
Insights on the Regeneration Potential of Müller Glia in the Mammalian Retina


Abstract
:1. Introduction
2. Müller Glia Anatomy and Function
3. Müller Glia Response to Injury in Mammals
4. Current Understanding of the Regeneration Potential of Müller Glia in Mammals
5. Clinical Applications of Reprogrammed Mammalian Müller Glia
6. Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Enoch, J.; McDonald, L.; Jones, L.; Jones, P.; Crabb, D.P. Evaluating Whether Sight Is the Most Valued Sense. JAMA Ophthalmol. 2019, 137, 1317–1320. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.S.; Park, S.S.; Albini, T.A.; Canto-Soler, M.V.; Klassen, H.; MacLaren, R.E.; Takahashi, M.; Nagiel, A.; Schwartz, S.D.; Bharti, K. Retinal stem cell transplantation: Balancing safety and potential. Prog. Retin. Eye Res. 2020, 75, 100779. [Google Scholar] [CrossRef] [PubMed]
- Trapani, I.; Auricchio, A. Has retinal gene therapy come of age? From bench to bedside and back to bench. Hum. Mol. Genet. 2019, 28, R108–R118. [Google Scholar] [CrossRef] [Green Version]
- Kantor, A.; McClements, M.E.; MacLaren, R.E. CRISPR-Cas9 DNA Base-Editing and Prime-Editing. Int. J. Mol. Sci. 2020, 21, 6240. [Google Scholar] [CrossRef]
- McClements, M.E.; Staurenghi, F.; MacLaren, R.E.; Cehajic-Kapetanovic, J. Optogenetic Gene Therapy for the Degenerate Retina: Recent Advances. Front. Neurosci. 2020, 14, 570909. [Google Scholar] [CrossRef]
- MacLaren, R.E. Electronic retinal implant surgery. Eye 2017, 31, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherpa, T.; Fimbel, S.M.; Mallory, D.E.; Maaswinkel, H.; Spritzer, S.D.; Sand, J.A.; Li, L.; Hyde, D.R.; Stenkamp, D.L. Ganglion cell regeneration following whole-retina destruction in zebrafish. Dev. Neurobiol. 2008, 68, 166–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsey, A.E.; Powers, M.K. Visual behavior of adult goldfish with regenerating retina. Vis. Neurosci. 2007, 24, 247–255. [Google Scholar] [CrossRef]
- Mensinger, A.F.; Powers, M.K. Visual function in regenerating teleost retina following cytotoxic lesioning. Vis. Neurosci. 1999, 16, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Powell, C.; Grant, A.; Cornblath, E.; Goldman, D. Analysis of DNA methylation reveals a partial reprogramming of the Muller glia genome during retina regeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 19814–19819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, R.; Fausett, B.V.; Goldman, D. Ascl1a regulates Muller glia dedifferentiation and retinal regeneration through a Lin-28-dependent, let-7 microRNA signalling pathway (vol 12, pg 1101, 2010). Nat. Cell Biol. 2015, 17, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fausett, B.V.; Goldman, D. A role for alpha1 tubulin-expressing Muller glia in regeneration of the injured zebrafish retina. J. Neurosci. 2006, 26, 6303–6313. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Barthel, L.K.; Raymond, P.A. Genetic evidence for shared mechanisms of epimorphic regeneration in zebrafish. Proc. Natl. Acad. Sci. USA 2009, 106, 9310–9315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagashima, M.; Barthel, L.K.; Raymond, P.A. A self-renewing division of zebrafish Muller glial cells generates neuronal progenitors that require N-cadherin to regenerate retinal neurons. Development 2013, 140, 4510–4521. [Google Scholar] [CrossRef] [Green Version]
