
Dysregulation of miRNAs Targeting the IGF-1R Pathway in Pancreatic Ductal Adenocarcinoma


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Evaluation of miRNAs
2.3. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foucher, E.D.; Ghigo, C.; Chouaib, S.; Galon, J.; Iovanna, J.; Olive, D. Pancreatic Ductal Adenocarcinoma: A strong imbalance of good and bad immunological cops in the tumor microenvironment. Front. Immunol. 2018, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Rawla, P.; Sunkara, T.; Gaduputi, V. Epidemiology of pancreatic cancer: Global trends, etiology and risk factors. World J. Oncol. 2019, 10, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Fanta, P.T.; Lowy, A.M. Adenocarcinoma of the Pancreas. In Yamada’ s Textbook of Gastroenterology; John Wiley & Sons, Ltd.: Oxford, UK, 2015; pp. 1761–1781. [Google Scholar]
- Kim, J.Y.; Hong, S.-M. Precursor lesions of pancreatic cancer. Oncol. Res. Treat. 2018, 41, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Mutgan, A.C.; Besikcioglu, H.E.; Wang, S.; Friess, H.; Ceyhan, G.O.; Demir, I.E. Insulin/IGF-driven cancer cell-stroma crosstalk as a novel therapeutic target in pancreatic cancer. Mol. Cancer 2018, 17, 66. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Yin, Z.; Tao, K.; Wang, G.; Gao, J. Function of insulin-like growth factor 1 receptor in cancer resistance to chemotherapy. Oncol. Lett. 2018, 15, 41–47. [Google Scholar] [CrossRef]
- Kopantzev, E.P.; Kopantseva, M.R.; Grankina, E.V.; Mikaelyan, A.; Egorov, V.I.; Sverdlov, E.D. Activation of IGF/IGF-IR signaling pathway fails to induce epithelial-mesenchymal transition in pancreatic cancer cells. Pancreatology 2019, 19, 390–396. [Google Scholar] [CrossRef]
- Hirakawa, T.; Yashiro, M.; Murata, A.; Hirata, K.; Kimura, K.; Amano, R.; Yamada, N.; Nakata, B.; Hirakawa, K. IGF-1 receptor and IGF binding protein-3 might predict prognosis of patients with resectable pancreatic cancer. BMC Cancer 2013, 13, 392. [Google Scholar] [CrossRef] [Green Version]
- Camblin, A.J.; Pace, E.A.; Adams, S.; Curley, M.D.; Rimkunas, V.; Nie, L.; Tan, G.; Bloom, T.; Iadevaia, S.; Baum, J.; et al. Dual Inhibition of IGF-1R and ErbB3 Enhances the Activity of Gemcitabine and Nab-Paclitaxel in Preclinical Models of Pancreatic Cancer. Clin. Cancer Res. 2018, 24, 2873–2885. [Google Scholar] [CrossRef] [Green Version]
