Cdc42. J. Biol. Chem. 1997, 272, 29579–29583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanger, G.R.; Johnson, N.L.; Johnson, G.L. MEK kinases are regulated by EGF and selectively interact with Rac/Cdc42. EMBO J. 1997, 16, 4961–4972. [Google Scholar] [CrossRef] [Green Version]
- Van Aelst, L.; Joneson, T.; Bar-Sagi, D. Identification of a novel Rac1-interacting protein involved in membrane ruffling. EMBO J. 1996, 15, 3778–3786. [Google Scholar] [CrossRef]
- Diekmann, D.; Abo, A.; Johnston, C.; Segal, A.W.; Hall, A. Interaction of Rac with p67phox and regulation of phagocytic NADPH oxidase activity. Science 1994, 265, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Bokoch, G.M.; Vlahos, C.J.; Wang, Y.; Knaus, U.G.; Traynor-Kaplan, A.E. Rac GTPase interacts specifically with phosphatidylinositol 3-kinase. Biochem. J. 1996, 315, 775–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolias, K.F.; Cantley, L.C.; Carpenter, C.L. Rho family GTPases bind to phosphoinositide kinases. J. Biol. Chem. 1995, 270, 17656–17659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houssa, B.; De Widt, J.; Kranenburg, O.; Moolenaar, W.H.; Van Blitterswijk, W.J. Diacylglycerol kinase θ binds to and is negatively regulated by active RhoA. J. Biol. Chem. 1999, 274, 6820–6822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illenberger, D.; Schwald, F.; Pimmer, D.; Binder, W.; Maier, G.; Dietrich, A.; Gierschik, P. Stimulation of phospholipase C-β2 by the Rho GTPases Cdc42Hs and Rac1. EMBO J. 1998, 17, 6241–6249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Karnoub, A.E.; Palmby, T.R.; Lengyel, E.; Sondek, J.; Der, C.J. Rac1b, a tumor associated, constitutively active Rac1 splice variant, promotes cellular transformation. Oncogene 2004, 23, 9369–9380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arana, E.; Vehlow, A.; Harwood, N.E.; Vigorito, E.; Henderson, R.; Turner, M.; Tybulewicz, V.L.L.J.; Batista, F.D. Activation of the Small GTPase Rac2 via the B Cell Receptor Regulates B Cell Adhesion and Immunological-Synapse Formation. Immunity 2008, 28, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Hajdo-Milasinovic, A.; van der Kammen, R.A.; Moneva, Z.; Collard, J.G. Rac3 inhibits adhesion and differentiation of neuronal cells by modifying GIT1 downstream signaling. J. Cell Sci. 2009, 122, 2127–2136. [Google Scholar] [CrossRef] [Green Version]
- Vignal, E.; Blangy, A.; Martin, M.; Gauthier-Rouvière, C.; Fort, P. Kinectin Is a Key Effector of RhoG Microtubule-Dependent Cellular Activity. Mol. Cell. Biol. 2001, 21, 8022–8034. [Google Scholar] [CrossRef] [Green Version]
- Rohatgi, R.; Ma, L.; Miki, H.; Lopez, M.; Kirchhausen, T.; Takenawa, T.; Kirschner, M.W. The interaction between N-WASP and the Arp2/3 complex links Cdc42-dependent signals to actin assembly. Cell 1999, 97, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Abo, A.; Qu, J.; Cammarano, M.S.; Dan, C.; Fritsch, A.; Baud, V.; Belisle, B.; Minden, A. PAK4, a novel effector for Cdc42Hs, is implicated in the reorganization of the actin cytoskeleton and in the formation of filopodia. EMBO J. 1998, 17, 6527–6540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, T.; Chen, X.-Q.; Tan, I.; Manser, E.; Lim, L. Myotonic Dystrophy Kinase-Related Cdc42-Binding Kinase Acts as a Cdc42 Effector in Promoting Cytoskeletal Reorganization. Mol. Cell. Biol. 1998, 18, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Wilson, E.; Leszczynska, K.; Poulter, N.S.; Edelmann, F.; Salisbury, V.A.; Noy, P.J.; Bacon, A.; Rappoport, J.Z.; Heath, J.K.; Bicknell, R.; et al. RhoJ interacts with the GIT-PIX complex and regulates focal adhesion disassembly. J. Cell Sci. 2014, 127, 3039–3051. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, K.L.; Florke, R.R.; Reyes, S.S.; Tader, B.R.; Hamann, M.J. TCL/RhoJ plasma membrane localization and nucleotide exchange is coordinately regulated by amino acids within the N terminus and a distal loop region. J. Biol. Chem. 2016, 291, 23604–23617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aspenström, P. Atypical Rho GTPases RhoD and Rif integrate cytoskeletal dynamics and membrane trafficking. Biol. Chem. 2014, 395, 477–484. [Google Scholar] [CrossRef]