- Kassen, S.C.; Ramanan, V.; Montgomery, J.E.; Burket, C.T.; Liu, C.-G.; Vihtelic, T.S.; Hyde, D.R. Time course analysis of gene expression during light-induced photoreceptor cell death and regeneration in albino zebrafish. Dev. Neurobiol. 2007, 67, 1009–1031. [Google Scholar] [CrossRef]
- Reichenbach, A.; Bringmann, A. Glia of the human retina. Glia 2020, 68, 768–796. [Google Scholar] [CrossRef]
- Turner, D.L.; Cepko, C.L. A common progenitor for neurons and glia persists in rat retina late in development. Nature 1987, 328, 131–136. [Google Scholar] [CrossRef]
- Jadhav, A.P.; Roesch, K.; Cepko, C.L. Development and neurogenic potential of Muller glial cells in the vertebrate retina. Prog. Retin. Eye Res. 2009, 28, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Jadhav, A.P.; Cho, S.-H.; Cepko, C.L. Notch activity permits retinal cells to progress through multiple progenitor states and acquire a stem cell property. Proc. Natl. Acad. Sci. USA 2006, 103, 18998–19003. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, T.; Mukherjee, S.; Bao, Z.-Z.; Morrow, E.; Cepko, C.L. rax, Hes1, and notch1 promote the formation of Muller glia by postnatal retinal progenitor cells. Neuron 2000, 26, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Goureau, O.; Rhee, K.D.; Yang, X.-J. Ciliary neurotrophic factor promotes muller glia differentiation from the postnatal retinal progenitor pool. Dev. Neurosci. 2004, 26, 359–370. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Das, A.V.; Mallya, K.B.; Ahmad, L. Ciliary neurotrophic factor-mediated signaling regulates neuronal versus glial differentiation of retinal stem cells/progenitors by concentration-dependent recruitment of mitogen-activated protein kinase and Janus kinase-signal transducer and activator of transcription pathways in conjunction with Notch signaling. Stem Cells 2008, 26, 2611–2624. [Google Scholar]
- Sidman, R.L. Histogenesis of Mouse Retina Studied with Thymidine-H-3. Anat. Record 1960, 136, 276–277. [Google Scholar]
- Reichenbach, A.; Bringmann, A. New functions of Muller cells. Glia 2013, 61, 651–678. [Google Scholar] [CrossRef]
- Bringmann, A.; Grosche, A.; Pannicke, T.; Reichenbach, A. GABA and Glutamate Uptake and Metabolism in Retinal Glial (Muller) Cells. Front. Endocrinol. 2013, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Biedermann, B.; Biedermann, A.; Franze, K.; Faude, F.; Wiedemann, P.; Reichenbach, A. GABA(A) receptors in Muller glial cells of the human retina. Glia 2004, 46, 302–310. [Google Scholar] [CrossRef]
- Brew, H.; Attwell, D. Electrogenic Glutamate Uptake Is a Major Current Carrier in the Membrane of Axolotl Retinal Glial-Cells. Nature 1987, 327, 707–709. [Google Scholar] [CrossRef]