- Pereira, N.P.; Corrêa, J.R. Pancreatic cancer: Treatment approaches and trends. J. Cancer Metastasis Treat. 2018, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Słotwiński, R.; Lech, G.; Słotwińska, S.M. MicroRNAs in pancreatic cancer diagnosis and therapy. Cent. J. Immunol. 2018, 43, 314–324. [Google Scholar] [CrossRef] [Green Version]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—MicroRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in control of gene expression: An overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Li, J.; Chi, D.; Sahnoune, I.; Calin, S.; Girnita, L.; Calin, G.A. Non-Coding RNAs in IGF-1R signaling regulation: The underlying pathophysiological link between diabetes and cancer. Cells 2019, 8, 1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nweke, E.E.; Brand, M. Downregulation of the let-7 family of microRNAs may promote insulin receptor/insulin-like growth factor signalling pathways in pancreatic ductal adenocarcinoma. Oncol. Lett. 2020, 20, 2613–2620. [Google Scholar] [CrossRef]
- Huang, J.S.; Egger, M.E.; Grizzle, W.E.; McNally, L.R. MicroRNA-100 regulates IGF1-receptor expression in metastatic pancreatic cancer cells. Biotech. Histochem. 2013, 88, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, C.; George Priya Doss, C.; Bandyopadhyay, S. miRNAs in insulin resistance and diabetes-associated pancreatic cancer: The “minute and miracle” molecule moving as a monitor in the “genomic galaxy”. Curr. Drug Targets 2013, 14, 1110–1117. [Google Scholar] [CrossRef]
- Farhana, L.; Dawson, M.I.; Murshed, F.; Das, J.K.; Rishi, A.K.; Fontana, J.A. Upregulation of miR-150* and miR-630 induces apoptosis in pancreatic cancer cells by targeting IGF-1R. PLoS ONE 2013, 8, e61015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, T.; Nakata, B.; Amano, R.; Kimura, K.; Shimizu, S.; Ohira, G.; Yamada, N.; Ohira, M.; Hirakawa, K. HER3 overexpression as an independent indicator of poor prognosis for patients with curatively resected pancreatic cancer. Oncology 2011, 81, 192–198. [Google Scholar] [CrossRef]
- Komoto, M.; Nakata, B.; Nishii, T.; Kawajiri, H.; Shinto, O.; Amano, R.; Yamada, N.; Yashiro, M.; Hirakawa, K. In vitro and in vivo evidence that a combination of lapatinib plus S-1 is a promising treatment for pancreatic cancer. Cancer Sci. 2010, 101, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Nair, P.N.; De Armond, D.T.; Adamo, M.L.; Strodel, W.E.; Freeman, J.W. Aberrant expression and activation of insulin-like growth factor-1 receptor (IGF-1R) are mediated by an induction of IGF-1R promoter activity and stabilization of IGF-1R mRNA and contributes to growth factor independence and increased survival of the panc. Oncogene 2001, 20, 8203–8214. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Rong, L.; Zhao, J.; Lin, R.; Li, S. MicroRNA-539 inhibits cell proliferation, colony formation and invasion in pancreatic ductal adenocarcinoma by directly targeting IGF-1R. Mol. Med. Rep. 2018, 18, 1804–1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, C.; da Silva, A.; Morales-Oyarvide, V.; Dias Costa, A.; Kozak, M.M.; Dunne, R.F.; Rubinson, D.A.; Perez, K.; Masugi, Y.; Hamada, T.; et al. Insulin-like growth factor-1 receptor expression and disease recurrence and survival in patients with resected pancreatic ductal adenocarcinoma. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1586–1595. [Google Scholar] [CrossRef] [PubMed]
- Valsecchi, M.E.; McDonald, M.; Brody, J.R.; Hyslop, T.; Freydin, B.; Yeo, C.J.; Solomides, C.; Peiper, S.C.; Witkiewicz, A.K. Epidermal growth factor receptor and insulinlike growth factor 1 receptor expression predict poor survival in pancreatic ductal adenocarcinoma. Cancer 2012, 118, 3484–3493. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Sharon, E. IGF-1R as an anti-cancer target--trials and tribulations. Chin. J. Cancer 2013, 32, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.Y.; Calin, G.A. MicroRNAs as therapeutic targets in human cancers. Wiley Interdiscip. Rev. RNA 2014, 5, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Henson, B.J.; Bhattacharjee, S.; O’Dee, D.M.; Feingold, E.; Gollin, S.M. Decreased expression of miR-125b and miR-100 in oral cancer cells contributes to malignancy. Genes. Chromosomes Cancer 2009, 48, 569–582. [Google Scholar] [CrossRef] [Green Version]
- Motawi, T.K.; Rizk, S.M.; Ibrahim, T.M.; Ibrahim, I.A.-R. Circulating microRNAs, miR-92a, miR-100 and miR-143, as non-invasive biomarkers for bladder cancer diagnosis. Cell Biochem. Funct. 2016, 34, 142–148. [Google Scholar] [CrossRef]
- Wang, M.; Ren, D.; Guo, W.; Wang, Z.; Huang, S.; Du, H.; Song, L.; Peng, X. Loss of miR-100 enhances migration, invasion, epithelial-mesenchymal transition and stemness properties in prostate cancer cells through targeting Argonaute 2. Int. J. Oncol. 2014, 45, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; He, T.; Yang, L.; Yang, G.; Chen, Y.; Zhang, X. The role of miR-100 in regulating apoptosis of breast cancer cells. Sci. Rep. 2015, 5, 11650. [Google Scholar] [CrossRef] [Green Version]