- Gad, A.K.B.; Aspenström, P. Rif proteins take to the RhoD: Rho GTPases at the crossroads of actin dynamics and membrane trafficking. Cell. Signal. 2010, 22, 183–189. [Google Scholar] [CrossRef]
- Li, Y.H.; Ghavampur, S.; Bondallaz, P.; Will, L.; Grenningloh, G.; Puschel, A.W. Rndl regulates axon extension by enhancing the microtubule destabilizing activity of SCG10. J. Biol. Chem. 2009, 284, 363–371. [Google Scholar] [CrossRef] [Green Version]
- Fujita, H.; Katoh, H.; Ishikawa, Y.; Mori, K.; Negishi, M. Rapostlin is a novel effector of Rnd2 GTPase inducing neurite branching. J. Biol. Chem. 2002, 277, 45428–45434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, H.; Harada, A.; Mori, K.; Negishi, M. Socius Is a Novel Rnd GTPase-Interacting Protein Involved in Disassembly of Actin Stress Fibers. Mol. Cell. Biol. 2002, 22, 2952–2964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riento, K.; Totty, N.; Villalonga, P.; Garg, R.; Guasch, R.; Ridley, A.J. RhoE function is regulated by ROCK I-mediated phosphorylation. EMBO J. 2005, 24, 1170–1180. [Google Scholar] [CrossRef] [Green Version]
- Riou, P.; Villalonga, P.; Ridley, A.J. Rnd proteins: Multifunctional regulators of the cytoskeleton and cell cycle progression. BioEssays 2010, 32, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, M.; Dvorsky, R.; Ahmadian, M.R. Deciphering the molecular and functional basis of Dbl family proteins: A novel systematic approach toward classification of selective activation of the Rho family proteins. J. Biol. Chem. 2013, 288, 4486–4500. [Google Scholar] [CrossRef] [Green Version]
- Wittinghofer, A.; Vetter, I.R. Structure-Function Relationships of the G Domain, a Canonical Switch Motif. Annu. Rev. Biochem. 2011, 80, 943–971. [Google Scholar] [CrossRef] [PubMed]
- Roberts, P.J.; Mitin, N.; Keller, P.J.; Chenette, E.J.; Madigan, J.P.; Currin, R.O.; Cox, A.D.; Wilson, O.; Kirschmeier, P.; Der, C.J. Rho family GTPase modification and dependence on CAAX motif-signaled posttranslational modification. J. Biol. Chem. 2008, 283, 25150–25163. [Google Scholar] [CrossRef] [Green Version]
- Fiegen, D.; Blumenstein, L.; Stege, P.; Vetter, I.R.; Ahmadian, M.R. Crystal structure of Rnd3/RhoE: Functional implications. FEBS Lett. 2002, 525, 100–104. [Google Scholar] [CrossRef] [Green Version]
- Fiegen, D.; Haeusler, L.C.; Blumenstein, L.; Herbrand, U.; Dvorsky, R.; Vetter, I.R.; Ahmadian, M.R. Alternative Splicing of Rac1 Generates Rac1b, a Self-activating GTPase. J. Biol. Chem. 2004, 279, 4743–4749. [Google Scholar] [CrossRef] [Green Version]
- Hemsath, L.; Dvorsky, R.; Fiegen, D.; Carlier, M.F.; Ahmadian, M.R. An electrostatic steering mechanism of Cdc42 recognition by Wiskott-Aldrich syndrome proteins. Mol. Cell 2005, 20, 313–324. [Google Scholar] [CrossRef]
- Kazemein Jasemi, N.; Herrmann, C.; Estirado, E.M.; Gremer, L.; Willbold, D.; Brunsveld, L.; Dvorsky, R.; Ahmadian, M. The intramolecular allostery of GRB2 governing its interaction with SOS1 is modulated by phosphotyrosine ligands. Biochem. J. 2021. [Google Scholar] [CrossRef]
- Ahmadian, M.R.; Jaiswal, M.; Fansa, E.K.; Dvorsky, R. New insight into the molecular switch mechanism of human Rho family proteins: Shifting a paradigm. Biol. Chem. 2013, 394, 89–95. [Google Scholar] [CrossRef]
- Winter-Vann, A.M.; Casey, P.J. Post-prenylation-processing enzymes as new targets in oncogenesis. Nat. Rev. Cancer 2005, 5, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Lang, P.; Gesbert, F.; Delespine-Carmagnat, M.; Stancou, R.; Pouchelet, M.; Bertoglio, J. Protein kinase A phosphorylation of RhoA mediates the morphological and functional effects of cyclic AMP in cytotoxic lymphocytes. EMBO J. 1996, 15, 510–519. [Google Scholar] [CrossRef]
- Forget, M.A.; Desrosiers, R.R.; Gingras, D.; Béliveau, R. Phosphorylation states of Cdc42 and RhoA regulate their interactions with Rho GDP dissociation inhibitor and their extraction from biological membranes. Biochem. J. 2002, 361, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding proteins of the Rho/Rac family: Regulation, effectors and functions in vivo. BioEssays 2007, 29, 356–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.; Weyand, M.; Lammers, M.; Ishizaki, T.; Ahmadian, M.R.; Wittinghofer, A. Structural and mechanistic insights into the interaction between Rho and mammalian Dia. Nature 2005, 435, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Troeger, A.; Chae, H.D.; Senturk, M.; Wood, J.; Williams, D.A. A unique carboxyl-Terminal insert domain in the hematopoietic-Specific, gtpase-Deficient rho gtpase rhoh regulates post-Translational processing. J. Biol. Chem. 2013, 288, 36451–36462. [Google Scholar] [CrossRef] [Green Version]
