
Alterations in Dendritic Spine Maturation and Neurite Development Mediated by FAM19A1



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Handling
2.2. Dendritic Spine Analysis In Vivo
2.3. Immunohistochemical Analysis of Excitatory Synapses
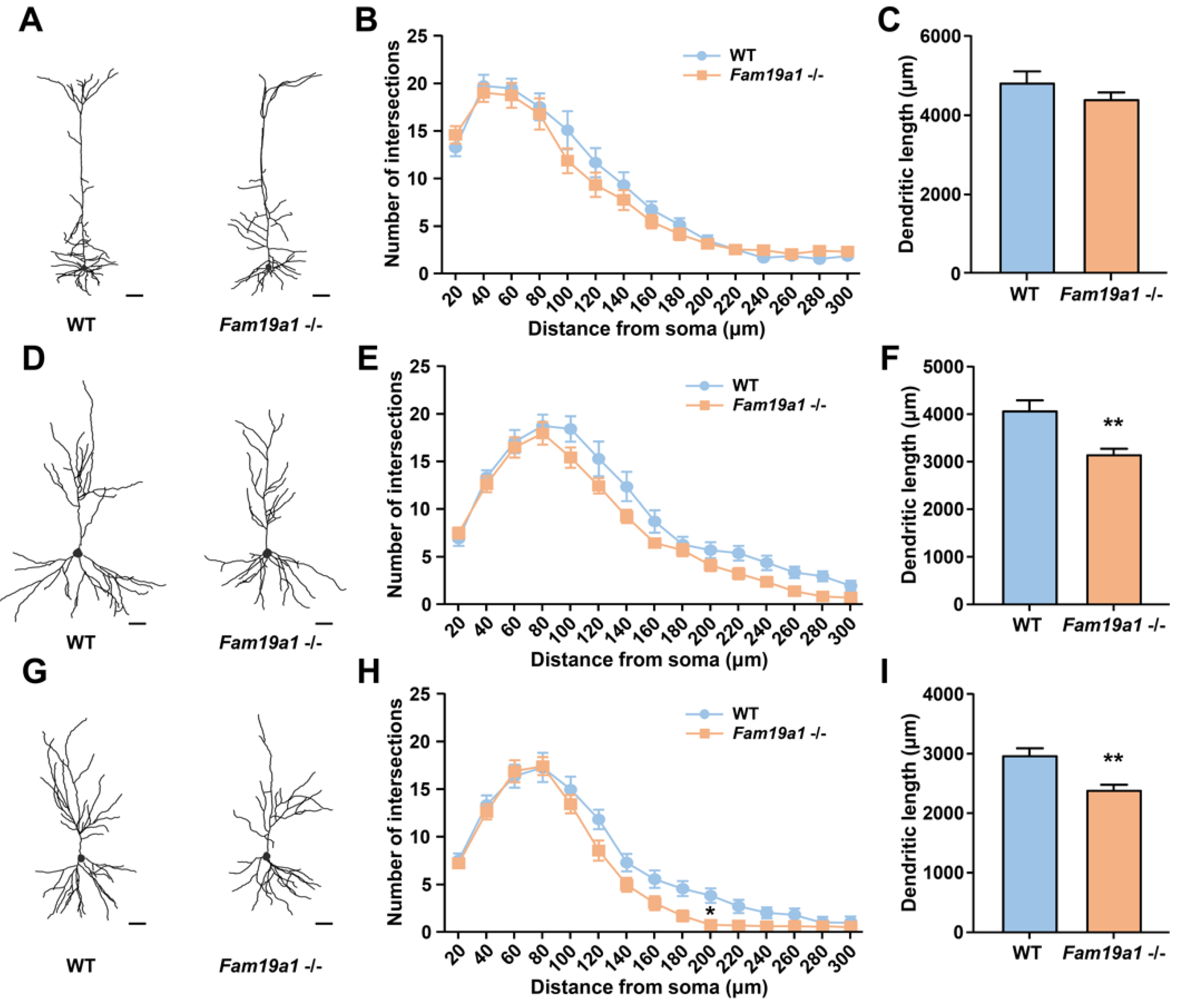
2.4. Dendritic Arborization Analysis In Vivo
2.5. Primary Neuronal Culture
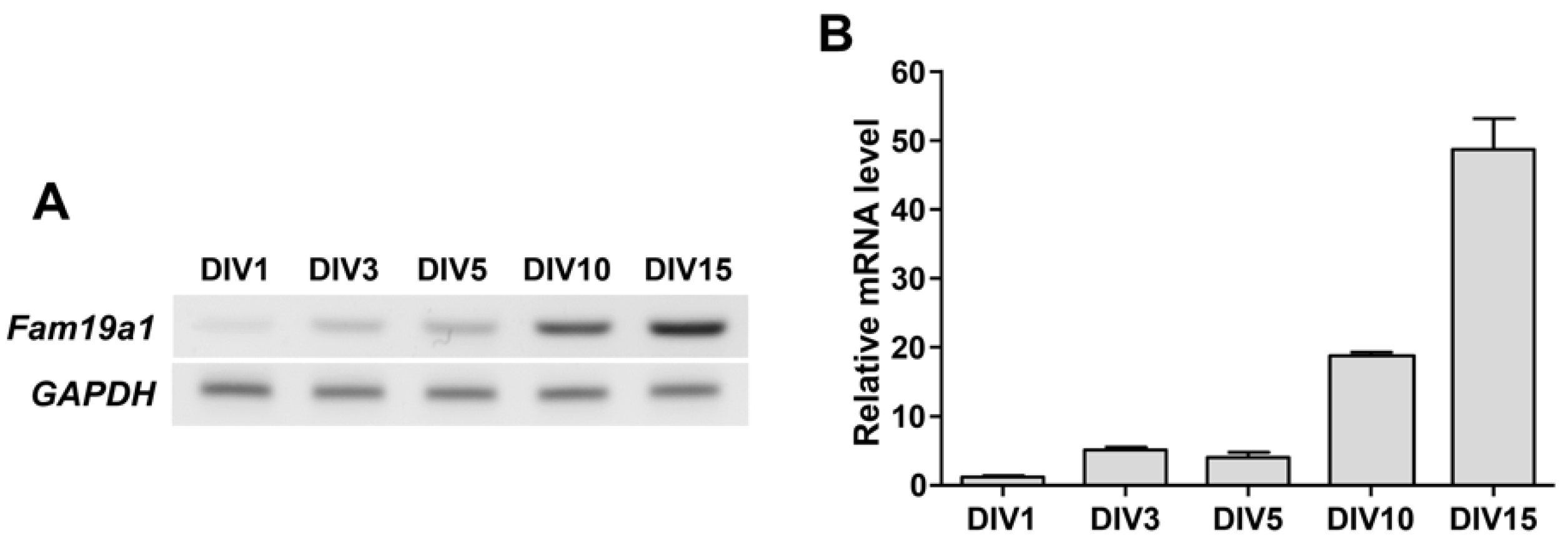
2.6. RNA Isolation and Quantitative PCR
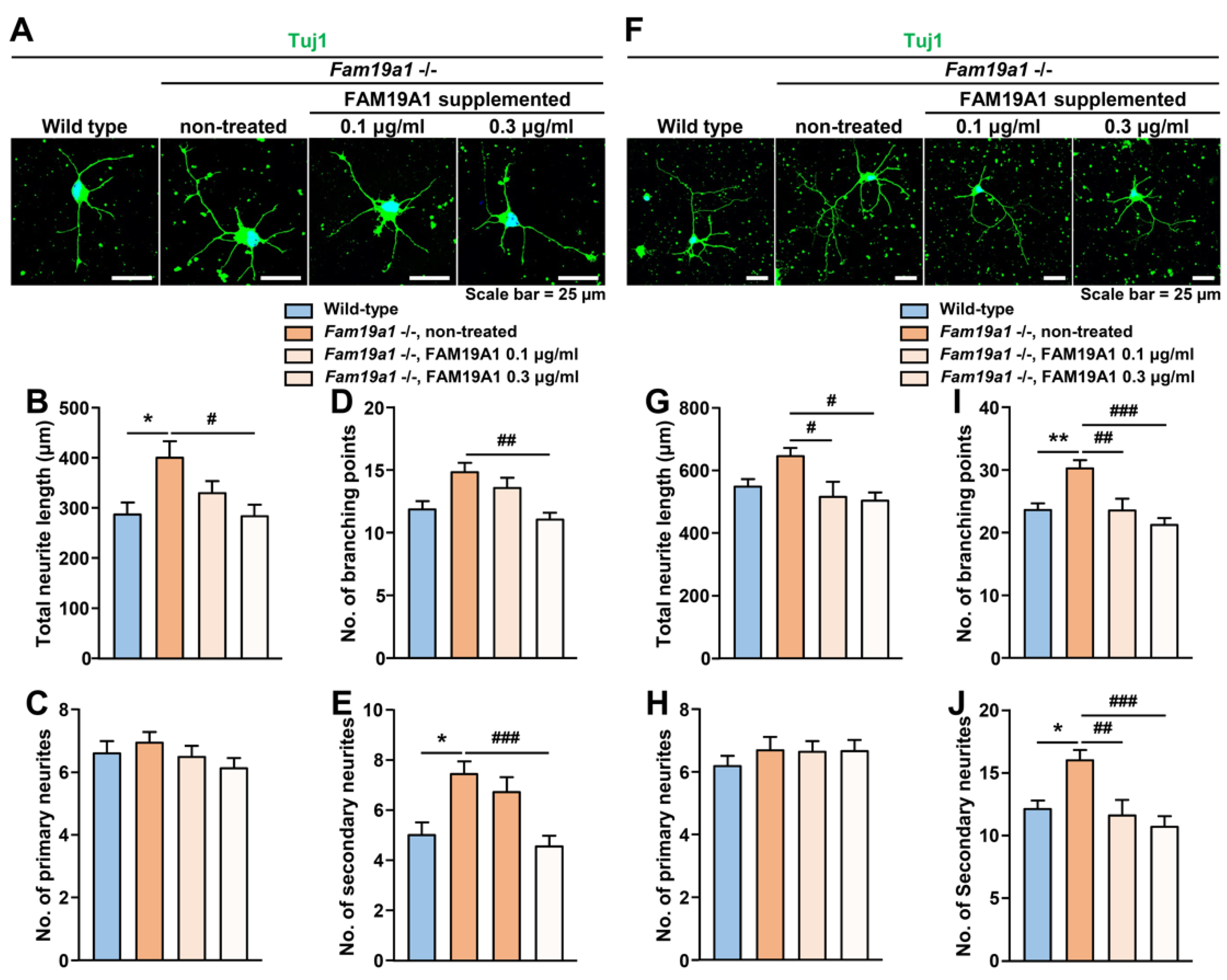
2.7. Recombinant His-Tagged FAM19A1 Protein Generation and Purification
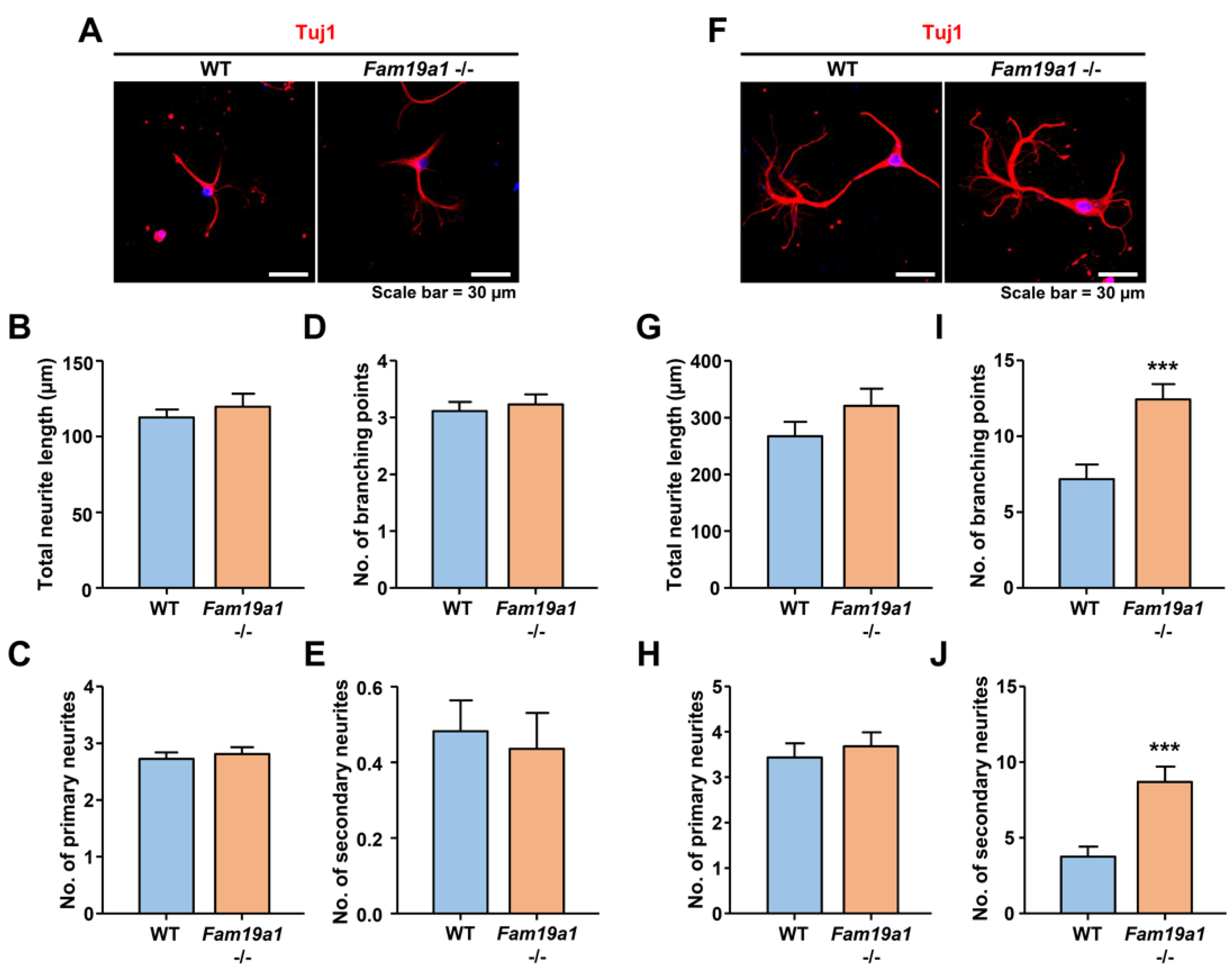
2.8. Immunocytochemical Analysis for Neuronal Morphology
2.9. Dendritic Spine Analysis In Vitro
2.10. Statistical Analysis
3. Results
3.1. Dendritic Spine Abnormalities in Cortical Neurons of Adult Fam19a1-Deficient Mice
3.2. Fewer Mature Spines in Fam19a1 −/− Mice during Postnatal Neurodevelopment
3.3. Dendritic Morphology of Neurons in Fam19a1-Expressing Brain Regions
3.4. Fam19a1 Gene Expression in Primary Neurons In Vitro
3.5. Increased Dendritic Complexity in Fam19a1-Deficient Primary Neurons
3.6. Reduced Dendritic Arborization in Primary Neurons upon Treatment with Recombinant FAM19A1
3.7. Increase in Neurite Complexity in Fam19a1-Deficient Primary Neurons Was Reversed by Recombinant FAM19A1 Supplementation
3.8. Morphological Analysis of Dendritic Spines in Primary Neurons Supplemented with FAM19A1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sernagor, E.; Chabrol, F.; Bony, G.; Cancedda, L. GABAergic control of neurite outgrowth and remodeling during development and adult neurogenesis: General rules and differences in diverse systems. Front. Cell. Neurosci. 2010, 4. [Google Scholar] [CrossRef] [Green Version]