- Assmann, T.S.; Recamonde-Mendoza, M.; De Souza, B.M.; Crispim, D. MicroRNA expression profiles and type 1 diabetes mellitus: Systematic review and bioinformatic analysis. Endocr. Connect. 2017, 6, 773–790. [Google Scholar] [CrossRef] [Green Version]
- Pek, S.L.T.; Sum, C.F.; Lin, M.X.; Cheng, A.K.S.; Wong, M.T.K.; Lim, S.C.; Tavintharan, S. Circulating and visceral adipose miR-100 is down-regulated in patients with obesity and Type 2 diabetes. Mol. Cell. Endocrinol. 2016, 427, 112–123. [Google Scholar] [CrossRef]
- Gong, R.; Jiang, Y. Non-coding RNAs in Pancreatic Ductal Adenocarcinoma. Front. Oncol. 2020, 10, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Ebeling, M.C.; Zaman, M.S.; Sikander, M.; Yallapu, M.M.; Chauhan, N.; Yacoubian, A.M.; Behrman, S.W.; Zafar, N.; Kumar, D.; et al. MicroRNA-145 targets MUC13 and suppresses growth and invasion of pancreatic cancer. Oncotarget 2014, 5, 7599–7609. [Google Scholar] [CrossRef] [PubMed]
- Namkung, J.; Kwon, W.; Choi, Y.; Yi, S.G.; Han, S.; Kang, M.J.; Kim, S.-W.; Park, T.; Jang, J.-Y. Molecular subtypes of pancreatic cancer based on miRNA expression profiles have independent prognostic value. J. Gastroenterol. Hepatol. 2016, 31, 1160–1167. [Google Scholar] [CrossRef]
- Ding, Y.; Cao, F.; Sun, H.; Wang, Y.; Liu, S.; Wu, Y.; Cui, Q.; Mei, W.; Li, F. Exosomes derived from human umbilical cord mesenchymal stromal cells deliver exogenous miR-145-5p to inhibit pancreatic ductal adenocarcinoma progression. Cancer Lett. 2019, 442, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, M.; Safarzadeh, A.; Beyranvand, F.; Ahmadpour, F.; Hajiasgharzadeh, K.; Baghbanzadeh, A.; Baradaran, B. The potential role of miR-29 in health and cancer diagnosis, prognosis, and therapy. J. Cell. Physiol. 2019, 234, 19280–19297. [Google Scholar] [CrossRef]
- Tréhoux, S.; Lahdaoui, F.; Delpu, Y.; Renaud, F.; Leteurtre, E.; Torrisani, J.; Jonckheere, N.; Van Seuningen, I. Micro-RNAs miR-29a and miR-330-5p function as tumor suppressors by targeting the MUC1 mucin in pancreatic cancer cells. Biochim. Biophys. Acta 2015, 1853, 2392–2403. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.J.; Willy, J.A.; Quirin, K.A.; Wek, R.C.; Korc, M.; Yin, X.-M.; Kota, J. Novel role of miR-29a in pancreatic cancer autophagy and its therapeutic potential. Oncotarget 2016, 7, 71635–71650. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-H.; Huang, J.; Wu, C.-R.; Huang, L.-Y.; Cui, J.; Xing, Z.-Z.; Zhao, C.-Y. Downregulation of miR-29b targets DNMT3b to suppress cellular apoptosis and enhance proliferation in pancreatic cancer. Mol. Med. Rep. 2018, 17, 2113–2120. [Google Scholar] [CrossRef]