- Garavini, H.; Riento, K.; Phelan, J.P.; McAlister, M.S.B.; Ridley, A.J.; Keep, N.H. Crystal structure of the core domain of RhoE/Rnd3: A constitutively activated small G protein. Biochemistry 2002, 41, 6303–6310. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Shao, S.; Zhang, B.; Wang, H.; Liu, B. Role of Rho-specific guanine nucleotide dissociation inhibitor α regulation in cell migration. Acta Histochem. 2017, 119, 183–189. [Google Scholar] [CrossRef]
- Griner, E.M.; Theodorescu, D. The faces and friends of RhoGDI2. Cancer Metastasis Rev. 2012, 31, 519–528. [Google Scholar] [CrossRef] [PubMed]
- De León-Bautista, M.P.; Del Carmen Cardenas-Aguayo, M.; Casique-Aguirre, D.; Almaraz-Salinas, M.; Parraguirre-Martinez, S.; Olivo-Diaz, A.; Del Rocío Thompson-Bonilla, M.; Vargas, M. Immunological and functional characterization of RhoGDI3 and its molecular targets RhoG and RhoB in human pancreatic cancerous and normal cells. PLoS One 2016, 11, e0166370. [Google Scholar] [CrossRef]
- Brunet, N.; Morin, A.; Olofsson, B. RhoGDI-3 regulates RhoG and targets this protein to the Golgi complex through its unique N-terminal domain. Traffic 2002, 3, 342–358. [Google Scholar] [CrossRef]
- Garcia-Mata, R.; Boulter, E.; Burridge, K. The “invisible hand”: Regulation of RHO GTPases by RHOGDIs. Nat. Rev. Mol. Cell Biol. 2011, 12, 493–504. [Google Scholar] [CrossRef] [Green Version]
- DerMardirossian, C.; Bokoch, G.M. GDIs: Central regulatory molecules in Rho GTPase activation. Trends Cell Biol. 2005, 15, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Moissoglu, K.; Schwartz, M.A. Spatial and temporal control of Rho GTPase functions. Cell. Logist. 2014, 4, e943618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tnimov, Z.; Abankwa, D.; Alexandrov, K. RhoGDI facilitates geranylgeranyltransferase-I-mediated RhoA prenylation. Biochem. Biophys. Res. Commun. 2014, 452, 967–973. [Google Scholar] [CrossRef]
- Grizot, S.; Fauré, J.; Fieschi, F.; Vignais, P.V.; Dagher, M.C.; Pebay-Peyroula, E. Crystal structure of the Rac1 - RhoGDI complex involved in NADPH oxidase activation. Biochemistry 2001, 40, 10007–10013. [Google Scholar] [CrossRef] [PubMed]
- Scheffzek, K.; Stephan, I.; Jensen, O.N.; Illenberger, D.; Gierschik, P. The Rac-RhoGDI complex and the structural basis for the regulation of Rho proteins by RhoGDI. Nat. Struct. Biol. 2000, 7, 122–126. [Google Scholar] [CrossRef]
- Dransart, E.; Olofsson, B.; Cherfils, J. RhoGDIs revisited: Novel roles in Rho regulation. Traffic 2005, 6, 957–966. [Google Scholar] [CrossRef]
- Hoffman, G.R.; Nassar, N.; Cerione, R.A. Structure of the Rho family GTP-binding protein Cdc42 in complex with the multifunctional regulator RhoGDI. Cell 2000, 100, 345–356. [Google Scholar] [CrossRef] [Green Version]
- Longenecker, K.; Read, P.; Derewenda, U.; Dauter, Z.; Liu, X.; Garrard, S.; Walker, L.; Somlyo, A.V.; Nakamoto, R.K.; Somlyo, A.P.; et al. How RhoGDI binds Rho. Acta Crystallogr. Sect. D Biol. Crystallogr. 1999, 55, 1503–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosaddeghzadeh, N.; Kazemein Jasemi, N.S.; Majoleé, J.; Zhang, S.C.; Hordijk, P.L.; Dvorsky, R.; Ahmadian, M.R. New insights into the specificity of the RHO GTPase-GDI interaction: An electrostatic force mechanism. J. Biol. Chem. 2021. Under revision. [Google Scholar]
- Cherfils, J.; Chardin, P. GEFs: Structural basis for their activation of small GTP-binding proteins. Trends Biochem. Sci. 1999, 24, 306–311. [Google Scholar] [CrossRef]
- Guo, Z.; Ahmadian, M.R.; Goody, R.S. Guanine nucleotide exchange factors operate by a simple allosteric competitive mechanism. Biochemistry 2005, 44, 15423–15429. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, J.P.; Eccleston, J.F. Mechanism of nucleotide release from Rho by the GDP dissociation stimulator protein. Biochemistry 2000, 39, 11348–11359. [Google Scholar] [CrossRef]