- Azari, H.; Reynolds, B.A. In Vitro Models for Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a021279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, M.; Hayashi, Y. Structural plasticity of dendritic spines. Curr. Opin. Neurobiol. 2012, 22, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Dotti, C.G.; Sullivan, C.A.; Banker, G.A. The establishment of polarity by hippocampal neurons in culture. J. Neurosci. 1988, 8, 1454. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Liu, S.-S.; Zhan, M.-Y.; Luo, J.-H.; Zhu, L.-J. Interleukin-2 Enhances Dendritic Development and Spinogenesis in Cultured Hippocampal Neurons. Anat. Rec. 2010, 293, 1017–1023. [Google Scholar] [CrossRef]
- Behnke, J.; Cheedalla, A.; Bhatt, V.; Bhat, M.; Teng, S.; Palmieri, A.; Windon, C.C.; Thakker-Varia, S.; Alder, J. Neuropeptide VGF Promotes Maturation of Hippocampal Dendrites That Is Reduced by Single Nucleotide Polymorphisms. Int. J. Mol. Sci. 2017, 18, 612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, L.K.; Cheng, H.-J. Axon pruning: An essential step underlying the developmental plasticity of neuronal connections. Philos. Trans. R Soc. Lond. B Biol. Sci. 2006, 361, 1531–1544. [Google Scholar] [CrossRef] [Green Version]
- Bonhoeffer, T.; Yuste, R. Spine motility. Phenomenology, mechanisms, and function. Neuron 2002, 35, 1019–1027. [Google Scholar] [CrossRef] [Green Version]
- Ethell, I.M.; Pasquale, E.B. Molecular mechanisms of dendritic spine development and remodeling. Prog. Neurobiol. 2005, 75, 161–205. [Google Scholar] [CrossRef] [PubMed]
- Bian, W.-J.; Miao, W.-Y.; He, S.-J.; Qiu, Z.; Yu, X. Coordinated Spine Pruning and Maturation Mediated by Inter-Spine Competition for Cadherin/Catenin Complexes. Cell 2015, 162, 808–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burbach, J.P. What are neuropeptides? Methods Mol. Biol. 2011, 789, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Deverman, B.E.; Patterson, P.H. Cytokines and CNS development. Neuron 2009, 64, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Prieto, G.A.; Cotman, C.W. Cytokines and cytokine networks target neurons to modulate long-term potentiation. Cytokine Growth Factor