- Rauen, T.; Taylor, W.R.; Kuhlbrodt, K.; Wiessner, M. High-affinity glutamate transporters in the rat retina: A major role of the glial glutamate transporter GLAST-1 in transmitter clearance. Cell Tissue Res. 1998, 291, 19–31. [Google Scholar] [CrossRef]
- Pow, D.V.; Robinson, S.R. Glutamate in Some Retinal Neurons Is Derived Solely from Glia. Neuroscience 1994, 60, 355–366. [Google Scholar] [CrossRef]
- Hasegawa, H.; Ma, T.; Skach, W.; Matthay, A.M.; Verkman, A.S. Molecular cloning of a mercurial-insensitive water channel expressed in selected water-transporting tissues. J. Biol. Chem. 1994, 269, 5497–5500. [Google Scholar] [CrossRef]
- Hasegawa, H.; Ma, T.; Skach, W.; Matthay, A.M.; Verkman, A.S. Immunogold evidence suggests that coupling of K+ siphoning and water transport in rat retinal Muller cells is mediated by a coenrichment of Kir4.1 and AQP4 in specific membrane domains. Glia 1999, 26, 47–54. [Google Scholar]
- Hasegawa, H.; Ma, T.; Skach, W.; Matthay, A.M.; Verkman, A.S. Role of retinal glial cells in neurotransmitter uptake and metabolism. Neurochem. Int. 2009, 54, 143–160. [Google Scholar]
- Newman, E.A. A physiological measure of carbonic anhydrase in Muller cells. Glia 1994, 11, 291–299. [Google Scholar] [CrossRef]
- Tout, S.; Chan-Ling, T.; Holländer, H.; Stone, J. The role of Muller cells in the formation of the blood-retinal barrier. Neuroscience 1993, 55, 291–301. [Google Scholar] [CrossRef]
- Choi, Y.K.; Kim, K.W. Blood-neural barrier: Its diversity and coordinated cell-to-cell communication. BMB Rep. 2008, 41, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Metea, M.R.; Newman, E.A. Glial cells dilate and constrict blood vessels: A mechanism of neurovascular coupling. J. Neurosci. 2006, 26, 2862–2870. [Google Scholar] [CrossRef]
- Bernardos, R.; Barthel, L.K.; Meyers, J.R.; Raymond, P.A. Late-stage neuronal progenitors in the retina are radial Muller glia that function as retinal stem cells. J. Neurosci. 2007, 27, 7028–7040. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, J.M.; Singhal, S.; Bhatia, B.; Keegan, D.J.; Reh, T.A.; Luthert, P.J.; Khaw, P.T.; Limb, G.A. MIO-M1 cells and similar Muller glial cell lines derived from adult human retina exhibit neural stem cell characteristics. Stem Cells 2007, 25, 2033–2043. [Google Scholar] [CrossRef]
- Lawrence, J.M.; Singhal, S.; Bhatia, B.; Keegan, D.J.; Reh, T.A.; Luthert, P.J.; Khaw, P.T.; Limb, G.A. Human Muller Glia with Stem Cell Characteristics Differentiate into Retinal Ganglion Cell (RGC) Precursors In Vitro and Partially Restore RGC Function In Vivo Following Transplantation. Stem Cells Transl. Med. 2012, 1, 188–199. [Google Scholar]
- Jayaram, H.; Jones, M.F.; Eastlake, K.; Cottrill, P.B.; Becker, S.; Wiseman, J.; Khaw, P.T.; Limb, G.A. Transplantation of Photoreceptors Derived From Human Muller Glia Restore Rod Function in the P23H Rat. Stem Cells Transl. Med. 2014, 3, 323–333. [Google Scholar] [CrossRef]