- Li, J.; Chan, M.C.; Yu, Y.; Bei, Y.; Chen, P.; Zhou, Q.; Cheng, L.; Chen, L.; Ziegler, O.; Rowe, G.C.; et al. miR-29b contributes to multiple types of muscle atrophy. Nat. Commun. 2017, 8, 15201. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Li, J.; Chen, Z.; Li, X.; Zheng, S.; Yi, D.; Zhong, A.; Chen, J. miR-29c suppresses pancreatic cancer liver metastasis in an orthotopic implantation model in nude mice and affects survival in pancreatic cancer patients. Carcinogenesis 2015, 36, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Maurer, B.; Stanczyk, J.; Jüngel, A.; Akhmetshina, A.; Trenkmann, M.; Brock, M.; Kowal-Bielecka, O.; Gay, R.E.; Michel, B.A.; Distler, J.H.W.; et al. MicroRNA-29, a key regulator of collagen expression in systemic sclerosis. Arthritis Rheum. 2010, 62, 1733–1743. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.J.; Nabinger, S.C.; Vega, Z.; Sahu, S.S.; Alluri, R.K.; Abdul-Sater, Z.; Yu, Z.; Gore, J.; Nalepa, G.; Saxena, R.; et al. Pathophysiological role of microRNA-29 in pancreatic cancer stroma. Sci. Rep. 2015, 5, 11450. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wu, Y.; He, Y. Silencing circRNA LRP6 down-regulates PRMT1 to improve the streptozocin-induced pancreatic β-cell injury and insulin secretion by sponging miR-9-5p. J. Bioenerg. Biomembr. 2021, 53, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, B.; Ren, H.; Chen, W. miR-9-5p inhibits pancreatic cancer cell proliferation, invasion and glutamine metabolism by targeting GOT1. Biochem. Biophys. Res. Commun. 2019, 509, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Liu, F.; Liu, Y.; Shao, M.; Li, X.; Qin, G. Linc00210 enhances the malignancy of thyroid cancer cells by modulating miR-195-5p/IGF1R/Akt axis. J. Cell. Physiol. 2020, 235, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Sun, C.; Hu, X.; Zhan, H.; Zou, H.; Feng, Y.; Qiu, F.; Zhang, S.; Wu, L.; Zhang, B. MicroRNA-195 suppresses the progression of pancreatic cancer by targeting DCLK1. Cell. Physiol. Biochem. 2017, 44, 1867–1881. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xia, T.; Cao, M.; Zhang, P.; Shi, G.; Chen, L.; Zhang, J.; Yin, J.; Wu, P.; Cai, B.; et al. LncRNA BANCR promotes pancreatic cancer tumorigenesis via modulating MiR-195-5p/Wnt/β-catenin signaling pathway. Technol. Cancer Res. Treat. 2019, 18, 1533033819887962. [Google Scholar] [CrossRef]