Rev. 2017, 34, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Trettel, F.; Di Castro, M.A.; Limatola, C. Chemokines: Key Molecules that Orchestrate Communication among Neurons, Microglia and Astrocytes to Preserve Brain Function. Neuroscience 2020, 439, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Lobentanzer, S.; Hanin, G.; Klein, J.; Soreq, H. Integrative Transcriptomics Reveals Sexually Dimorphic Control of the Cholinergic/Neurokine Interface in Schizophrenia and Bipolar Disorder. Cell Rep. 2019, 29, 764–777.e765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatkova, M.; Reichova, A.; Bacova, Z.; Strbak, V.; Kiss, A.; Bakos, J. Neurite Outgrowth Stimulated by Oxytocin Is Modulated by Inhibition of the Calcium Voltage-Gated Channels. Cell. Mol. Neurobiol. 2018, 38, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Björnström, K.; Turina, D.; Strid, T.; Sundqvist, T.; Eintrei, C. Orexin A inhibits propofol-induced neurite retraction by a phospholipase D/protein kinase Cε-dependent mechanism in neurons. PLoS ONE 2014, 9, e97129. [Google Scholar] [CrossRef] [Green Version]
- Mody, M.; Cao, Y.; Cui, Z.; Tay, K.-Y.; Shyong, A.; Shimizu, E.; Pham, K.; Schultz, P.; Welsh, D.; Tsien, J.Z. Genome-wide gene expression profiles of the developing mouse hippocampus. Proc. Natl. Acad. Sci. USA 2001, 98, 8862. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.K.; Jiang, Y.; Chen, S.; Xia, Y.; Maciejewski, D.; McNamara, R.K.; Streit, W.J.; Salafranca, M.N.; Adhikari, S.; Thompson, D.A.; et al. Role for neuronally derived fractalkine in mediating interactions between neurons and CX3CR1-expressing microglia. Proc. Natl. Acad. Sci. USA 1998, 95, 10896–10901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milior, G.; Lecours, C.; Samson, L.; Bisht, K.; Poggini, S.; Pagani, F.; Deflorio, C.; Lauro, C.; Alboni, S.; Limatola, C.; et al. Fractalkine receptor deficiency impairs microglial and neuronal responsiveness to chronic stress. Brain Behav. Immun. 2016, 55, 114–125. [Google Scholar] [CrossRef]