- Heasman, S.J.; Ridley, A.J. Mammalian Rho GTPases: New insights into their functions from in vivo studies. Nat. Rev. Mol. Cell Biol. 2008, 9, 690–701. [Google Scholar] [CrossRef]
- Rossman, K.L.; Der, C.J.; Sondek, J. GEF means go: Turning on Rho GTPases with guanine nucleotide-exchange factors. Nat. Rev. Mol. Cell Biol. 2005, 6, 167–180. [Google Scholar] [CrossRef]
- Kunimura, K.; Uruno, T.; Fukui, Y. DOCK family proteins: Key players in immune surveillance mechanisms. Int. Immunol. 2020, 32, 5–15. [Google Scholar] [CrossRef]
- Cook, D.R.; Rossman, K.L.; Der, C.J. Rho guanine nucleotide exchange factors: Regulators of Rho GTPase activity in development and disease. Oncogene 2014, 33, 4021–4035. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.K.; Wheelock, R.H.P.; Aaronson, S.A.; Eva, A. Identification of the protein encoded by the human diffuse B-cell lymphoma (dbl) oncogene. Proc. Natl. Acad. Sci. USA 1986, 83, 8868–8872. [Google Scholar] [CrossRef] [Green Version]
- Hart, M.J.; Eva, A.; Evans, T.; Aaronson, S.A.; Cerione, R.A. Catalysis of guanine nucleotide exchange on the CDC42Hs protein by the dbloncogene product. Nature 1991, 354, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Aittaleb, M.; Boguth, C.A.; Tesmer, J.J.G. Structure and function of heterotrimeric G protein-regulated Rho guanine nucleotide exchange factors. Mol. Pharmacol. 2010, 77, 111–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, G.R.; Cerione, R.A. Signaling to the Rho GTPases: Networking with the DH domain. FEBS Lett. 2002, 513, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, M.; Dubey, B.N.; Koessmeier, K.T.; Gremer, L.; Ahmadian, M.R. Biochemical assays to characterize rho GTPases. Methods Mol. Biol. 2012, 827, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Viaud, J.; Gaits-Iacovoni, F.; Payrastre, B. Regulation of the DH-PH tandem of guanine nucleotide exchange factor for Rho GTPases by phosphoinositides. Adv. Biol. Regul. 2012, 52, 303–314. [Google Scholar] [CrossRef]
- Dubash, A.D.; Wennerberg, K.; García-Mata, R.; Menold, M.M.; Arthur, W.T.; Burridge, K. A novel role for Lsc/p115 RhoGEF and LARG in regulating RhoA activity downstream of adhesion to fibronectin. J. Cell Sci. 2007, 120, 3989–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bos, J.L.; Rehmann, H.; Wittinghofer, A. GEFs and GAPs: Critical Elements in the Control of Small G Proteins. Cell 2007, 129, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Vigil, D.; Cherfils, J.; Rossman, K.L.; Der, C.J. Ras superfamily GEFs and GAPs: Validated and tractable targets for cancer therapy? Nat. Rev. Cancer 2010, 10, 842–857. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, M.; Gremer, L.; Dvorsky, R.; Haeusler, L.C.; Cirstea, I.C.; Uhlenbrock, K.; Ahmadian, M.R. Mechanistic insights into specificity, activity, and regulatory elements of the regulator of G-protein signaling (RGS)-containing Rho-specific guanine nucleotide exchange factors (GEFs) p115, PDZ-RhoGEF (PRG), and leukemia-associated RhoGEF (LARG). J. Biol. Chem. 2011, 286, 18202–18212. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, H.; Eberstadt, M.; Schnuchel, A.; Olejniczak, E.T.; Meadows, R.P.; Schkeryantz, J.M.; Janowick, D.A.; Harlan, J.E.; Harris, E.A.S.; et al. NMR structure and mutagenesis of the N-terminal Dbl homology domain of the nucleotide exchange factor Trio. Cell 1998, 95, 269–277. [Google Scholar] [CrossRef] [Green Version]
- Erickson, J.W.; Cerione, R.A. Structural Elements, Mechanism, and Evolutionary Convergence of Rho Protein-Guanine Nucleotide Exchange Factor Complexes. Biochemistry 2004, 43, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Ferguson, K.M.; Abrams, C.S. Pleckstrin homology domains and the cytoskeleton. FEBS Lett. 2002, 513, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Haslam, R.J.; Koide, H.B.; Hemmings, B.A. Pleckstrin domain homology. Nature 1993, 363, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Haslam, R.J.; Rachubinski, R.A.; Harley, C.B. Molecular analysis of pleckstrin: The major protein kinase c substrate of platelets. J. Cell. Biochem. 1989, 40, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Dinitto, J.; Lambright, D. Membrane and juxtamembrane targeting by PH and PTB domains. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2006, 1761, 850–867. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Luby-Phelps, K.; Das, B.; Shu, X.; Xia, Y.; Mosteller, R.D.; Krishna, U.M.; Falck, J.R.; White, M.A.; Broek, D. Role of substrates and products of PI 3-kinase in regulating activation of Rac-related guanosine triphosphatases by Vav. Science 1998, 279, 558–560. [Google Scholar] [CrossRef]