- Becker, S.; Singhal, S.; Jones, M.F.; Eastlake, K.; Cottrill, P.B.; Jayaram, H.; Limb, G.A. Acquisition of RGC phenotype in human Muller glia with stem cell characteristics is accompanied by upregulation of functional nicotinic acetylcholine receptors. Mol. Vis. 2013, 19, 1925–1936. [Google Scholar]
- Kiyama, T.; Li, H.; Gupta, M.; Lin, Y.-P.; Chuang, A.Z.; Otteson, D.; Wang, S.W. Distinct neurogenic potential in the retinal margin and the pars plana of mammalian eye. J. Neurosci. 2012, 32, 12797–12807. [Google Scholar] [CrossRef] [Green Version]
- Jayakody, S.A.; Cordero, A.G.; Ali, R.; Pearson, R.A. Cellular strategies for retinal repair by photoreceptor replacement. Prog. Retin. Eye Res. 2015, 46, 31–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salero, E.; Blenkinsop, T.A.; Corneo, B.; Harris, A.; Rabin, D.; Stern, J.H.; Temple, S. Adult Human RPE Can Be Activated into a Multipotent Stem Cell that Produces Mesenchymal Derivatives. Cell Stem Cell 2012, 10, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.Z.; Yan, R.T. The Retinal Pigment Epithelium: A Convenient Source of New Photoreceptor cells? J. Ophthalmic Vis. Res. 2014, 9, 83–93. [Google Scholar] [PubMed]
- Dyer, M.A.; Cepko, C.L. Control of Muller glial cell proliferation and activation following retinal injury. Nat. Neurosci. 2000, 3, 873–880. [Google Scholar] [CrossRef]
- Bringmann, A.; Iandiev, I.; Pannicke, T.; Wurm, A.; Hollborn, M.; Wiedemann, P.; Osborne, N.N.; Reichenbach, A. Cellular signaling and factors involved in Muller cell gliosis: Neuroprotective and detrimental effects. Prog. Retin. Eye Res. 2009, 28, 423–451. [Google Scholar] [CrossRef]
- Bringmann, A.; Iandiev, I.; Pannicke, T.; Wurm, A.; Hollborn, M.; Wiedemann, P.; Osborne, N.N.; Reichenbach, A. Potential for neural regeneration after neurotoxic injury in the adult mammalian retina. Proc. Natl. Acad. Sci. USA 2004, 101, 13654–13659. [Google Scholar]
- Karl, M.; Hayes, S.; Nelson, B.R.; Tan, K.; Buckingham, B.; Reh, T.A. Stimulation of neural regeneration in the mouse retina. Proc. Natl. Acad. Sci. USA 2008, 105, 19508–19513. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Zheng, H.; Chen, Z.-L.; Xiao, H.-L.; Shen, Z.-J.; Zhou, G.-M. Preferential regeneration of photoreceptor from Muller glia after retinal degeneration in adult rat. Vis. Res. 2008, 48, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Osakada, F.; Ooto, S.; Akagi, T.; Mandai, M.; Akaike, A.; Takahashi, M. Wnt signaling promotes regeneration in the retina of adult mammals. J. Neurosci. 2007, 27, 4210–4219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angbohang, A.; Wu, N.; Charalambous, T.; Eastlake, K.; Lei, Y.; Kim, Y.S.; Sun, X.H.; Limb, G.A. Downregulation of the Canonical WNT Signaling Pathway by TGFbeta1 Inhibits Photoreceptor Differentiation of Adult Human Muller Glia with Stem Cell Characteristics. Stem Cells Dev. 2016, 25, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannelli, S.G.; Demontis, G.C.; Pertile, G.; Rama, P.; Broccoli, V. Adult human Muller glia cells are a highly efficient source of rod photoreceptors. Stem Cells 2011, 29, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Joly, S.; Pernet, V.; Samardzija, M.; Grimm, C. PAX6-Positive Muller Glia Cells Express Cell Cycle Markers but Do Not Proliferate After Photoreceptor Injury in the Mouse Retina. Glia 2011, 59, 1033–1046. [Google Scholar] [CrossRef] [PubMed]