- Zhou, W.-Y.; Zhang, M.-M.; Liu, C.; Kang, Y.; Wang, J.-O.; Yang, X.-H. Long noncoding RNA LINC00473 drives the progression of pancreatic cancer via upregulating programmed death-ligand 1 by sponging microRNA-195-5p. J. Cell. Physiol. 2019, 234, 23176–23189. [Google Scholar] [CrossRef] [PubMed]
- Zeöld, A.; Sándor, G.O.; Kiss, A.; Soós, A.Á.; Tölgyes, T.; Bursics, A.; Szűcs, Á.; Harsányi, L.; Kittel, Á.; Gézsi, A.; et al. Shared extracellular vesicle miRNA profiles of matched ductal pancreatic adenocarcinoma organoids and blood plasma samples show the power of organoid technology. Cell. Mol. Life Sci. 2021, 78, 3005–3020. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Features | PDAC (n = 38) | CTRL (n = 5 P, n = 6 NT) | Significance |
|---|---|---|---|
| Age (mean ± SD) | 65.08 ± 9.93 | CTRL = 58.00 ± 13.93 | PDAC vs. CTRL, p = 0.064 |
| P = 65.80 ± 13.36 | PDAC vs. P, p = 0.884 | ||
| NT = 51.50 ± 11.55 | PDAC vs. NT, p = 0.004 | ||
| Sex (%F) | 55.30% | CTRL = 36.4% | PDAC vs. CTRL, χ = 1.219, p = 0.269 |
| P = 40% | PDAC vs. P, χ = 0.414, p = 0.520 | ||
| NT = 33.3% | PDAC vs. NT, χ = 0.999, p = 0.318 | ||
| Smokers (%) | 39.50% | CTRL = 27.3% | PDAC vs. CTRL, χ = 0.546, p = 0.460 |
| P = 0% | PDAC vs. P, χ = 3.031, p = 0.082 | ||
| NT = 50% | PDAC vs. NT, χ = 0.238, p = 0.626 | ||
| Coffee consumers (%) | 39.50% | CTRL = 63.7% | PDAC vs. CTRL, χ = 2.013, p = 0.156 |
| P = 60% | PDAC vs. P, χ = 0.765, p = 0.382 | ||
| NT = 66.7% | PDAC vs. NT, χ = 1.562, p = 0.211 | ||
| Alcohol consumers (%) | 27.30% | CTRL = 15.8 % | PDAC vs. CTRL, χ = 0.750, p = 0.386 |
| P = 40% | PDAC vs. P, χ = 1.710, p = 0.191 | ||
| NT = 16.7% | PDAC vs. NT, χ = 0.003, p = 0.956 |
| PDAC (n = 38) | ||||||
|---|---|---|---|---|---|---|
| Localization | Clinical Stage | Lymph Node Invasion | Metastasis | Diabetes | Jaundice | Family History of PDAC |
| Head = 23 | T2 = 7 | 18 | 14 | 10 | 15 | 2 |
| T3 = 6 | ||||||
| T4 = 10 | ||||||
| Neck = 3 | T2 = 1 | 2 | 1 | 2 | 0 | 0 |
| T3 = 2 | ||||||
| Body = 6 | T2 = 2 | 5 | 5 | 3 | 0 | 0 |
| T3 = 2 | ||||||
| T4 = 2 | ||||||
| Body-Tail = 4 | T2 = 2 | 2 | 2 | 2 | 0 | 1 |
| T3 = 1 | ||||||
| T4 = 1 | ||||||
| Tail = 2 | T3 = 1 | 2 | 2 | 0 | 0 | 0 |
| T4 = 1 | ||||||
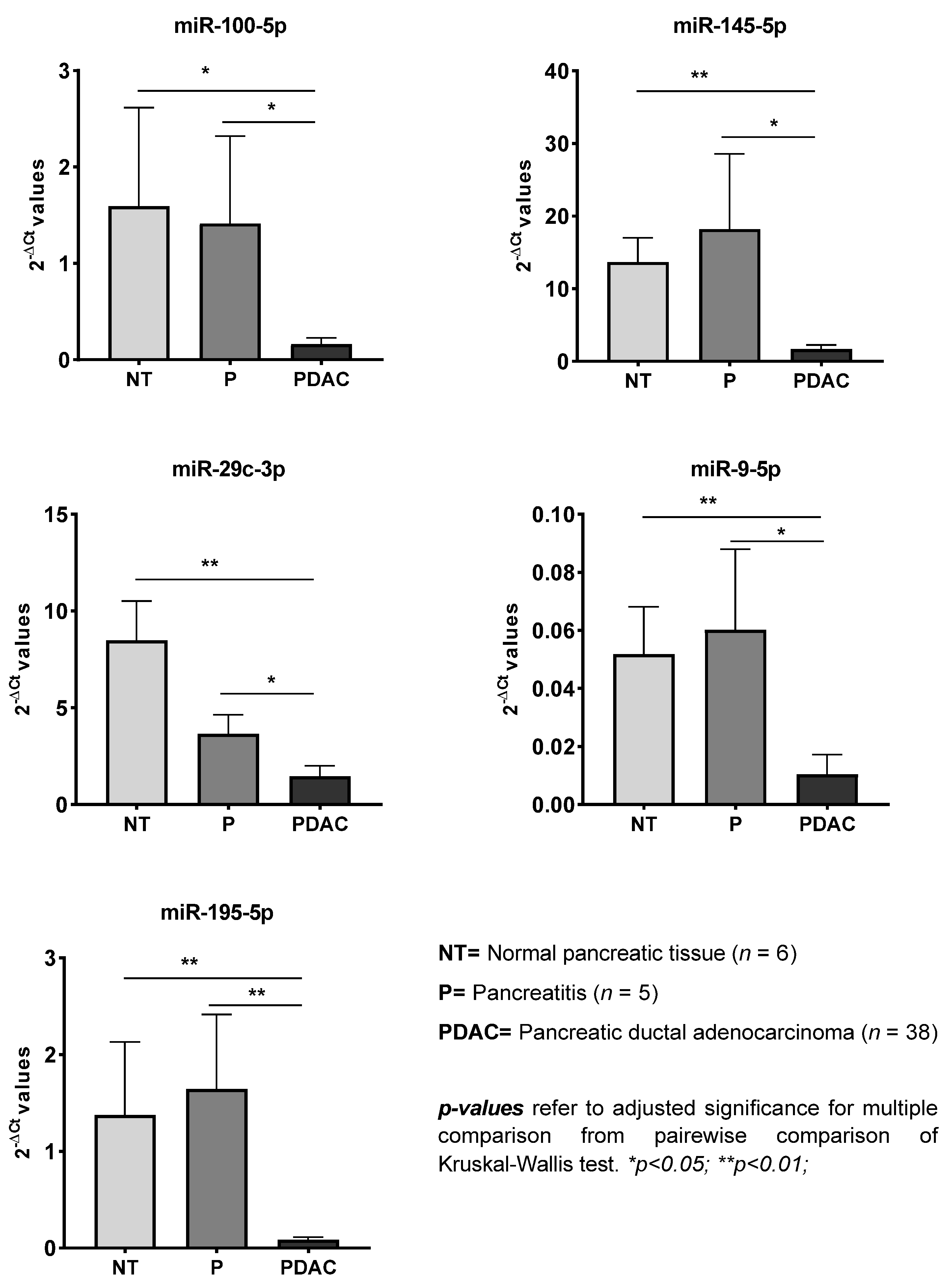
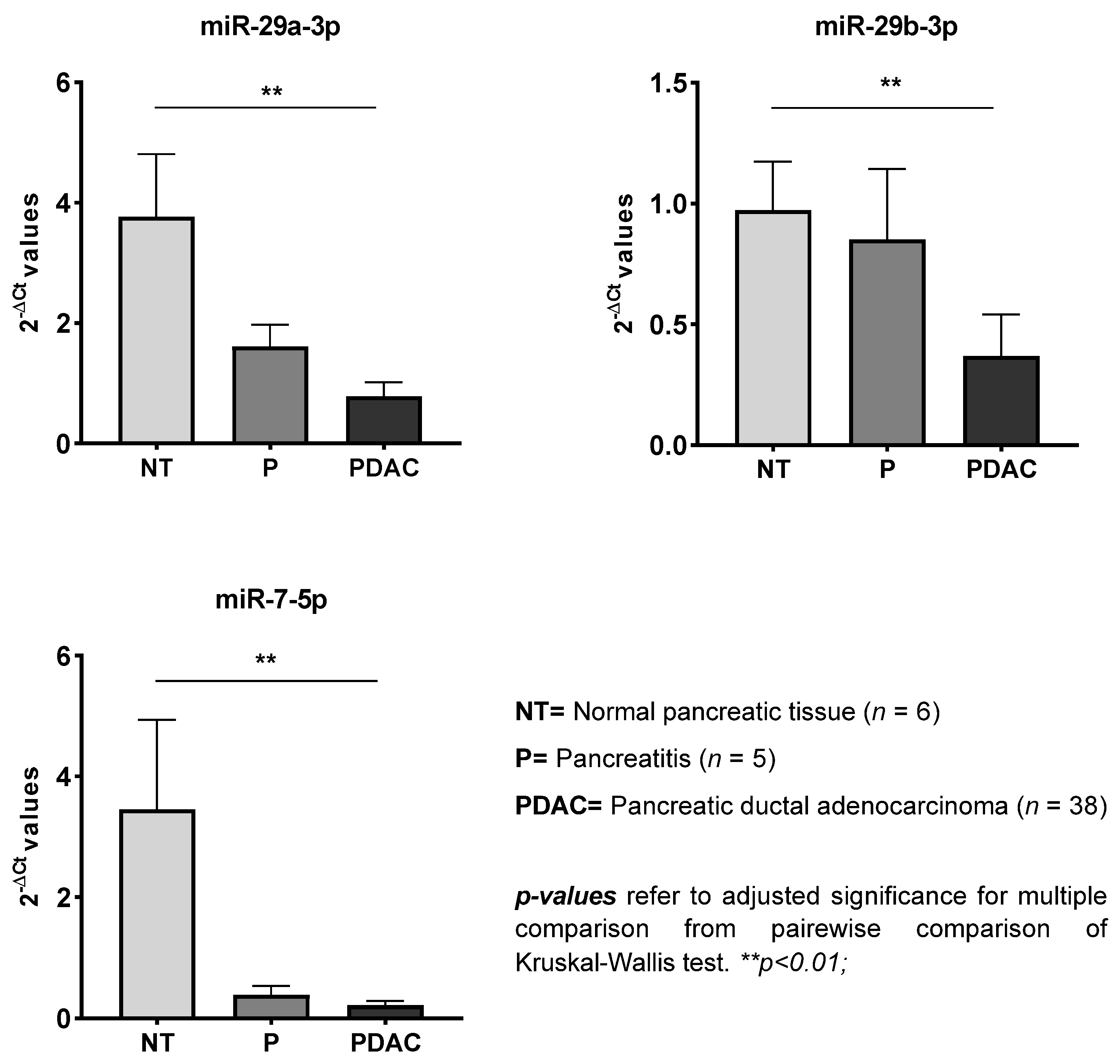