- Nimnual, A.S. Coupling of Ras and Rac Guanosine Triphosphatases Through the Ras Exchanger Sos. Science 1998, 279, 560–563. [Google Scholar] [CrossRef]
- Lemmon, M.A. Pleckstrin homology domains: Not just for phosphoinositides. Biochem. Soc. Trans. 2004, 32, 707–711. [Google Scholar] [CrossRef]
- Shi, L. Dock protein family in brain development and neurological disease. Commun. Integr. Biol. 2013, 6, e26839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goicoechea, S.M.; Awadia, S.; Garcia-Mata, R. I’m coming to GEF you: Regulation of RhoGEFs during cell migration. Cell Adhes. Migr. 2014, 8, 535–549. [Google Scholar] [CrossRef]
- Chang, L.; Yang, J.; Jo, C.H.; Boland, A.; Zhang, Z.; McLaughlin, S.H.; Abu-Thuraia, A.; Killoran, R.C.; Smith, M.J.; Côté, J.F.; et al. Structure of the DOCK2−ELMO1 complex provides insights into regulation of the auto-inhibited state. Nat. Commun. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Laurin, M.; Côté, J.F. Insights into the biological functions of Dock family guanine nucleotide exchange factors. Genes Dev. 2014, 28, 533–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Côté, J.F.; Vuori, K. Identification of an evolutionary conserved superfamily of DOCK180-related proteins with guanine nucleotide exchange activity. J. Cell Sci. 2002, 115, 4901–4913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, Y.; Yamauchi, J. Cellular signaling of Dock family proteins in neural function. Cell. Signal. 2010, 22, 175–182. [Google Scholar] [CrossRef]
- Li, H.; Yang, L.; Fu, H.; Yan, J.; Wang, Y.; Guo, H.; Hao, X.; Xu, X.; Jin, T.; Zhang, N. ARTICLE Association between Gai2 and ELMO1/Dock180 connects chemokine signalling with Rac activation and metastasis. Nat. Commun. 2013. [Google Scholar] [CrossRef] [Green Version]
- Cimino, P.J.; Sokal, I.; Leverenz, J.; Fukui, Y.; Montine, T.J. DOCK2 is a microglial specific regulator of central nervous system innate immunity found in normal and Alzheimer’s disease brain. Am. J. Pathol. 2009, 175, 1622–1630. [Google Scholar] [CrossRef] [Green Version]
- Cimino, P.J.; Yang, Y.; Li, X.; Hemingway, J.F.; Cherne, M.K.; Khademi, S.B.; Fukui, Y.; Montine, K.S.; Montine, T.J.; Keene, C.D. Ablation of the microglial protein DOCK2 reduces amyloid burden in a mouse model of Alzheimer’s disease. Exp. Mol. Pathol. 2013, 94, 366–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, E.; Tohme, M.; Hedayat, M.; Leick, M.; Kumari, S.; Ramesh, N.; Massaad, M.J.; Ullas, S.; Azcutia, V.; Goodnow, C.C.; et al. A DOCK8-WIP-WASp complex links T cell receptors to the actin cytoskeleton. J. Clin. Investig. 2016, 126, 3837–3851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eberth, A.; Dvorsky, R.; Becker, C.F.W.; Beste, A.; Goody, R.S.; Ahmadian, M.R. Monitoring the real-time kinetics of the hydrolysis reaction of guanine nucleotide-binding proteins. Biol. Chem. 2005, 386, 1105–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidyk, N.J.; Cerione, R.A. Understanding the catalytic mechanism of GTPase-activating proteins: Demonstration of the importance of switch domain stabilization in the stimulation of GTP hydrolysis. Biochemistry 2002, 41, 15644–15653. [Google Scholar] [CrossRef] [PubMed]
- Amin, E.; Jaiswal, M.; Derewenda, U.; Reis, K.; Nouri, K.; Koessmeier, K.T.; Aspenström, P.; Somlyo, A.V.; Dvorsky, R.; Ahmadian, M.R. Deciphering the molecular and functional basis of RHOGAP family proteins: A systematic approach toward selective inactivation of RHO family proteins. J. Biol. Chem. 2016, 291, 20353–20371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheffzek, K.; Ahmadian, M.R. GTPase activating proteins: Structural and functional insights 18 years after discovery. Cell. Mol. Life Sci. 2005, 62, 3014–3038. [Google Scholar] [CrossRef] [PubMed]