- Löffler, K.; Schäfer, P.; Völkner, M.; Holdt, T.; Karl, M.O. Age-dependent Muller glia neurogenic competence in the mouse retina. Glia 2015, 63, 1809–1824. [Google Scholar] [CrossRef] [PubMed]
- Ueki, Y.; Reh, T.A. EGF stimulates Muller glial proliferation via a BMP-dependent mechanism. Glia 2013, 61, 778–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Close, J.L.; Liu, J.; Gumuscu, B.; Reh, T.A. Epidermal growth factor receptor expression regulates proliferation in the postnatal rat retina. Glia 2006, 54, 94–104. [Google Scholar] [CrossRef]
- Close, J.L.; Gumuscu, B.; Reh, T.A. Retinal neurons regulate proliferation of postnatal progenitors and Muller glia in the rat retina via TGF beta signaling. Development 2005, 132, 3015–3026. [Google Scholar] [CrossRef] [Green Version]
- Takeda, M.; Takamiya, A.; Jiao, J.-W.; Cho, K.-S.; Trevino, S.G.; Matsuda, T.; Chen, D.F. alpha-Aminoadipate induces progenitor cell properties of Muller glia in adult mice. Investig. Ophthalmol. Vis. Sci. 2008, 49, 1142–1150. [Google Scholar] [CrossRef]
- Fausett, B.V.; Gumerson, J.D.; Goldman, D. The proneural basic helix-loop-helix gene ascl1a is required for retina regeneration. J. Neurosci. 2008, 28, 1109–1117. [Google Scholar] [CrossRef]
- Pollak, J.; Wilken, M.S.; Ueki, Y.; Cox, K.E.; Sullivan, J.M.; Taylor, R.J.; Levine, E.M.; Reh, T.A. ASCL1 reprograms mouse Muller glia into neurogenic retinal progenitors. Development 2013, 140, 2619–2631. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.; Zheng, H.; Xiao, H.-L.; She, Z.-J.; Zhou, G.-M. Sonic hedgehog promotes stem-cell potential of Muller glia in the mammalian retina. Biochem. Biophys. Res. Commun. 2007, 363, 347–354. [Google Scholar] [CrossRef] [PubMed]
- MacLaren, R.E. Development and role of retinal glia in regeneration of ganglion cells following retinal injury. Br. J. Ophthalmol. 1996, 80, 458–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorstad, N.L.; Wilken, M.S.; Grimes, W.N.; Wohl, S.G.; VandenBosch, L.S.; Yoshimatsu, T.; Wong, R.O.; Rieke, F.; Reh, T.A. Stimulation of functional neuronal regeneration from Muller glia in adult mice. Nature 2017, 548, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Ghai, K.; Zelinka, C.; Fischer, A.J. Notch signaling influences neuroprotective and proliferative properties of mature Muller glia. J. Neurosci. 2010, 30, 3101–3112. [Google Scholar] [CrossRef]