| miRNA | Significance | PDAC vs. NT | PDAC vs. P | P vs. NT | |||
|---|---|---|---|---|---|---|---|
| p-Value | FR | p-Value | FR | p-Value | FR | p-Value | |
| miR-let-7a-5p | 0.006 | / | ns | −19.01 | 0.005 | ns | ns |
| miR-let-7b-5p | 0.002 | / | ns | −19.70 | 0.006 | ns | ns |
| miR-let-7c-5p | 0.004 | / | ns | −12.58 | 0.005 | ns | ns |
| miR-let-7d-5p | 0.012 | / | ns | −13.48 | 0.009 | ns | ns |
| miR-let-7g-5p | 0.034 | / | ns | −12.64 | 0.044 | ns | ns |
| miR-100-5p | 0.001 | −9.83 | 0.012 | −8.72 | 0.035 | ns | ns |
| miR-let-7f-5p | 0.015 | / | ns | −12.44 | 0.015 | ns | ns |
| miR-126-3p | 0.002 | / | ns | −29.16 | 0.007 | ns | ns |
| miR-145-5p | <0.001 | −8.15 | 0.003 | −10.84 | 0.019 | ns | ns |
| miR-150-5p | 0.002 | / | ns | −23.70 | 0.002 | ns | ns |
| miR-19a-3p | 0.007 | / | ns | −10.13 | 0.011 | ns | ns |
| miR-21-5p | 0.012 | / | ns | −2.96 | 0.025 | ns | ns |
| miR-29a-3p | 0.001 | −4.82 | 0.003 | / | ns | ns | ns |
| miR-29b-3p | 0.001 | −2.64 | 0.006 | / | ns | ns | ns |
| miR-29c-3p | <0.001 | −5.78 | 0.001 | −2.49 | 0.04 | ns | ns |
| miR-7-5p | 0.001 | −16.41 | 0.001 | / | ns | ns | ns |
| miR-9-5p | <0.001 | −4.96 | 0.002 | −5.76 | 0.013 | ns | ns |
| miR-let-7e-5p | 0.006 | / | ns | −5.52 | 0.015 | ns | ns |
| miR-195-5p | <0.001 | −16.09 | 0.001 | −19.26 | 0.004 | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobre, M.; Herlea, V.; Vlăduţ, C.; Ciocîrlan, M.; Balaban, V.D.; Constantinescu, G.; Diculescu, M.; Milanesi, E. Dysregulation of miRNAs Targeting the IGF-1R Pathway in Pancreatic Ductal Adenocarcinoma. Cells 2021, 10, 1856. https://doi.org/10.3390/cells10081856
Dobre M, Herlea V, Vlăduţ C, Ciocîrlan M, Balaban VD, Constantinescu G, Diculescu M, Milanesi E. Dysregulation of miRNAs Targeting the IGF-1R Pathway in Pancreatic Ductal Adenocarcinoma. Cells. 2021; 10(8):1856. https://doi.org/10.3390/cells10081856
Chicago/Turabian StyleDobre, Maria, Vlad Herlea, Cătălina Vlăduţ, Mihai Ciocîrlan, Vasile Daniel Balaban, Gabriel Constantinescu, Mircea Diculescu, and Elena Milanesi. 2021. "Dysregulation of miRNAs Targeting the IGF-1R Pathway in Pancreatic Ductal Adenocarcinoma" Cells 10, no. 8: 1856. https://doi.org/10.3390/cells10081856
APA StyleDobre, M., Herlea, V., Vlăduţ, C., Ciocîrlan, M., Balaban, V. D., Constantinescu, G., Diculescu, M., & Milanesi, E. (2021). Dysregulation of miRNAs Targeting the IGF-1R Pathway in Pancreatic Ductal Adenocarcinoma. Cells, 10(8), 1856. https://doi.org/10.3390/cells10081856