- Paolicelli, R.C.; Bolasco, G.; Pagani, F.; Maggi, L.; Scianni, M.; Panzanelli, P.; Giustetto, M.; Ferreira, T.A.; Guiducci, E.; Dumas, L.; et al. Synaptic Pruning by Microglia Is Necessary for Normal Brain Development. Science 2011, 333, 1456. [Google Scholar] [CrossRef] [Green Version]
- Han, K.A.; Woo, D.; Kim, S.; Choii, G.; Jeon, S.; Won, S.Y.; Kim, H.M.; Heo, W.D.; Um, J.W.; Ko, J. Neurotrophin-3 Regulates Synapse Development by Modulating TrkC-PTPσ Synaptic Adhesion and Intracellular Signaling Pathways. J. Neurosci. 2016, 36, 4816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tom Tang, Y.; Emtage, P.; Funk, W.D.; Hu, T.; Arterburn, M.; Park, E.E.; Rupp, F. TAFA: A novel secreted family with conserved cysteine residues and restricted expression in the brain. Genomics 2004, 83, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.J.; Ha, N.; Cho, E.B.; Yun, S.; Kim, H.; Hwang, J.-I.; Seong, J.Y. The unique expression profile of FAM19A1 in the mouse brain and its association with hyperactivity, long-term memory and fear acquisition. Sci. Rep. 2020, 10, 3969. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.; Liu, L.; Terrillion, C.E.; Karuppagounder, S.S.; Cisternas, P.; Lay, M.; Martinelli, D.C.; Aja, S.; Dong, X.; Pletnikov, M.V.; et al. FAM19A1, a brain-enriched and metabolically responsive neurokine, regulates food intake patterns and mouse behaviors. FASEB J. 2019, 33, 14734–14747. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Chen, D.; Zhang, Y.; Bai, Y.; Huang, S.; Zheng, D.; Liang, W.; She, S.; Peng, X.; Wang, P.; et al. FAM19A1 is a new ligand for GPR1 that modulates neural stem-cell proliferation and differentiation. FASEB J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Khalaj, A.J.; Sterky, F.H.; Sclip, A.; Schwenk, J.; Brunger, A.T.; Fakler, B.; Südhof, T.C. Deorphanizing FAM19A proteins as pan-neurexin ligands with an unusual biosynthetic binding mechanism. J. Cell Biol. 2020, 219. [Google Scholar] [CrossRef]