- Fischer, A.J.; Reh, T.A. Exogenous growth factors stimulate the regeneration of ganglion cells in the chicken retina. Dev. Biol. 2002, 251, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.J.; McGuire, C.R.; Dierks, B.D.; Reh, T.A. Insulin and fibroblast growth factor 2 activate a neurogenic program in Muller glia of the chicken retina. J. Neurosci. 2002, 22, 9387–9398. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.J.; Scott, M.A.; Tuten, W. Mitogen-activated protein kinase-signaling stimulates Muller glia to proliferate in acutely damaged chicken retina. Glia 2009, 57, 166–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, A.J.; Reh, T.A. Muller glia are a potential source of neural regeneration in the postnatal chicken retina. Nat. Neurosci. 2001, 4, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Wang, S.; Zhang, S.; Zhang, P.; Zhou, G. Directed transdifferentiation of Muller glial cells to photoreceptors using the sonic hedgehog signaling pathway agonist purmorphamine. Mol. Med. Rep. 2017, 16, 7993–8002. [Google Scholar] [CrossRef] [Green Version]
- Yao, K.; Qiu, S.; Wang, Y.V.; Park, S.J.H.; Mohns, E.J.; Mehta, B.; Liu, X.; Chang, B.; Zenisek, D.; Crair, M.C.; et al. Restoration of vision after de novo genesis of rod photoreceptors in mammalian retinas. Nature 2018, 560, 484–488. [Google Scholar] [CrossRef]
- Yao, K.; Qiu, S.; Tian, L.; Snider, W.D.; Flannery, J.; Schaffer, D.V.; Chen, B. Wnt Regulates Proliferation and Neurogenic Potential of Muller Glial Cells via a Lin28/let-7 miRNA-Dependent Pathway in Adult Mammalian Retinas. Cell Rep. 2016, 17, 165–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohl, S.G.; Hooper, M.J.; Reh, T.A. MicroRNAs miR-25, let-7 and miR-124 regulate the neurogenic potential of Muller glia in mice. Development 2019, 146, dev179556. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.-P.; Xiong, Y.; Song, W.-T.; Zhang, E.-D.; Gao, Z.-L.; Yao, F.; Su, T.; Zhou, R.-R.; Xia, X.-B. MicroRNA-28 potentially regulates the photoreceptor lineage commitment of Muller glia-derived progenitors. Sci. Rep. 2017, 7, 11374. [Google Scholar] [CrossRef]
- Sanges, D.; Romo, N.; Simonte, G.; Vicino, U.D.; Tahoces, A.D.; Fernández, E.; Cosma, M.P. Wnt/beta-catenin signaling triggers neuron reprogramming and regeneration in the mouse retina. Cell Rep. 2013, 4, 271–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanges, D.; Simonte, G.; Di Vicino, U.; Romo, N.; Pinilla, I.; Nicolás, M.; Cosma, M.P. Reprogramming Muller glia via in vivo cell fusion regenerates murine photoreceptors. J. Clin. Investig. 2016, 126, 3104–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, K.; Fukuda, A.; Hisatake, K. Mechanisms of the Metabolic Shift during Somatic Cell Reprogramming. Int. J. Mol. Sci. 2019, 20, 2254. [Google Scholar] [CrossRef] [Green Version]
- Naik, P.P.; Birbrair, A.; Bhutia, S.K. Mitophagy-driven metabolic switch reprograms stem cell fate. Cell. Mol. Life Sci. 2019, 76, 27–43. [Google Scholar] [CrossRef]