- Scheffzek, K.; Ahmadian, M.R.; Kabsch, W.; Wiesmüller, L.; Lautwein, A.; Schmitz, F.; Wittinghofer, A. The Ras-RasGAP complex: Structural basis for GTPase activation and its loss in oncogenic ras mutants. Science 1997, 277, 333–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, M.; Dvorsky, R.; Amin, E.; Risse, S.L.; Fansa, E.K.; Zhang, S.C.; Taha, M.S.; Gauhar, A.R.; Nakhaei-Rad, S.; Kordes, C.; et al. Functional cross-talk between ras and rho pathways: A ras-specific gtpase-activating protein (p120RasGAP) competitively inhibits the rhogap activity of deleted in liver cancer (DLC) tumor suppressor by masking the catalytic arginine finger. J. Biol. Chem. 2014, 289, 6839–6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.Y.; Guan, M.; Vigil, D.; Der, C.J.; Lowy, D.R.; Popescu, N.C. p120Ras-GAP binds the DLC1 Rho-GAP tumor suppressor protein and inhibits its RhoA GTPase and growth-suppressing activities. Oncogene 2009, 28, 1401–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, M.D.; Self, A.J.; Van Oers, C.; Hall, A. Identification of distinct cytoplasmic targets for ras/R-ras and rho regulatory proteins. J. Biol. Chem. 1989, 264, 10–13. [Google Scholar] [CrossRef]
- Eberth, A.; Lundmark, R.; Gremer, L.; Dvorsky, R.; Koessmeier, K.T.; McMahon, H.T.; Ahmadian, M.R. A BAR domain-mediated autoinhibitory mechanism for RhoGAPs of the GRAF family. Biochem. J. 2009, 417, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Minoshima, Y.; Kawashima, T.; Hirose, K.; Tonozuka, Y.; Kawajiri, A.; Bao, Y.C.; Deng, X.; Tatsuka, M.; Narumiya, S.; May, W.S.; et al. Phosphorylation by Aurora B converts MgcRacGAP to a RhoGAP during cytokinesis. Dev. Cell 2003, 4, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Ligeti, E.; Dagher, M.C.; Hernandez, S.E.; Koleske, A.J.; Settleman, J. Phospholipids Can Switch the GTPase Substrate Preference of a GTPase-activating Protein. J. Biol. Chem. 2004, 279, 5055–5058. [Google Scholar] [CrossRef] [Green Version]
- Ahmadian, M.R.; Mittal, R.; Hall, A.; Wittinghofer, A. Aluminium fluoride associates with the small guanine nucleotide binding proteins. FEBS Lett. 1997, 408, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.L.; Eccleston, J.F.; Lowe, P.N. The conserved arginine in Rho-GTPase-activating protein is essential for efficient catalysis but not for complex formation with Rho·GDP and aluminum fluoride. Biochemistry 1999, 38, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Trahey, M.; Mccormick, F. A cytoplasmic protein stimulates normal N-ras p21 GTPase, but does not affect oncogenic mutants. Science 1987, 238, 542–545. [Google Scholar] [CrossRef]
- Diekmann, D.; Brill, S.; Garrett, M.D.; Totty, N.; Hsuan, J.; Monfries, C.; Hall, C.; Lim, L.; Hall, A. Bcr encodes a GTPase-activating protein for p21rac. Nature 1991, 351, 400–402. [Google Scholar] [CrossRef]
- Lancaster, C.A.; Taylor-Harris, P.M.; Self, A.J.; Brill, S.; Van Erp, H.E.; Hall, A. Characterization of rhoGAP. A GTPase-activating protein for rho-related small GTPases. J. Biol. Chem. 1994, 269, 1137–1142. [Google Scholar] [CrossRef]
- Rittinger, K.; Taylor, W.R.; Smerdon, S.J.; Gamblin, S.J. Support for shared ancestry of GAPs. Nature 1998, 392, 448–449. [Google Scholar] [CrossRef]
- Scheffzek, K.; Ahmadian, M.R.; Wittinghofer, A. GTPase-activating proteins: Helping hands to complement an active site. Trends Biochem. Sci. 1998, 23, 257–262. [Google Scholar] [CrossRef]
- Scheffzek, K.; Lautwein, A.; Kabscht, W.; Ahmadian, M.R.; Wittinghofer, A. Crystal structure of the GTPase-activating domain of human p120GAP and implications for the interaction with Ras. Nature 1996, 384, 591–596. [Google Scholar] [CrossRef]
- Barrett, T.; Xiao, B.; Dodson, E.J.; Dodson, G.; Ludbrook, S.B.; Nurmahomed, K.; Gamblin, S.J.; Musacchio, A.; Smerdon, S.J.; Eccleston, J.F. The structure of the GTPase-activating domain from p50rhoGAP. Nature 1997, 385, 458–461. [Google Scholar] [CrossRef]
- Graham, D.L.; Lowe, P.N.; Grime, G.W.; Marsh, M.; Rittinger, K.; Smerdon, S.J.; Gamblin, S.J.; Eccleston, J.F. MgF3- as a Transition State Analog of Phosphoryl Transfer. Chem. Biol. 2002, 9, 375–381. [Google Scholar] [CrossRef]
- Vetter, I.R.; Wittinghofer, A. The guanine nucleotide-binding switch in three dimensions. Science 2001, 294, 1299–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittinger, K.; Walker, P.A.; Eccleston, J.F.; Nurmahomed, K.; Owen, D.; Laue, E.; Gamblin, S.J.; Smerdon, S.J. Crystal structure of a small G protein in complex with the GTPase- activating protein rhoGAP. Nature 1997, 388, 693–697. [Google Scholar] [CrossRef]