- Choi, S.Y.; Han, K.; Cutforth, T.; Chung, W.; Park, H.; Lee, D.; Kim, R.; Kim, M.-H.; Choi, Y.; Shen, K.; et al. Mice lacking the synaptic adhesion molecule Neph2/Kirrel3 display moderate hyperactivity and defective novel object preference. Front. Cell. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [Green Version]
- Risher, W.C.; Ustunkaya, T.; Singh Alvarado, J.; Eroglu, C. Rapid Golgi analysis method for efficient and unbiased classification of dendritic spines. PLoS ONE 2014, 9, e107591. [Google Scholar] [CrossRef] [Green Version]
- Sholl, D.A. Dendritic organization in the neurons of the visual and motor cortices of the cat. J. Anat. 1953, 87, 387–406. [Google Scholar] [PubMed]
- Beaudoin, G.M., 3rd; Lee, S.H.; Singh, D.; Yuan, Y.; Ng, Y.G.; Reichardt, L.F.; Arikkath, J. Culturing pyramidal neurons from the early postnatal mouse hippocampus and cortex. Nat. Protoc. 2012, 7, 1741–1754. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yuste, R.; Denk, W. Dendritic spines as basic functional units of neuronal integration. Nature 1995, 375, 682–684. [Google Scholar] [CrossRef]
- Nimchinsky, E.A.; Sabatini, B.L.; Svoboda, K. Structure and function of dendritic spines. Annu. Rev. Physiol. 2002, 64, 313–353. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.C.; Lu, J.; Zuo, Y. Spatiotemporal dynamics of dendritic spines in the living brain. Front. Neuroanat. 2014, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Yang, G.; Kwon, E.; Gan, W.B. Long-term sensory deprivation prevents dendritic spine loss in primary somatosensory cortex. Nature 2005, 436, 261–265. [Google Scholar] [CrossRef]