- Palazzo, I.; Deistler, K.; Hoang, T.V.; Blackshaw, S.; Fischer, A.J. NF-kappaB signaling regulates the formation of proliferating Muller glia-derived progenitor cells in the avian retina. Development 2020, 147, dev183418. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Slembrouck-Brec, A.; Rodrigues, A.; Rabesandratana, O.; Gagliardi, G.; Nanteau, C.; Fouquet, S.; Thuret, G.; Reichman, S.; Orieux, G.; Goureau, O. Reprogramming of Adult Retinal Muller Glial Cells into Human-Induced Pluripotent Stem Cells as an Efficient Source of Retinal Cells. Stem Cells Int. 2019, 2019, 7858796. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.H.; Shen, W.; Davidson, K.C.; Pébay, A.; Wong, R.C.-B.; Yau, B.; Gillies, M. Differentiation of Retinal Glial Cells From Human Embryonic Stem Cells by Promoting the Notch Signaling Pathway. Front. Cell. Neurosci. 2019, 13, 527. [Google Scholar] [CrossRef] [PubMed]
- Del Debbio, C.B.; Balasubramanian, S.; Parameswaran, S.; Chaudhuri, A.; Qiu, F.; Ahmad, I. Notch and Wnt signaling mediated rod photoreceptor regeneration by Muller cells in adult mammalian retina. PLoS ONE 2010, 5, e12425. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
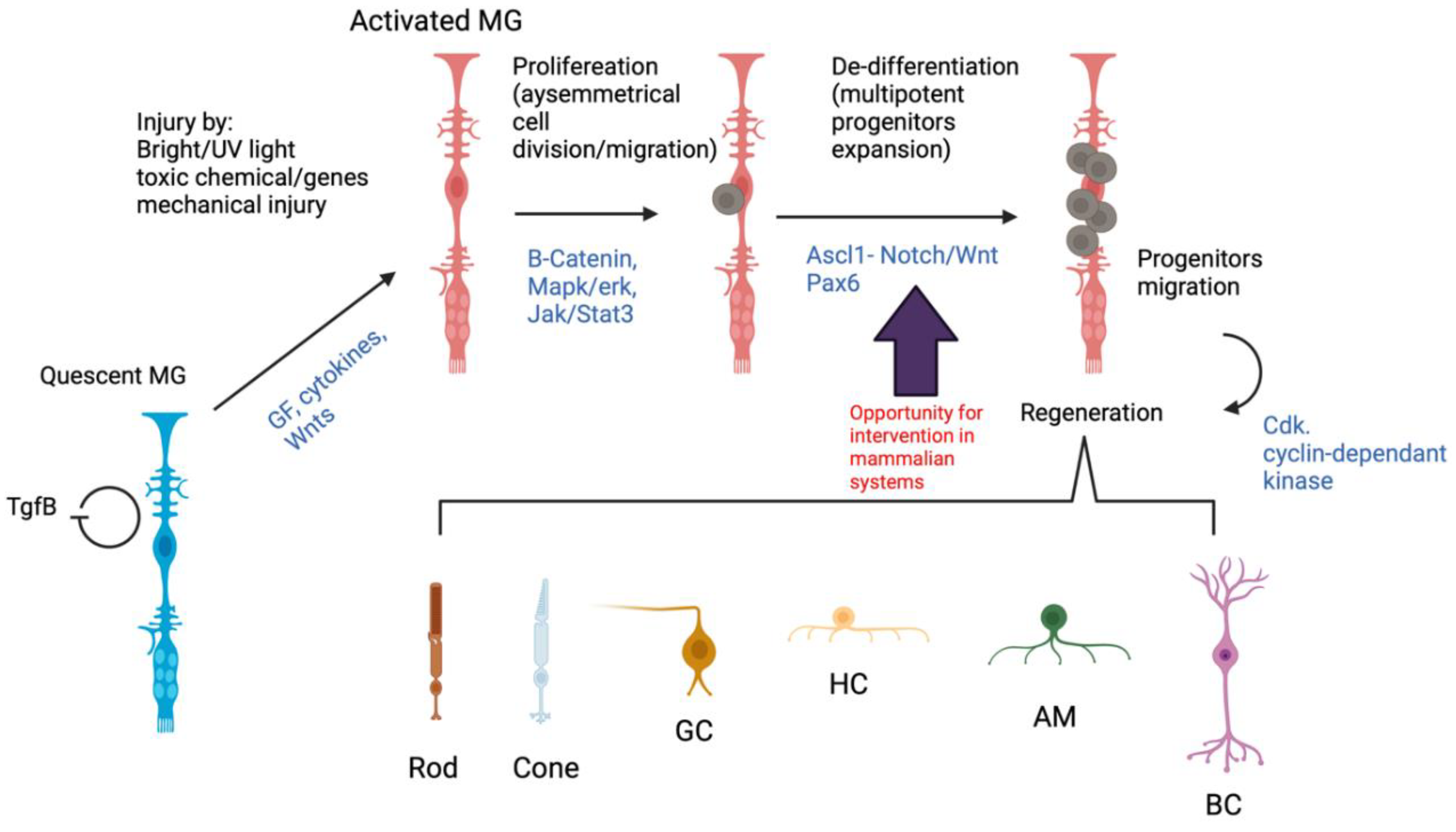{kind=link}
| Signalling Pathway/ Factor | Function | Species Tested | Effect on Müller Glia Regeneration | References |
|---|---|---|---|---|
| Ascl1 | Transcription factor | Birds, rodents | Stimulates | [49,55,56] |
| BMP-SMAD | Signalling pathways activated by a secreted factor (BMP) when binding to a transcription factor (SMAD) | Rodents | Stimulates | [56] |
| EGF/EGFR | Signalling pathways activated by a secreted factor (EGF) when binding to its receptor (EGFR) | Rodents | Stimulates | [49,57,58] |
| Glutamate | Neurotransmitter | Rodents | Stimulates | [59] |
| FGF2-FGFR-MAPK | Secreted factor (FGF2) binding to its receptor (FGFR) to activate a signalling pathway (MAPK) | Birds, rodents | Stimulates | [57,65,66,67,68] |
| Delta-Notch | Signalling cascade activated by a transmembrane ligand (Delta) binding to its receptor | Birds, rodents | Stimulates | [20,49,65] |
| Pax6 | Transcription factor | Birds, rodents | Unknown | [49,69] |
| Insulin-IGF1-PI3K | Secreted factor (FGF2) binding to its receptor (FGFR) to activate a signalling pathway (MAPK) | Birds, rodents | Stimulates | [66,67,68] |
| SHH | Secreted factor | Rodents | Stimulates | [62,70] |
| TGF-B | Secreted factor | Rodents | Inhibits | [52,57,58] |
| Wnt/B-catenin | Signal transduction pathway | Rodents | Stimulates | [51,52,71,72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salman, A.; McClements, M.E.; MacLaren, R.E. Insights on the Regeneration Potential of Müller Glia in the Mammalian Retina. Cells 2021, 10, 1957. https://doi.org/10.3390/cells10081957
Salman A, McClements ME, MacLaren RE. Insights on the Regeneration Potential of Müller Glia in the Mammalian Retina. Cells. 2021; 10(8):1957. https://doi.org/10.3390/cells10081957
Chicago/Turabian StyleSalman, Ahmed, Michelle E. McClements, and Robert E. MacLaren. 2021. "Insights on the Regeneration Potential of Müller Glia in the Mammalian Retina" Cells 10, no. 8: 1957. https://doi.org/10.3390/cells10081957
APA StyleSalman, A., McClements, M. E., & MacLaren, R. E. (2021). Insights on the Regeneration Potential of Müller Glia in the Mammalian Retina. Cells, 10(8), 1957. https://doi.org/10.3390/cells10081957