- Rittinger, K.; Walker, P.A.; Eccleston, J.F.; Smerdon, S.J.; Gamblin, S.J. Structure at 1.65 Å of RhoA and its GTPase-activating protein in complex with a transition-state analogue. Nature 1997, 389, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, A.B.; Majerus, P.W. Properties of type II inositol polyphosphate 5-phosphatase. J. Biol. Chem. 1995, 270, 9370–9377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernards, A. GAPs galore! A survey of putative Ras superfamily GTPase activating proteins in man and Drosophila. Biochim. Biophys. Acta Rev. Cancer 2003, 1603, 47–82. [Google Scholar] [CrossRef]
- Amin, E.; Dubey, B.N.; Zhang, S.C.; Gremer, L.; Dvorsky, R.; Moll, J.M.; Taha, M.S.; Nagel-Steger, L.; Piekorz, R.P.; Somlyo, A.V.; et al. Rho-kinase: Regulation, (dys)function, and inhibition. Biol. Chem. 2013, 394, 1399–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narumiya, S.; Tanji, M.; Ishizaki, T. Rho signaling, ROCK and mDia1, in transformation, metastasis and invasion. Cancer Metastasis Rev. 2009, 28, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.S.; Manser, E. PAK and other Rho-associated kinases - Effectors with surprisingly diverse mechanisms of regulation. Biochem. J. 2005, 386, 201–214. [Google Scholar] [CrossRef]
- Hedman, A.C.; Smith, J.M.; Sacks, D.B. The biology of IQGAP proteins: Beyond the cytoskeleton. EMBO Rep. 2015, 16, 427–446. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.A.; Wang, M.J.; Chi, C.W.; Wu, C.W.; Chen, J.Y. Rho/Rhotekin-mediated NF-κB activation confers resistance to apoptosis. Oncogene 2004, 23, 8731–8742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maesaki, R.; Ihara, K.; Shimizu, T.; Kuroda, S.; Kaibuchi, K.; Hakoshima, T. The structural basis of Rho effector recognition revealed by the crystal structure of human RhoA complexed with the effector domain of PKN/PRK1. Mol. Cell 1999, 4, 793–803. [Google Scholar] [CrossRef]
- Dvorsky, R.; Blumenstein, L.; Vetter, I.R.; Ahmadian, M.R. Structural Insights into the Interaction of ROCKI with the Switch Regions of RhoA. J. Biol. Chem. 2004, 279, 7098–7104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Manan, N.; Aghazadeh, B.; Liu, G.A.; Majumdar, A.; Ouerfelli, O.; Simlnovitch, K.A.; Rosen, M.K. Structure of Cdc42 in complex with the GTPase-binding domain of the “Wiskott-Aldrich syndrome” protein. Nature 1999, 399, 379–383. [Google Scholar] [CrossRef]
- Mott, H.R.; Owen, D.; Nietlispach, D.; Lowe, P.N.; Manser, E.; Lim, L.; Laue, E.D. Structure of the small G protein Cdc42 bound to the GTpasebinding domain of ACK. Nature 1999, 399, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Gizachew, D.; Guo, W.; Chohan, K.K.; Sutcliffe, M.J.; Oswald, R.E. Structure of the complex of Cdc42Hs with a peptide derived from P-21 activated kinase. Biochemistry 2000, 39, 3963–3971. [Google Scholar] [CrossRef]
- Morreale, A.; Venkatesan, M.; Mott, H.R.; Owen, D.; Nietlispach, D.; Lowe, P.N.; Laue, E.D. Structure of Cdc42 bound to the GTPase binding domain of PAK. Nat. Struct. Biol. 2000, 7, 384–388. [Google Scholar] [CrossRef]
- Garrard, S.M.; Capaldo, C.T.; Gao, L.; Rosen, M.K.; Macara, I.G.; Tomchick, D.R. Structure of Cdc42 in a complex with the GTPase-binding domain of the cell polarity protein, Par6. EMBO J. 2003, 22, 1125–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, S.; Krumbach, O.H.F.; Pantaleoni, F.; Coppola, S.; Amin, E.; Pannone, L.; Nouri, K.; Farina, L.; Dvorsky, R.; Lepri, F.; et al. Functional Dysregulation of CDC42 Causes Diverse Developmental Phenotypes. Am. J. Hum. Genet. 2018, 102, 309–320. [Google Scholar] [CrossRef] [Green Version]
- Lapouge, K.; Smith, S.J.M.; Walker, P.A.; Gamblin, S.J.; Smerdon, S.J.; Rittinger, K. Structure of the TPR domain of p67(phox) in complex with Rac·GTP. Mol. Cell 2000, 6, 899–907. [Google Scholar] [CrossRef]
- Tarricone, C.; Xiao, B.; Justin, N.; Walker, P.A.; Rittinger, K.; Gamblin, S.J.; Smerdon, S.J. The structural basis of Arfaptin-mediated cross-talk between Rac and Arf signalling pathways. Nature 2001, 411, 215–219. [Google Scholar] [CrossRef]