- Tang, G.; Gudsnuk, K.; Kuo, S.H.; Cotrina, M.L.; Rosoklija, G.; Sosunov, A.; Sonders, M.S.; Kanter, E.; Castagna, C.; Yamamoto, A.; et al. Loss of mTOR-dependent macroautophagy causes autistic-like synaptic pruning deficits. Neuron 2014, 83, 1131–1143. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Ash, R.T.; Baker, S.A.; Suter, B.; Ferguson, A.; Park, J.; Rudy, J.; Torsky, S.P.; Chao, H.-T.; Zoghbi, H.Y.; et al. Dendritic arborization and spine dynamics are abnormal in the mouse model of MECP2 duplication syndrome. J. Neurosci. 2013, 33, 19518–19533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Govek, E.E.; Newey, S.E.; Van Aelst, L. The role of the Rho GTPases in neuronal development. Genes Dev. 2005, 19, 1–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baj, G.; Patrizio, A.; Montalbano, A.; Sciancalepore, M.; Tongiorgi, E. Developmental and maintenance defects in Rett syndrome neurons identified by a new mouse staging system in vitro. Front. Cell. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuste, R. Dendritic Spines and Distributed Circuits. Neuron 2011, 71, 772–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borczyk, M.; Śliwińska, M.A.; Caly, A.; Bernas, T.; Radwanska, K. Neuronal plasticity affects correlation between the size of dendritic spine and its postsynaptic density. Sci. Rep. 2019, 9, 1693. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Penzes, P. Common mechanisms of excitatory and inhibitory imbalance in schizophrenia and autism spectrum disorders. Curr. Mol. Med. 2015, 15, 146–167. [Google Scholar] [CrossRef]
- Han, K.; Holder, J.L., Jr.; Schaaf, C.P.; Lu, H.; Chen, H.; Kang, H.; Tang, J.; Wu, Z.; Hao, S.; Cheung, S.W.; et al. SHANK3 overexpression causes manic-like behaviour with unique pharmacogenetic properties. Nature 2013, 503, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, K.; Fujita, Y.; Kasai, A.; Onaka, Y.; Hashimoto, H.; Okada, H.; Yamashita, T. Deletion of JMJD2B in neurons leads to defective spine maturation, hyperactive behavior and memory deficits in mouse. Transl. Psychiatry 2016, 6, e766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gipson, C.D.; Olive, M.F. Structural and functional plasticity of dendritic spines—root or result of behavior? Genes Brain Behav. 2017, 16, 101–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reissner, C.; Runkel, F.; Missler, M. Neurexins. Genome Biol. 2013, 14, 213. [Google Scholar] [CrossRef]
- Gangwar, S.P.; Zhong, X.; Seshadrinathan, S.; Chen, H.; Machius, M.; Rudenko, G. Molecular Mechanism of MDGA1: Regulation of Neuroligin 2:Neurexin Trans-synaptic Bridges. Neuron 2017, 94, 1132–1141.e1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koleske, A.J. Molecular mechanisms of dendrite stability. Nat. Rev. Neurosci. 2013, 14, 536–550. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Koleske, A.J. Mechanisms of synapse and dendrite maintenance and their disruption in psychiatric and neurodegenerative disorders. Annu. Rev. Neurosci. 2010, 33, 349–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charych, E.I.; Akum, B.F.; Goldberg, J.S.; Jörnsten, R.J.; Rongo, C.; Zheng, J.Q.; Firestein, B.L. Activity-Independent Regulation of Dendrite Patterning by Postsynaptic Density Protein PSD-95. J. Neurosci. 2006, 26, 10164. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yong, H.-J.; Hwang, J.-I.; Seong, J.-Y. Alterations in Dendritic Spine Maturation and Neurite Development Mediated by FAM19A1. Cells 2021, 10, 1868. https://doi.org/10.3390/cells10081868
Yong H-J, Hwang J-I, Seong J-Y. Alterations in Dendritic Spine Maturation and Neurite Development Mediated by FAM19A1. Cells. 2021; 10(8):1868. https://doi.org/10.3390/cells10081868
Chicago/Turabian StyleYong, Hyo-Jeong, Jong-Ik Hwang, and Jae-Young Seong. 2021. "Alterations in Dendritic Spine Maturation and Neurite Development Mediated by FAM19A1" Cells 10, no. 8: 1868. https://doi.org/10.3390/cells10081868
APA StyleYong, H. -J., Hwang, J. -I., & Seong, J. -Y. (2021). Alterations in Dendritic Spine Maturation and Neurite Development Mediated by FAM19A1. Cells, 10(8), 1868. https://doi.org/10.3390/cells10081868