- Cherfils, J. Structural mimicry of DH domains by Arfaptin suggests a model for the recognition of Rac-GDP by its guanine nucleotide exchange factors. FEBS Lett. 2001, 507, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Diebold, B.A.; Bokoch, G.M. Molecular basis for Rac2 regulation of phagocyte NADPH oxidase. Nat. Immunol. 2001, 2, 211–215. [Google Scholar] [CrossRef]
- Hoffman, G.R.; Cerione, R.A. Rac inserts its way into the immune response. Nat. Immunol. 2001, 2, 194–196. [Google Scholar] [CrossRef] [PubMed]
- Blumenstein, L.; Ahmadian, M.R. Models of the cooperative mechanism for Rho effector recognition: Implications for RhoA-mediated effector activation. J. Biol. Chem. 2004, 279, 53419–53426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A. Rho GTpases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, C.D.; Ridley, A.J. Rho GTPase signaling complexes in cell migration and invasion. J. Cell Biol. 2018, 217, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Guan, X.; Dong, C.; Jiao, Z. Rho GTPases and related signaling complexes in cell migration and invasion. Exp. Cell Res. 2020, 388. [Google Scholar] [CrossRef]
- Hall, A. Rho family GTPases. Biochem. Soc. Trans. 2012, 40, 1378–1382. [Google Scholar] [CrossRef] [Green Version]
- Schaks, M.; Giannone, G.; Rottner, K. Actin dynamics in cell migration. Essays Biochem. 2019, 63, 483–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devreotes, P.; Horwitz, A.R. Signaling networks that regulate cell migration. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zegers, M.M.; Friedl, P. Rho GTPases in collective cell migration. Small GTPases 2014, 5. [Google Scholar] [CrossRef]
- Sadok, A.; Marshall, C.J. Rho gtpases masters of cell migration. Small GTPases 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Raftopoulou, M.; Hall, A. Cell migration: Rho GTPases lead the way. Dev. Biol. 2004, 265, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Miralles, F.; Posern, G.; Zaromytidou, A.I.; Treisman, R. Actin dynamics control SRF activity by regulation of its coactivator MAL. Cell 2003, 113, 329–342. [Google Scholar] [CrossRef] [Green Version]
- Hordijk, P.L. Regulation of NADPH oxidases: The role of Rac proteins. Circ. Res. 2006, 98, 453–462. [Google Scholar] [CrossRef]
- Voncken, J.W.; van Schaick, H.; Kaartinen, V.; Deemer, K.; Coates, T.; Landing, B.; Pattengale, P.; Dorseuil, O.; Bokoch, G.M.; Groffen, J.; et al. Increased neutrophil respiratory burst in bcr-null mutants. Cell 1995, 80, 719–728. [Google Scholar] [CrossRef] [Green Version]
- Lyon, A.S.; Peeples, W.B.; Rosen, M.K. A framework for understanding the functions of biomolecular condensates across scales. Nat. Rev. Mol. Cell Biol. 2021, 22, 215–235. [Google Scholar] [CrossRef] [PubMed]
- Araki, Y.; Hong, I.; Gamache, T.R.; Ju, S.; Collado-Torres, L.; Shin, J.H.; Huganir, R.L. Syngap isoforms differentially regulate synaptic plasticity and dendritic development. Elife 2020, 9, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Pudewell, S.; Wittich, C.; Kazemein Jasemi, N.S.; Bazgir, F.; Ahmadian, M.R. Accessory proteins of the RAS-MAPK pathway: Moving from the side line to the front line. Commun. Biol. 2021, 4, 696. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Banjade, S.; Cheng, H.C.; Kim, S.; Chen, B.; Guo, L.; Llaguno, M.; Hollingsworth, J.V.; King, D.S.; Banani, S.F.; et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 2012, 483, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Case, L.B.; Zhang, X.; Ditlev, J.A.; Rosen, M.K. Stoichiometry controls activity of phase-separated clusters of actin signaling proteins. Science 2019, 363, 1093–1097. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mosaddeghzadeh, N.; Ahmadian, M.R. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells 2021, 10, 1831. https://doi.org/10.3390/cells10071831
Mosaddeghzadeh N, Ahmadian MR. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells. 2021; 10(7):1831. https://doi.org/10.3390/cells10071831
Chicago/Turabian StyleMosaddeghzadeh, Niloufar, and Mohammad Reza Ahmadian. 2021. "The RHO Family GTPases: Mechanisms of Regulation and Signaling" Cells 10, no. 7: 1831. https://doi.org/10.3390/cells10071831
APA StyleMosaddeghzadeh, N., & Ahmadian, M. R. (2021). The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells, 10(7), 1831. https://doi.org/10.3390/cells10071831